
Effects of Liming on the Morphologies and Nutrients of Different Functional Fine Roots of Cunninghamia lanceolata Seedlings

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Pot Experiment
2.3. Root Trait Analysis
2.4. Data Analysis
3. Results
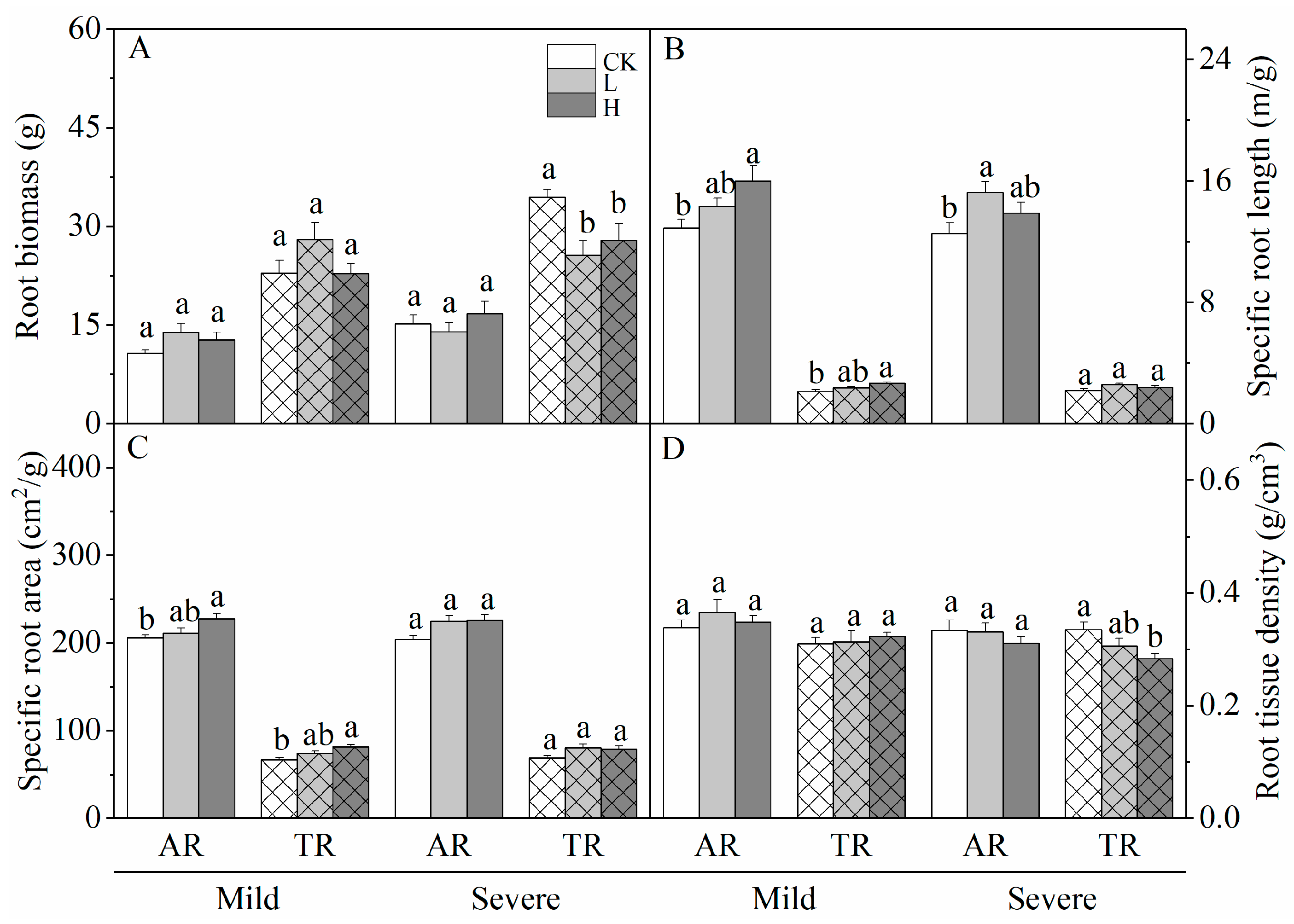
3.1. Effects of Liming on Root Morphology
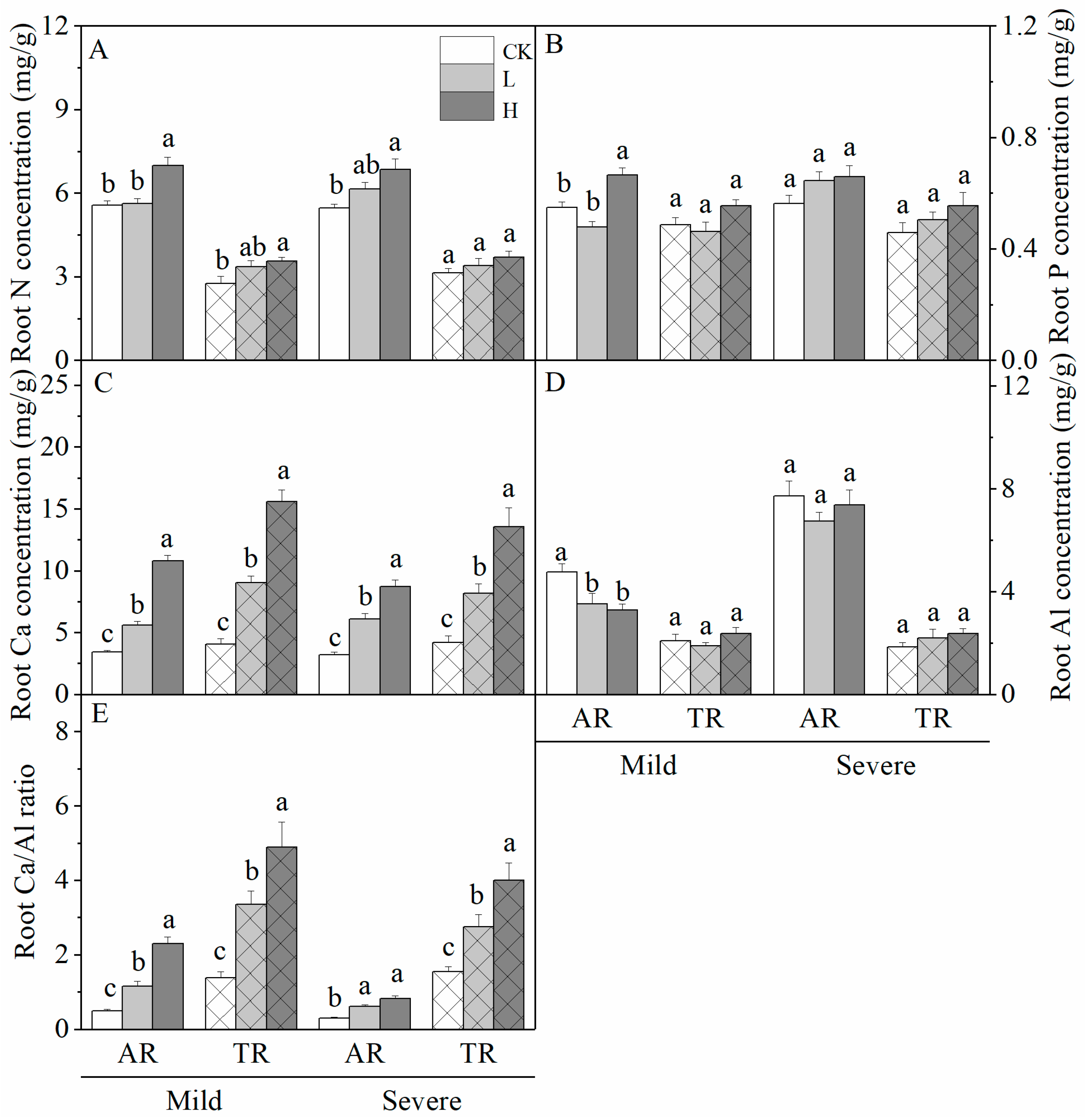
3.2. Effects of Liming on Root Nutrient Concentrations
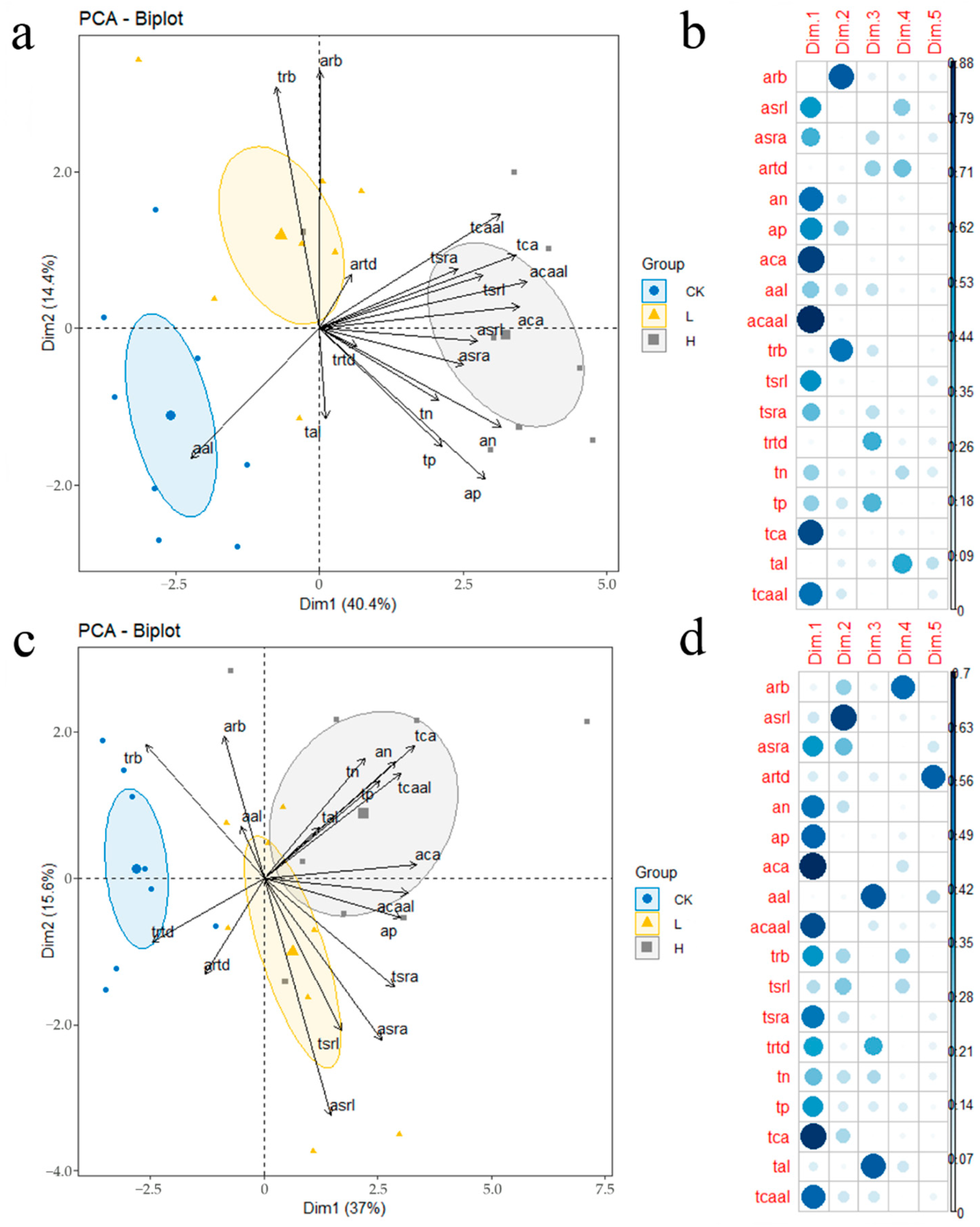
3.3. Relationships among Root Traits under Liming
4. Discussion
4.1. Effect of Liming on Root Traits
4.2. Responses of Different Functional Roots to Liming
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, Y.; Yang, J.; Zeng, S.; Wu, D.; Jacobs, D.F.; Sloan, J.L. Soil pH, organic matter, and nutrient content change with the continuous cropping of Cunninghamia lanceolata plantations in South China. J. Soils Sediments 2017, 17, 2230–2238. [Google Scholar] [CrossRef]
- Farooq, T.H.; Yan, W.; Rashid, M.H.U.; Tigabu, M.; Gilani, M.M.; Zou, X.H.; Wu, P.F. Chinese fir (Cunninghamia lanceolata) a green gold of China with continues decline in its productivity over the successive rotations: A review. Appl. Ecol. Environ. Res. 2019, 17, 11055–11067. [Google Scholar] [CrossRef]
- Tian, D.; Xiang, W.; Chen, X.; Yan, W.; Fang, X.; Kang, W.; Dan, X.; Peng, C.; Peng, Y. A long-term evaluation of biomass production in first and second rotations of Chinese fir plantations at the same site. Forestry 2011, 84, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Bohan, L.; Larssen, T.; Seip, H.M. Response of five Chinese forest soils to acidic inputs: Batch experiment. Geoderma 1998, 86, 295–316. [Google Scholar] [CrossRef]
- Liu, X.; Fu, Z.; Zhang, B.; Zhai, L.; Meng, M.; Lin, J.; Zhuang, J.; Wang, G.G.; Zhang, J. Effects of sulfuric, nitric, and mixed acid rain on Chinese fir sapling growth in Southern China. Ecotoxicol. Environ. Saf. 2018, 160, 154–161. [Google Scholar] [CrossRef]
- Fan, H. A review of research on interactions between acid rain and forest decline. J. Fujian Coll. For. 2003, 23, 88–92. [Google Scholar]
- Leys, B.A.; Likens, G.E.; Johnson, C.E.; Craine, J.M.; Lacroix, B.; McLauchlan, K.K. Natural and anthropogenic drivers of calcium depletion in a northern forest during the last millennium. Proc. Natl. Acad. Sci. USA 2016, 113, 6934–6938. [Google Scholar] [CrossRef] [Green Version]
- Battles, J.J.; Fahey, T.J.; Driscoll, C.T.; Blum, J.D.; Johnson, C.E. Restoring soil calcium reverses forest decline. Environ. Sci. Technol. Lett. 2014, 1, 15–19. [Google Scholar] [CrossRef]
- Rosi-Marshall, E.J.; Bernhardt, E.S.; Buso, D.C.; Driscoll, C.T.; Likens, G.E. Acid rain mitigation experiment shifts a forested watershed from a net sink to a net source of nitrogen. Proc. Natl. Acad. Sci. USA 2016, 113, 7580–7583. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.; Driscoll, C.T.; Johnson, C.E.; Siccama, T.G. Chemical changes in soil and soil solution after calcium silicate addition to a northern hardwood forest. Biogeochemistry 2010, 100, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.-W.; Wu, F.-H.; Wang, W.-H.; Chen, J.; Li, Z.-J.; Dong, X.-J.; Patton, J.; Pei, Z.-M.; Zheng, H.-L. Effects of calcium on seed germination, seedling growth and photosynthesis of six forest tree species under simulated acid rain. Tree Physiol. 2011, 31, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Freschet, G.T.; Roumet, C.; Comas, L.H.; Weemstra, M.; Bengough, A.G.; Rewald, B.; Bardgett, R.D.; De Deyn, G.B.; Johnson, D.; Klimesova, J.; et al. Root traits as drivers of plant and ecosystem functioning: Current understanding, pitfalls and future research needs. New Phytol. 2021, 232, 1123–1158. [Google Scholar] [CrossRef] [PubMed]
- Ostonen, I.; Püttsepp, Ü.; Biel, C.; Alberton, O.; Bakker, M.R.; Lõhmus, K.; Majdi, H.; Metcalfe, D.; Olsthoorn, A.F.M.; Pronk, A.; et al. Specific root length as an indicator of environmental change. Plant Biosyst. 2007, 141, 426–442. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A. Root decisions. Plant Cell Environ. 2009, 32, 628–640. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Wells, C.E.; Yanai, R.D.; Whitbeck, J.L. Building roots in a changing environment: Implications for root longevity. New Phytol. 2000, 147, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Kochian, L.V.; Pineros, M.A.; Liu, J.; Magalhaes, J.V. Plant adaptation to acid soils: The molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 2015, 66, 571–598. [Google Scholar] [CrossRef]
- Bakker, M.R.; Kerisit, R.; Verbist, K.; Nys, C. Effects of liming on rhizosphere chemistry and growth of fine roots and of shoots of sessile oak (Quercus petraea). Plant Soil 1999, 217, 243–255. [Google Scholar] [CrossRef]
- Miyatani, K.; Mizusawa, Y.; Okada, K.; Tanikawa, T.; Makita, N.; Hirano, Y. Fine root traits in Chamaecyparis obtusa forest soils with different acid buffering capacities. Trees 2015, 30, 415–429. [Google Scholar] [CrossRef]
- Guo, D.; Xia, M.; Wei, X.; Chang, W.; Liu, Y.; Wang, Z. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytol. 2008, 180, 673–683. [Google Scholar] [CrossRef]
- Pregitzer, K.S. Tree root architecture—Form and function. New Phytol. 2008, 180, 562–564. [Google Scholar] [CrossRef] [PubMed]
- Zadworny, M.; McCormack, M.L.; Zytkowiak, R.; Karolewski, P.; Mucha, J.; Oleksyn, J. Patterns of structural and defense investments in fine roots of Scots pine (Pinus sylvestris L.) across a strong temperature and latitudinal gradient in Europe. Glob. Chang. Biol. 2017, 23, 1218–1231. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Estrada, L.R.; Vera-Caraballo, V.; Ruth, L.E.; Eissenstat, D.M. Root anatomy, morphology, and longevity among root orders in Vaccinium corymbosum (ericaceae). Am. J. Bot. 2008, 95, 1506–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pu, X.; Yin, C.; Xiao, Q.; Qiao, M.; Liu, Q. Fine roots branch orders of Abies faxoniana respond differentially to warming in a subalpine coniferous forest ecosystem. Agrofor. Syst. 2016, 91, 955–966. [Google Scholar] [CrossRef]
- Guo, D.L.; Mitchell, R.J.; Hendricks, J.J. Fine root branch orders respond differentially to carbon source-sink manipulations in a longleaf pine forest. Oecologia 2004, 140, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Yan, G.; Xing, Y.; Chen, F.; Zhang, X.; Wang, J.; Zhang, J.; Dai, G.; Zheng, X.; Sun, W.; et al. Nitrogen deposition and decreased precipitation does not change total nitrogen uptake in a temperate forest. Sci. Total Environ. 2019, 651, 32–41. [Google Scholar] [CrossRef]
- Kramer-Walter, K.R.; Laughlin, D.C. Root nutrient concentration and biomass allocation are more plastic than morphological traits in response to nutrient limitation. Plant Soil 2017, 416, 539–550. [Google Scholar] [CrossRef]
- Haling, R.E.; Simpson, R.J.; Delhaize, E.; Hocking, P.J.; Richardson, A.E. Effect of lime on root growth, morphology and the rhizosheath of cereal seedlings growing in an acid soil. Plant Soil 2010, 327, 199–212. [Google Scholar] [CrossRef]
- Halman, J.M.; Schaberg, P.G.; Hawley, G.J.; Pardo, L.H.; Fahey, T.J. Calcium and aluminum impacts on sugar maple physiology in a northern hardwood forest. Tree Physiol. 2013, 33, 1242–1251. [Google Scholar] [CrossRef] [Green Version]
- Vanguelova, E.I.; Hirano, Y.; Eldhuset, T.D.; Sas-Paszt, L.; Bakker, M.R.; Puttsepp, U.; Brunner, I.; Lohmus, K.; Godbold, D. Tree fine root Ca/Al molar ratio—Indicator of Al and acidity stress. Plant Biosyst. 2007, 141, 460–480. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Tjoelker, M.G.; Pregitzer, K.S.; Wright, I.J.; Oleksyn, J.; Machado, J.L. Scaling of respiration to nitrogen in leaves, stems and roots of higher land plants. Ecol. Lett. 2008, 11, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Comas, L.; Bouma, T.; Eissenstat, D. Linking root traits to potential growth rate in six temperate tree species. Oecologia 2002, 132, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Son, Y.; Kim, C.; Yi, M.-J.; Kim, Z.-S.; Lee, W.-K.; Hong, S.-K. Fine root dynamics in thinned and limed pitch pine and Japanese larch plantations. J. Plant Nutr. 2007, 30, 1821–1839. [Google Scholar] [CrossRef]
- Hettiarachchi, L.S.K.; Sinclair, A.H. Effects of addition of magnesium and calcium supplied in liming and non-liming materials on the growth of Camellia japonica in an acid soil, and its soil ph changes, nutrient uptake, and availability. Commun. Soil Sci. Plant Anal. 2002, 33, 2965–2988. [Google Scholar] [CrossRef]
- Chen, G.T.; Zheng, J.; Peng, T.C.; Li, S.; Qiu, X.R.; Chen, Y.Q.; Ma, H.Y.; Tu, L.H. Fine root morphology and chemistry characteristics in different branch orders of Castanopsis platyacantha and their responses to nitrogen addition. Chin. J. Appl. Ecol. 2017, 28, 3461–3468. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, Y.F.; Fang, X.M.; Chen, F.S.; Zong, Y.Y.; Gu, H.J.; Hu, X.F. Influence of simulated acid rain on nitrogen and phosphorus contents and their stoichiometric ratios of tea organs in a red soil region, China. Chin. J. Appl. Ecol. 2017, 28, 1309–1316. (In Chinese) [Google Scholar] [CrossRef]
- Lambers, H.; Shane, M.W.; Cramer, M.D.; Pearse, S.J.; Veneklaas, E.J. Root structure and functioning for efficient acquisition of phosphorus: Matching morphological and physiological traits. Ann. Bot. 2006, 98, 693–713. [Google Scholar] [CrossRef] [Green Version]
- Bakker, M.R. The effect of lime and gypsum applications on a sessile oak (Quercus petraea (M.) Liebl.) stand at La Croix-Scaille (French Ardennes) II. Fine root dynamics. Plant Soil 1999, 206, 109–121. [Google Scholar] [CrossRef]
- Rengel, Z.; Zhang, W.H. Role of dynamics of intracellular calcium in aluminium-toxicity syndrome. New Phytol. 2003, 159, 295–314. [Google Scholar] [CrossRef]
- Hepler, P.K. Calcium: A central regulator of plant growth and development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef]
- Ma, J.F.; Chen, Z.C.; Shen, R.F. Molecular mechanisms of Al tolerance in gramineous plants. Plant Soil 2014, 381, 1–12. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Y.; Liu, Y.; Guo, H.; Li, T.; Li, Z.-H.; Shi, G. Long-term effects of liming on health and growth of a Masson pine stand damaged by soil acidification in Chongqing, China. PLoS ONE 2014, 9, e94230. [Google Scholar] [CrossRef]
- Burton, A.J.; Jarvey, J.C.; Jarvi, M.P.; Zak, D.R.; Pregitzer, K.S. Chronic N deposition alters root respiration-tissue N relationship in northern hardwood forests. Glob. Chang. Biol. 2012, 18, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Heyburn, J.; McKenzie, P.; Crawley, M.J.; Fornara, D.A. Long-term belowground effects of grassland management: The key role of liming. Ecol. Appl. 2017, 27, 2001–2012. [Google Scholar] [CrossRef] [PubMed]
- DeForest, J.L.; Snell, R.S. Tree growth response to shifting soil nutrient economy depends on mycorrhizal associations. New Phytol. 2020, 225, 2557–2566. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.K.; Raynal, D.J. Liming influences growth and nutrient balances in sugar maple (Acer saccharum) seedlings on an acidic forest soil. Environ. Exp. Bot. 1998, 39, 105–116. [Google Scholar] [CrossRef]
- Jiang, J.; Sheng, X.; Shao, J.; Lin, Y.; He, Q.; Han, W. Responses of the major functional traits of Pinus massoniana and Schima superba seedlings to nitrogen, phosphorus, and lime addition. Chin. J. Appl. Environ. Biol. 2020, 26, 410–416. [Google Scholar]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Helmisaari, H.S.; Hallbacken, L. Fine-root biomass and necromass in limed and fertilized Norway spruce (Picea abies (L.) Karst.) stands. For. Ecol. Manag. 1999, 119, 99–110. [Google Scholar] [CrossRef]
- Clivot, H.; Pagnout, C.; Aran, D.; Devin, S.; Bauda, P.; Poupin, P.; Guerold, F. Changes in soil bacterial communities following liming of acidified forests. Appl. Soil Ecol. 2012, 59, 116–123. [Google Scholar] [CrossRef]
- Yu, J.; Xia, L.; Yin, D.; Zhou, C. Effects of phosphorus on aluminum tolerance of Chinese fir seedlings. Sci. Silvae Sin. 2018, 54, 36–47. (In Chinese) [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Soil Acidification | pH | C (%) | N (%) | Available P (mg/kg) | Exchangeable Al (mg/kg) | Exchangeable Ca (mg/kg) | Ca/Al Molar Ratio |
|---|---|---|---|---|---|---|---|
| Mild | 4.28 ± 0.05 | 1.47 ± 0.02 | 0.13 ± 0.00 | 1.56 ± 0.34 | 44.35 ± 1.11 | 125.62 ± 12.89 | 1.93 ± 0.23 |
| Severe | 3.61 ± 0.02 | 1.41 ± 0.03 | 0.13 ± 0.01 | 0.86 ± 0.05 | 48.52 ± 0.99 | 19.50 ± 4.46 | 0.27 ± 0.06 |
| Treatment | RB | SRL | SRA | RTD | N | P | Al | Ca | Ca/Al |
|---|---|---|---|---|---|---|---|---|---|
| CaCO3 | ns | <0.01 | <0.001 | ns | <0.001 | <0.001 | ns | <0.001 | <0.001 |
| Root | <0.001 | <0.001 | <0.001 | <0.01 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Acid | <0.001 | ns | ns | ns | ns | <0.05 | <0.001 | ns | ns |
| Ca × R | ns | <0.05 | ns | ns | ns | ns | <0.05 | ns | ns |
| Ca × A | <0.01 | ns | ns | <0.05 | ns | ns | ns | ns | <0.05 |
| R × A | ns | ns | ns | ns | ns | ns | <0.001 | ns | ns |
| Ca × R × A | ns | ns | ns | ns | ns | ns | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Guan, X.; Xiao, F.; Zhang, W.; Yang, Q.; Wang, Q.; Wang, S.; Chen, L. Effects of Liming on the Morphologies and Nutrients of Different Functional Fine Roots of Cunninghamia lanceolata Seedlings. Forests 2022, 13, 822. https://doi.org/10.3390/f13060822
Yu X, Guan X, Xiao F, Zhang W, Yang Q, Wang Q, Wang S, Chen L. Effects of Liming on the Morphologies and Nutrients of Different Functional Fine Roots of Cunninghamia lanceolata Seedlings. Forests. 2022; 13(6):822. https://doi.org/10.3390/f13060822
Chicago/Turabian StyleYu, Xin, Xin Guan, Fuming Xiao, Weidong Zhang, Qingpeng Yang, Qingkui Wang, Silong Wang, and Longchi Chen. 2022. "Effects of Liming on the Morphologies and Nutrients of Different Functional Fine Roots of Cunninghamia lanceolata Seedlings" Forests 13, no. 6: 822. https://doi.org/10.3390/f13060822
APA StyleYu, X., Guan, X., Xiao, F., Zhang, W., Yang, Q., Wang, Q., Wang, S., & Chen, L. (2022). Effects of Liming on the Morphologies and Nutrients of Different Functional Fine Roots of Cunninghamia lanceolata Seedlings. Forests, 13(6), 822. https://doi.org/10.3390/f13060822