Forest Phenology Shifts in Response to Climate Change over China–Mongolia–Russia International Economic Corridor



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data
2.3. Methods and Analyses
3. Results
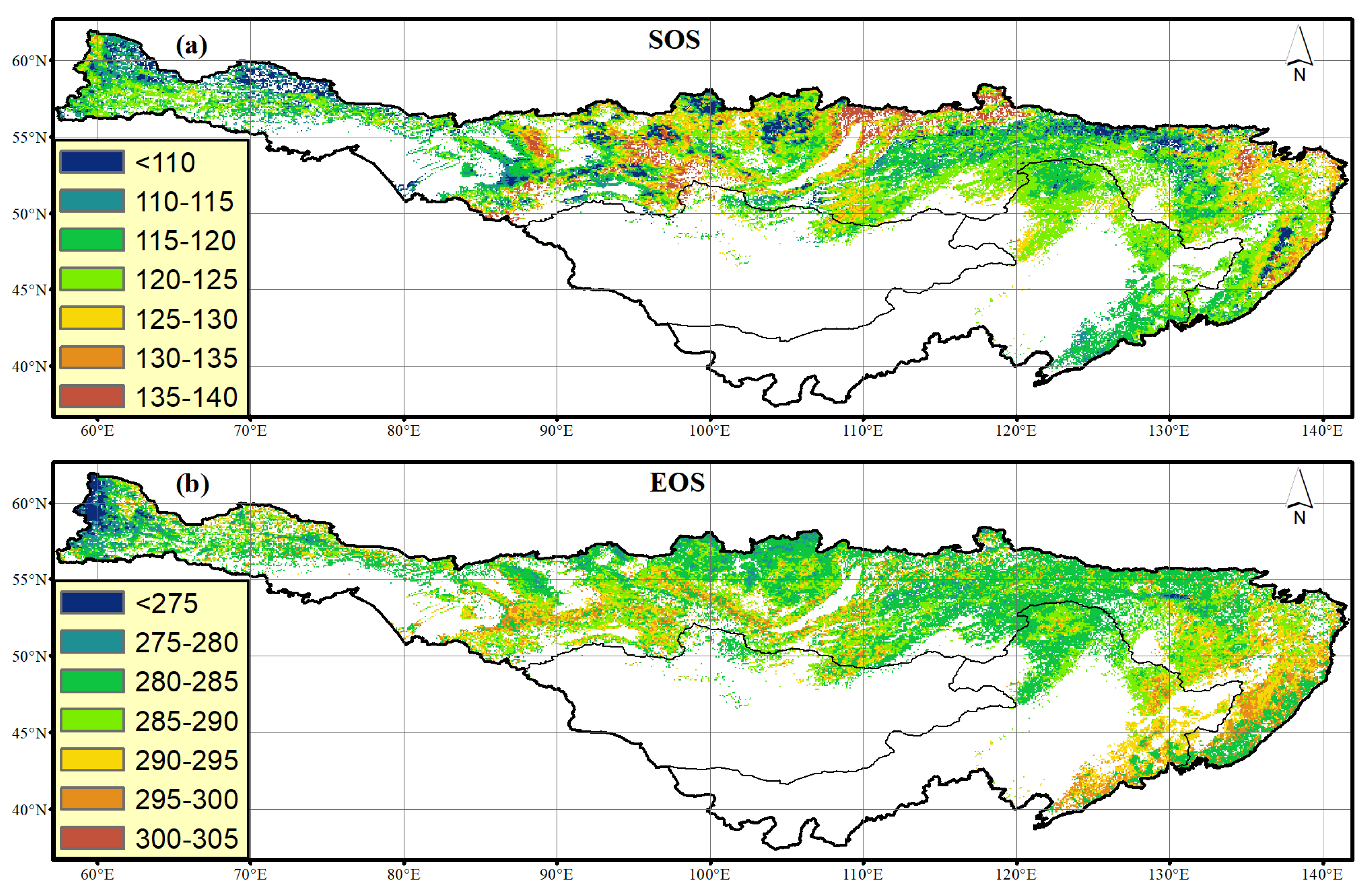
3.1. Characterization of Forest Phenology
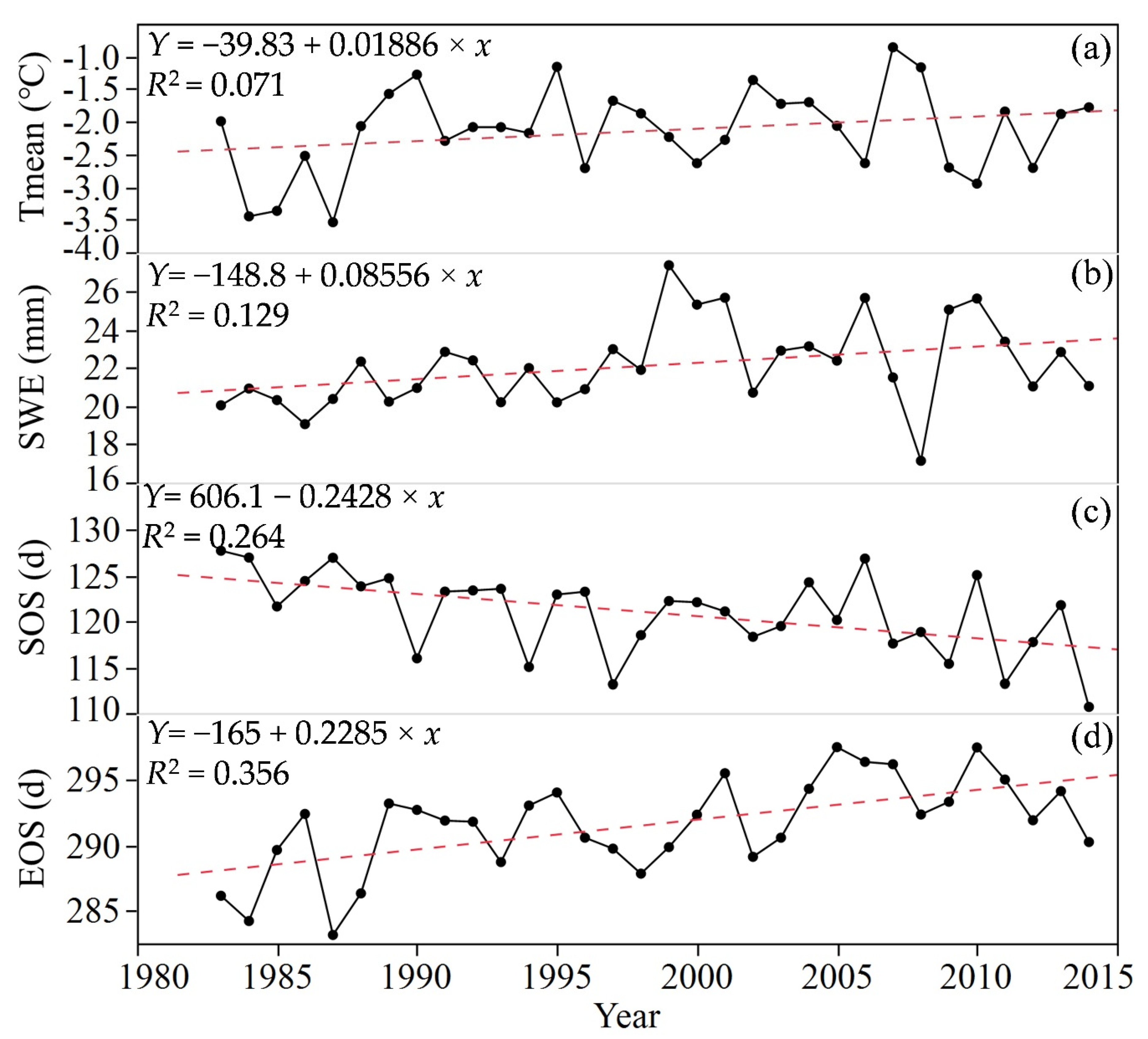
3.2. Trends in Growing Season of Forest Phenology
3.3. SOS and EOS Trends in Relation to Climate
4. Discussion
4.1. Trends and Spatiotemporal Variations in Forest Phenology
4.2. Relationship between SOS/EOS and Climatic Factors
4.3. Uncertainties and Future Works
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Menzel, A.; Fabian, P. Growing Season Extended in Europe. Nature 1999, 397, 659. [Google Scholar] [CrossRef]
- Stockli, R.; Vidale, P.L. European Plant Phenology and Climate as Seen in a 20-Year Avhrr Land-Surface Parameter Dataset. Int. J. Remote Sens. 2004, 25, 3303–3330. [Google Scholar] [CrossRef]
- Christiansen, D.E.; Markstrom, S.L.; Hay, L.E. Impacts of Climate Change on the Growing Season in the United States. Earth Interact. 2011, 15, 1–17. [Google Scholar] [CrossRef]
- Cook, K.H.; Vizy, E.K. Impact of Climate Change on Mid-Twenty-First Century Growing Seasons in Africa. Clim. Dyn. 2012, 39, 2937–2955. [Google Scholar] [CrossRef]
- Vujadinovic, M.; Vukovic, A.; Djurdjevic, V.; Rankovic-Vasic, Z.; Atanackovic, Z.; Sivcev, B.; Markovic, N.; Petrovic, N. Impact of Climate Change on Growing Season and Dormant Period Characteristics for the Balkan Region. Acta Hortic. 2012, 931, 87–94. [Google Scholar] [CrossRef]
- Bradley, N.L.; Leopold, A.C.; Ross, J.; Huffaker, W. Phenological Changes Reflect Climate Change in Wisconsin. Proc. Natl. Acad. Sci. USA 1999, 96, 9701–9704. [Google Scholar] [CrossRef]
- Euskirchen, E.S.; Mcguire, A.D.; Chapin, F.S. Energy Feedbacks of Northern High-Latitude Ecosystems to the Climate System Due to Reduced Snow Cover During 20th Century Warming. Glob. Chang. Biol. 2007, 13, 2425–2438. [Google Scholar] [CrossRef]
- Loarie, S.R.; Duffy, P.B.; Hamilton, H.; Asner, G.P.; Field, C.B.; Ackerly, D.D. The Velocity of Climate Change. Nature 2009, 462, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change. A Simple Method for Reconstructing a High-Quality Ndvi Time-Series Data Set Based on the Savitzky-Golay Filter. Remote Sens. Environ. 2013, 91, 332–344. [Google Scholar]
- Beerling, D.J.; Woodward, F.I. The Climate-Change Experiment (Climex)—Phenology and Gas-Exchange Responses of Boreal Vegetation to Global Change. Glob. Ecol. Biogeogr. Lett. 1994, 4, 17–26. [Google Scholar] [CrossRef]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased Plant Growth in the Northern High Latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- White, M.A.; Thornton, P.E.; Running, S.W. A Continental Phenology Model for Monitoring Vegetation Responses to Interannual Climatic Variability. Glob. Biogeochem. Cycles 1997, 11, 217–234. [Google Scholar] [CrossRef]
- Piao, S.L.; Fang, J.Y.; Zhou, L.M.; Ciais, P.; Zhu, B. Variations in Satellite-Derived Phenology in China‘s Temperate Vegetation. Glob. Chang. Biol. 2006, 12, 672–685. [Google Scholar] [CrossRef]
- Jeong, S.J.; Ho, C.H.; Gim, H.J.; Brown, M.E. Phenology Shifts at Start Vs. End of Growing Season in Temperate Vegetation over the Northern Hemisphere for the Period 1982-2008. Glob. Chang. Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H. Climate Controls on Vegetation Phenological Patterns in Northern Mid- and High Latitudes Inferred from Modis Data. Glob. Chang. Biol. 2004, 10, 1133–1145. [Google Scholar] [CrossRef]
- Piao, S.L.; Tan, J.G.; Chen, A.P.; Fu, Y.H.; Ciais, P.; Liu, Q.; Janssens, I.A.; Vicca, S.; Zeng, Z.Z.; Jeong, S.J.; et al. Pennuelas. Leaf Onset in the Northern Hemisphere Triggered by Daytime Temperature. Nat. Commun. 2015, 6, 6911. [Google Scholar] [CrossRef]
- Deng, G.R.; Zhang, H.Y.; Guo, X.Y.; Shan, Y.; Ying, H.; Wu, R.H.; Li, H.; Han, Y.L. Asymmetric Effects of Daytime and Nighttime Warming on Boreal Forest Spring Phenology. Remote Sens. 2019, 11, 1651. [Google Scholar] [CrossRef]
- Yu, H.; Luedeling, E.; Xu, J. Winter and Spring Warming Result in Delayed Spring Phenology on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2010, 107, 22151–22156. [Google Scholar] [CrossRef]
- Shen, M.; Piao, S.; Cong, N.; Zhang, G.; Jassens, I.A. Precipitation Impacts on Vegetation Spring Phenology on the Tibetan Plateau. Glob. Chang. Biol. 2015, 21, 3647–3656. [Google Scholar] [CrossRef]
- Shen, X.; Liu, B.; Henderson, M.; Wang, L.; Wu, Z.; Wu, H.; Jiang, M.; Lu, X. Asymmetric Effects of Daytime and Nighttime Warming on Spring Phenology in the Temperate Grasslands of China. Agric. For. Meteorol. 2018, 259, 240–249. [Google Scholar] [CrossRef]
- Groffman, P.M.; Driscoll, C.T.; Fahey, T.J.; Hardy, J.P.; Fitzhugh, R.D.; Tierney, G.L. Colder Soils in a Warmer World: A Snow Manipulation Study in a Northern Hardwood Forest Ecosystem. Biogeochemistry 2001, 56, 135–150. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Ciais, P.; Fang, J.; Wang, X. Change in Winter Snow Depth and Its Impacts on Vegetation in China. Glob. Chang. Biol. 2010, 16, 3004–3013. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate Change, Phenology, and Phenological Control of Vegetation Feedbacks to the Climate System. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Grippa, M.; Kergoat, L.; Le Toan, T.; Mognard, N.M.; Delbart, N.; L’Hermitte, J.; Vicente-Serrano, S.M. The Impact of Snow Depth and Snowmelt on the Vegetation Variability over Central Siberia. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Belt and Road Initiative. Overview-Belt and Road Initiative Forum 2019; Belt and Road Forum for International Cooperation: Beijing, China, 2019. [Google Scholar]
- Yu, L.X.; Liu, T.X.; Zhang, S.W. Temporal and Spatial Changes in Snow Cover and the Corresponding Radiative Forcing Analysis in Siberia from the 1970s to the 2010s. Adv. Meteorol. 2017, 2017, 1–11. [Google Scholar] [CrossRef]
- European Space Agency. Land Cover Cci Product User Guide Version 2. Tech. Rep. Available online: Maps.Elie.Ucl.Ac.Be/Cci/Viewer/Download/Esacci-Lc-Ph2-Pugv2_2.0.Pdf (accessed on 25 March 2020).
- Tucker, C.; Pinzón, J.; Brown, M.E.; Slayback, D.; Pak, E.W.; Mahoney, R.; Vermote, E.; Saleous, N. An Extended Avhrr 8-Km Ndvi Dataset Compatible with Modis and Spot Vegetation Ndvi Data. Int. J. Remote.Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C. Sensitivity of vegetation phenology detection to the temporal resolution of satellite data. Int. J. Remot. Sens. 2009, 30, 2061–2074. [Google Scholar] [CrossRef]
- Duveiller, G.; Hooker, J.; Cescatti, A. The mark of vegetation change on Earth’s surface energy balance. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Li, C.; Li, M.; Liu, J.; Li, Y.; Dai, Q.S. Comparative Analysis of Seasonal Landsat 8 Images for Forest Aboveground Biomass Estimation in a Subtropical Forest. Forests 2019, 11, 45. [Google Scholar] [CrossRef]
- Qiu, T.; Song, C.; Zhang, Y.; Liu, H.; Vose, J.M. Urbanization and climate change jointly shift land surface phenology in the northern mid-latitude large cities. Remote Sens. Environ. 2020, 236, 111477. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, S.; Xu, X.; Bu, K.; Ning, J.; Chang, L. High resolution land cover datasets integration and application based on Landsat and Globcover data from 1975 to 2010 in Siberia. Chin. Geogr. Sci. 2016, 26, 429–438. [Google Scholar] [CrossRef]
- Liu, J.; Kuang, W.; Zhang, Z.; Xu, X.; Qin, Y.; Ning, J.; Zhou, W.; Zhang, S.; Li, R.; Yan, C.; et al. Spatiotemporal characteristics, patterns, and causes of land-use changes in China since the late 1980s. J. Geogr. Sci. 2014, 24, 195–210. [Google Scholar] [CrossRef]
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Luojus, K. European Space Agency (Esa) Globsnow Snow Water Equivalent (Swe) V2.0 L3b Monthly Data (1979–2013); Finnish Meteorological Institute: Helsinki, Finland, 2015.
- Zhang, X.; Friedl, M.A.; Schaaf, C.; Strahler, A.H.; Hodges, J.C.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Liang, L.; Schwartz, M.; Fei, S. Validating satellite phenology through intensive ground observation and landscape scaling in a mixed seasonal forest. Remote Sens. Environ. 2011, 115, 143–157. [Google Scholar] [CrossRef]
- Wu, C.; Gonsamo, A.; Gough, C.M.; Chen, J.M.; Xu, S. Modeling growing season phenology in North American forests using seasonal mean vegetation indices from MODIS. Remote Sens. Environ. 2014, 147, 79–88. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Atzberger, C.; Høgda, K.A.; Johansen, B.; Skidmore, A. Improved monitoring of vegetation dynamics at very high latitudes: A new method using Modis Ndvi. Remote Sens. Environ. 2006, 100, 321–334. [Google Scholar] [CrossRef]
- Yu, L.; Liu, T.; Bu, K.; Yan, F.; Yang, J.; Chang, L.; Zhang, S. Monitoring the long term vegetation phenology change in Northeast China from 1982 to 2015. Sci. Rep. 2017, 7, 14770. [Google Scholar] [CrossRef]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Ciais, P.; Myneni, R.; Chen, A.; Chevallier, F.; Dolman, H.; Janssens, I.A.; Peñuelas, J.; Zhang, G.; et al. Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature 2013, 501, 88–92. [Google Scholar] [CrossRef]
- Shabanov, N.; Zhou, L.; Knyazikhin, Y.; Myneni, R.; Tucker, C. Analysis of interannual changes in northern vegetation activity observed in AVHRR data from 1981 to 1994. IEEE Trans. Geosci. Remote Sens. 2002, 40, 115–130. [Google Scholar] [CrossRef]
- Dye, D.G.; Tucker, C.J. Seasonality and trends of snow-cover, vegetation index, and temperature in northern Eurasia. Geophys. Res. Lett. 2003, 30, 30. [Google Scholar] [CrossRef]
- Doi, H.; Takahashi, M. Latitudinal patterns in the phenological responses of leaf colouring and leaf fall to climate change in Japan. Glob. Ecol. Biogeogr. 2008, 17, 556–561. [Google Scholar] [CrossRef]
- Estrella, N.; Menzel, A. Responses of leaf colouring in four deciduous tree species to climate and weather in Germany. Clim. Res. 2006, 32, 253–267. [Google Scholar] [CrossRef]
- Zhang, X.; Tarpley, D.; Sullivan, J.T. Diverse responses of vegetation phenology to a warming climate. Geophys. Res. Lett. 2007, 34, 34. [Google Scholar] [CrossRef]
- Migliavacca, M.; Sonnentag, O.; Keenan, T.F.; Cescatti, A.; O’Keefe, J.; Richardson, A.D.; O’keefe, J. On the uncertainty of phenological responses to climate change, and implications for a terrestrial biosphere model. Biogeosciences 2012, 9, 2063–2083. [Google Scholar] [CrossRef]
- Nichol, C.J.; Lloyd, J.; Shibistova, O.; Arneth, A.; Roser, C.; Knohl, A.; Matsubara, S.; Grace, J. Remote Sensing of Photosynthetic-Light-Use Efficiency of a Siberian Boreal Forest. Tellus Ser. B-Chem. Phys. Meteorol. 2002, 54, 677–687. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and Climate Change: Forcings, Feedbacks, and the Climate Benefits of Forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Forest Type | Count | Area (104 km2) | SOS (mean) | SOS (std) | EOS (mean) | EOS (std) |
|---|---|---|---|---|---|---|---|
| 1 | Broadleaved deciduous forest | 16,996 | 118.03 | 120.67 | 5.28 | 291.95 | 9.07 |
| 2 | Needleleaved evergreen forest | 9695 | 67.33 | 119.73 | 11.08 | 290.75 | 8.94 |
| 3 | Needleleaved deciduous forest | 26,068 | 181.03 | 121.81 | 7.53 | 291.22 | 9.86 |
| 4 | Mixed forest | 5080 | 35.28 | 120.02 | 8.25 | 288.07 | 9.08 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Yan, Z.; Zhang, S. Forest Phenology Shifts in Response to Climate Change over China–Mongolia–Russia International Economic Corridor. Forests 2020, 11, 757. https://doi.org/10.3390/f11070757
Yu L, Yan Z, Zhang S. Forest Phenology Shifts in Response to Climate Change over China–Mongolia–Russia International Economic Corridor. Forests. 2020; 11(7):757. https://doi.org/10.3390/f11070757
Chicago/Turabian StyleYu, Lingxue, Zhuoran Yan, and Shuwen Zhang. 2020. "Forest Phenology Shifts in Response to Climate Change over China–Mongolia–Russia International Economic Corridor" Forests 11, no. 7: 757. https://doi.org/10.3390/f11070757
APA StyleYu, L., Yan, Z., & Zhang, S. (2020). Forest Phenology Shifts in Response to Climate Change over China–Mongolia–Russia International Economic Corridor. Forests, 11(7), 757. https://doi.org/10.3390/f11070757