Most Southern Scots Pine Populations Are Locally Adapted to Drought for Tree Height Growth

 , , and
, , and
Abstract
1. Introduction
2. Material and Methods
2.1. Phenotypic Data and Common Gardens
2.2. Climate Data
2.3. Selection of Climate Variables
2.4. Linear Mixed-Effect Models of Tree Height Growth
α5(clim_s)ik × (clim_p)ij + β1(Planting.site/Block) + β2 (Population) + εijk,
2.5. Assessment of the Local Adaptation of Tree Height Growth for Spanish Scots Pine Populations under the Current Climate
2.6. Spatial Predictions of the Local Adaptation of Tree Height Growth for Southern Scots Pine Populations under the Impacts of the Current and Future Climate
3. Results
3.1. Linear Mixed-Effect Models of Tree Height Growth
3.2. Assessment of the Local Adaptation of Tree Height Growth for Spanish Scots Pine Populations under the Current Climate
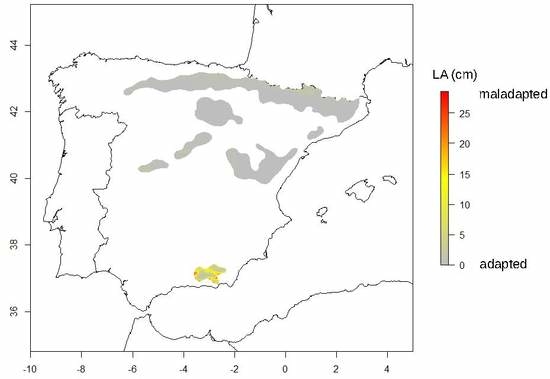
3.3. Spatial Predictions of the Local Adaptation of Tree Height Growth for Southern Scots Pine Populations under the Impacts of the Current and Future Climate
4. Discussion
4.1. The Main Climatic Drivers Shaping Among-Population Differentiation and Phenotypic Plasticity Responses of Tree Height Growth
4.2. Most Southern Scots Pine Populations are Locally Adapted to the Current Climate
4.3. The Importance of Considering Genetic and Plastic Effects for Evaluating Tree Height Growth for Future Climates
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Climate at the Planting Site | Climate at the Population Origin | ||||
|---|---|---|---|---|---|
| Variables | Pearson | Spearman | Variables | Pearson | Spearman |
| TMF.jan | −0.565 | −0.384 | TMF.jan | 0.025 | 0.019 |
| TMF.feb | −0.581 | −0.497 | TMF.feb | 0.006 | 0 |
| TMF.mar | −0.62 | −0.554 | TMF.mar | 0.007 | −0.002 |
| TMF.apr | −0.65 | −0.616 | TMF.apr | 0.004 | −0.004 |
| TMF.may | −0.371 | −0.382 | TMF.may | −0.007 | −0.01 |
| TMF.jun | −0.055 | −0.111 | TMF.jun | −0.002 | 0.003 |
| TMF.jul | −0.107 | −0.038 | TMF.jul | −0.016 | −0.025 |
| TMF.ago | −0.171 | 0.009 | TMF.ago | −0.021 | −0.036 |
| TMF.sep | −0.534 | −0.536 | TMF.sep | −0.014 | −0.029 |
| TMF.oct | −0.633 | −0.554 | TMF.oct | 0.005 | −0.009 |
| TMF.nov | −0.636 | −0.594 | TMF.nov | 0.01 | 0.006 |
| TMF.dec | −0.577 | −0.479 | TMF.dec | 0.024 | 0.01 |
| TMC.jan | −0.32 | −0.174 | TMC.jan | −0.017 | −0.031 |
| TMC.feb | −0.478 | −0.47 | TMC.feb | −0.03 | −0.023 |
| TMC.mar | −0.628 | −0.644 | TMC.mar | −0.043 | −0.039 |
| TMC.apr | −0.648 | −0.644 | TMC.apr | −0.034 | −0.037 |
| TMC.may | −0.607 | −0.695 | TMC.may | −0.055 | −0.057 |
| TMC.jun | −0.469 | −0.345 | TMC.jun | −0.055 | −0.064 |
| TMC.jul | −0.494 | −0.478 | TMC.jul | −0.075 | −0.087 |
| TMC.ago | −0.379 | −0.378 | TMC.ago | −0.075 | −0.086 |
| TMC.sep | −0.438 | −0.44 | TMC.sep | −0.077 | −0.086 |
| TMC.oct | −0.443 | −0.521 | TMC.oct | −0.034 | −0.024 |
| TMC.nov | −0.269 | −0.208 | TMC.nov | −0.028 | −0.027 |
| TMC.dec | −0.287 | −0.174 | TMC.dec | −0.016 | −0.014 |
| T.jan | −0.457 | −0.407 | T.jan | 0.003 | 0 |
| T.feb | −0.569 | −0.497 | T.feb | −0.014 | −0.018 |
| T.mar | −0.66 | −0.684 | T.mar | −0.023 | −0.022 |
| T.apr | −0.648 | −0.616 | T.apr | −0.019 | −0.027 |
| T.may | −0.586 | −0.616 | T.may | −0.034 | −0.037 |
| T.jun | −0.337 | −0.345 | T.jun | −0.03 | −0.029 |
| T.jul | −0.358 | −0.282 | T.jul | −0.048 | −0.052 |
| T.ago | −0.311 | −0.378 | T.ago | −0.051 | −0.049 |
| T.sep | −0.492 | −0.378 | T.sep | −0.05 | −0.042 |
| T.oct | −0.534 | −0.521 | T.oct | −0.016 | −0.027 |
| T.nov | −0.466 | −0.521 | T.nov | −0.011 | −0.023 |
| T.dec | −0.426 | −0.407 | T.dec | 0.001 | −0.006 |
| PREC.annl | 0.675 | 0.663 | PREC.ann | 0.073 | 0.105 |
| PREC.win | 0.575 | 0.516 | PREC.win | 0.035 | 0.038 |
| PREC.spr | 0.779 | 0.663 | PREC.spr | 0.073 | 0.088 |
| PREC.sum | 0.609 | 0.497 | PREC.sum | 0.08 | 0.079 |
| PREC.son | 0.611 | 0.487 | PREC.son | 0.079 | 0.107 |
| MT | −0.544 | −0.458 | MT | −0.024 | −0.031 |
| MCMT | −0.262 | −0.282 | MCMT | −0.056 | −0.057 |
| MAXWMT | −0.416 | −0.378 | MAXWMT | −0.072 | −0.083 |
| MCMT | −0.446 | −0.407 | MCMT | 0.01 | 0.007 |
| MINCMT | −0.561 | −0.351 | MINCMT | 0.031 | 0.027 |
| FP | 0.651 | 0.594 | FP | −0.016 | 0.009 |
| DD5 | −0.613 | −0.548 | DD5 | −0.004 | −0.012 |
| TD | 0.282 | 0.33 | TD | −0.103 | −0.107 |
| AHM | −0.611 | −0.663 | AHM | −0.083 | −0.087 |
| SHM | −0.376 | −0.554 | SHM | −0.108 | −0.103 |
| PREC.ann_p | PREC.sum_p | PREC.aut_p | TD_p | SHM_p | |
|---|---|---|---|---|---|
| TMF.jan_s | 49,559.63 | 49,551.74 | 49,554.15 | 49,557.09 | 49,546.44 |
| TMF.feb_s | 49,560.72 | 49,551.38 | 49,555.08 | 49,557.30 | 49,547.21 |
| TMF.mar_s | 49,560.71 | 49,550.99 | 49,555.01 | 49,557.11 | 49,546.92 |
| TMF.apr_s | 49,560.68 | 49,551.40 | 49,555.01 | 49,557.05 | 49,546.58 |
| TMF.sep_s | 49,559.38 | 49,550.93 | 49,554.04 | 49,557.11 | 49,546.46 |
| TMF.ocT_s | 49,560.05 | 49,550.89 | 49,554.45 | 49,556.87 | 49,546.21 |
| TMF.nov_s | 49,560.59 | 49,550.80 | 49,554.90 | 49,557.00 | 49,546.73 |
| TMF.dec_s | 49,558.35 | 49,551.16 | 49,553.06 | 49,556.43 | 49,545.56 |
| TMC.mar_s | 49,559.41 | 49,551.82 | 49,553.99 | 49,556.32 | 49,545.58 |
| TMC.apr_s | 49,560.07 | 49,551.58 | 49,554.50 | 49,556.48 | 49,545.95 |
| TMC.may_s | 49,558.67 | 49,550.63 | 49,553.19 | 49,555.64 | 49,544.68 |
| TMC.ocT_s | 49,560.10 | 49,551.94 | 49,554.42 | 49,557.14 | 49,546.54 |
| T.feb_s | 49,560.11 | 49,551.59 | 49,554.50 | 49,557.14 | 49,546.43 |
| T.mar_s | 49,559.29 | 49,550.86 | 49,553.77 | 49,556.02 | 49,545.26 |
| T.abr_s | 49,560.58 | 49,551.67 | 49,554.94 | 49,556.90 | 49,546.43 |
| T.may_s | 49,559.93 | 49,551.30 | 49,554.34 | 49,556.65 | 49,545.82 |
| T.ocT_s | 49,560.85 | 49,552.14 | 49,555.19 | 49,557.78 | 49,547.12 |
| T.nov_s | 49,560.74 | 49,552.16 | 49,554.99 | 49,557.50 | 49,547.12 |
| PREC.ann_s | 49,556.66 | 49,547.51 | 49,551.56 | 49,554.96 | 49,542.98 |
| PREC.win_s | 49,557.89 | 49,548.82 | 49,553.03 | 49,557.20 | 49,544.34 |
| PREC.spr_s | 49,551.91 | 49,542.89 | 49,546.79 | 49,549.92 | 49,537.76 |
| PREC.sum_s | 49,559.40 | 49,550.01 | 49,553.93 | 49,556.48 | 49,545.86 |
| PREC.aut_s | 49,558.02 | 49,548.82 | 49,552.83 | 49,555.99 | 49,544.56 |
| TM_s | 49,560.08 | 49,551.91 | 49,554.55 | 49,557.53 | 49,546.59 |
| MINCMT_s | 49,559.40 | 49,551.46 | 49,553.88 | 49,556.69 | 49,546.14 |
| FP_s | 49,559.60 | 49550.39 | 49,553.99 | 49,556.29 | 49,545.69 |
| DD5_s | 49,559.15 | 49,550.99 | 49,553.67 | 49,556.52 | 49,545.53 |
| AHM_s | 49,553.34 | 49,546.99 | 49,548.73 | 49,553.97 | 49,541.36 |
| SHM_s | 49,557.06 | 49,551.09 | 49,551.97 | 49,556.55 | 49,545.83 |



References
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef] [PubMed]
- McMahon, S.M.; Parker, G.G.; Miller, D.R. Evidence for a recent increase in forest growth. Proc. Natl. Acad. Sci. USA 2010, 107, 3611–3615. [Google Scholar] [CrossRef] [PubMed]
- Machar, I.; Vlckova, V.; Bucek, A.; Vozenilek, V.; Salek, L.; Jerabkova, L.; Machar, I.; Vlckova, V.; Bucek, A.; Vozenilek, V.; et al. Modelling of Climate Conditions in Forest Vegetation Zones as a Support Tool for Forest Management Strategy in European Beech Dominated Forests. Forests 2017, 8, 82. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Lévesque, M.; Siegwolf, R.; Saurer, M.; Eilmann, B.; Rigling, A. Increased water-use efficiency does not lead to enhanced tree growth under xeric and mesic conditions. New Phytol. 2014, 203, 94–109. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; García-Valdés, R.; Ruiz-Benito, P.; Zavala, M.A. Disentangling the relative importance of climate, size and competition on tree growth in Iberian forests: Implications for forest management under global change. Glob. Chang. Biol. 2011, 17, 2400–2414. [Google Scholar] [CrossRef]
- Kunstler, G.; Falster, D.; Coomes, D.A.; Hui, F.; Kooyman, R.M.; Laughlin, D.C.; Poorter, L.; Vanderwel, M.; Vieilledent, G.; Wright, S.J.; et al. Plant functional traits have globally consistent effects on competition. Nature 2016, 529, 204–207. [Google Scholar]
- Cornelius, J. Heritabilities and additive genetic coefficients of variation in forest trees. Can. J. For. Res. 1994, 24, 372–379. [Google Scholar] [CrossRef]
- Leites, L.P.; Robinson, A.P.; Rehfeldt, G.E.; Marshall, J.D.; Crookston, N.L. Height-growth response to climatic changes differs among populations of Douglas-fir: A novel analysis of historic data. Ecol. Appl. 2012, 22, 154–165. [Google Scholar] [CrossRef]
- Rehfeldt, G.; Ying, C.; Spittlehouse, D.; Hamilton, D. Genetic responses to climate in Pinus contorta: Niche breadth, climate change, and reforestation. Ecol. Monogr. 1999, 69, 375–407. [Google Scholar] [CrossRef]
- Gárate-Escamilla, H.; Hampe, A.; Vizcaíno-Palomar, N.; Robson, T.M.; Benito Garzón, M. Range-wide variation in local adaptation and phenotypic plasticity of fitness-related traits in Fagus sylvatica and their implications under climate change. Glob. Ecol. Biogeogr. 2019. [Google Scholar] [CrossRef]
- Vizcaíno-Palomar, N.; Ibáñez, I.; González-Martínez, S.C.; Zavala, M.A.; Alía, R. Adaptation and plasticity in aboveground allometry variation of four pine species along environmental gradients. Ecol. Evol. 2016, 6, 7561–7573. [Google Scholar] [CrossRef]
- Wang, T.; O’Neill, G.A.; Aitken, S.N. Integrating environmental and genetic effects to predict responses of tree populations to climate. Ecol. Appl. 2010, 20, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene flow and local adaptation in trees. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef]
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef]
- Martínez-Meyer, E.; Díaz-Porras, D.; Peterson, A.T.; Yáñez-Arenas, C. Ecological niche structure and range wide abundance patterns of species. Biol. Lett. 2012, 9, 20120637. [Google Scholar] [CrossRef]
- Purves, D.W. The demography of range boundaries versus range cores in eastern US tree species. Proc. R. Soc. B Biol. Sci. 2009, 276, 1477–1484. [Google Scholar] [CrossRef]
- Pedlar, J.H.; McKenney, D.W.; Parra, J.; Jones, P.; Jarvis, A. Assessing the anticipated growth response of northern conifer populations to a warming climate. Sci. Rep. 2017, 7, 43881. [Google Scholar] [CrossRef]
- Doak, D.F.; Morris, W.F. Demographic compensation and tipping points in climate-induced range shifts. Nature 2010, 467, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Benito Garzón, M.; Ruiz-Benito, P.; Zavala, M.A. Interspecific differences in tree growth and mortality responses to environmental drivers determine potential species distributional limits in Iberian forests. Glob. Ecol. Biogeogr. 2013, 22, 1141–1151. [Google Scholar] [CrossRef]
- Peterson, M.L.; Doak, D.F.; Morris, W.F. Both life-history plasticity and local adaptation will shape range-wide responses to climate warming in the tundra plant Silene Acaulis. Glob. Chang. Biol. 2018, 24, 1614–1625. [Google Scholar] [CrossRef] [PubMed]
- Morgenstern, E.K. Geographic Variation in Forest Trees: Genetic Basis and Application of Knowledge in Silviculture; University of British Columbia: Vancouver, BC, Canada, 1996. [Google Scholar]
- Eckenwalder, J.E. Conifers of the World: The Complete Reference; Timber Press: Portland, OR, USA, 2009; ISBN 0881929743. [Google Scholar]
- Sánchez-Salguero, R.; Camarero, J.J.; Hevia, A.; Madrigal-González, J.; Linares, J.C.; Ballesteros-Cánovas, J.A.; Sánchez-Miranda, A.; Alfaro-Sánchez, R.; Sangüesa-Barreda, G.; Galván, J.D.; et al. What drives growth of Scots pine in continental Mediterranean climates: Drought, low temperatures or both? Agric. For. Meteorol. 2015, 206, 151–162. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Camarero, J.J.; Gutiérrez, E.; González Rouco, F.; Gazol, A.; Sangüesa-Barreda, G.; Andreu-Hayles, L.; Linares, J.C.; Seftigen, K. Assessing forest vulnerability to climate warming using a process-based model of tree growth: Bad prospects for rear-edges. Glob. Chang. Biol. 2017, 23, 2705–2719. [Google Scholar] [CrossRef] [PubMed]
- Kujala, S.T.; Savolainen, O. Sequence variation patterns along a latitudinal cline in Scots pine (Pinus sylvestris): Signs of clinal adaptation? Tree Genet. Genomes 2012, 8, 1451–1467. [Google Scholar] [CrossRef]
- Soranzo, N.; Alía, R.; Provan, J.; Powell, W. Patterns of variation at a mitochondrial sequence-tagged-site locus provides new insights into the postglacial history of European Pinus sylvestris populations. Mol. Ecol. 2000, 9, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Robledo-Arnuncio, J.J.; Collada, C.; Alía, R.; Gil, L. Genetic structure of montane isolates of Pinus sylvestris L. in a Mediterranean refugial area. J. Biogeogr. 2005, 32, 595–605. [Google Scholar] [CrossRef]
- Prus-Glowacki, W.; Stephan, B.R.; Bujas, E.; Alía, R.; Marciniak, A. Genetic differentiation of autochthonous populations of Pinus sylvestris (Pinaceae) from the Iberian Peninsula. Plant Syst. Evol. 2003, 239, 55–66. [Google Scholar] [CrossRef]
- Nahal, I. The Mediterranean climate from a biological viewpoint. In Ecosystems of the World 11: Mediterranean-Type Shrublands; Di Castri, F., Goodall, W., Specht, R.L., Eds.; Elsevier Scientific Publishing Co.: Amsterdam, The Netherlands, 1981; pp. 63–86. [Google Scholar]
- Sánchez-Salguero, R.; Navarro-Cerrillo, R.M.; Camarero, J.J.; Fernández-Cancio, Á. Selective drought-induced decline of pine species in southeastern Spain. Clim. Chang. 2012, 113, 767–785. [Google Scholar] [CrossRef]
- Castro, J.; Zamora, R.; Hódar, A.J.; Gómez, J. Seedling establishment of a boreal tree species (Pinus sylvestris) at its southernmost distribution limit: Consequences of being in a marginal Mediterranean habitat. J. Ecol. 2004, 92, 266–277. [Google Scholar] [CrossRef]
- Herrero, A.; Rigling, A.; Zamora, R. Varying climate sensitivity at the dry distribution edge of Pinus sylvestris and P. nigra. For. Ecol. Manag. 2013, 308, 50–61. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Falster, D.S.; Westoby, M. Plant height and evolutionary games. Trends Ecol. Evol. 2003, 18, 337–343. [Google Scholar] [CrossRef]
- Moles, A.T.; Leishman, M.R. The seedling as part of a plant’s life history strategy. In Seedling Ecology and Evolution; Leck, A., Parker, V.T., Simpson, R.L., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 217–238. [Google Scholar]
- Alía, R.; Moro, J.; Notivol, E.; Moro-Serrano, J. Genetic variability of Scots pine (Pinus sylvestris L.) provenances in Spain: Growth traits and survival. Silva Fenn. 2001, 35, 27–38. [Google Scholar] [CrossRef]
- Agúndez, D.; Alía, R.; Diez, R. Variación de Pinus sylvestris en España: Características de piñas y piñones. Investig. Agrar. Sist. Recur. For. 1992, 1, 151–162. [Google Scholar]
- González-Martínez, S.C.; Burczyk, J.; Nathan, R.; Nanos, N.; Gil, L.; Alía, R. Effective gene dispersal and female reproductive success in Mediterranean maritime pine (Pinus pinaster Aiton). Mol. Ecol. 2006, 15, 4577–4588. [Google Scholar] [CrossRef]
- Gonzalo-Jiménez, J. Diagnosis Fitoclimática de la España Peninsular Hacia un Modelo de Clasificación Funcional de la Vegetación y de los Ecosistemas Peninsulares Españoles; Organismo Autónomo Parques Nacionales: Madrid, Spain, 2010; ISBN 978-84-8014-787-3. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- IPCC Climate Change. Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In Breakthroughs in Statistics; Kotz, S., Johnson, N., Eds.; Springer-Verlag: London, UK, 1992; pp. 610–624. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-0-387-87458-6. [Google Scholar]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Rune, H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Br. 2017, 12, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.-F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Agúndez, D.; Alía, R.; Stephan, R.; Gil, L.; Pardos, J.A. Ensayo de Procedencias Españolas y Alemanas de Pinus Sylvestris L.: Comportamiento en Vivero y Supervivencia en Monte. Ecología 1994, 8, 245–257. [Google Scholar]
- Valladares, F.; Gianoli, E.; Gómez, J.M. Ecological limits to plant phenotypic plasticity. New Phytol. 2007, 176, 749–763. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Gómez, J.M.; García, D.; Zamora, R.; Hódar, J.A. Seed predation and dispersal in relict Scots pine forests in southern Spain. Plant Ecol. 1999, 145, 115–123. [Google Scholar] [CrossRef]
- Benito Garzón, M.; Robson, T.M.; Hampe, A. ΔTrait SDMs: Species distribution models that account for local adaptation and phenotypic plasticity. New Phytol. 2019, 222, 1757–1765. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.; Feild, T.; Gleason, S.; Hacke, U.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Montes, F.; Gavilán, R.G.; Cañellas, I.; Rubio, A. Is this the end? Dynamics of a relict stand from pervasively deforested ancient Iberian pine forests. Eur. J. For. Res. 2015, 134, 525–536. [Google Scholar] [CrossRef]



| Temperature-related | Monthly temperature (T, °C) | T.jan, T.feb, T.mar, T.apr, T.may, T.jun, T.jul, T.ago, T.sep, T.oct, T.nov, T.dec |
| Mean annual temperature | MT | |
| Monthly minimum temperature (TMF, °C) | TMF.jan, TMF.feb, TMF.mar, TMF.apr, TMF.may, TMF.jun, TMF.jul, TMF.ago, TMF.sep, TMF.oct, TMF.nov, TMF.dec | |
| Temperature of the coldest month |
| |
| Monthly maximum temperature (TMC, °C) | TMC.jan, TMC.feb, TMC.mar, TMC.apr, TMC.may, TMC.jun, TMC.jul, TMC.ago, TMC.sep, TMC.oct, TMC.nov, TMC.dec | |
| Temperature of the warmest month |
| |
| Continentality (°C) | TD = MWMT − MCMT | |
| Growing degree-days with temperature above 5 ℃ | DD5 | |
| Frost period (month) | FP | |
| Precipitation-related | Annual precipitation (mm) | PREC.annl |
| Seasonal precipitation (mm) | PREC.win, PREC.spr, PREC.sum, PREC.son | |
| Annual heat-moisture index | AHM = MT + 10/(PREC.annl/1000), based upon [10] | |
| Summer heat-moisture index | SHM = MAXWMT/(PREC.sum/1000), based upon [10] |
| Fixed Effect Selection | d.f. | AIC | ∆AIC | MR2 | CR2 |
|---|---|---|---|---|---|
| (Round 1) Two-way interaction | |||||
| Tree height growth ~ full model | 10 | 50,802.8 | 0 | ||
| Tree height growth ~ full model minus interaction (PREC.spr_s × SHM_p) | 9 | 50,824.2 | 21.44 | ||
| * (Round 2) Quadratic effect planting site | |||||
| Tree height growth ~ full model | 10 | 50,802.8 | 0 | ||
| Tree height growth ~ full model minus quadratic effect (PREC.spr_s) | 9 | 50,801.4 | −1.31 | ||
| * (Round 3) Quadratic effect population | |||||
| Tree height growth ~ full model | 9 | 50,801.4 | 0 | ||
| Tree height growth ~ full model minus quadratic effect (SHM_p) | 8 | 50,804.2 | 2.77 | ||
| * (Round 4) Main effects | |||||
| Tree height growth ~ full model | 9 | 50,801.4 | 0 | 62.36 | 75.14 |
| Tree height growth ~ full model minus linear term (PREC.spr_s) | 7 | 50,835.4 | 33.94 | ||
| Tree height growth ~ full model minus linear term (no SHM_p) | 6 | 50,830.1 | 28.69 | ||
| Tree height growth ~ intercept model | 5 | 50,842.6 | 41.13 |
| Fixed Effects | ||||
| Mean | SE | t-Value | p-Value | |
| (Intercept) | 335.414 | 19.873 | 16.878 | 1.34 × 10−5 *** |
| PREC.spr_s | 124.186 | 19.189 | 6.472 | 2.96 × 10−3 ** |
| SHM_p | −9.091 | 5.694 | −1.597 | 1.34 × 10−1 n.s. |
| (SHM_p)2 | −8.562 | 3.894 | −2.199 | 4.64 × 10−2 * |
| PREC.spr_s × SHM_p | −5.794 | 1.195 | −4.848 | 1.29 × 10−6 *** |
| Random Effects | ||||
| SD | ||||
| Block/Planting site | 31.33 | |||
| Population | 20.2 | |||
| Planting site | 42.8 | |||
| Residual | 79.15 |
| Population | PREC.spr_p | CLL | HL (cm) | CLOPT | HOPT (cm) | LAH (cm) | CLH |
|---|---|---|---|---|---|---|---|
| Baza | 265 | 325.00 | 239.17 | 152.34 | 309.24 | 70.07 | −172.66 |
| Navarredonda de Gredos | 237 | 282.83 | 229.54 | 159.87 | 264.98 | 35.43 | −122.96 |
| Campisábalos | 211 | 239.66 | 211.75 | 166.87 | 224.09 | 12.34 | −72.79 |
| Galve de Sorbe | 227 | 221.77 | 241.20 | 162.56 | 249.22 | 8.02 | −59.21 |
| Morrano | 185 | 205.41 | 181.13 | 173.86 | 183.40 | 2.27 | −31.54 |
| Castell de Cabrés | 205 | 197.74 | 212.78 | 168.48 | 214.68 | 1.90 | −29.26 |
| La Cenia | 298 | 179.43 | 359.13 | 143.45 | 361.79 | 2.66 | −35.98 |
| Navafría | 327 | 175.18 | 404.95 | 135.66 | 408.12 | 3.17 | −39.52 |
| La Granja | 377 | 164.96 | 485.11 | 122.21 | 488.71 | 3.60 | −42.76 |
| Gúdar | 202 | 155.77 | 209.40 | 169.30 | 209.92 | 0.52 | 13.53 |
| San Zadornil | 267 | 154.55 | 312.41 | 151.80 | 312.41 | 0.00 | −2.74 |
| Covaleda | 327 | 148.13 | 407.92 | 135.66 | 408.12 | 0.20 | −12.47 |
| Orihuela del Tremedal | 297 | 147.56 | 360.13 | 143.73 | 360.13 | 0.00 | −3.83 |
| Puebla de Lillo | 477 | 118.18 | 651.63 | 95.28 | 652.34 | 0.70 | −22.90 |
| Borau | 400 | 92.37 | 524.20 | 116.01 | 526.11 | 1.92 | 23.64 |
| Pobla de Lillet | 226 | 91.69 | 234.84 | 162.84 | 247.59 | 12.75 | 71.15 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizcaíno-Palomar, N.; González-Muñoz, N.; González-Martínez, S.C.; Alía, R.; Benito Garzón, M. Most Southern Scots Pine Populations Are Locally Adapted to Drought for Tree Height Growth. Forests 2019, 10, 555. https://doi.org/10.3390/f10070555
Vizcaíno-Palomar N, González-Muñoz N, González-Martínez SC, Alía R, Benito Garzón M. Most Southern Scots Pine Populations Are Locally Adapted to Drought for Tree Height Growth. Forests. 2019; 10(7):555. https://doi.org/10.3390/f10070555
Chicago/Turabian StyleVizcaíno-Palomar, Natalia, Noelia González-Muñoz, Santiago C. González-Martínez, Ricardo Alía, and Marta Benito Garzón. 2019. "Most Southern Scots Pine Populations Are Locally Adapted to Drought for Tree Height Growth" Forests 10, no. 7: 555. https://doi.org/10.3390/f10070555
APA StyleVizcaíno-Palomar, N., González-Muñoz, N., González-Martínez, S. C., Alía, R., & Benito Garzón, M. (2019). Most Southern Scots Pine Populations Are Locally Adapted to Drought for Tree Height Growth. Forests, 10(7), 555. https://doi.org/10.3390/f10070555