Environmental Controls on the Seasonal Variation in Gas Exchange and Water Balance in a Near-Coastal Mediterranean Pinus halepensis Forest

 ,
,  , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Environmental Conditions
2.3. Measurement Campaigns
2.4. Gas Exchange and Needle Water Potential
2.5. Needle δ13C Signature
2.6. Tree Sap Flow and Canopy Stomatal Conductance
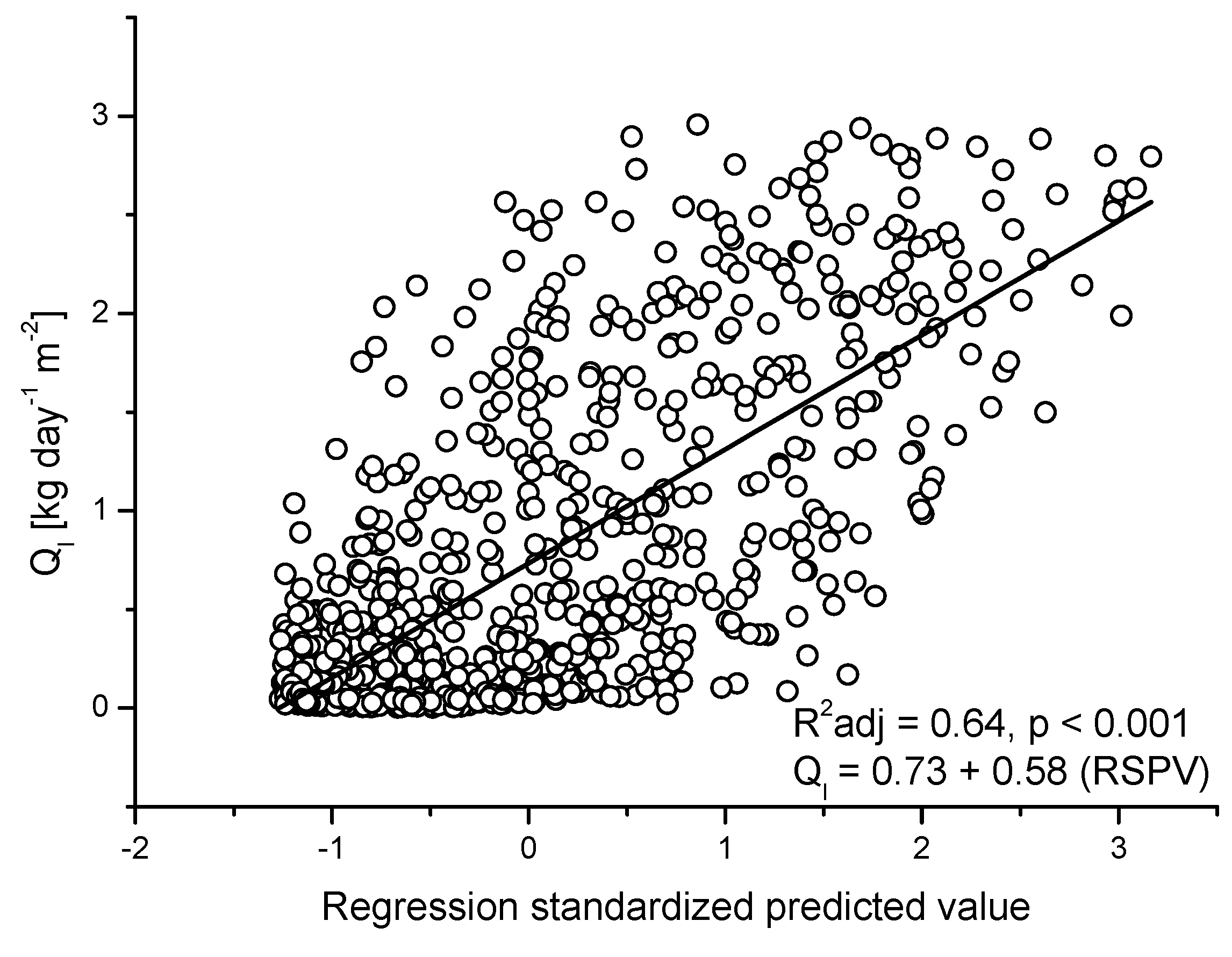
2.7. Statistical Analysis
3. Results
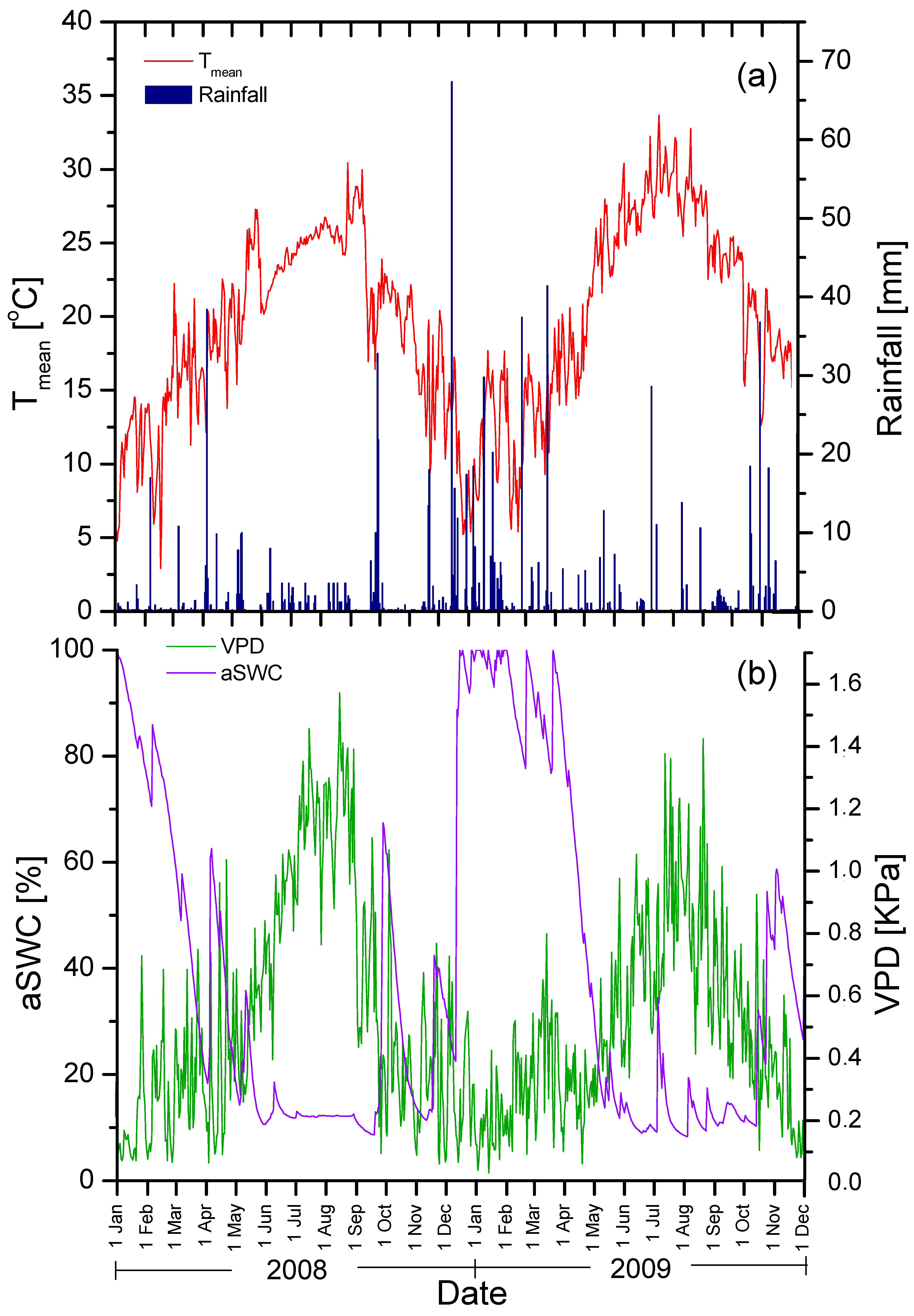
3.1. Climatic Conditions
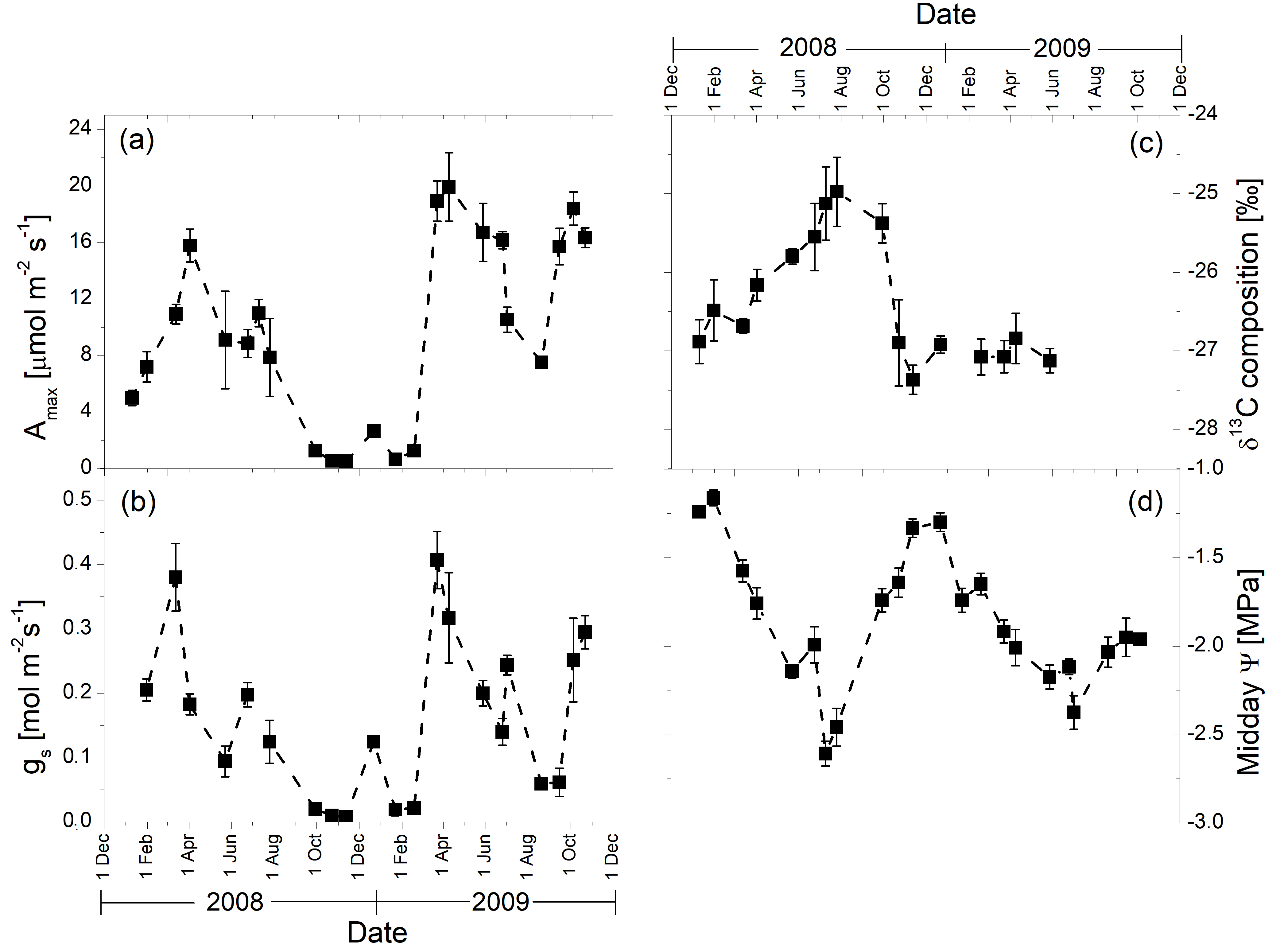
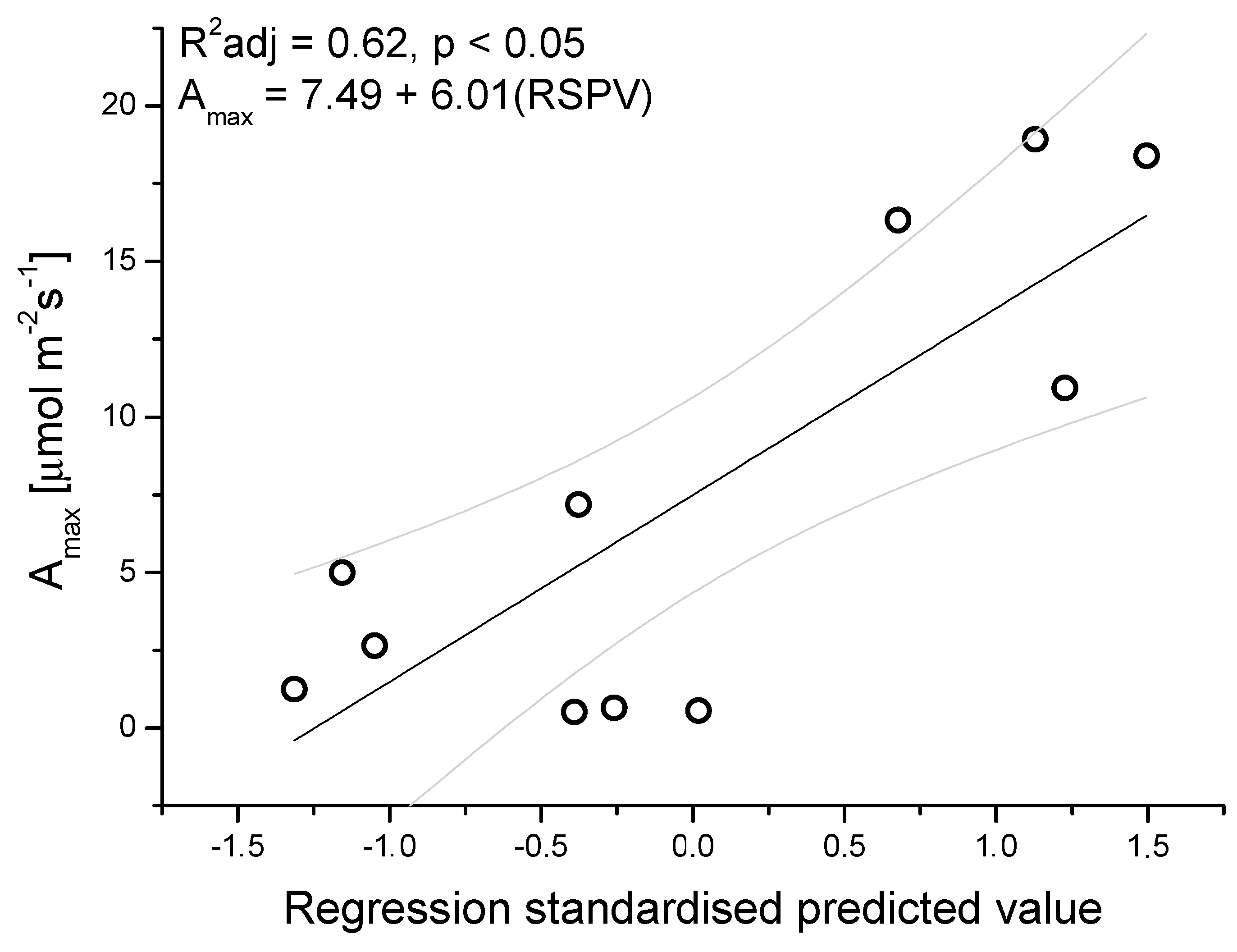
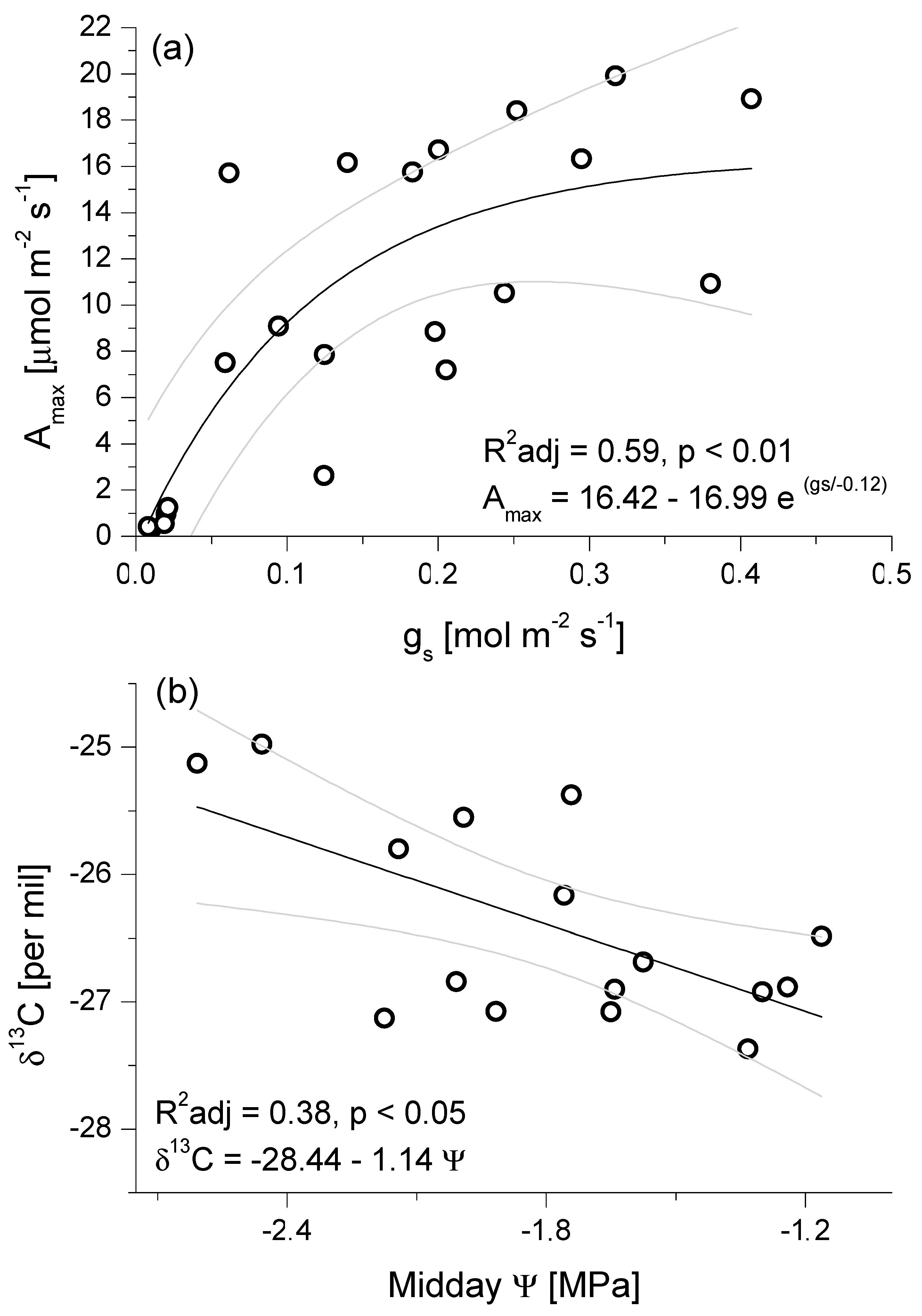
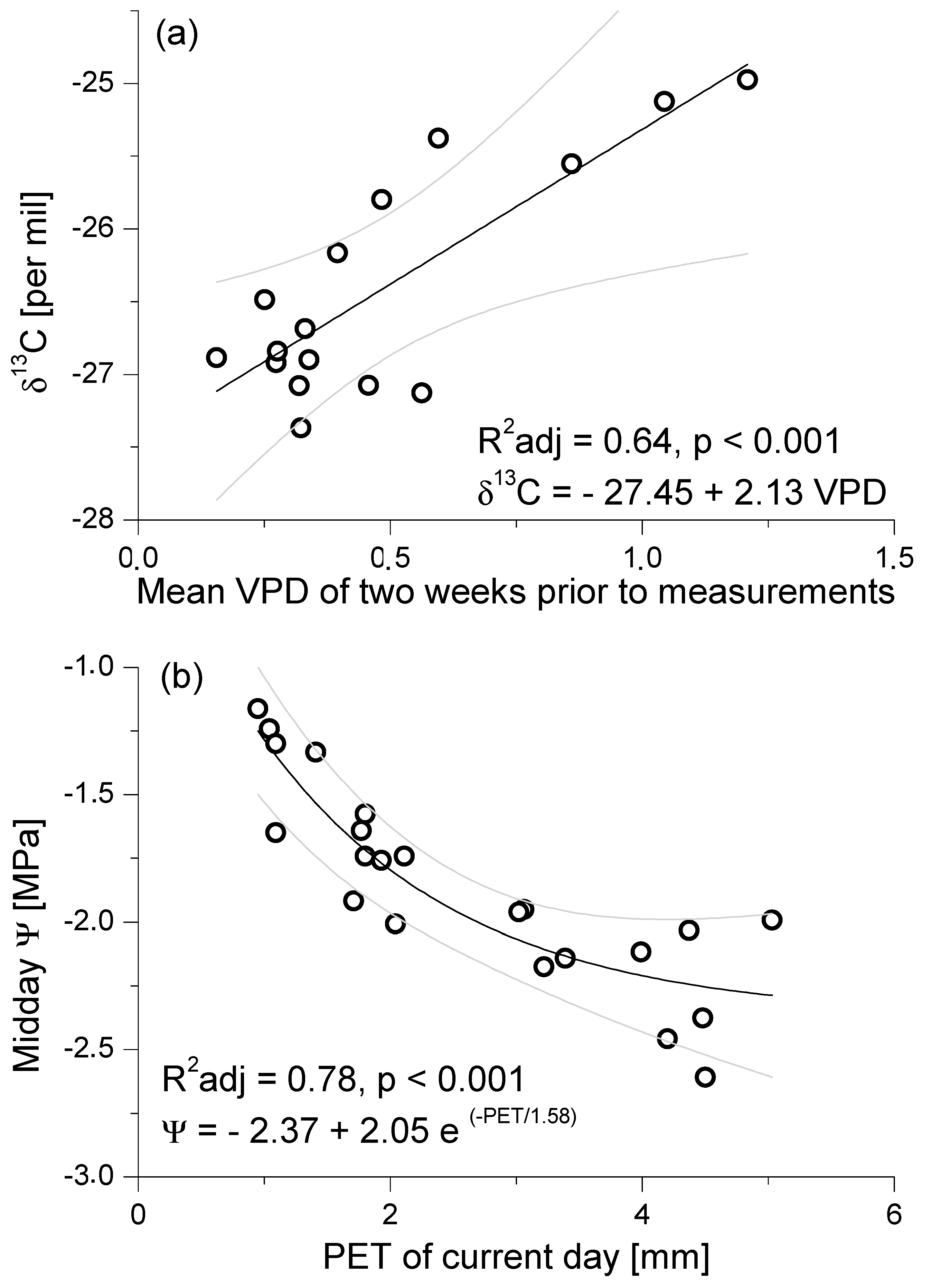
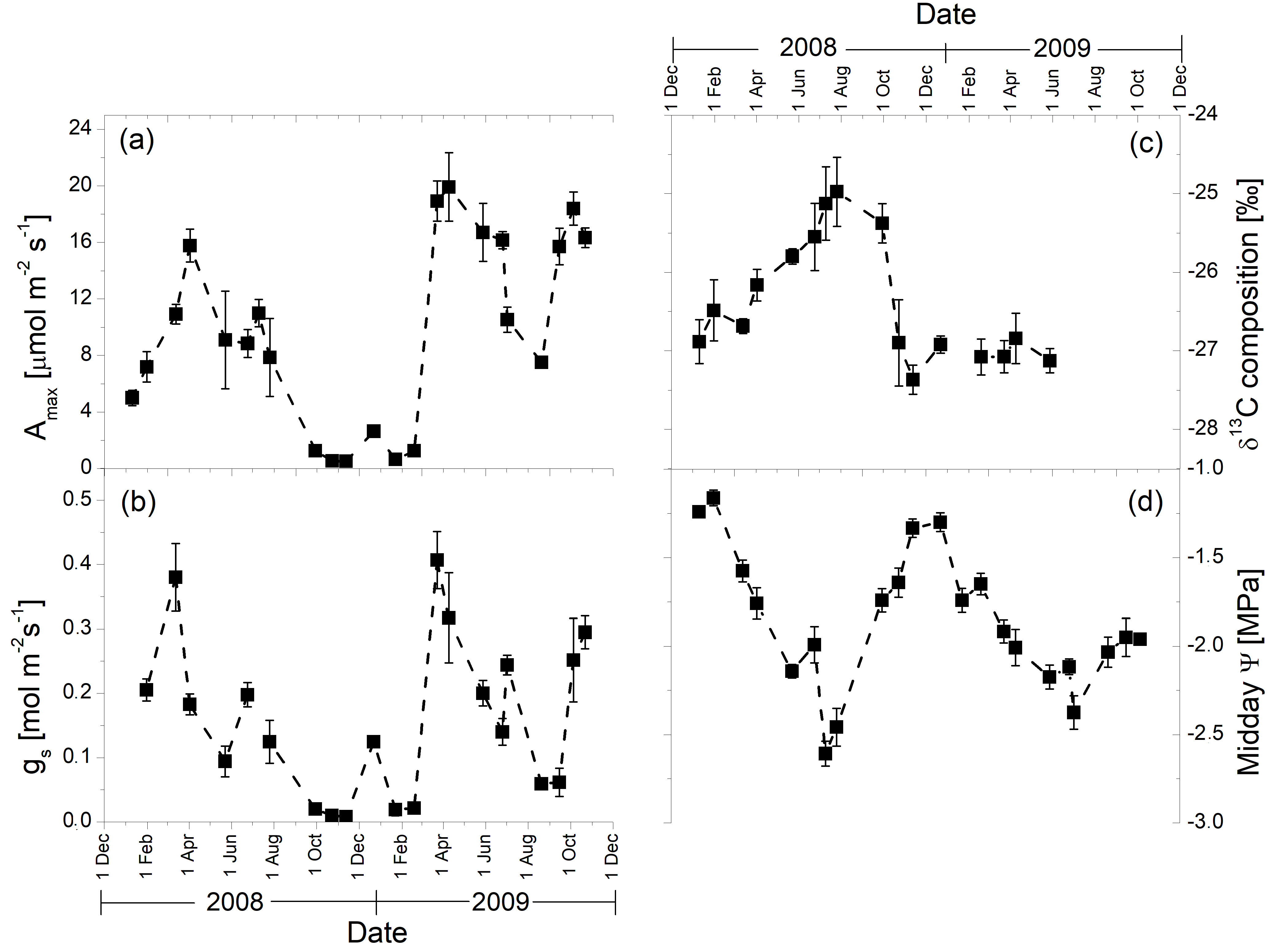
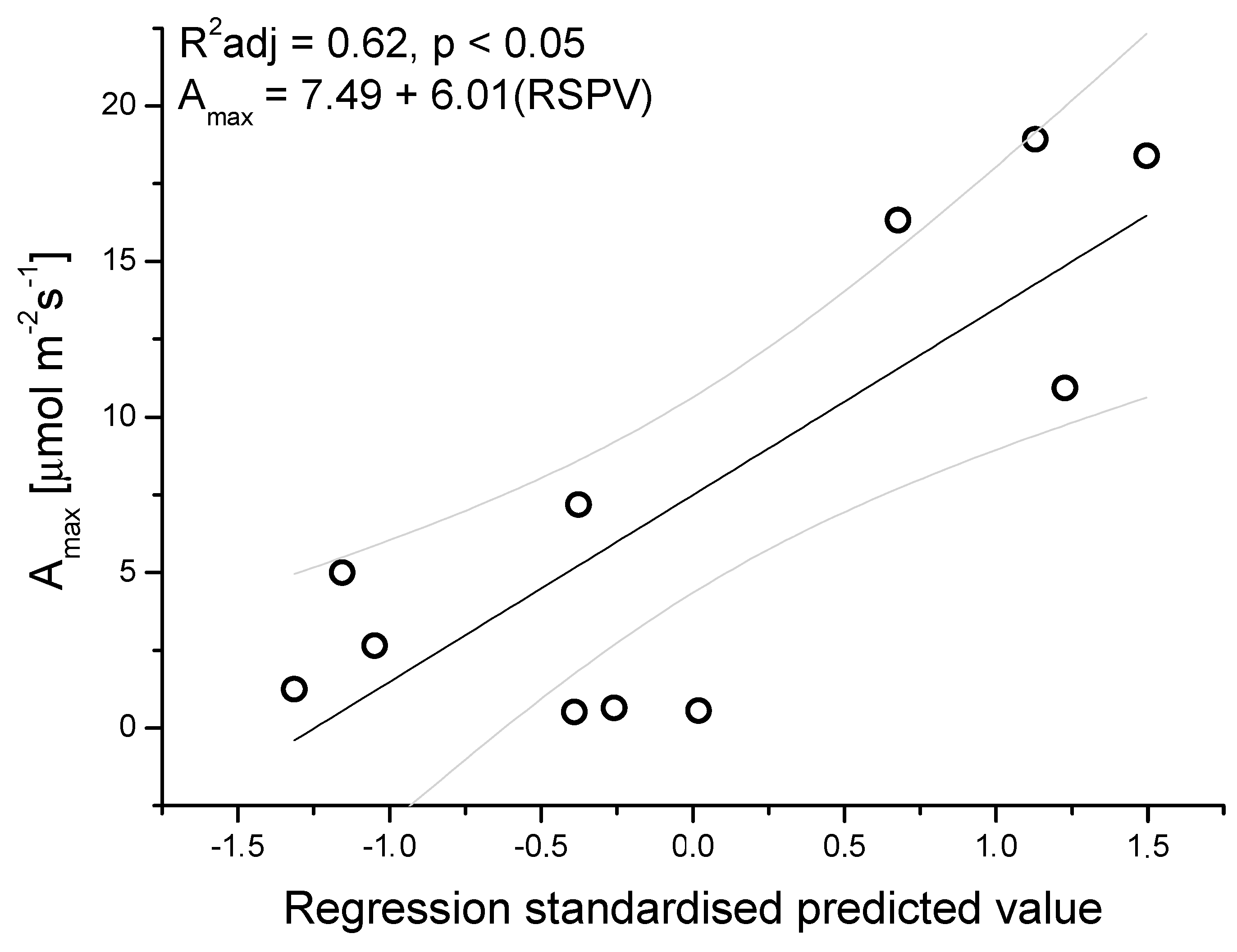
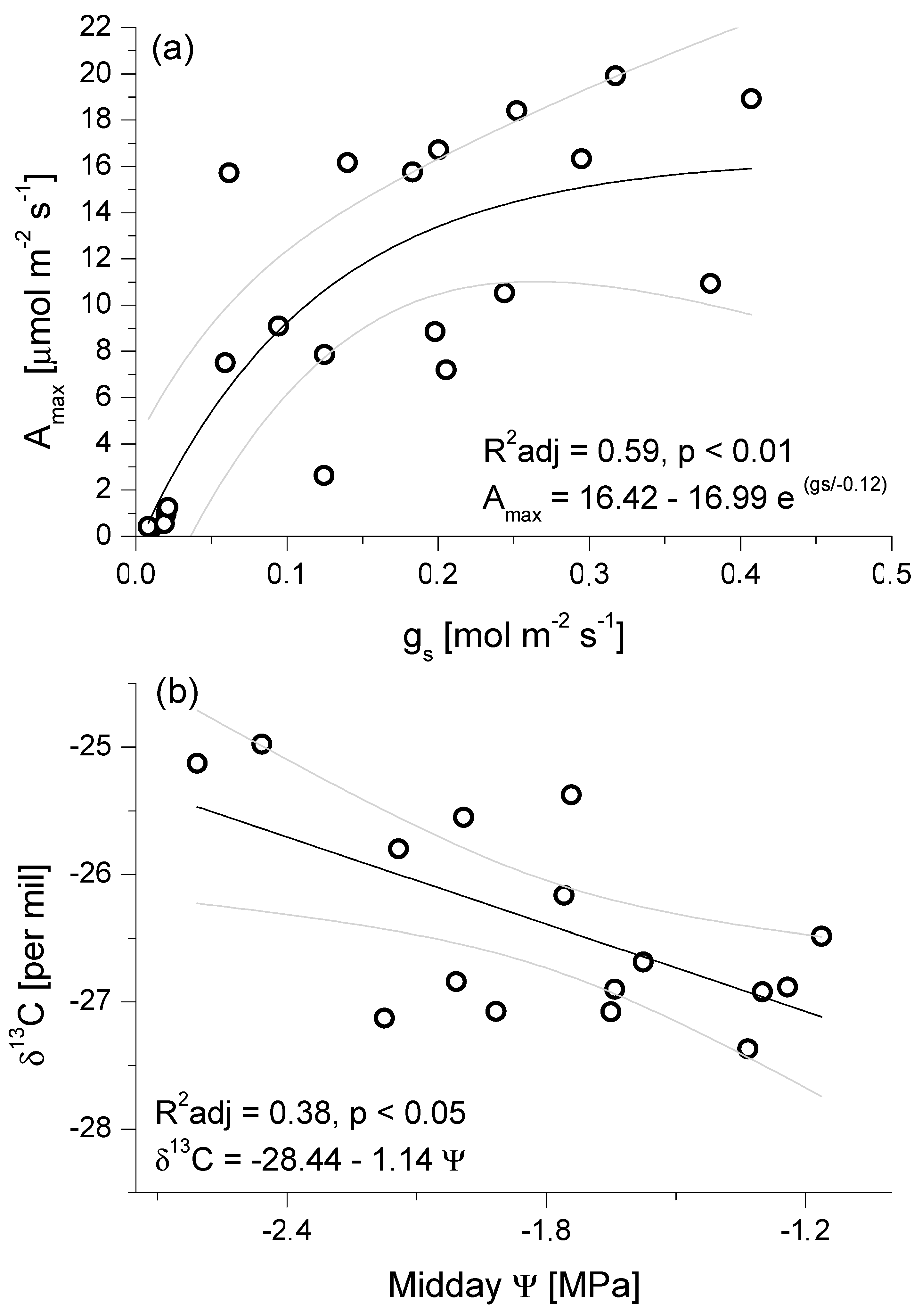
3.2. Seasonal Patterns of Gas Exchange, Needle Water Potential, and δ13C Composition
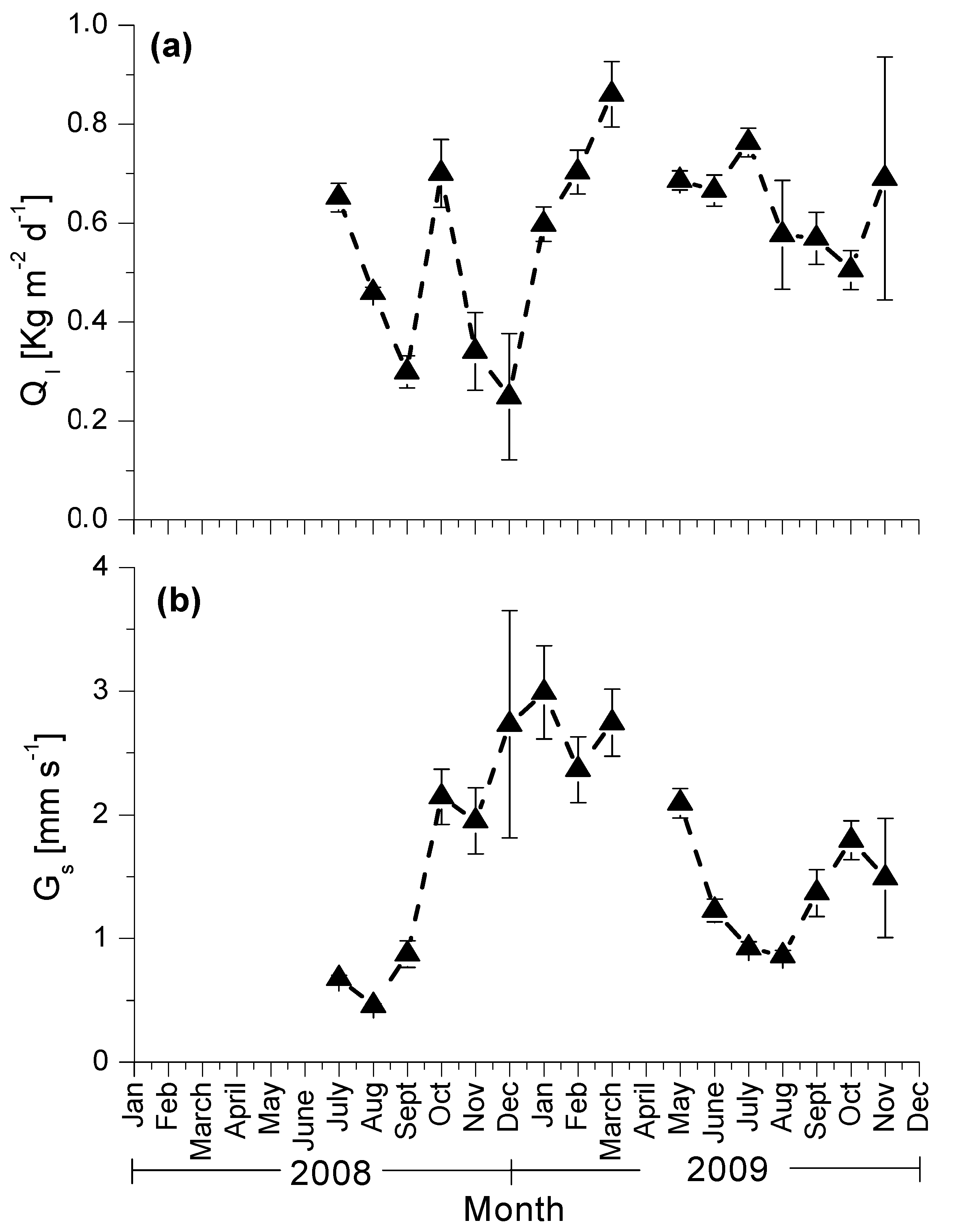
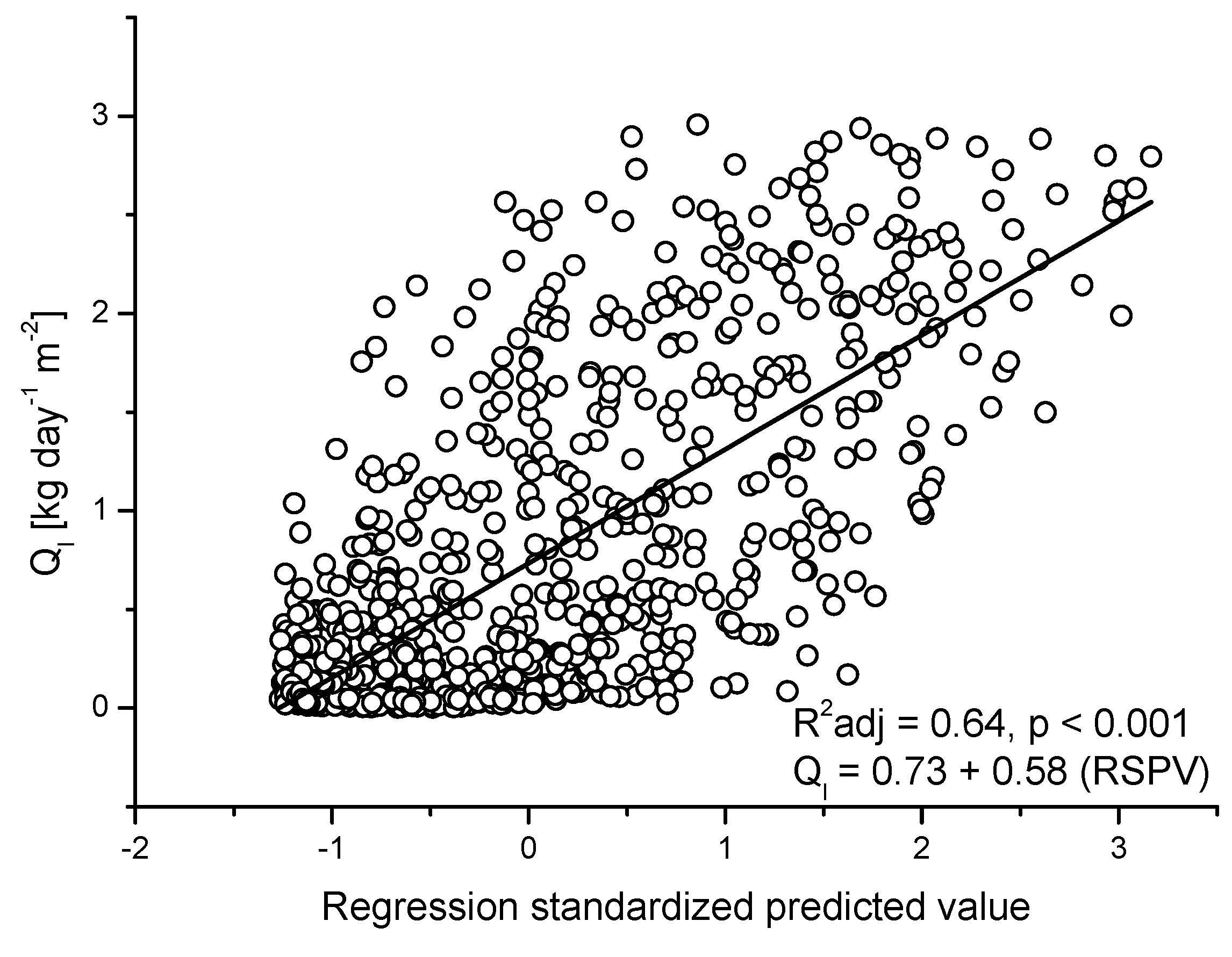
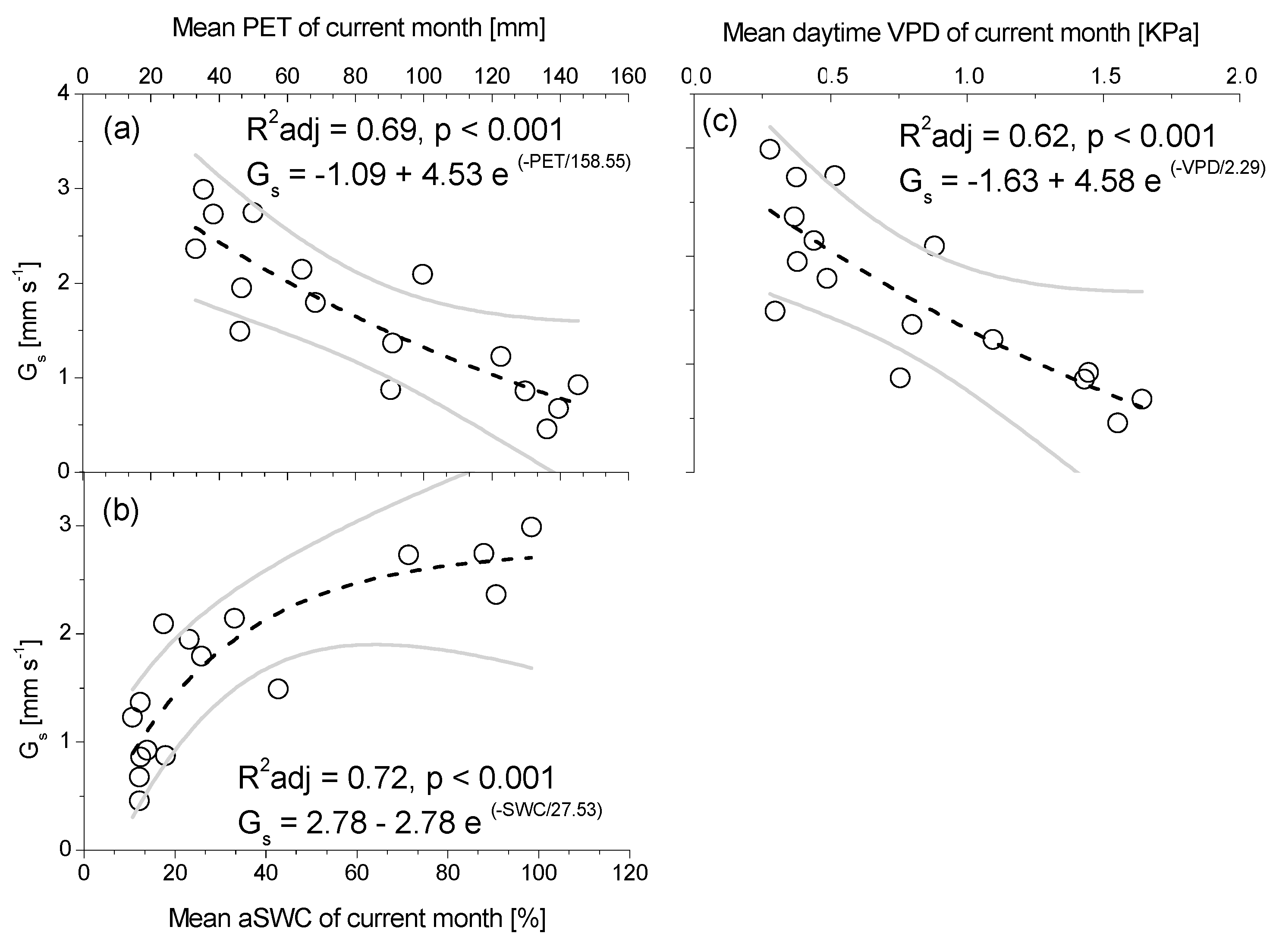
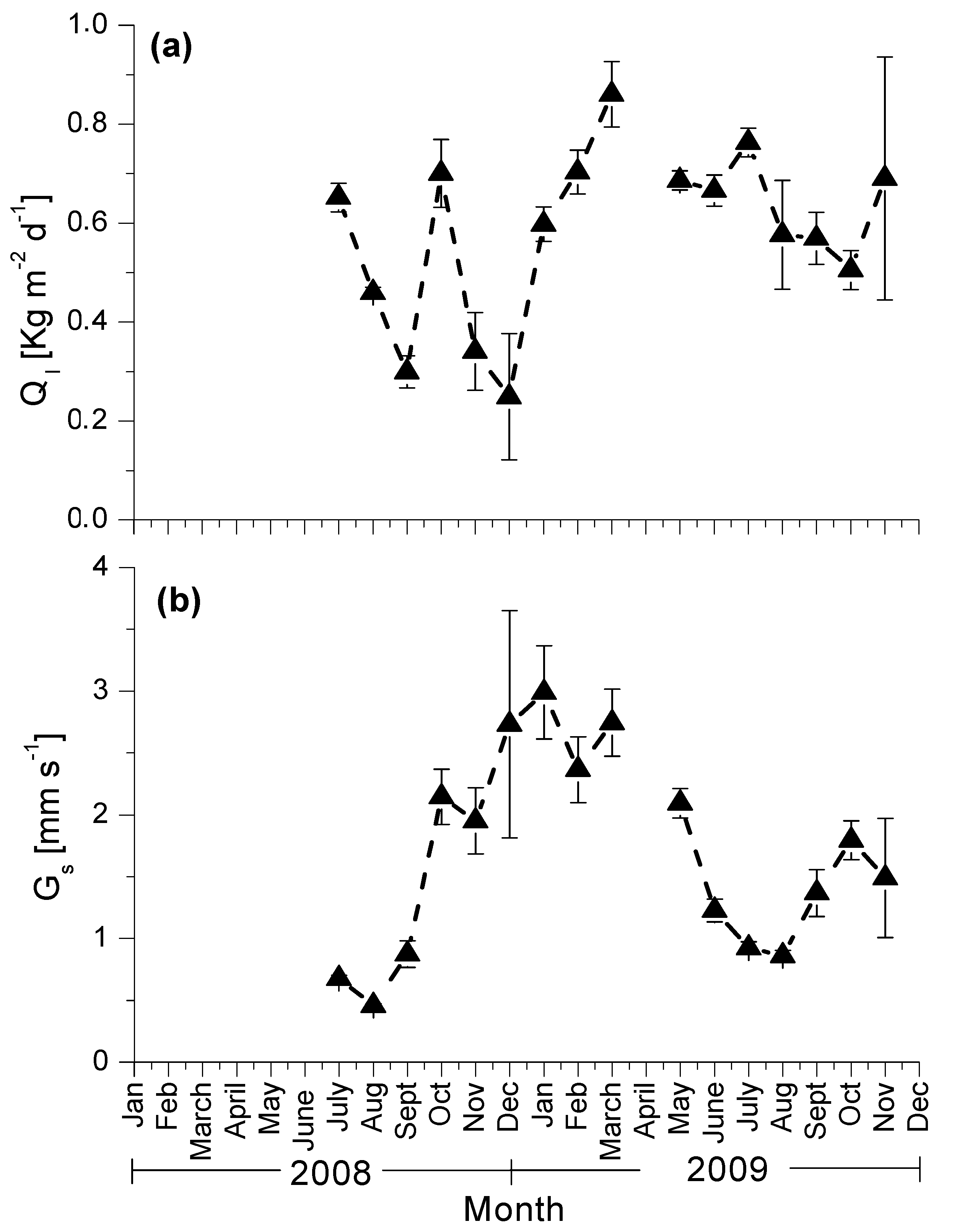
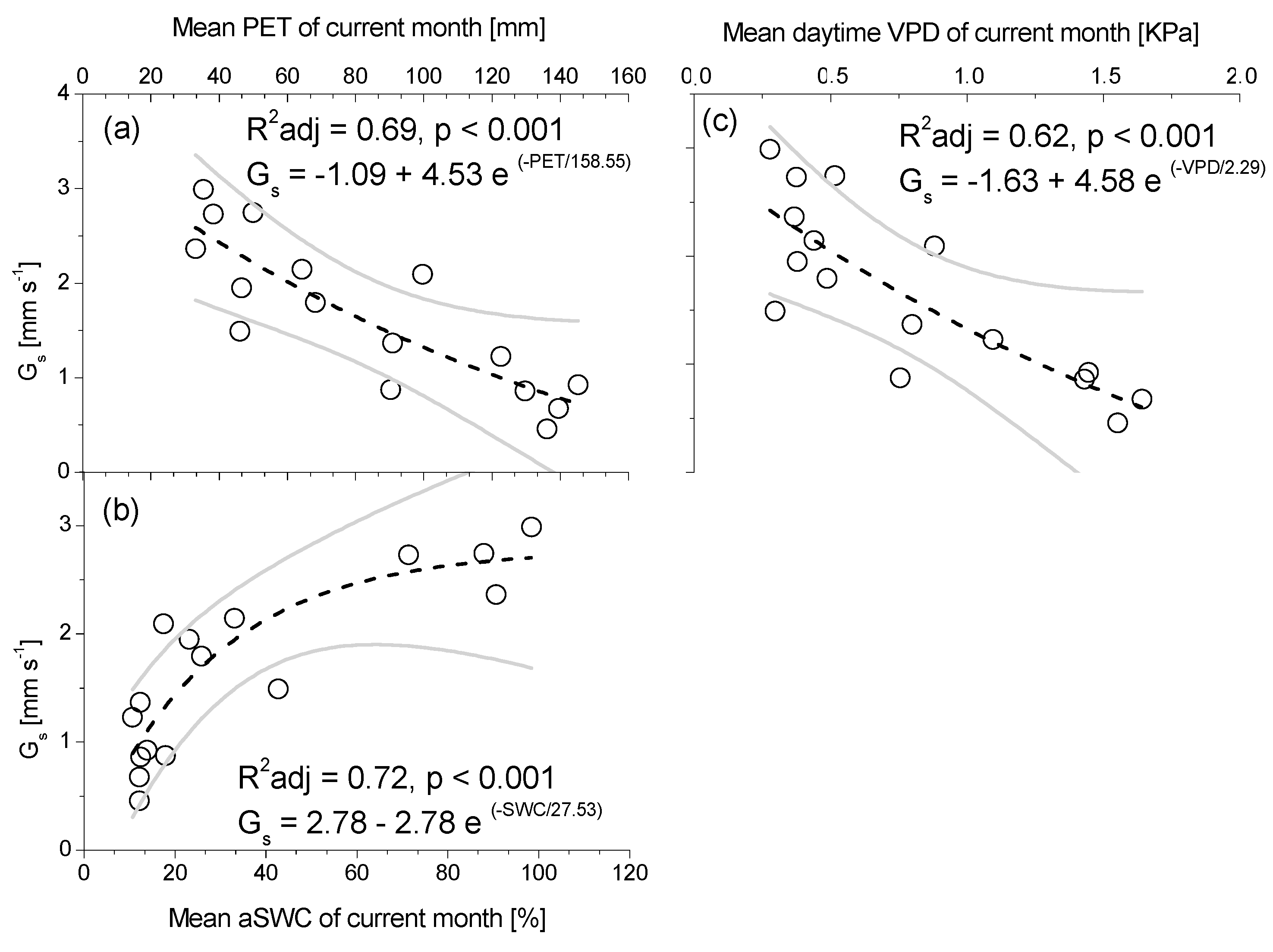
3.3. Seasonal Patterns of Sap Flow and Canopy Conductance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chambel, M.R.; Climent, J.; Pichot, C.; Ducci, F. Mediterranean Pines (Pinus halepensis Mill. and brutia Ten.). In Forest Tree Breeding in Europe: Current State-of-the-Art and Perspectives; Pâques, L.E., Ed.; Springer: Dordrecht, The Netherlands, 2013; Volume 25, pp. 229–265. ISBN 978-2-11128054-0. [Google Scholar]
- Euro + Med PlantBase—The Information Resource for Euro-Mediterranean Plant Diversity. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 12 February 2019).
- Dafis, S.; Moulopoulos, C. Einwirkung der Bodenfeuchtigkeit auf das Wachstum und Wurzelbildung von Samlingen der Aleppo und Hartkiefer (Pinus halepensis Mill und P. brutia Ten). Beih. Z. Schweiz. Forstver. 1969, 46, 225–260. [Google Scholar]
- Klein, T.; Cohen, S.; Yakir, D. Hydraulic adjustments underlying drought resistance of Pinus halepensis. Tree Physiol. 2011, 31, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Cohen, S.; Paudel, I.; Preisler, Y.; Rotenberg, E.; Yakir, D. Diurnal dynamics of water transport, storage and hydraulic conductivity in pine trees under seasonal drought. iForest 2016, 9, e1–e10. [Google Scholar] [CrossRef]
- Oliveras, I.; Martinez Vilalta, J.; Jimenez-Ortiz, T.; Lledo, J.; Escarre, A.; Piñol, J. Hydraulic properties of Pinus halepensis, Pinus pinea and Tetraclinis articulata in a dune ecosystem of Eastern Spain. Plant Ecol. 2003, 169, 131–141. [Google Scholar] [CrossRef]
- Pacheco, A.; Camarero, J.J.; Ribas, M.; Gazol, A.; Gutierrez, E.; Carrer, M. Disentangling the climate-driven bimodal growth pattern in coastal and continental Mediterranean pine stands. Sci. Total Environ. 2017, 615, 1518–1526. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; de Luis, M.; Novak, K.; del Castillo, M.E.; Schmitt, U.; Koch, G.; Štrus, J.; Mrak, P.; Žnidarič, M.T.; et al. Annual cambial rhythm in Pinus halepensis and Pinus sylvestris as indicator for climate adaptation. Front. Plant Sci. 2016, 7, 1923. [Google Scholar] [CrossRef]
- Gazol, A.; Ribas, M.; Gutiérrez, E.; Camarero, J.J. Aleppo pine forests from across Spain show drought-induced growth decline and partial recovery. Agric. For. Meteorol. 2017, 232, 186–194. [Google Scholar] [CrossRef]
- Schiller, G.; Cohen, Y. Water regime of a pine forest under a Mediterranean climate. Agric. For. Meteorol. 1995, 74, 181–193. [Google Scholar] [CrossRef]
- Klein, T.; Shpringer, I.; Fikler, B.; Elbaz, G.; Cohen, S.; Yakir, D. Relationships between stomatal regulation, water-use, and water-use efficiency of two coexisting key Mediterranean tree species. For. Ecol. Manag. 2013, 302, 34–42. [Google Scholar] [CrossRef]
- Birami, B.; Gattmann, M.; Heyer, A.G.; Grote, R.; Arneth, A.; Ruehr, N.K. Heat Waves Alter Carbon Allocation and Increase Mortality of Aleppo Pine Under Dry Conditions. Front. For. Glob. Chang. 2018, 1, 8. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Wolf, A.; Arango-Velez, A.; Choat, B.; Chmura, D.J.; Jansen, S.; Kolb, T.; Li, S.; Meinzer, F.C.; Pita, P.; et al. Woody plants optimise stomatal behaviour relative to hydraulic risk. Ecol. Lett. 2018, 21, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Del Castillio, J.; Voltas, J.; Ferrio, J.P. Carbon isotope discrimination, radial growth, and NDVI share spatiotemporal responses to precipitation in Aleppo pine. Trees Struct. Funct. 2015, 29, 223–233. [Google Scholar] [CrossRef]
- García-Ruiz, J.M.; López-Moreno, J.I.; Vicente-Serrano, S.M.; Lasanta-Martínez, T.; Beguería, S. Mediterranean water resources in a global change scenario. Earth-Sci. Rev. 2011, 105, 121–139. [Google Scholar] [CrossRef]
- Sarris, D.; Christodoulakis, D.; Körner, C. Recent decline in precipitation and tree growth in the eastern Mediterranean. Glob. Chang. Biol. 2007, 13, 1187–1200. [Google Scholar] [CrossRef]
- Grünwald, C.; Schiller, G. Needle xylem water potential and water saturation deficit in provenances of Pinus halepensis Mill. and P. brutia Ten. Forêt Méditerranéenne 1988, 10, 407–414. [Google Scholar]
- Michelozzi, M.; Loreto, F.; Colom, R.; Rossi, F.; Calamassi, R. Drought responses in Aleppo pine seedlings from two wild provenances with different climatic features. Photosynthetica 2011, 49, 564–572. [Google Scholar] [CrossRef]
- Klein, T.; Di Matteo, G.; Rotenberg, E.; Cohen, S.; Yakir, D. Differential ecophysiological response of a major Mediterranean pine species across a climatic gradient. Tree Physiol. 2012, 33, 26–36. [Google Scholar] [CrossRef]
- David-Schwartz, R.; Paudel, I.; Mizrachi, M.; Delzon, S.; Cochard, H.; Lukyanov, V.; Badel, E.; Capdeville, G.; Shklar, G.; Cohen, S. Indirect Evidence for Genetic Differentiation in Vulnerability to Embolism in Pinus halepensis. Front. Plant Sci. 2016, 7, 768. [Google Scholar] [CrossRef]
- European Soil Bureau Network—European Commission. Soil Atlas of Europe, 11th ed.; Office for Official Publications of the European Communities: Luxembourg, 2005; p. 128. ISBN 92-894-8120-X. [Google Scholar]
- Orfanoudakis, M.; Democritus University of Thrace, Nea Orestiada, Greece. Personal communication, 2019.
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Dingman, S.L. Physical Hydrology, 2nd ed.; Prentice Hall: Upper Saddler River, NJ, USA, 2002. [Google Scholar]
- Paparrizos, S. The Effect of Climate on the Hydrological Regime of Selected Greek Areas with Different Climate Conditions. Ph.D. Thesis, Faculty of Environment and Natural Resources, Albert-Ludwigs-University of Freiburg, Freiburg, Germany, 2016. [Google Scholar]
- Matzarakis, A.; Rutz, F.; Mayer, H. Modelling radiation fluxes in simple and complex environments—Application of the RayMan model. Int. J. Biometeorol. 2007, 51, 323–334. [Google Scholar] [CrossRef]
- Matzarakis, A.; Rutz, F.; Mayer, H. Modelling radiation fluxes in simple and complex environments: Basics of the RayMan model. Int. J. Biometeorol. 2010, 54, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Matzarakis, A.; Mayer, H.; Schindler, D.; Fritsch, J. Simulation des Wasserhaushaltes eines Buchenwaldes mit dem forstlichen Wasserhaushaltsmodell WBS3. Bericht Des Meteorologischen Instituts der Universität Freiburg 2000, 5, 137–146. [Google Scholar]
- Fotelli, M.N.; Nahm, M.; Radoglou, K.; Rennenberg, H.; Matzarakis, A. Seasonal and interannual ecophysiological responses of beech (Fagus sylvatica) at its south-eastern distribution limit in Europe. For. Ecol. Manag. 2009, 257, 1157–1164. [Google Scholar] [CrossRef]
- United Nations Environment Programme (UNEP). World Atlas of Desertification; Edward Arnold: London, UK, 1992; p. 69. ISBN 0 340 55512 2. [Google Scholar]
- Paparrizos, S.; Maris, F.; Matzarakis, A. Integrated analysis and mapping of aridity over Greek areas with different climate conditions. Glob. NEST J. 2016, 18, 131–145. [Google Scholar]
- Food and Agriculture Organization (FAO). Forest Resources Assessment 1990: Tropical Countries; FAO-UN: Rome, Italy, 1993; ISBN 92-5-103390-0. [Google Scholar]
- Granier, A. A new method of sap flow measurement in tree stems. Ann. Sci. For. 1985, 42, 193–200. [Google Scholar] [CrossRef]
- Granier, A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiol. 1987, 3, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Do, F.; Rocheteau, A. Influence of natural temperature gradients on measurements of xylem sap flow with thermal dissipation probes. 1. Field observations and possible remedies. Tree Physiol. 2002, 22, 641–648. [Google Scholar] [CrossRef]
- Mitsopoulos, I.D.; Dimitrakopoulos, A.P. Allometric equations for crown fuel biomass of Aleppo pine (Pinus halepensis Mill.) in Greece. Int. J. Wildland Fire 2007, 16, 642–647. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Mangirón, M.; Ogaya, R.; Sauret, M.; Serrano, L.; Peñuelas, J.; Piñol, J. Sap flow of three co-occurring Mediterranean woody species under varying atmospheric and soil water conditions. Tree Physiol. 2003, 23, 747–758. [Google Scholar] [CrossRef]
- Phillips, N.; Oren, R. A comparison of daily representations of canopy conductance based on two conditional time-averaging methods and dependence of daily conductance on environmental factors. Ann. For. Sci. 1998, 55, 217–235. [Google Scholar] [CrossRef]
- Salazar-Tortosa, D.; Castro, J.; Rubio de Casas, R.; Viñegla, B.; Sánchez-Cañete, E.P.; Villar-Salvador, P. Gas exchange at whole plant level shows that a less conservative water use is linked to a higher performance in three ecologically distinct pine species. Environ. Res. Lett. 2018, 13, 045004. [Google Scholar] [CrossRef]
- Chang, C.Y.; Unda, F.; Zubilewich, A.; Mansfield, S.D.; Ensminger, I. Sensitivity of cold acclimation to elevated autumn temperature in field-grown Pinus strobus seedlings. Front. Plant Sci. 2015, 6, 165. [Google Scholar] [CrossRef]
- Awada, T.; Radoglou, K.; Fotelli, M.N.; Constantinidou, H.-I.A. Ecophysiology of three Mediterranean pine species under contrasting light regimes. Tree Physiol. 2002, 23, 33–41. [Google Scholar] [CrossRef]
- Pflug, E.; Brüggemann, W. Frost-acclimation of photosynthesis in overwintering Mediterranean holm oak, grown in Central Europe. Int. J. Plant Biol. 2012, 3, e1. [Google Scholar] [CrossRef]
- Fotelli, M.N.; Geβler, A.; Peuke, A.D.; Rennenberg, H. Drought affects the competition between Fagus sylvatica L. seedlings and an early successional species (Rubus fruticosus): Growth, water status and δ13C composition. New Phytol. 2001, 151, 427–435. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Keitel, C.; Matzarakis, A.; Rennenberg, H.; Geßler, A. Carbon isotopic composition and oxygen isotopic enrichment in phloem and total leaf organic matter of European beech (Fagus sylvatica L.) along a climate gradient. Plant Cell Environ. 2006, 29, 1492–1507. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Costa, E.; Poyatos, R.; Sabaté, S. Contrasting growth and water use strategies in four co-occurring Mediterranean tree species revealed by concurrent measurements of sap flow and stem diameter variations. Agric. For. Meteorol. 2015, 207, 24–37. [Google Scholar] [CrossRef]
- Oren, R.; Phillips, N.; Ewers, B.E.; Pataki, D.E.; Megonigal, J.P. Sap- flux scaled transpiration response to light, vapor pressure deficit, and leaf area reduction in a flooded Taxodium distichum forest. Tree Physiol. 1999, 19, 337–347. [Google Scholar] [CrossRef]
- Sperry, J.S. Hydraulic constraints on plant gas exchange. Agric. For. Meteorol. 2000, 104, 13–23. [Google Scholar] [CrossRef]
- Tognetti, R.A.; Michelozzi, M.; Giovanelli, A. Geographical variation in water relations, hydraulic architecture and terpene composition of Aleppo pine seedlings from Italian provenances. Tree Physiol. 1997, 17, 241–250. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | Rainfall (mm) | Aridity Index | aSWC (%) | VPD (KPa) |
|---|---|---|---|---|
| 2008 | 542.1 | 0.77 | 36.9 | 0.55 |
| 2009 | 680.2 | 1.16 | 46.6 | 0.46 |
| 2008–2017 | 544.9 | 0.84 | 37.5 | 0.48 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fotelli, M.N.; Korakaki, E.; Paparrizos, S.A.; Radoglou, K.; Awada, T.; Matzarakis, A. Environmental Controls on the Seasonal Variation in Gas Exchange and Water Balance in a Near-Coastal Mediterranean Pinus halepensis Forest. Forests 2019, 10, 313. https://doi.org/10.3390/f10040313
Fotelli MN, Korakaki E, Paparrizos SA, Radoglou K, Awada T, Matzarakis A. Environmental Controls on the Seasonal Variation in Gas Exchange and Water Balance in a Near-Coastal Mediterranean Pinus halepensis Forest. Forests. 2019; 10(4):313. https://doi.org/10.3390/f10040313
Chicago/Turabian StyleFotelli, Mariangela N., Evangelia Korakaki, Spyridon A. Paparrizos, Kalliopi Radoglou, Tala Awada, and Andreas Matzarakis. 2019. "Environmental Controls on the Seasonal Variation in Gas Exchange and Water Balance in a Near-Coastal Mediterranean Pinus halepensis Forest" Forests 10, no. 4: 313. https://doi.org/10.3390/f10040313
APA StyleFotelli, M. N., Korakaki, E., Paparrizos, S. A., Radoglou, K., Awada, T., & Matzarakis, A. (2019). Environmental Controls on the Seasonal Variation in Gas Exchange and Water Balance in a Near-Coastal Mediterranean Pinus halepensis Forest. Forests, 10(4), 313. https://doi.org/10.3390/f10040313