Occurrence, Seasonal Abundance, and Superparasitism of Ooencyrtus kuvanae (Hymenoptera: Encyrtidae) as an Egg Parasitoid of the Spotted Lanternfly (Lycorma delicatula) in North America

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Plots
2.2. Egg Collection
2.3. Egg Incubation
2.4. Occurrence of O. kuvanae
2.5. Seasonal Abundance of O. kuvanae
2.6. Parasitism of O. kuvanae
2.7. Superparasitism in O. kuvanae
2.8. Identification of O. kuvanae
2.9. Data Analysis
3. Results
3.1. Egg Collection
3.2. Occurrence of O. kuvanae
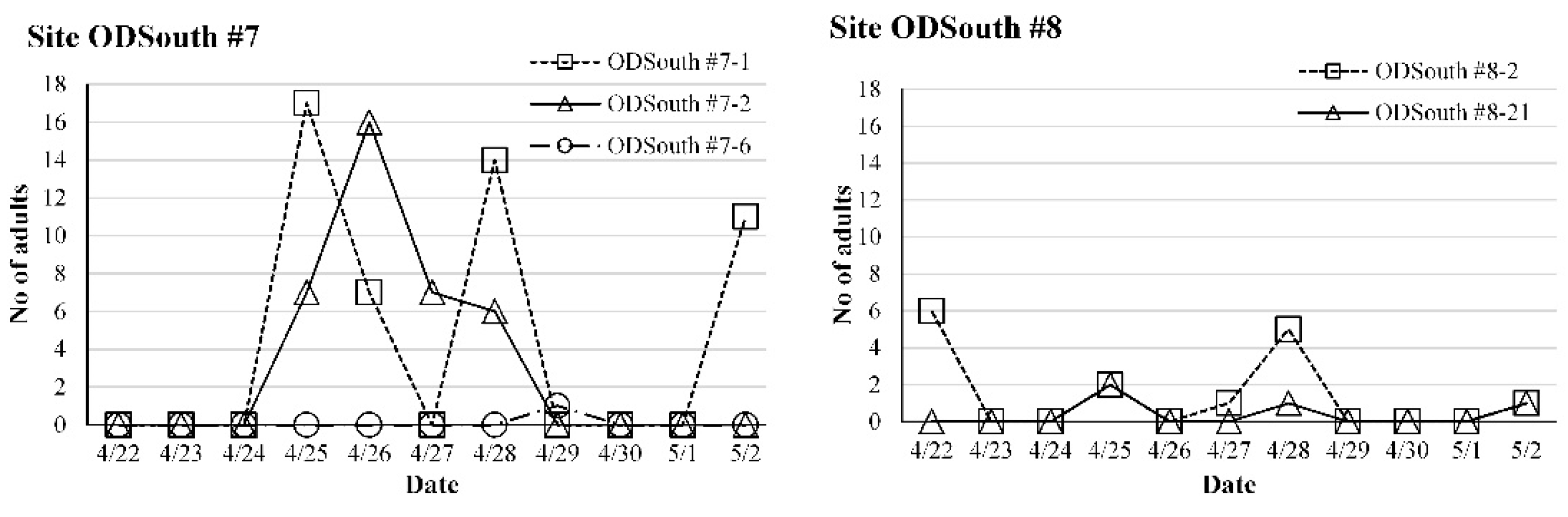
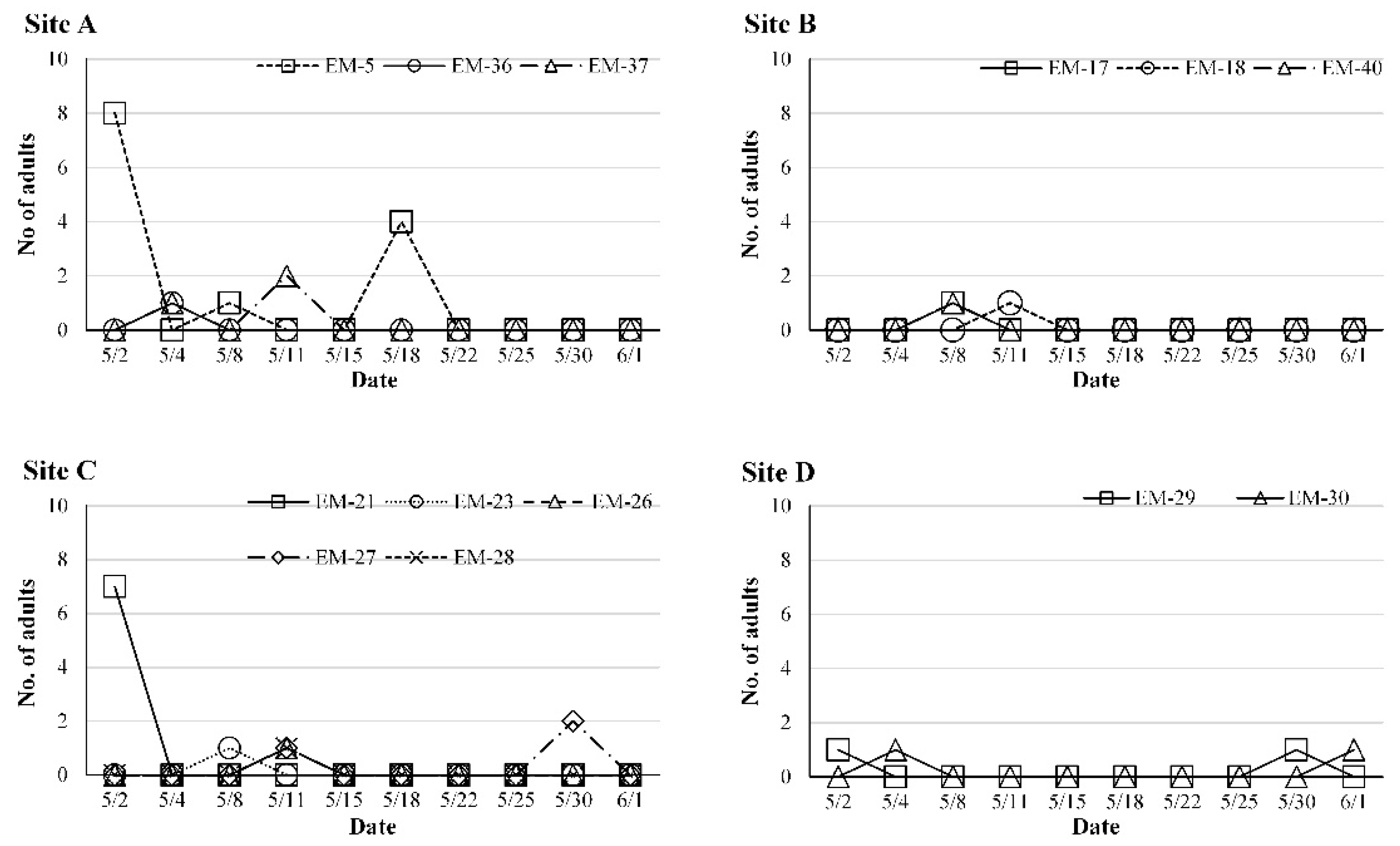
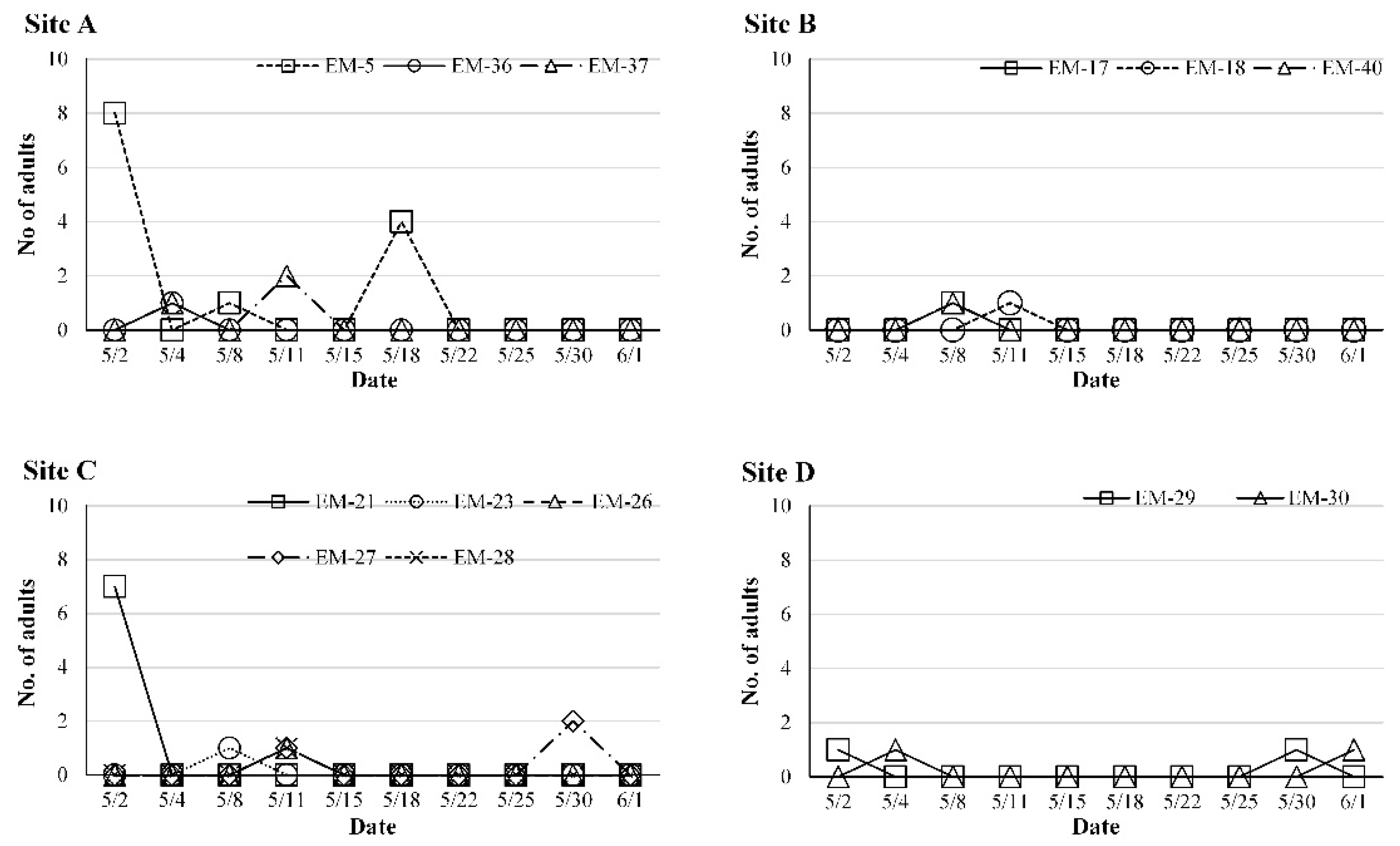
3.3. Seasonal Abundance of O. kuvanae
3.4. Parasitism of O. kuvanae
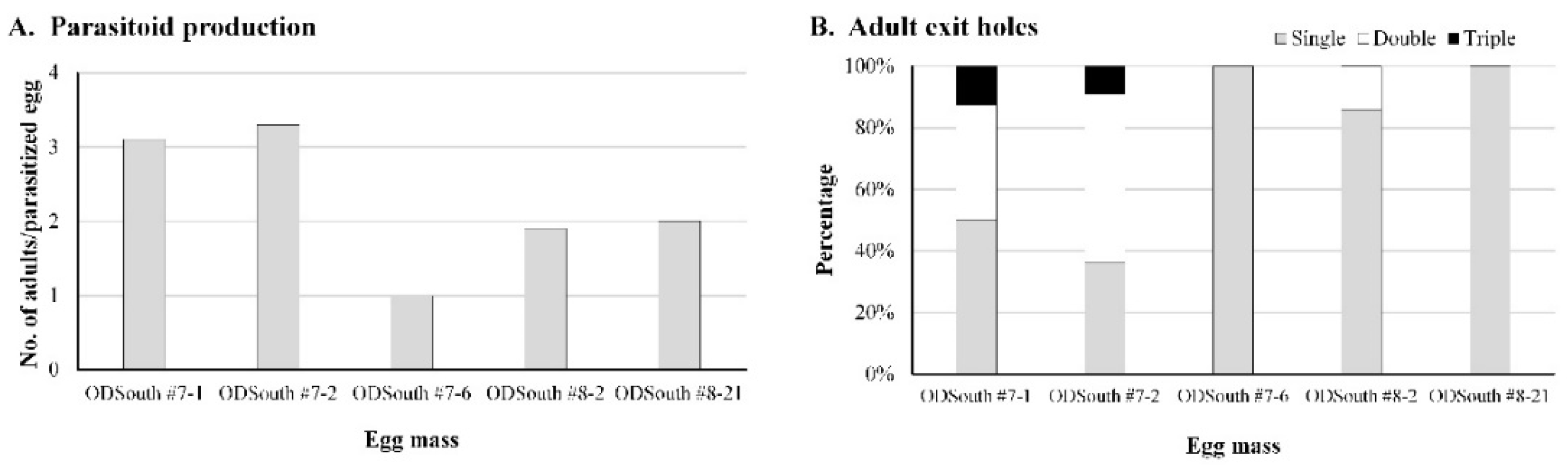
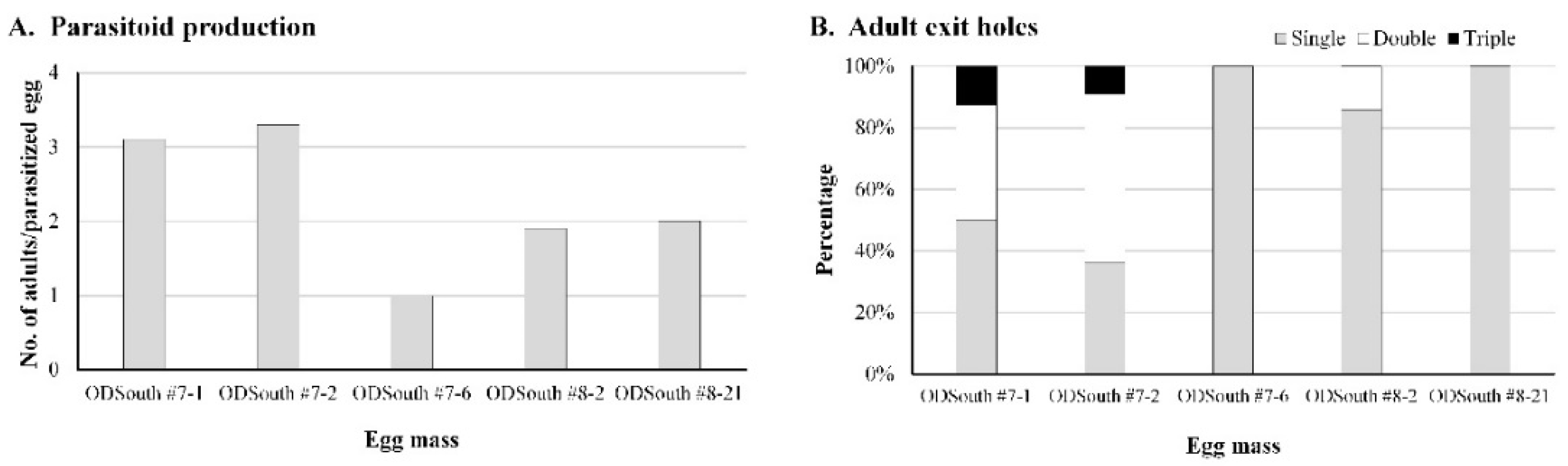
3.5. Superparasitism
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, A. Description of a new genus and some new species of homopterous insects from the East in the collection of British Museum. Ann. Mag. Nat. Hist. 1845, 15, 34–37. [Google Scholar] [CrossRef]
- Liu, G. Some extracts from the history of entomology in China. Psyche 1939, 46, 23–28. [Google Scholar] [CrossRef]
- Zhou, J.X. Lycorma delicatula (White) (Homoptera: Fulgoridae). In Forest Insects of China, 2nd ed.; Xiao, G.R., Ed.; Chinese Forestry Publishing House: Beijing, China, 1992; pp. 169–170. [Google Scholar]
- Han, J.M.; Kim, H.J.; Lim, E.J.; Lee, S.H.; Kwon, Y.J.; Cho, S.W. Lycorma delicatula (Hemiptera: Auchenorrhyncha: Fulgoridae: Aphaeninae) finally, but suddenly arrived in Korea. Entomol. Res. 2008, 38, 281–286. [Google Scholar] [CrossRef]
- Barringer, L.E.; Donovall, L.R.; Spichiger, S.-E.; Lynch, D.; Henry, D. The first New World record of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae). Entomol. News 2015, 125, 20–23. [Google Scholar] [CrossRef]
- Dara, S.K.; Barringer, L.; Arthurs, S.P. Lycorma delicatula (Hemiptera: Fulgoridae): A new invasive pest in the United States. J. Integr. Pest Manag. 2015, 6, 20. [Google Scholar] [CrossRef]
- Pennsylvania Department of Agriculture. Spotted Lanternfly. Available online: https://www.agriculture.pa.gov/Plants_Land_Water/PlantIndustry/Entomology/spotted_lanternfly/Pages/default.aspx (accessed on 21 November 2018).
- Virginia Cooperative Extension. Spotted Lanternfly in Virginia. Available online: https://ext.vt.edu/agriculture/commercial-horticulture/spotted-lanternfly.html (accessed on 21 November 2018).
- New Jersey Department of Agriculture. Spotted Lanternfly. Available online: https://www.nj.gov/agriculture/divisions/pi/prog/spottedlanternfly.html (accessed on 21 November 2018).
- New York Department of Environmental Conservation. Spotted Lanternfly. Available online: http://www.dec.ny.gov/animals/113303.html (accessed on 21 November 2018).
- Delaware Department of Agriculture. Spotted Lanternfly. Available online: https://agriculture.delaware.gov/plant-industries/spotted-lanternfly/ (accessed on 21 November 2018).
- Kim, J.; Lee, E.H.; Seo, Y.M.; Kim, N.Y. Cyclic behavior of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae) on host plants. J. Insect Behav. 2011, 24, 423–435. [Google Scholar] [CrossRef]
- Lee, J.E.; Moon, S.R.; Ahn, H.G.; Cho, S.R.; Yang, J.O.; Yoon, C.; Kim, J.H. Feeding behavior of Lycorma delicatula (Hemiptera: Fulgoridae) and response on feeding stimulants of some plants. Korean J. Appl. Entomol. 2009, 48, 467–477. [Google Scholar] [CrossRef]
- Park, J.D.; Kim, M.Y.; Lee, S.G.; Shin, S.C.; Kim, J.H.; Park, I.K. Biological characteristics of Lycorma delicatula and the control effects of some insecticides. Korean J. Appl. Entomol. 2009, 48, 53–57. [Google Scholar] [CrossRef]
- Pennsylvania Department of Agriculture. PA Agriculture Department Adds Seven Counties to Spotted Lanternfly Quarantine Zone. Available online: http://www.media.pa.gov/pages/Agriculture_details.aspx?newsid=616 (accessed on 28 September 2018).
- Lee, K.Y.; Kim, S.K.; Kim, I.H.; Kim, K.S. Seasonal occurrence of spot clothing wax cicada, Lycorma delicatula (Hemiptera: Fulgoridae) and its control efficacy using EFAM at the vineyards. Korean J. Pestic. Sci. 2011, 15, 303–309. [Google Scholar]
- Howard, L.O. Technical results from the gypsy moth parasite laboratory. I. The parasites reared or supposed to have been reared from the eggs of gypsy moth. U.S. Dep. Agric. Tech. Ser. Bull. 1910, 19, 1–12. [Google Scholar]
- Hirose, Y. The activity of the egg parasites of the pinemoth, Dendrolimus spectabilis Butler, in Japanese black pine forest on the seacoast. Sci. Bull. Fac. Agric. Kyushi Univ. 1964, 21, 13–24. [Google Scholar]
- Koidzumi, K.; Shobata, K. Studies on Eriogyna pyretorum Westwood and its fishing thread. XI. Epiparasites. J. Soc. Trop. Agric. Formosa 1940, 12, 259–265. [Google Scholar]
- Howard, L.O.; Fiske, W.F. The Importation into the United States of the Parasites of the Gypsy Moth and the Brown-Tailed Moth: A Report of Progress with Some Consideration of Previous and Concurrent Efforts of this Kind; USDA: Washington, DC, USA, 1911.
- Muesebeck, C.F.W.; Dohanian, S.M. A Study in Hyperparasitism with Particular Reference to the Parasites of Apanteles melanoscelus (Ratzeburg); USDA: Washington, DC, USA, 1927.
- Crossman, S.S. Some methods of colonizing imported parasites and determining their increase and spread. J. Econ. Entomol. 1917, 10, 177–183. [Google Scholar] [CrossRef]
- Britton, W.E. The gypsy moth. Conn. Agric. Exp. Stn. Bull. 1935, 375, 625–647. [Google Scholar]
- Crossman, S.S. Two imported egg parasites of the gypsy moth, Anastatus bifasciatus Fonsc. and Schedius kuvanae Howard. J. Agric. Res. 1925, 30, 643–675. [Google Scholar]
- Brown, M.W. Literature review of Ooencyrtus kuvanae (Hym.: Encyrtidae), an egg parasite of Lymantria dispar (Lep.: Lymantriidae). Entomophaga 1984, 29, 249–265. [Google Scholar] [CrossRef]
- Smilowitz, Z.; Rhoads, L. An assessment of gypsy moth natural enemies in Pennsylvania. Environ. Entomol. 1973, 2, 797–799. [Google Scholar] [CrossRef]
- Liu, H.P.; Mottern, J. An old remedy for a new problem? Identification of Ooencyrtus kuvanae (Hymenoptera: Encyrtidae), an egg parasitoid of Lycorma delicatula (Hemiptera: Fulgoridae) in North America. J. Insect Sci. 2017, 17, 1–6. [Google Scholar] [CrossRef]
- van Alphen, J.J.M.; Visser, M.E. Superparasitism as an adaptive strategy for insect parasitoids. Annu. Rev. Entomol. 1990, 35, 59–79. [Google Scholar] [CrossRef]
- Gandon, S.; Rivero, A.; Varaldi, J. Superparasitism evolution: Adaptation and manipulation? Am. Nat. 2006, 176, 1–22. [Google Scholar] [CrossRef]
- Godfray, H.C.J. Parasitoids: Behavioral and Evolutionary Ecology; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Gu, H.; Wang, Q.; Dorn, S. Superparasitism in Cotesia glomerata: Response of hosts and consequences for parasitoids. Ecol. Entomol. 2003, 28, 422–431. [Google Scholar] [CrossRef]
- Dorn, S.; Beckage, N. Superparasitism in gregarious hymenopteran parasitoids: Ecological, behavioural and physiological perspectives. Physiol. Entomol. 2007, 32, 199–211. [Google Scholar] [CrossRef]
- Tunca, H.; Colombel, E.-A.; Venard, M.; Tabone, E. Incidence of superparasitism in the egg parasitoid, Ooencyrtus kuvanae Howard (Hymenoptera: Encyrtidae). Biocontrol Sci. Technol. 2017, 27, 796–808. [Google Scholar] [CrossRef]
- Hofstetter, R.W.; Raffa, K.F. New host record for Ooencyrtus kuvanae (Hymenoptera: Encyrtidae). Entomol. News 1997, 108, 63–65. [Google Scholar]
- Huang, D.W.; Noyes, J.S. A revision of the Indo-Pacific species of Ooencyrtus (Hymenoptera: Encyrtidae), parasitoids of the immature stages of economically important insect species (mainly hemipteran and Lepidoptera). Bull. Nat. Hist. Mus. Lond. 1994, 63, 1–136. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. Available online: http://www.R-project.org/ (accessed on 27 November 2018).
- Hu, S. Ailanthus. Arnoldia 1979, 39, 29–50. [Google Scholar]
- Swingle, W.T. The early European history and the botanical name of the tree-of-heaven, Ailanthus altissima. J. Wash. Acad. Sci. 1916, 6, 409–498. [Google Scholar]
- Zasada, J.C.; Little, S. Ailanthus altissima (P. Mill.) Swingle: Ailanthus. In Woody Plant Seed Manual; Agric. Handbook No. 727; Bonner, F.T., Karrfalt, R.P., Eds.; USDA Forest Service: Washington, DC, USA, 2008; pp. 224–226. [Google Scholar]
- Howard, J.L. Ailanthus altissima. In Fire Effects Information System; USDA Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, 2004. Available online: https://www.fs.fed.us/database/feis/plants/tree/ailalt/all.html (accessed on 29 October 2018).
- Cooperband, M.F.; USDA APHIS, Buzzards Bay, MA, USA. Personal communication, 2018.
- Liu, H.-P.; Pennsylvania Department of Conservation and Natural Resources, Harrisburg, PA, USA. Personal observation, 2018.
- Hoover, K.; Pennsylvania State University, State College, PA, USA. Personal communication, 2018.
- Leach, H.; Krawczyk, G. Spotted Lanternfly Management for Homeowners. Penn State Extension: State College, PA, USA. Available online: https://extension.psu.edu/spotted-lanternfly-management-for-homeowners (accessed on 31 October 2018).
- Cooperband, M.F.; Mack, R.; Spichiger, S.-E. Chipping to destroy egg masses of the spotted lanternfly, Lycorma delicatula (Hempitera: Fulgoridae). J. Insect Sci. 2018, 18, 1–3. [Google Scholar] [CrossRef]
- Yang, Z.-Q.; Choi, W.-Y.; Cao, L.-M.; Wang, X.-Y.; Hou, Z.-R. A new species of Anastatus (Hymenoptera: Eulpelmidae) from China, parasitizing eggs of Lycorma delicatula (Homoptera: Fulgoridae). Zool. Syst. 2015, 40, 290–302. [Google Scholar]
- Choi, M.-Y.; Yang, Z.-Q.; Wang, X.-Y.; Hou, Z.-R.; Kim, J.H.; Byeon, Y.W. Parasitism rate of egg parasitoid Anastatus orientalis (Hymenoptera: Eupelmidae) on Lycorma delicatula (Hemiptera: Fulgoridae) in China. Korea J. Appl. Entomol. 2014, 53, 135–139. [Google Scholar] [CrossRef]
- Yan, J.H.; Yu, X.M.; Qin, X.B.; Wang, F.Y.; Bo, L.L. Study on the biology of Dryinus browni. Shandong For. Sci. Technol. 2008, 1, 16–18. [Google Scholar]
- Kim, I.-K.; Koh, S.-H.; Lee, J.-S.; Choi, W.I.; Shin, S.-C. Discovery of an egg parasitoid of Lycorma delicatula (Hemiptera: Fulgoridae) an invasive species in South Korea. J. Asia Pac. Entomol. 2015, 14, 213–215. [Google Scholar] [CrossRef]
- Hokkanen, H.; Pimentel, D. New approach for selecting biological control agents. Can. Entomol. 1984, 116, 1109–1121. [Google Scholar] [CrossRef]
- Hokkanen, H.; Pimentel, D. New associations in biological control: Theory and practice. Can. Entomol. 1989, 121, 829–840. [Google Scholar] [CrossRef]
- Irvin, N.A.; Hoddle, M.S. Comparative assessments of Gonatocerus ashmeadi and the ‘new association’ parasitoid Gonatocerus tuberculifemur (Hymenoptera: Mymaridae) as biological control agents of Homalodisca vitripennis (Hemiptera: Cicadellidae). Biol. Control 2010, 55, 186–196. [Google Scholar] [CrossRef]
- O’Connell, D.M.; Wratten, S.D.; Pugh, A.R.; Barnes, A.-M. ‘New species association’ biological control? Two coccinellid species and an invasive psyllid pest in New Zealand. Biol. Control 2012, 62, 86–92. [Google Scholar] [CrossRef]
- Brown, M.W.; Cameron, E.A.; Williams, F.M. Population model for the gypsy moth (Lepidoptera: Lymantriidae) egg parasite, Ooencyrtus kuvanae (Hymenoptera: Encyrtidae). Environ. Entomol. 1982, 11, 1299–1304. [Google Scholar] [CrossRef]
- Hofstetter, R.W.; Raffa, K.F. Endogenous and exogenous factors affecting parasitism of gypsy moth egg masses by Ooencyrtus kuvanae. Entomol. Exp. Appl. 1998, 88, 123–135. [Google Scholar] [CrossRef]
- Cappaert, D.; McCullough, D.G. Occurrence and seasonal abundance of Atanycolus cappaerti (Hymenoptera: Braconidae), a native parasitoid of emerald ash borer, Agrilus planipennis (Coleoptera: Buprestidae). Great Lakes Entomol. 2009, 42, 16–29. [Google Scholar]
- Ademokoya, B.; Balusu, R.; Ray, C.; Mottern, J.; Fadamiro, H. The first record of Ooencyrtus nezarae (Hymenoptera: Encyrtidae) on kudzu bug (Hemiptera: Plataspidae) in North America. J. Insect Sci. 2018, 18, 1–7. [Google Scholar] [CrossRef]
- Tunca, H.; Venard, M.; Colombel, E.-A.; Tabone, E. A new substitute host and its effects on some biological properties of Ooencyrtus kuvanae. Bull. Entomol. Res. 2017, 107, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Parker, D.L. The interactions of two hymenopterous egg parasites of the gipsy moth, with notes on the larval instars of each. J. Agric. Res. 1933, 46, 23–34. [Google Scholar]
- Weseloh, R.M. Spatial distribution of the gypsy moth (Lepidoptera: Lymantriidae) and some of its parasitoids within the forest environment. Entomophaga 1972, 17, 339–351. [Google Scholar] [CrossRef]
- Zhang, B.; Li, B.-P.; Meng, L. Effects of self-superparasitism and host age on fitness-correlated traits in solitary endoparasitoid wasp Meteorus pulchricornis. J. Insect Sci. 2014, 14, 1–11. [Google Scholar]
- Tunca, H.; Buradino, M.; Colombel, E.-A.; Tabone, E. Tendency and consequences of superparasitism for the parasitoid Ooencyrtus pityocampae (Hymenoptera: Encyrtidae) in parasitizing a new laboratory host, Philosamia ricini (Lepidoptera: Saturniidae). Eur. J. Entomol. 2016, 113, 51–59. [Google Scholar] [CrossRef]
- Tunca, H.; Colombel, E.-A.; Buradino, M.; Galio, F.; Tabone, E. Optimal biological parameters for rearing Ooencyrtus pityocampae on the new laboratory host Philosamia ricini. J. Appl. Entomol. 2015, 140, 1–9. [Google Scholar]
- Montoya, P.; Perez-Lachaud, G.; Liedo, P. Superparasitism in the fruit fly parasitoid Diachasmimorpha longicaudata (Hymenoptera: Braconidae) and the implications for mass rearing and augmentative release. Insects 2012, 3, 900–911. [Google Scholar] [CrossRef]
- Jaramillo, J.; Borgemeister, C.; Setamou, M. Field superparasitism by Phymastichus coffea, a parasitoid of adult coffee berry borer Hypothenemus hampei. Entomol. Exp. Appl. 2006, 119, 231–237. [Google Scholar] [CrossRef]
- Bourgoin, T. FLOW (Fulgoromorpha Lists on The Web): A Knowledge and a Taxonomy Database Dedicated to Planthoppers (Insecta, Hemiptera, Fulgoromorpha, Fulgoroidea) Version 8. French National Museum of Natural History: Paris, France. Available online: https://www.hemiptera-databases.org/flow/ (accessed on 29 October 2018).
- Bartlett, C.R.; O’Brien, L.B.; Wilson, S.W. A review of the planthoppers (Hemiptera: Fulgoroidea) of the United States. Mem. Am. Entomol. Soc. 2014, 50, 1–287. [Google Scholar]
- Planthopper Parasite Moth, Fulgoraecia exigua (H. Edwards, 1882). Butterflies and Moths of North America. Available online: https://www.butterfliesandmoths.org/species/Fulgoraecia-exigua (accessed on 14 November 2018).
 2016;
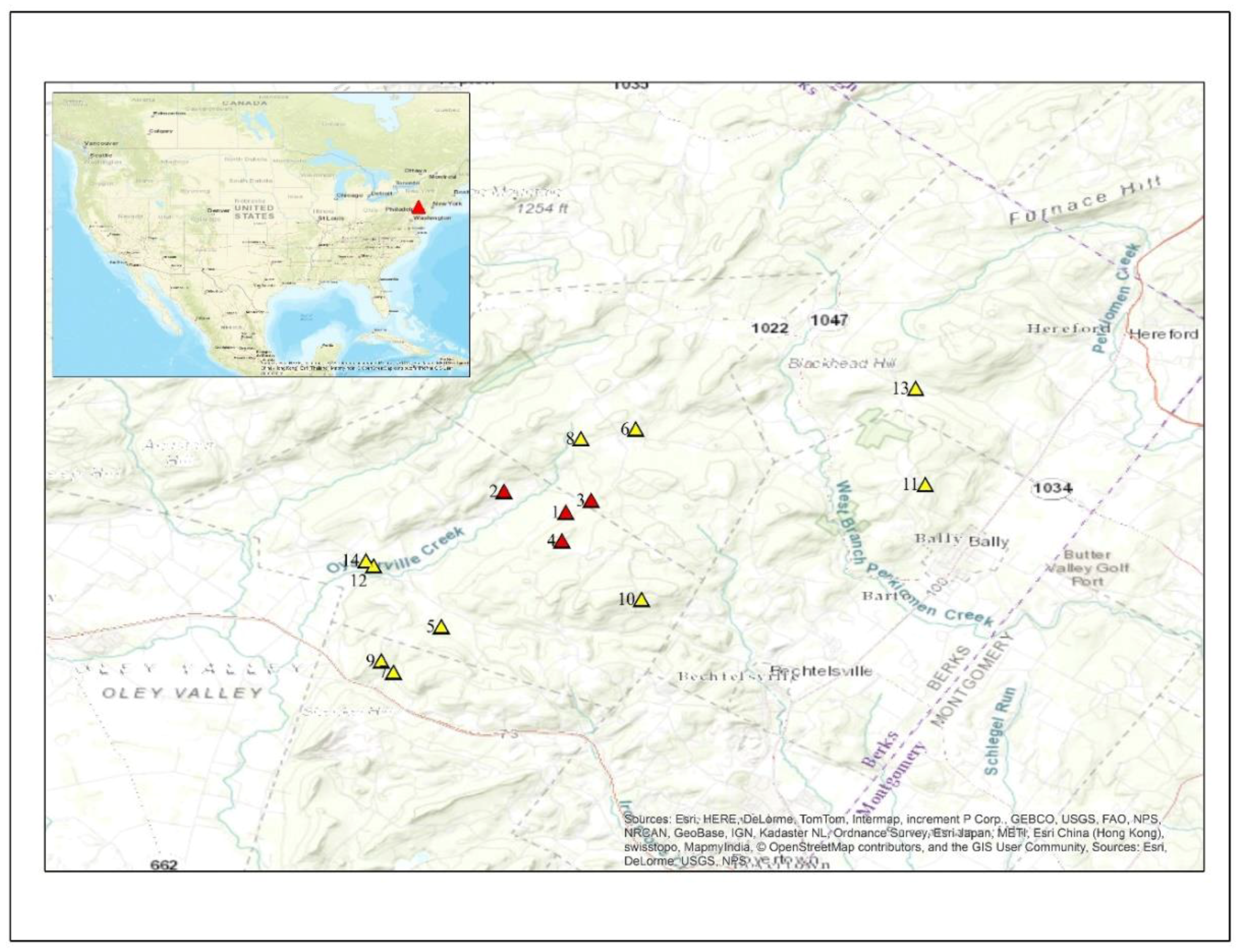
2016;  2017; 1. Rolling (epicenter); 2. ODSouth; 3. Rock; 4. Lutz; 5. Straub; 6. Conrad; 7. WSEast; 8. ODNorth; 9. WSWest; 10. Nuss; 11. Kulps; 12. HCEast; 13. Huffs; 14. HCWest.
2016; 2017; 1. Rolling (epicenter); 2. ODSouth; 3. Rock; 4. Lutz; 5. Straub; 6. Conrad; 7. WSEast; 8. ODNorth; 9. WSWest; 10. Nuss; 11. Kulps; 12. HCEast; 13. Huffs; 14. HCWest.
2017; 1. Rolling (epicenter); 2. ODSouth; 3. Rock; 4. Lutz; 5. Straub; 6. Conrad; 7. WSEast; 8. ODNorth; 9. WSWest; 10. Nuss; 11. Kulps; 12. HCEast; 13. Huffs; 14. HCWest.
2016; 2017; 1. Rolling (epicenter); 2. ODSouth; 3. Rock; 4. Lutz; 5. Straub; 6. Conrad; 7. WSEast; 8. ODNorth; 9. WSWest; 10. Nuss; 11. Kulps; 12. HCEast; 13. Huffs; 14. HCWest.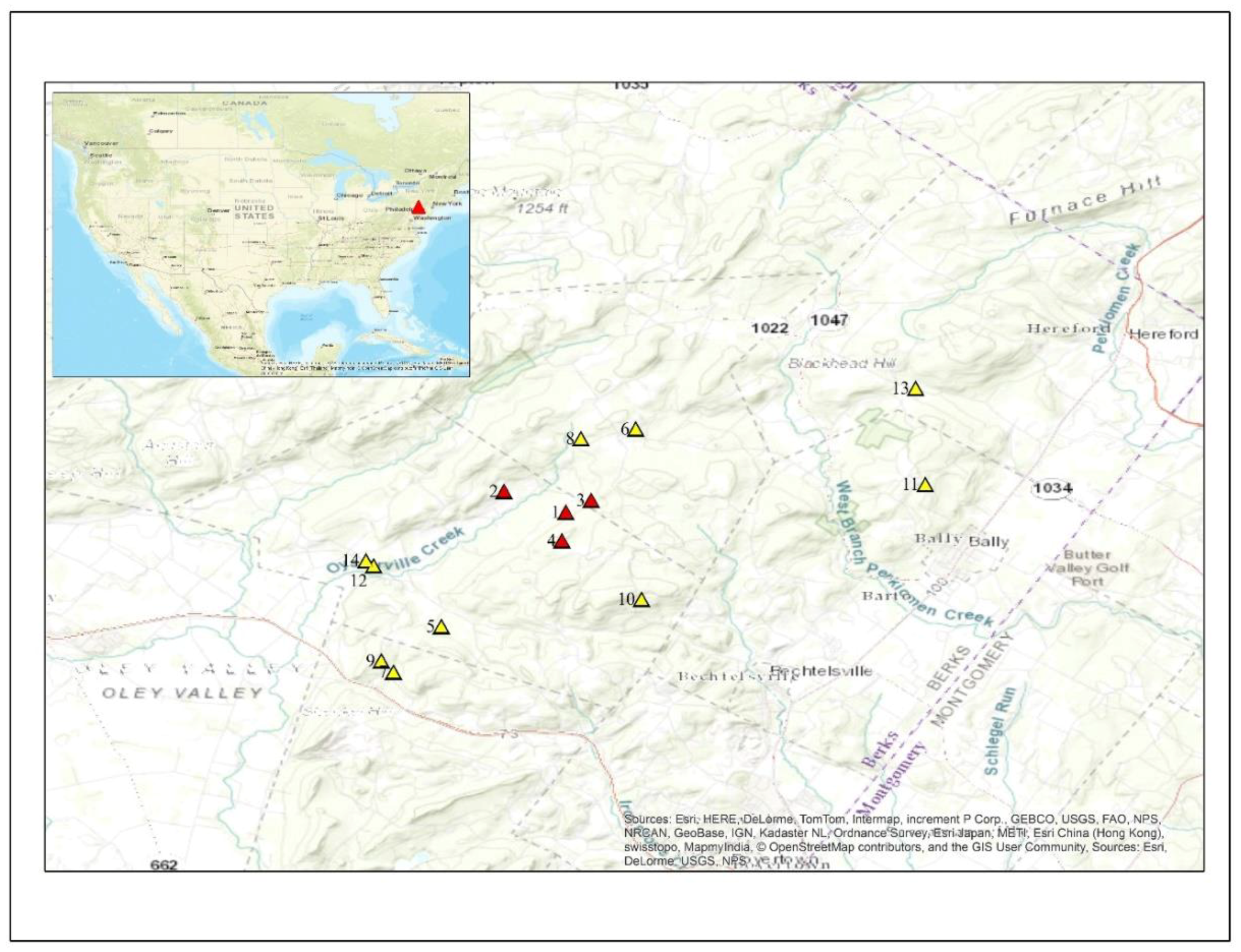


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Name | Oviposition Sites Examined | Dominant Site | No. Egg Masses | No. Eggs | Tree-of-Heaven | Note | |
|---|---|---|---|---|---|---|---|
| Distribution a | Treatment b | ||||||
| 2016 | |||||||
| Rolling | 50 | Black birch | 79 | 2482 | Pockets | Chemical | Infestation epicenter—mixed hardwood dominated by red maple and black cherry |
| ODSouth | 9 | Black cherry | 83 | 3013 | Single trees | Chemical | Family farm with hedgerows of flowering dogwood and black cherry |
| Rock | 25 | Yellow birch | 40 | 1108 | Scattered | Chemical | Mixed hardwood dominated by black cherry and red maple |
| Lutz | 13 | Tree-of-heaven | 60 | 1679 | Pockets | None | Mixed hardwood dominated by northern red oak and black cherry |
| 97 | 262 | 8282 | |||||
| 2017 | |||||||
| Straub | 16 | Tree-of-heaven | 43 | 1165 | Scattered | Mechanical | Mixed hardwood dominated by northern red oak and tulip tree |
| Conrad | 7 | Tree-of-heaven | 19 | 638 | Pockets | Mechanical | Mixed hardwood dominated by tree-of-heaven and tulip tree |
| WSEast | 12 | Metal fence post | 57 | 2085 | Single trees | Chemical | Single yard trees of tree-of-heaven and black walnut |
| ODNorth | 8 | Tulip tree | 18 | 490 | Scattered | Chemical | Mixed hardwood dominated by red maple and black cherry |
| WSWest | 1 | Sweet cherry | 52 | 2573 | Scattered | Chemical | Mixed hardwood dominated by red maple and northern red oak |
| Nuss | 18 | Black birch | 27 | 858 | Scattered | Chemical | Mixed hardwood dominated by red maple and northern red oak |
| Kulps | 22 | Tree-of-heaven | 57 | 1845 | Scattered | Mechanical | Mixed hardwood dominated by red maple and northern red oak |
| HCEast | 13 | Black cherry | 51 | 1919 | Pockets | None | Mixed hardwood dominated by tree-of-heaven and red maple |
| Huffs | 7 | Tree-of-heaven | 14 | 523 | Scattered | Mechanical | Mixed hardwood dominated by northern red oak and red maple |
| HCWest | 7 | Tree-of-heaven | 63 | 1824 | Pockets | Mechanical | Mixed hardwood dominated by tree-of-heaven and white ash |
| 111 | 401 | 13920 | |||||
| 208 | 663 | 22202 | |||||
| Site No. | Species | DBH a (cm) | By Egg Mass | By Egg | ||||
|---|---|---|---|---|---|---|---|---|
| Total | Parasitized | Parasitism (%) b | Total | Parasitized | Parasitism (%) c | |||
| 2016 | ||||||||
| #7 | Black cherry | 7.5 | 20 | 3 | 15.0 | 653 | 28 | 5.0 ± 45.3 |
| #8 | Black cherry | 10.0 | 21 | 2 | 9.5 | 854 | 9 | 1.3 ± 5.5 |
| 41 | 5 | 12.3 ± 1.9 | 1507 | 37 | 3.1 ± 11.7 | |||
| 2017 | ||||||||
| A | White ash | 25.4 | 16 | 3 | 18.8 | 600 | 17 | 2.5 ± 6.7 |
| B | Flowering dogwood (dead) | 30.8 | 11 | 3 | 27.3 | 503 | 3 | 0.5 ± 0.8 |
| C | Black cherry | 17.8 | 9 | 5 | 55.6 | 378 | 13 | 3.9 ± 6.9 |
| D | Flowering dogwood | 35.6 | 5 | 2 | 40.0 | 245 | 4 | 2.2 ± 4.3 |
| 41 | 13 | 35.4 ± 15.0 | 1726 | 37 | 2.2 ± 5.4 | |||
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H. Occurrence, Seasonal Abundance, and Superparasitism of Ooencyrtus kuvanae (Hymenoptera: Encyrtidae) as an Egg Parasitoid of the Spotted Lanternfly (Lycorma delicatula) in North America. Forests 2019, 10, 79. https://doi.org/10.3390/f10020079
Liu H. Occurrence, Seasonal Abundance, and Superparasitism of Ooencyrtus kuvanae (Hymenoptera: Encyrtidae) as an Egg Parasitoid of the Spotted Lanternfly (Lycorma delicatula) in North America. Forests. 2019; 10(2):79. https://doi.org/10.3390/f10020079
Chicago/Turabian StyleLiu, Houping. 2019. "Occurrence, Seasonal Abundance, and Superparasitism of Ooencyrtus kuvanae (Hymenoptera: Encyrtidae) as an Egg Parasitoid of the Spotted Lanternfly (Lycorma delicatula) in North America" Forests 10, no. 2: 79. https://doi.org/10.3390/f10020079
APA StyleLiu, H. (2019). Occurrence, Seasonal Abundance, and Superparasitism of Ooencyrtus kuvanae (Hymenoptera: Encyrtidae) as an Egg Parasitoid of the Spotted Lanternfly (Lycorma delicatula) in North America. Forests, 10(2), 79. https://doi.org/10.3390/f10020079