In the analyses, a graph with
and
was employed. In this graph, each node was connected to a maximum of eight neighbors, and edge weights were real numbers uniformly chosen from the range
. The simulations involved distributing
ants across
g groups (see Algorithm 1 line 7), with
g taking values from the set
and
representing the number of ants in the group. The specific set considered was a subset of all possible divisors of
n, that was selected based on practical considerations and to cover a range of scenarios. Smaller group sizes (
) provided insight into the performance of small groups, while larger group sizes (
) simulated scenarios where collaboration involved more ants. This approach struck a balance between granularity analysis and computational efficiency, ensuring a thorough exploration of various group sizes while maintaining practical feasibility in terms of computational resources. Exploration by groups started after a fixed time
following the previous group, and an overall time limit
was established for the entire colony to reach the exit. This limit was defined as
, with
c set to 150. Individual ants had specific time windows to reach the exit, determined by
. Pheromone reduction occurred at a fixed degradation interval of
, and the global updating rule took place every
ticks with evaporation rates of
and
. The initial pheromone value was
, as expressed in Algorithm 1 line 2, and the parameter determining the amount of pheromone released by ants after crossing an edge was
(as in Algorithm 1 line 32). To assess the impact of group size, 10 independent simulations were conducted for each value of
g. Three different kinds of analysis were performed: an
overall analysis to evaluate the performance of the colony, a
group analysis for every group configuration to evaluate the performance of the single groups, and a
pheromone distribution analysis to identify correlations between the temporal diffusion of pheromones and the navigation behavior of ants. All variables and parameters of interest in our model are listed in
Table 1 for clarity.
3.2. Group Analysis
In the group analysis phase, adjustments were made to the path costs considering each group’s success rate. The success rate, denoted as , is determined by the ratio of ants that successfully reached the exit to the total number of ants within the group.
When the colony is composed of a single group, as in
Table 3, the success rates at both evaporation rates are comparable (
at
and 0.848 at
), indicating consistent performance. The normalized cost considering the success rate allows for a more accurate evaluation. For
the cost is
, while at
the corresponding value is
. These results suggest that despite minor variations the colony maintains relatively consistent performance in terms of success rates and cost across different evaporation rates.
Let us analyze now the results when we have more than one group. In what follows, the index of the group will denote its turn in starting the exploration. Thus, group 1 starts first, then group 2, and so on.
When the colony is divided into two groups, as in
Table 4, for
group 1, which is the one that starts first, has the highest success rate of
and a lower cost of
. In comparison, the second group, group 2, exhibits a slightly lower success rate of
and a marginally higher cost of
. Shifting to
, group 1 maintains a consistently high success rate of
. However, this comes with a higher cost of
. On the other hand, group 2 shows a notable improvement, achieving the highest success rate in this context at
, accompanied by the lowest cost of
.
When the colony is divided into five distinct groups (
), as in
Table 5, under
group 1 shows a remarkable success rate of
accompanied by a cost of
. The subsequent groups (3, 4, and 5) show signs of performance degradation, as evidenced by varying success rates and costs. Transitioning to
, group 1 maintains the best performance across both metrics, with a success rate of
and a cost of
. However, similarly to the
scenario, groups 2, 3, 4, and 5 exhibit varying degrees of success rates and costs, denoting a slight general degradation in performance as we move across the groups.
When the colony is divided into ten groups (
), as in
Table 6, under
group 1 distinguishes itself with an impressive success rate of
, showing a high level of efficiency in reaching the exit. Following closely, group 2 achieves a slightly lower success rate of
but maintains competitive values for the cost. Group 3 stands out with the lowest cost of
, emphasizing its cost-effectiveness despite a moderate success rate. Transitioning to
, group 1 continues to lead with the highest success rate of
. Group 2 shows improvement, achieving a success rate of
with the lowest cost of
. Despite these strengths, there is a slight trend of performance degradation as we progress through the groups, as one can see in both
scenarios.
When the colony is divided into
groups, as in
Table 7, and
, group 1 emerges as the top performer, with a notable success rate of
accompanied by a competitive cost of
. Group 2 closely follows, achieving a success rate of
and showing competitive values for the cost. Noteworthy is group 6, standing out with the lowest cost of
, emphasizing its cost-effectiveness. Transitioning to
, group 1 maintains a high success rate of
with a slightly increased cost of
. Group 5 exhibits the lowest cost of
. Conversely, group 4 boasts the highest success rate of
coupled with a competitive value for the path cost. Additionally, there is a noticeable trend of performance degradation, especially under
.
Finally, when the colony is divided into
groups, as in
Table 8, under
group 1 stands out, with a perfect success rate of
along with a competitive cost of
. Subsequent groups closely follow with relatively high success rates and competitive values for cost and exit time. Notably, group 11 excels with the lowest path cost of
. However, there is a moderate degradation in performance across successive groups. Transitioning to
, group 2 maintains a perfect success rate of
, coupled with a path cost of
. Meanwhile, group 6 emerges as the top performer, showing a perfect success rate of
and the lowest cost of
. This highlights the group’s remarkable ability to efficiently navigate and exit the environment. Overall, for both values of
, a diverse range of success rates, costs, and exit times is observed across the groups. Some groups excel in achieving high success rates, while others prioritize minimizing costs or achieving faster exit times, providing valuable insights into optimizing the management of larger ant colonies.
3.3. Pheromone Distribution Analysis
We now study the correlation between the temporal distribution of pheromones on the edges and the number of ants navigating the network during the simulation. This involves the systematic computation of the mean and standard deviation of pheromones on the edges, as well as counting how many ants are still in the network.
The mean and standard deviation of pheromones over time provide insights into how ants distribute the pheromone along the edges. The average quantity of pheromone deposited by ants is directly proportional to the number of ants in the network. As the number of ants increases, the quantity of pheromone deposited along the edges also increases. Conversely, when some ants exit the network, the pheromone is deposited in a lower quantity. The standard deviation of the pheromone, on the other hand, measures the degree of heterogeneity of the pheromone on the edges. A low value indicates a higher uniformity of the pheromone, leading to greater difficulty for the ants to exit the network since the edges have more or less the same level of pheromone. Conversely, high values denote a higher degree of heterogeneity of the pheromone and this information can be used by ants to exit the network. Finally, the number of ants that are present in the network at each instant of time allows us to better visualize the groups of ants that enter the network at regular intervals and their persistence in the network. This process was carried out for each group configuration and both values of . The resulting data are presented through three plots discussed before, which are obtained by selecting the best experiment that maximizes the number of ants that exit the network, considering a total of 10 experiments performed for each configuration.
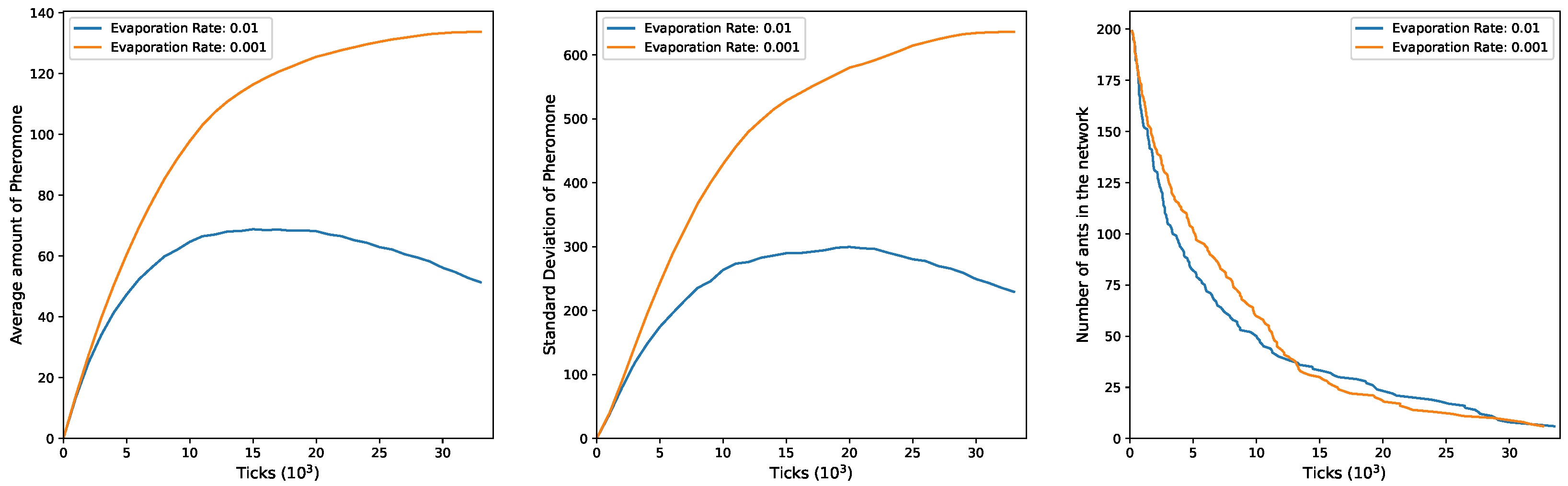
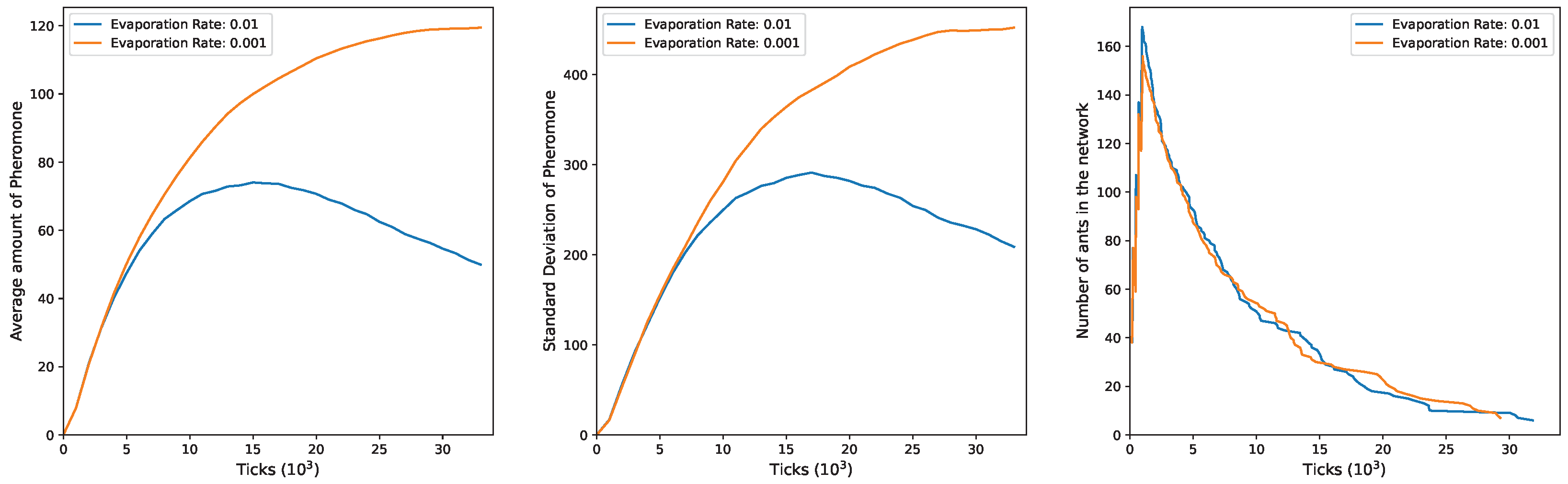
In
Figure 3, the outcomes for
and
are illustrated. In the initial phase of the simulation, there is an increase in the mean of pheromones, coinciding with active exploration by a substantial number of ants in the environment. Subsequently, a decline occurs, more pronounced, with an evaporation rate of
and a slower stabilization at
. This pattern underscores the deep connection between mean pheromone levels and the presence of ants in the network. As ants exit the network, there is a corresponding decrease in the pheromone quantity on edges, particularly clear at a higher evaporation rate. This decrease is attributed to the reduced ability of the remaining ants to counteract the ongoing pheromone evaporation. The second plot mirrors these dynamics, illustrating changes in the standard deviation. Also, in this scenario there is an initial rise in the value during the simulation’s early phase, signifying an increase in the heterogeneity of pheromones as ants release them at the edges. As the ants gradually exit the network, the standard deviation decreases when
and stabilizes when
. This is consistent with the idea that a smaller ant population in the network results in a diminished ability to counteract the evaporation rate, ultimately leading to a more uniform distribution of pheromone levels across the edges. The third plot depicts the evolving number of ants in the network over time, starting at 200, corresponding to the initial exploration group in the
configuration. As time progresses, ants exit with varied trends influenced by the
value. Notably, these trends undergo an inversion around
ticks.
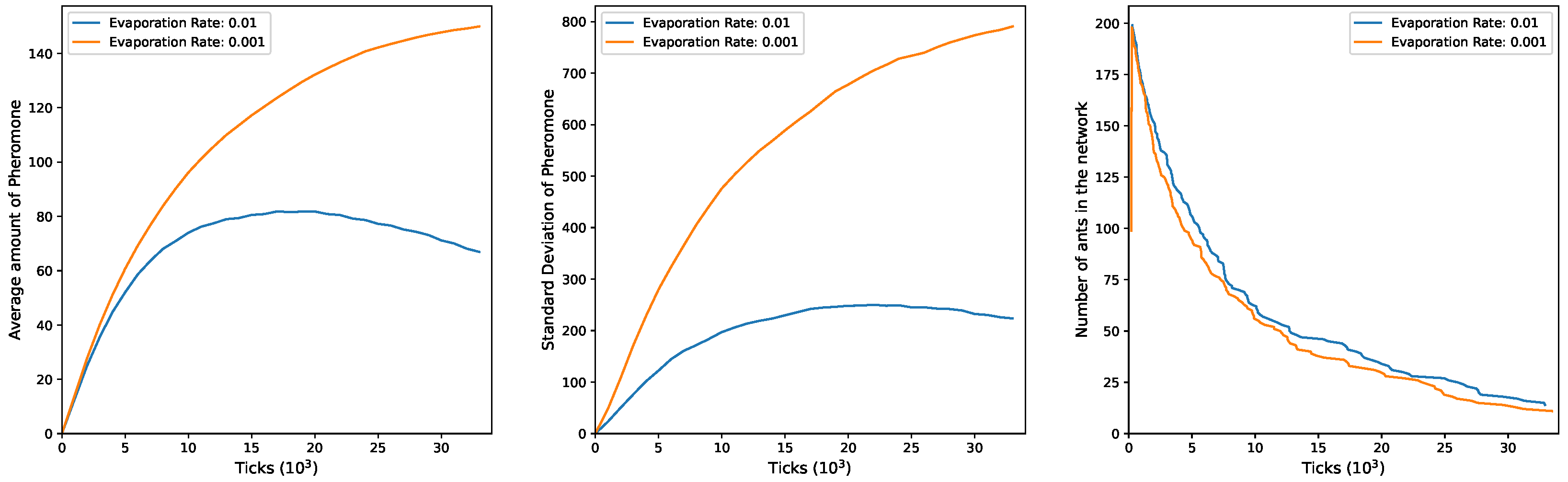
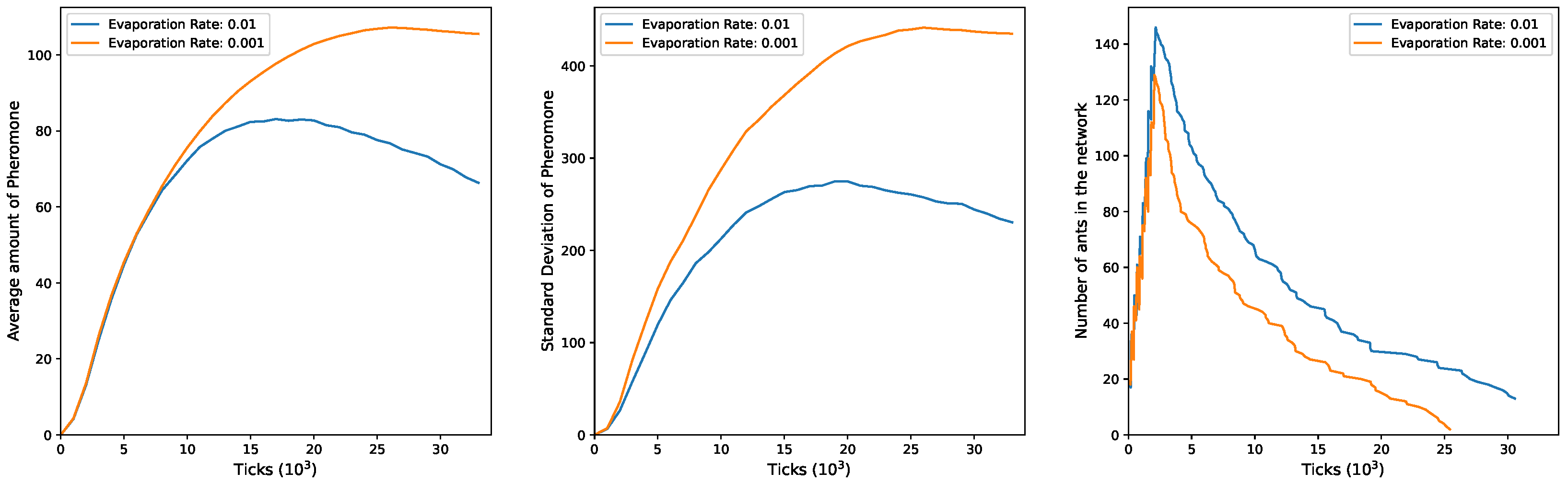
In
Figure 4, the plots for
and
exhibit a trend similar to the previous configuration. Both mean pheromones and standard deviation show an initial increase during the simulation. Subsequently, a decline is observed, but only when
. Under
, both mean and standard deviation continue to rise over time. This discrepancy can be attributed to the more efficient exploratory activity of ants in this setup, resulting in fewer ants being present in the network at any given moment to counteract ongoing pheromone evaporation, particularly with the higher evaporation rate of
. The third plot illustrates the evolving number of ants in the network over time, starting at 100 and peaking at 200, indicating the entry of groups 1 and 2. As time progresses, ants exit more rapidly under
and slower under
.
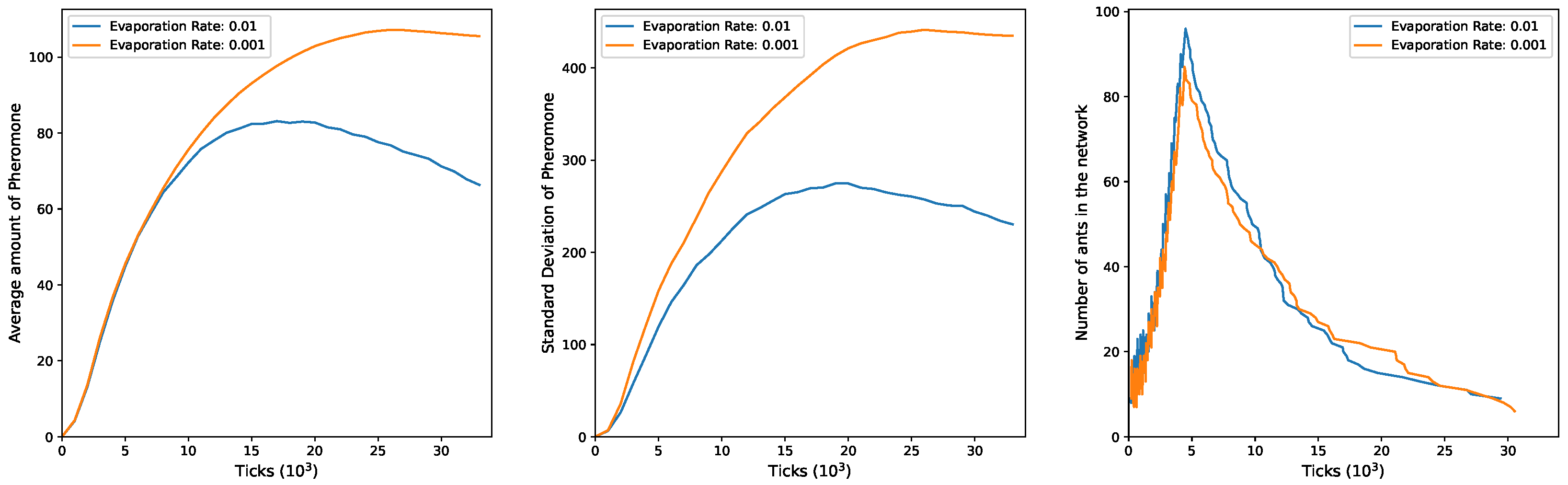
The plots for
and
in
Figure 5 follow a similar trend. Both the mean and the standard deviation exhibit an initial increase during the simulation, followed by a decline, but only when
. Under
, both metrics reach stabilization. As ants gradually exit the environment, their ability to counteract pheromone evaporation is more effective with a low evaporation rate (
), resulting in a higher degree of pheromone heterogeneity. Conversely, when the evaporation rate is higher (
), the remaining ants in the network struggle to counteract pheromone evaporation, leading to a decrease in the mean and heterogeneity. Concerning the number of ants in the environment, it initiates at 40, with the fluctuations in the plot representing the intermittent entry of the five groups. Over time, ants exit at varying rates depending on the value of
.
In
Figure 6 and
Figure 7, the plots for the configurations
and
, as well as
and
, exhibit a trend similar to previous configurations. For both configurations, both the mean and the standard deviation show an initial increase during the simulation, followed by a decline, but only under
. Under
, both metrics in both cases reach a stabilization. In other words, as ants gradually exit the environment their ability to counteract pheromone evaporation is more effective with a low evaporation rate (
), resulting in a prolonged period of higher pheromone heterogeneity. Conversely, when the evaporation rate is higher (
) the remaining ants in the network struggle to counteract pheromone evaporation, leading to a decrease in mean and standard deviation. Regarding the number of ants in the environment, they initiate at 20 and 10, respectively, with the fluctuations in the plot representing the intermittent entry of the other groups (10 and 20, respectively). For
, it is clear that over time ants exit more rapidly when
than when
, suggesting that the ants increasingly benefit from the remaining pheromones on the edges as time unfolds. In contrast, for
this pattern holds true but only until around 11,000 ticks, beyond which the two trends reverse.
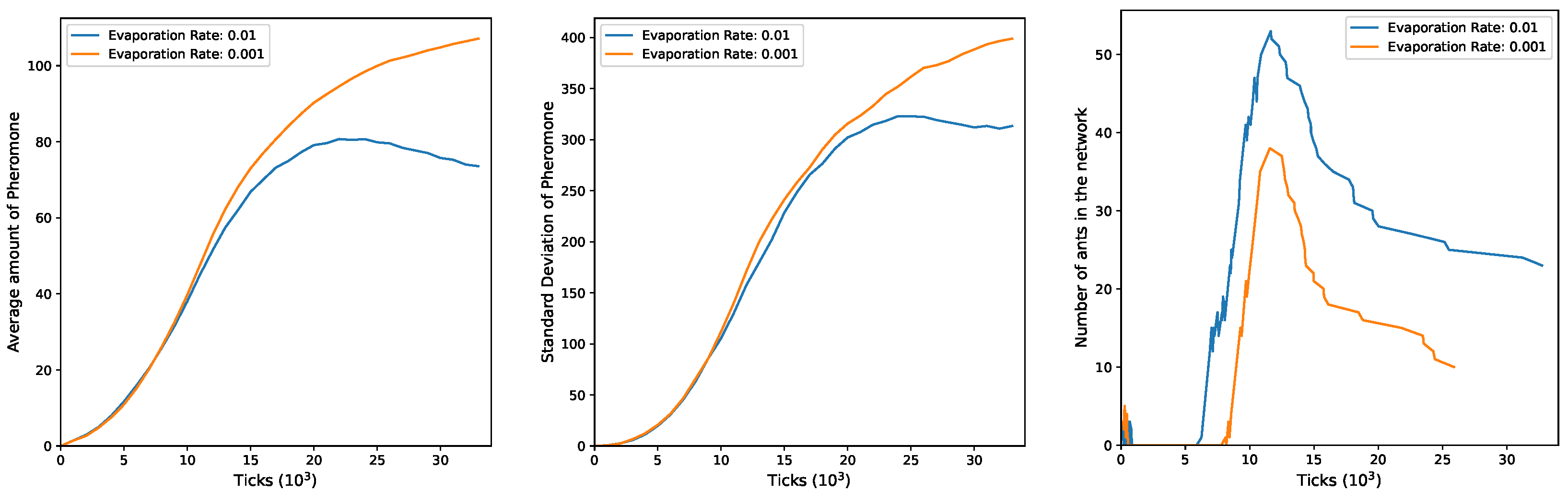
In
Figure 8, for the configuration
and
, both the mean and standard deviation show a continuous increase during the simulation when
. However, with
, the trend in the standard deviation, after an initial phase of increase, starts to decrease, followed by stabilization and a slight subsequent increase. This is in contrast to the mean, which experiences a more pronounced decrease. Furthermore, in this configuration a high degree of heterogeneity is sustained for a more extended period. These behaviors are closely linked to the evolving number of ants in the network, as observed in the third plot. Initially (for
), groups of four ants engage in exploration, swiftly exiting with no noticeable peaks in the curve. Concurrently, other groups are launched, but their rapid exit is not reflected in the curve. In the subsequent time interval (
10,000), there is an increase in the number of ants, marked by fluctuations in the curves. Notably, higher values are observed under
, while lower values are seen at
. Around 11,000 ticks, the number of ants gradually starts to decrease.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}