The Biological Fate of Silver Nanoparticles from a Methodological Perspective




Abstract
1. Introduction and Problem Description
2. Analyses of Analytical Techniques for Quantification of NP Biological Fate in Terms of Their Characteristics and Throughput
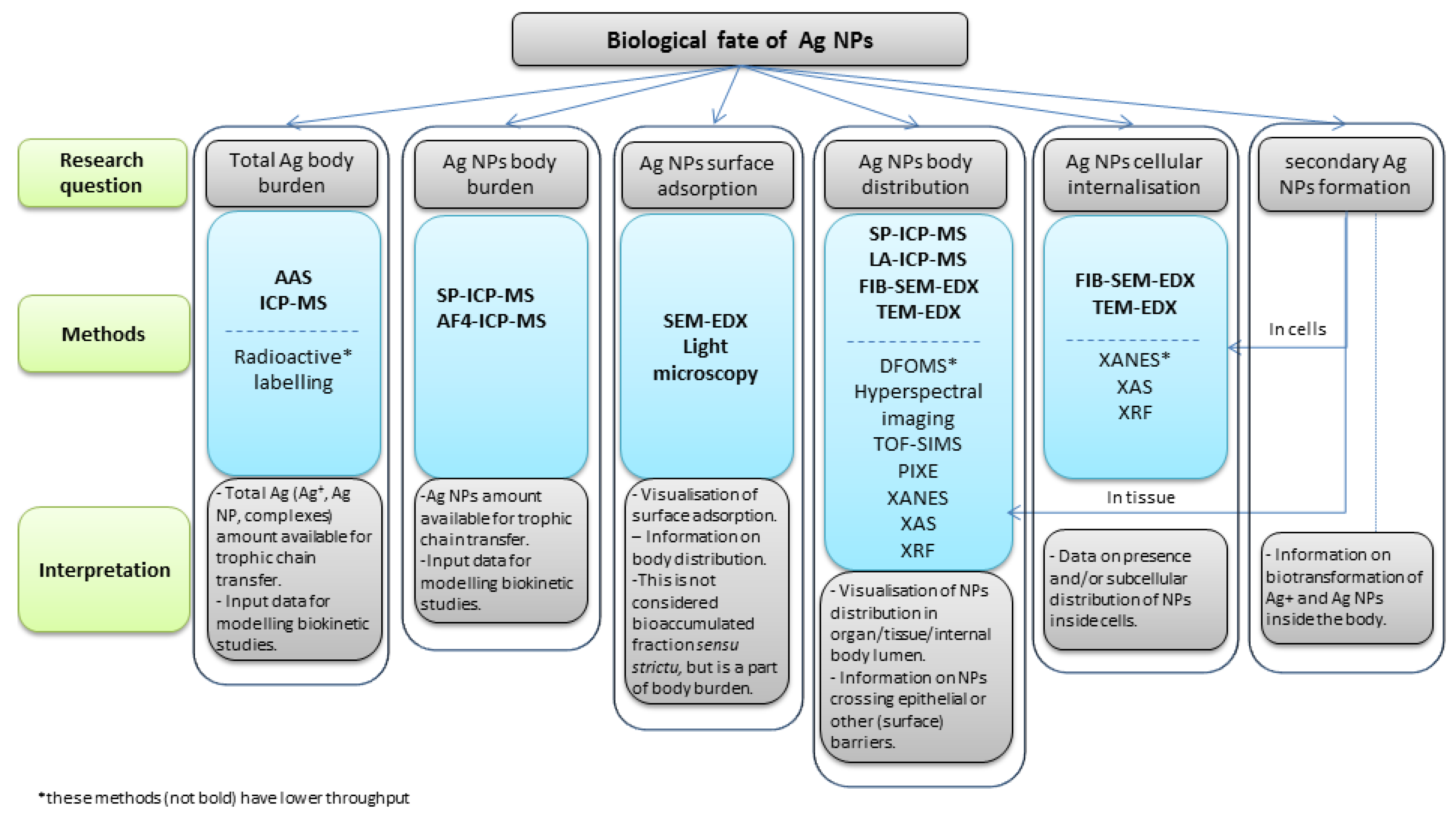
2.1. Characteristics of Techniques by Their Information Output
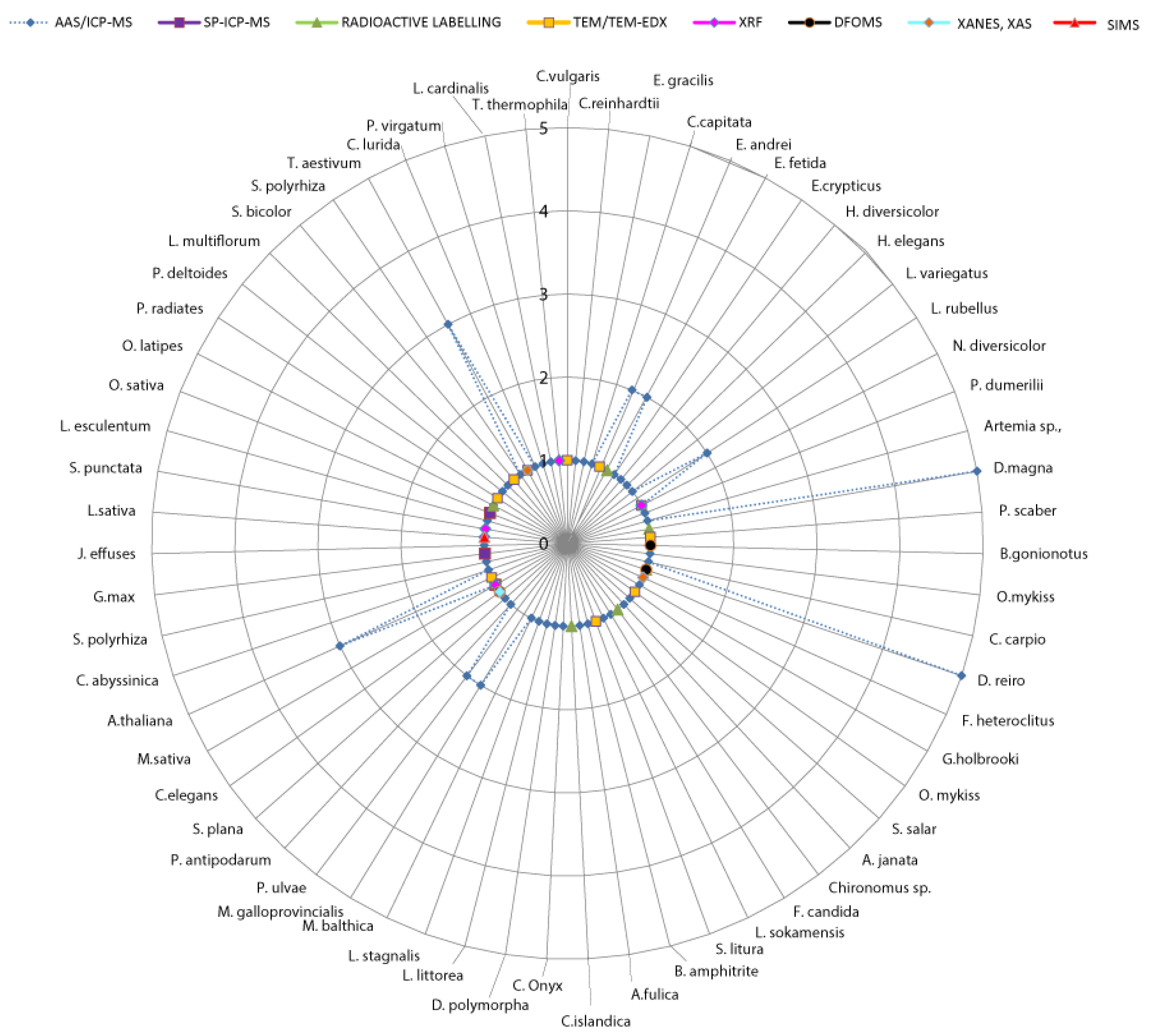
2.2. Throughput
3. Interpretation of Biological Fate Data for Ag NPs
3.1. Body Burden
3.2. Body Distribution
4. Final Conclusions and Outlook
Supplementary Materials
Funding
Conflicts of Interest
Abbreviations
| AAS | atomic absorption spectroscopy |
| AF4-ICP-MS | asymmetric flow field-flow fractionation inductively coupled plasma mass spectrometry |
| DFOM | dark field optical microscopy |
| DFOMS | dark field optical microscopy and spectroscopy |
| EDX | energy dispersive X-ray |
| FIB-SEM-EDX | focused ion beam scanning microscopy coupled with energy-dispersive X-ray spectroscopy |
| ICP-MS | inductively coupled plasma mass spectrometry |
| ICP-OES | inductively coupled plasma optical emission spectroscopy |
| LA-ICP-MS | laser ablation inductively coupled ion mass spectrometry |
| PIXE | proton induced X-ray emission |
| SEM | scanning electron microscopy |
| SIMS | secondary ion mass spectrometry |
| ToF-SIMS | time of flight-secondary ion mass spectrometry |
| SP-ICP-MS | single particle inductively coupled plasma mass spectrometry |
| STEM | scanning transmission electron microscopy |
| TEM | transmission electron microscopy |
| TEM-EDX | transmission electron microscopy coupled with energy-dispersive X-ray spectroscopy |
| XANES | X-ray absorption near edge structure spectroscopy |
| XAS | synchrotron X-ray absorption spectroscopy |
| XRF | X-ray fluorescence |
References
- Kahru, A.; Ivask, A. Mapping the dawn of nanoecotoxicological research. Acc. Chem. Res. 2013, 46, 823–833. [Google Scholar] [CrossRef] [PubMed]
- McGillicuddy, E.; Murray, I.; Kavanagh, S.; Morrison, L.; Fogarty, A.; Cormican, M.; Dockery, P.; Prendergast, M.; Rowan, N.; Morris, D. Silver nanoparticles in the environment: Sources, detection and ecotoxicology. Sci. Total Environ. 2017, 575, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.-M.; Wang, W.-X. Biokinetic uptake and efflux of silver nanoparticles in daphnia magna. Environ. Sci. Technol. 2010, 44, 7699–7704. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.R.; Paul, K.B.; Dybowska, A.D.; Valsami-Jones, E.; Lead, J.R.; Stone, V.; Fernandes, T.F. Accumulation dynamics and acute toxicity of silver nanoparticles to Daphnia magna and Lumbriculus variegatus: Implications for metal modeling approaches. Environ. Sci. Technol. 2015, 49, 4389–4397. [Google Scholar] [CrossRef] [PubMed]
- Sotiriou, G.A.; Meyer, A.; Knijnenburg, J.T.; Panke, S.; Pratsinis, S.E. Quantifying the origin of released Ag+ ions from nanosilver. Langmuir 2012, 28, 15929–15936. [Google Scholar] [CrossRef] [PubMed]
- Behra, R.; Sigg, L.; Clift, M.J.; Herzog, F.; Minghetti, M.; Johnston, B.; Petri-Fink, A.; Rothen-Rutishauser, B. Bioavailability of silver nanoparticles and ions: From a chemical and biochemical perspective. J. R. Soc. Interface 2013, 10, 20130396. [Google Scholar] [CrossRef] [PubMed]
- Ivask, A.; George, S.; Bondarenko, O.; Kahru, A. Metal-containing nano-antimicrobials: Differentiating the impact of solubilized metals and particles. In Nano-Antimicrobials; Springer: Berlin/Heidelberg, Germany, 2012; pp. 253–290. [Google Scholar]
- Levard, C.M.; Mitra, S.; Yang, T.; Jew, A.D.; Badireddy, A.R.; Lowry, G.V.; Brown, G.E., Jr. Effect of chloride on the dissolution rate of silver nanoparticles and toxicity to E. coli. Environ. Sci. Technol. 2013, 47, 5738–5745. [Google Scholar] [CrossRef] [PubMed]
- Levard, C.; Reinsch, B.C.; Michel, F.M.; Oumahi, C.; Lowry, G.V.; Brown, G.E., Jr. Sulfidation processes of PVP-coated silver nanoparticles in aqueous solution: Impact on dissolution rate. Environ. Sci. Technol. 2011, 45, 5260–5266. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.K.; Dybowska, A.; Berhanu, D.; Luoma, S.N.; Valsami-Jones, E. The complexity of nanoparticle dissolution and its importance in nanotoxicological studies. Sci. Total Environ. 2012, 438, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Blinova, I.; Niskanen, J.; Kajankari, P.; Kanarbik, L.; Käkinen, A.; Tenhu, H.; Penttinen, O.-P.; Kahru, A. Toxicity of two types of silver nanoparticles to aquatic crustaceans Daphnia magna and Thamnocephalus platyurus. Environ. Sci. Pollut. Res. 2013, 20, 3456–3463. [Google Scholar] [CrossRef] [PubMed]
- Izak-Nau, E.; Huk, A.; Reidy, B.; Uggerud, H.; Vadset, M.; Eiden, S.; Voetz, M.; Himly, M.; Duschl, A.; Dusinska, M. Impact of storage conditions and storage time on silver nanoparticles’ physicochemical properties and implications for their biological effects. RSC Adv. 2015, 5, 84172–84185. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; McLean, J.E.; Martineau, N.; Britt, D.W.; Haverkamp, R.; Anderson, A.J. Silver nanoparticles disrupt wheat (Triticum aestivum L.) growth in a sand matrix. Environ. Sci. Technol. 2013, 47, 1082–1090. [Google Scholar] [CrossRef] [PubMed]
- Geisler-Lee, J.; Wang, Q.; Yao, Y.; Zhang, W.; Geisler, M.; Li, K.; Huang, Y.; Chen, Y.; Kolmakov, A.; Ma, X. Phytotoxicity, accumulation and transport of silver nanoparticles by Arabidopsis thaliana. Nanotoxicology 2012, 7, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Larue, C.; Castillo-Michel, H.; Sobanska, S.; Cécillon, L.; Bureau, S.; Barthès, V.; Ouerdane, L.; Carrière, M.; Sarret, G. Foliar exposure of the crop Lactuca sativa to silver nanoparticles: Evidence for internalization and changes in Ag speciation. J. Hazard. Mater. 2014, 264, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R. Synthesis of Silver Nanoparticles in Photosynthetic Plants. J. Nanopart. 2014, 2014, 963961. [Google Scholar] [CrossRef]
- Juganson, K.; Mortimer, M.; Ivask, A.; Kasemets, K.; Kahru, A. Extracellular conversion of silver ions into silver nanoparticles by protozoan Tetrahymena thermophila. Environ. Sci. Process. Impacts 2013, 15, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Mohanpuria, P.; Rana, N.K.; Yadav, S.K. Biosynthesis of nanoparticles: Technological concepts and future applications. J. Nanopart. Res. 2008, 10, 507–517. [Google Scholar] [CrossRef]
- Quester, K.; Avalos-Borja, M.; Castro-Longoria, E. Biosynthesis and microscopic study of metallic nanoparticles. Micron 2013, 54, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Nam, S.-H.; An, Y.-J. Interaction of silver nanoparticles with biological surfaces of Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2012, 77, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.-Y.; Sim, S.J.; Yi, J.; Park, K.; Chung, K.H.; Ryu, D.-Y.; Choi, J. Ecotoxicity of silver nanoparticles on the soil nematode Caenorhabditis elegans using functional ecotoxicogenomics. Environ. Sci. Technol. 2009, 43, 3933–3940. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.N.; Lord, C.A.; Yang, X.Y.; Turner, E.A.; Badireddy, A.R.; Marinakos, S.M.; Chilkoti, A.; Wiesner, M.R.; Auffan, M. Intracellular uptake and associated toxicity of silver nanoparticles in Caenorhabditis elegans. Aquat. Toxicol. 2010, 100, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Jiang, C.; Hsu-Kim, H.; Badireddy, A.R.; Dykstra, M.; Wiesner, M.; Hinton, D.E.; Meyer, J.N. Silver nanoparticle behavior, uptake, and toxicity in Caenorhabditis elegans: Effects of natural organic matter. Environ. Sci. Technol. 2014, 48, 3486–3495. [Google Scholar] [CrossRef] [PubMed]
- Ivask, A.; Juganson, K.; Bondarenko, O.; Mortimer, M.; Aruoja, V.; Kasemets, K.; Blinova, I.; Heinlaan, M.; Slaveykova, V.; Kahru, A. Mechanisms of toxic action of Ag, ZnO and CuO nanoparticles to selected ecotoxicological test organisms and mammalian cells in vitro: A comparative review. Nanotoxicology 2014, 8 (Suppl. 1), 57–71. [Google Scholar] [CrossRef] [PubMed]
- Monopoli, M.P.; Åberg, C.; Salvati, A.; Dawson, K.A. Biomolecular coronas provide the biological identity of nanosized materials. Nat. Nanotechnol. 2012, 7, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.; Powell, K.; Crossley, A.; Jurkschat, K.; Kille, P.; Morgan, A.J.; Read, D.; Tyne, W.; Lahive, E.; Svendsen, C. Analytical approaches to support current understanding of exposure, uptake and distributions of engineered nanoparticles by aquatic and terrestrial organisms. Ecotoxicology 2015, 24, 239–261. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Advanced analytical techniques for the measurement of nanomaterials in food and agricultural samples: A review. Environ. Eng. Sci. 2013, 30, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, C.J.; Vermeire, T.G. Risk Assessment of Chemicals: An Introduction; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- OECD. Test No. 305: Bioaccumulation in Fish: Aqueous and Dietary Exposure; OECD Publishing: Paris, France, 2012; Available online: https://www.oecd-ilibrary.org/environment/test-no-305-bioaccumulation-in-fish-aqueous-and-dietary-exposure_9789264185296-en (accessed on 5 June 2018).
- Chen, Y.; Si, Y.; Zhou, D.; Dang, F. Differential bioaccumulation patterns of nanosized and dissolved silver in a land snail Achatina fulica. Environ. Pollut. 2017, 222, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Novak, S.; Drobne, D.; Golobič, M.; Zupanc, J.; Romih, T.; Gianoncelli, A.; Kiskinova, M.; Kaulich, B.; Pelicon, P.; Vavpetič, P. Cellular internalization of dissolved cobalt ions from ingested CoFe2O4 nanoparticles: In vivo experimental evidence. Environ. Sci. Technol. 2013, 47, 5400–5408. [Google Scholar] [CrossRef] [PubMed]
- Novak, S.; Drobne, D.; Valant, J.; Pipan-Tkalec, Ž.; Pelicon, P.; Vavpetič, P.; Grlj, N.; Falnoga, I.; Mazej, D.; Remškar, M. Cell membrane integrity and internalization of ingested TiO2 nanoparticles by digestive gland cells of a terrestrial isopod. Environ. Toxicol. Chem. 2012, 31, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Lešer, V.; Drobne, D.; Pipan, Ž.; Milani, M.; Tatti, F. Comparison of different preparation methods of biological samples for FIB milling and SEM investigation. J. Microsc. 2009, 233, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Lešer, V.; Milani, M.; Tatti, F.; Tkalec, Ž. P.; Štrus, J.; Drobne, D. Focused ion beam (FIB)/scanning electron microscopy (SEM) in tissue structural research. Protoplasma 2010, 246, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Laborda, F.; Bolea, E.; Jiménez-Lamana, J. Single Particle Inductively Coupled Plasma Mass Spectrometry: A Powerful Tool for Nanoanalysis; ACS Publications: Washington, DC, USA, 2013. [Google Scholar]
- Kaulich, B.; Gianoncelli, A.; Beran, A.; Eichert, D.; Kreft, I.; Pongrac, P.; Regvar, M.; Vogel-Mikuš, K.; Kiskinova, M. Low-energy X-ray fluorescence microscopy opening new opportunities for bio-related research. J. R. Soc. Interface 2009, 6 (Suppl. 5), S641–S647. [Google Scholar] [CrossRef] [PubMed]
- Eichert, D.; Gregoratti, L.; Kaulich, B.; Marcello, A.; Melpignano, P.; Quaroni, L.; Kiskinova, M. Imaging with spectroscopic micro-analysis using synchrotron radiation. Anal. Bioanal. Chem. 2007, 389, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, S.-I.; Fujii, S.-I.; Shigeta, K.; Inagaki, K. Single Cell Analysis by Using ICP-MS. In Metallomics; Springer: Berlin/Heidelberg, Germany, 2017; pp. 107–124. [Google Scholar]
- Drescher, D.; Giesen, C.; Traub, H.; Panne, U.; Kneipp, J.; Jakubowski, N. Quantitative imaging of gold and silver nanoparticles in single eukaryotic cells by laser ablation ICP-MS. Anal. Chem. 2012, 84, 9684–9688. [Google Scholar] [CrossRef] [PubMed]
- Benninghoven, A. Chemical analysis of inorganic and organic surfaces and thin films by static time-of-flight secondary ion mass spectrometry (TOF-SIMS). Angew. Chem. Int. Ed. Engl. 1994, 33, 1023–1043. [Google Scholar] [CrossRef]
- Mitrano, D.M.; Barber, A.; Bednar, A.; Westerhoff, P.; Higgins, C.P.; Ranville, J.F. Silver nanoparticle characterization using single particle ICP-MS (SP-ICP-MS) and asymmetrical flow field flow fractionation ICP-MS (AF4-ICP-MS). J. Anal. Atom. Spectrom. 2012, 27, 1131–1142. [Google Scholar] [CrossRef]
- Auffan, M.; Matson, C.W.; Rose, J.; Arnold, M.; Proux, O.; Fayard, B.; Liu, W.; Chaurand, P.; Wiesner, M.R.; Bottero, J.-Y. Salinity-dependent silver nanoparticle uptake and transformation by Atlantic killifish (Fundulus heteroclitus) embryos. Nanotoxicology 2014, 8 (Suppl. 1), 167–176. [Google Scholar] [CrossRef] [PubMed]
- Kwok, K.W.; Auffan, M.; Badireddy, A.R.; Nelson, C.M.; Wiesner, M.R.; Chilkoti, A.; Liu, J.; Marinakos, S.M.; Hinton, D.E. Uptake of silver nanoparticles and toxicity to early life stages of Japanese medaka (Oryzias latipes): Effect of coating materials. Aquat. Toxicol. 2012, 120, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Nallathamby, P.D.; Browning, L.M.; Osgood, C.J.; Xu, X.-H.N. In vivo imaging of transport and biocompatibility of single silver nanoparticles in early development of zebrafish embryos. ACS Nano 2007, 1, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.; Hussain, S.M.; Schrand, A.M.K.; Braydich-Stolle, L.; Hess, K.L.; Jones, R.L.; Schlager, J.J. Unique cellular interaction of silver nanoparticles: Size-dependent generation of reactive oxygen species. J. Phys. Chem. B 2008, 112, 13608–13619. [Google Scholar] [CrossRef] [PubMed]
- Miao, A.-J.; Luo, Z.; Chen, C.-S.; Chin, W.-C.; Santschi, P.H.; Quigg, A. Intracellular uptake: A possible mechanism for silver engineered nanoparticle toxicity to a freshwater alga Ochromonas danica. PLoS ONE 2010, 5, e15196. [Google Scholar] [CrossRef] [PubMed]
- García-Alonso, J.; Khan, F.R.; Misra, S.K.; Turmaine, M.; Smith, B.D.; Rainbow, P.S.; Luoma, S.N.; Valsami-Jones, E. Cellular internalization of silver nanoparticles in gut epithelia of the estuarine polychaete Nereis diversicolor. Environ. Sci. Technol. 2011, 45, 4630–4636. [Google Scholar] [CrossRef] [PubMed]
- Lahive, E.; Jurkschat, K.; Shaw, B.J.; Handy, R.D.; Spurgeon, D.J.; Svendsen, C. Toxicity of cerium oxide nanoparticles to the earthworm Eisenia fetida: Subtle effects. Environ. Chem. 2014, 11, 268–278. [Google Scholar] [CrossRef]
- Handy, R.D.; Henry, T.B.; Scown, T.M.; Johnston, B.D.; Tyler, C.R. Manufactured nanoparticles: Their uptake and effects on fish—A mechanistic analysis. Ecotoxicology 2008, 17, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Brun, N.R.; Beenakker, M.M.; Hunting, E.R.; Ebert, D.; Vijver, M.G. Brood pouch-mediated polystyrene nanoparticle uptake during Daphnia magna embryogenesis. Nanotoxicology 2017, 11, 1059–1069. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| (A) Characteristics of Techniques | AAS, ICP-MS (OES) | SP-ICP-MS, AF4-ICP-MS | Tracing Labelled NPs | LA-ICP-MS | XANES, XAS | XRF | SIMS | PIXE | FIB-SEM-EDX | TEM-EDX | Hyperspectral Imaging | DFOMS |
| Quantification in decomposed tissue | + | + | + | - | - | + | + | - | - | - | - | |
| Power to quantify Ag NPs in the sample | - | + | - | - | + | - | - | - | - | - | - | - |
| Power to discriminate between Ag ions and NPs | - | + | - | - | + | - | + | - | - | - | + | - |
| Visualization on tissue slices | - | - | + | + | + | + | + | + | + | + | + | + |
| Visualization of Ag species at the tissue level | - | + | + | + | + | + | + | + | + | + | + | + |
| Visualization of Ag species at the subcellular level | - | - | + | - | + | + | + | + | + | + | - | - |
| (B) Throughput | AAS, ICP-MS (OES) | SP-ICP-MS, AF4-ICP-MS | Tracing Labelled NPs | LA-ICP-MS | XANES, XAS | XRF | SIMS | PIXE | FIB-SEM-EDX | TEM-EDX | Hyperspectral Imaging | DFOMS |
| Time input for sample preparation * | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 3 | 0 | 0 |
| Time input for data acquisition; processing † | 0 | 0 | 0 | 2 | 3 | 3 | 3 | 1 | 1 | 1 | 2 | 2 |
| Difficulty to access ‡ | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| Final score $ | 1 | 1 | 3 | 5 | 6 | 6 | 6 | 4 | 4 | 4 | 3 | 3 |
| Throughput & | high | high | medium | low | low | low | low | medium | medium | medium | medium | medium |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drobne, D.; Novak, S.; Talaber, I.; Lynch, I.; Kokalj, A.J. The Biological Fate of Silver Nanoparticles from a Methodological Perspective. Materials 2018, 11, 957. https://doi.org/10.3390/ma11060957
Drobne D, Novak S, Talaber I, Lynch I, Kokalj AJ. The Biological Fate of Silver Nanoparticles from a Methodological Perspective. Materials. 2018; 11(6):957. https://doi.org/10.3390/ma11060957
Chicago/Turabian StyleDrobne, Damjana, Sara Novak, Iva Talaber, Iseult Lynch, and Anita Jemec Kokalj. 2018. "The Biological Fate of Silver Nanoparticles from a Methodological Perspective" Materials 11, no. 6: 957. https://doi.org/10.3390/ma11060957
APA StyleDrobne, D., Novak, S., Talaber, I., Lynch, I., & Kokalj, A. J. (2018). The Biological Fate of Silver Nanoparticles from a Methodological Perspective. Materials, 11(6), 957. https://doi.org/10.3390/ma11060957