
Assessing Different Inoculum Treatments for Improved Production of Hydrogen through Dark Fermentation

 ,
, 
Abstract
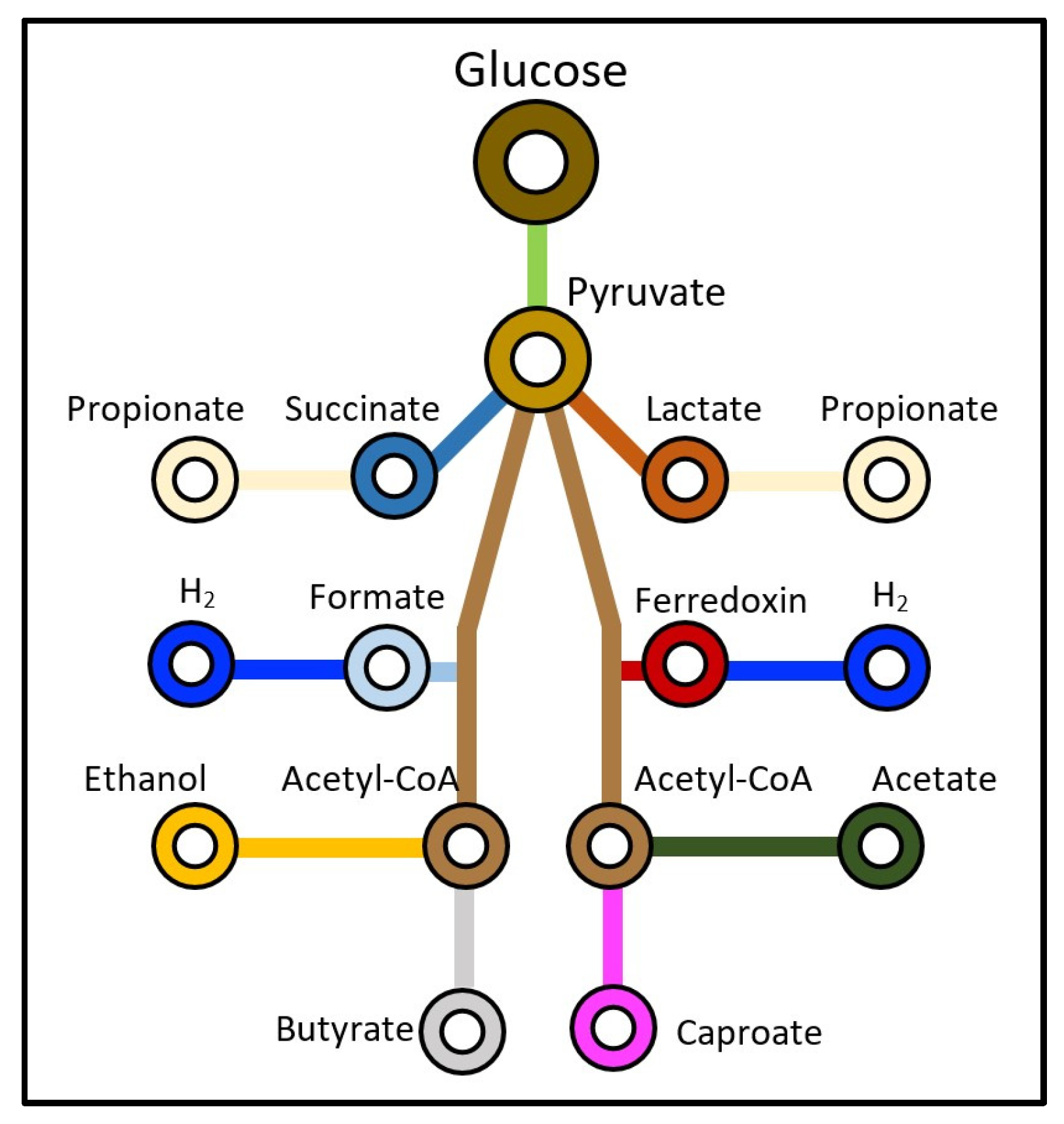
1. Introduction
2. Materials and Methods
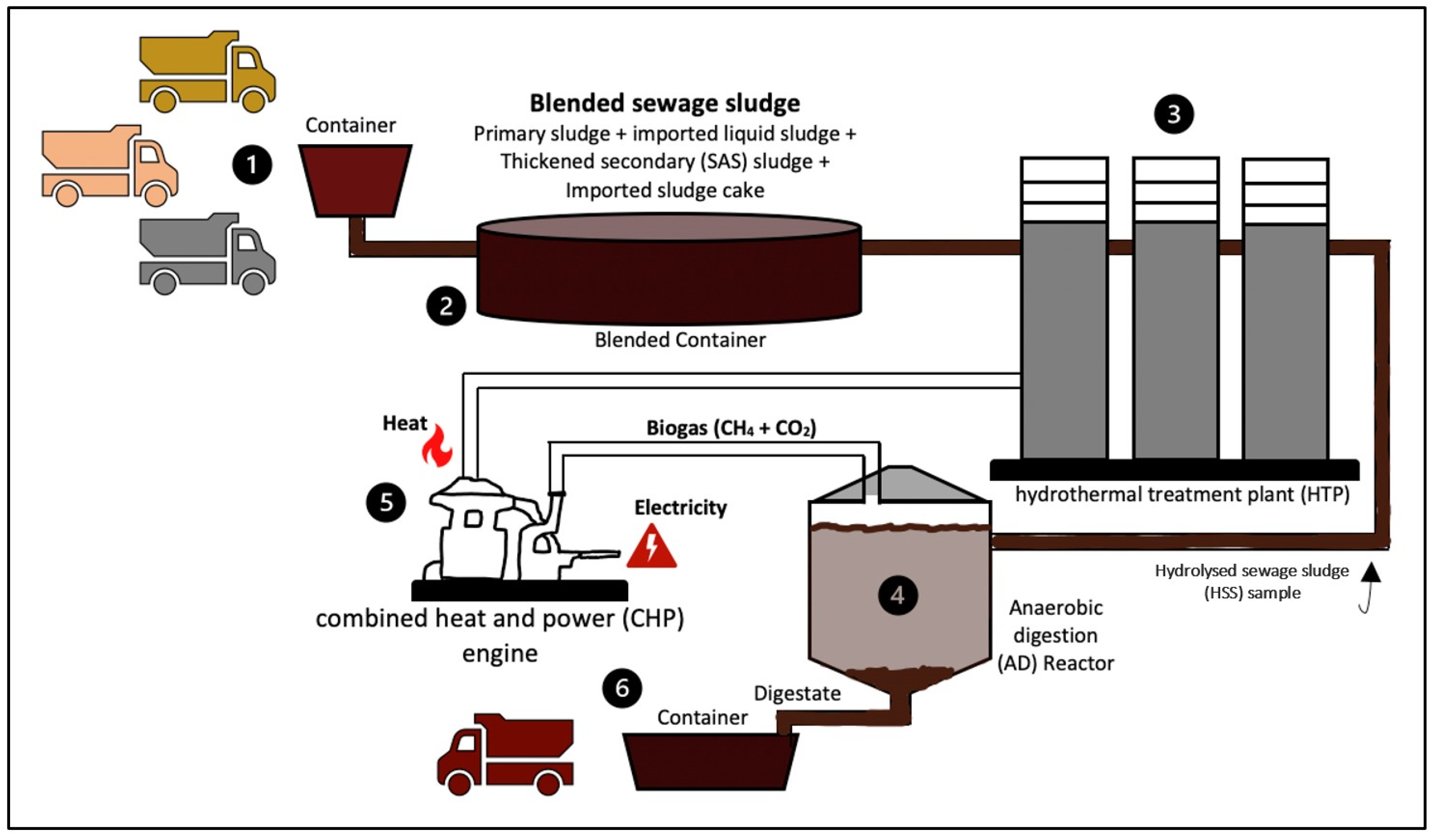
2.1. Inoculum and Substrate
2.2. Inoculum Pre-Treatment and Experimental Setup
- Acid-shock pre-treatment (AST) was performed by adjusting the pH of the digestate to pH 3 using 1 M HCl and storing it in a fridge at 4 °C for 24 h. After 24 h, the pH was returned to pH 7 using 1 M NaOH.
- Basic-shock pre-treatment (BST) was performed by adjusting the pH of the digestate to pH 10 using 1 M NaOH and storing it in a fridge at 4 °C for 24 h. After 24 h, the pH was returned to pH 7 using 1 M HCl.
- Heat-shock pre-treatment (HST) was conducted by heating the digestate for 20 min at 115 °C using a standard autoclave at approximately 1.5 bar.
2.3. Analytical Methods
2.3.1. Gas Analysis
2.3.2. Liquid Analysis
2.3.3. Statistical Analysis
2.3.4. Energy Balance of BHP Tests
3. Results and Discussion
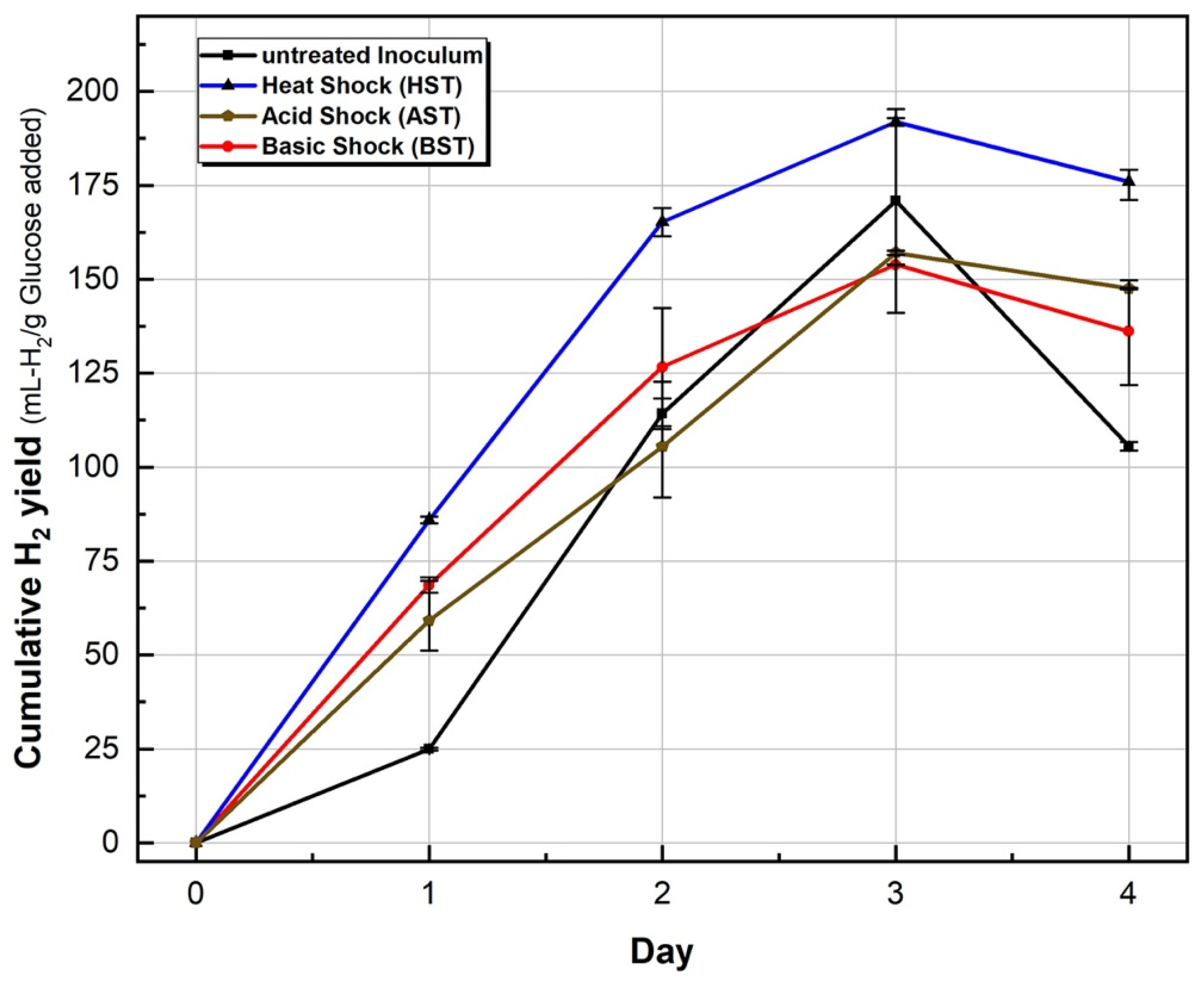
3.1. Hydrogen Yields
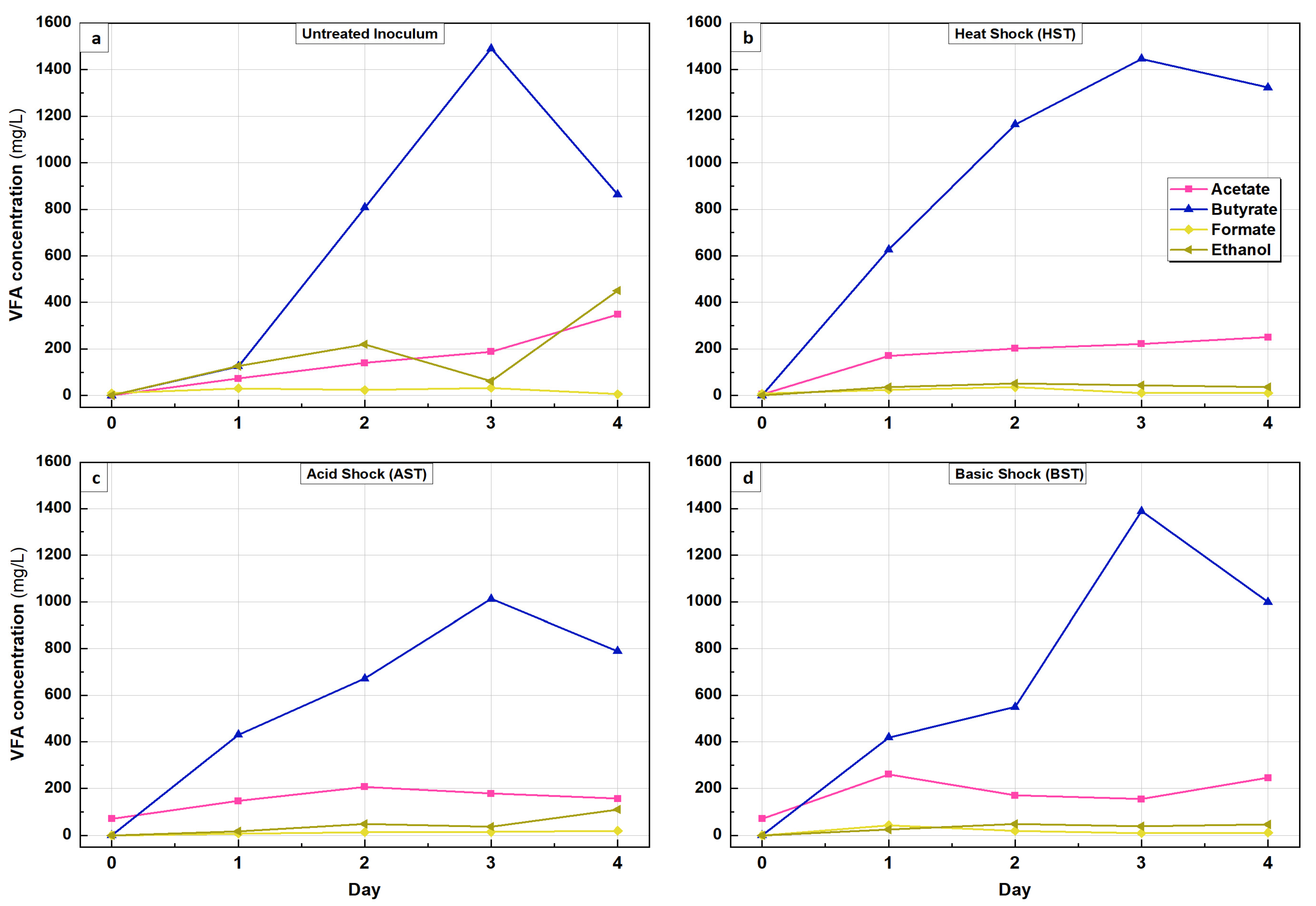
3.2. Volatile Fatty Acids and By-Product from Glucose Transformation
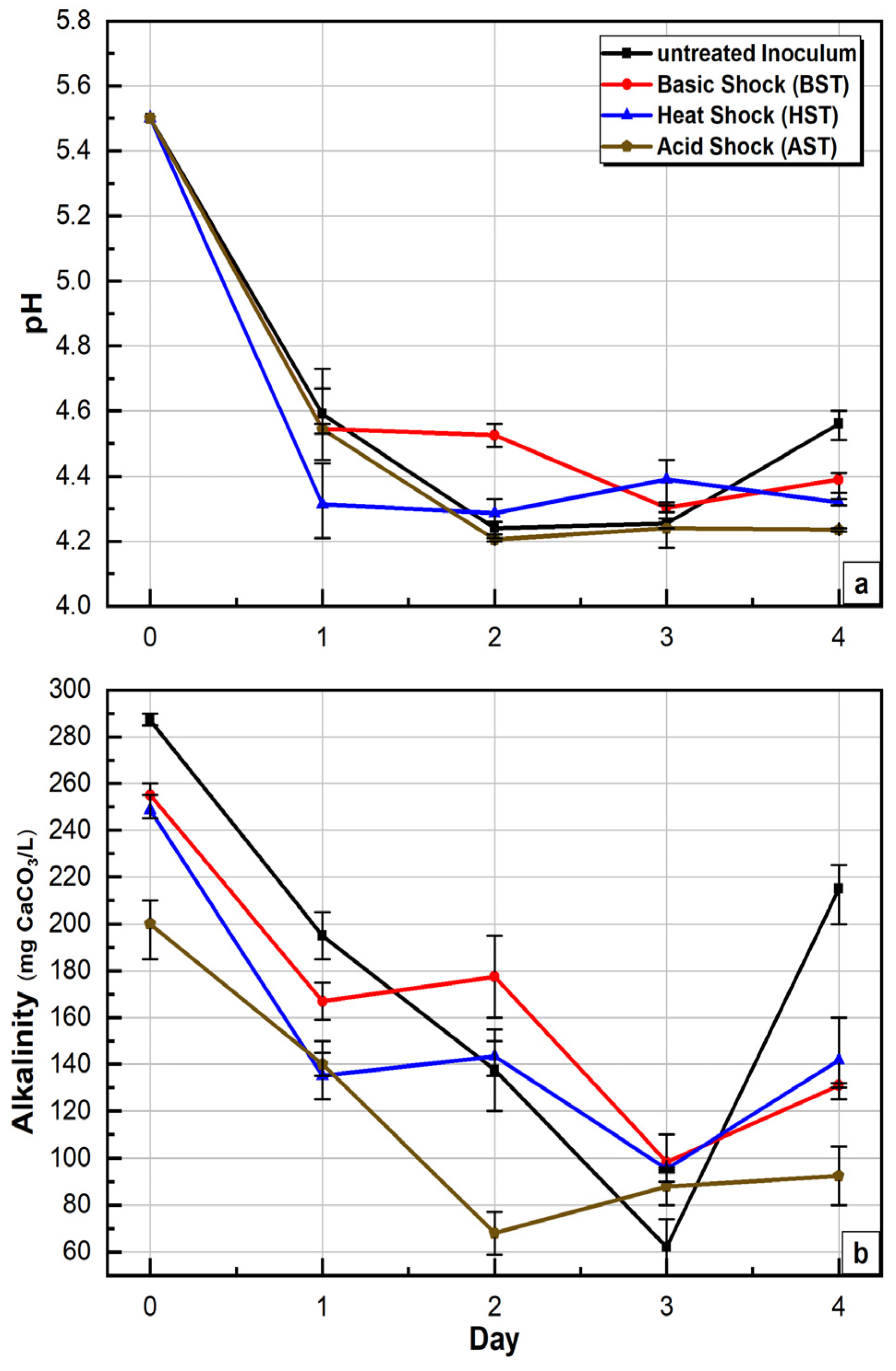
3.3. pH and Alkalinity
3.4. Process Kinetics
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Graboski, M.S.; McCormick, R.L. Combustion of fat and vegetable oil derived fuels in diesel engines. Prog. Energy Combust. Sci. 1998, 24, 125–164. [Google Scholar] [CrossRef]
- Zajic, J.; Kosaric, N.; Brosseau, J. Microbial production of hydrogen. In Advances in Biochemical Engineering; Springer: Berlin/Heidelberg, Germany, 1978; Volume 9, pp. 57–109. [Google Scholar]
- Łukajtis, R.; Hołowacz, I.; Kucharska, K.; Glinka, M.; Rybarczyk, P.; Przyjazny, A.; Kamiński, M. Hydrogen production from biomass using dark fermentation. Renew. Sustain. Energy Rev. 2018, 91, 665–694. [Google Scholar] [CrossRef]
- Momirlan, M.; Veziroglu, T. Current status of hydrogen energy. Renew. Sustain. Energy Rev. 2002, 6, 141–179. [Google Scholar] [CrossRef]
- Kapdan, I.K.; Kargi, F. Bio-hydrogen production from waste materials. Enzym. Microb. Technol. 2006, 38, 569–582. [Google Scholar] [CrossRef]
- Logan, B.E. Peer Reviewed: Extracting Hydrogen and Electricity from Renewable Resources. Environ. Sci. Technol. 2004, 38, 160A–167A. [Google Scholar] [CrossRef] [PubMed]
- Bertuccioli, L.; Chan, A.; Hart, D.; Lehner, F.; Madden, B.; Standen, E. Study on Development of Water Electrolysis in the EU. Fuel Cells Hydrogen Joint Undertakings: Lausanne, Switzerland, 2014; pp. 1–160. [Google Scholar]
- Ghimire, A.; Frunzo, L.; Pontoni, L.; D’Antonio, G.; Lens, P.N.L.; Esposito, G.; Pirozzi, F. Dark fermentation of complex waste biomass for biohydrogen production by pretreated thermophilic anaerobic digestate. J. Environ. Manag. 2015, 152, 43–48. [Google Scholar] [CrossRef]
- Nath, K.; Muthukumar, M.; Kumar, A.; Das, D. Kinetics of two-stage fermentation process for the production of hydrogen. Int. J. Hydrogen Energy 2008, 33, 1195–1203. [Google Scholar] [CrossRef]
- Antonopoulou, G.; Ntaikou, I.; Stamatelatou, K.; Lyberatos, G. Biological and Fermentative Production of Hydrogen. In Handbook of Biofuels Production; Woodhead Publishing: Cambridge, UK, 2011; pp. 305–346. [Google Scholar] [CrossRef]
- Wang, J.; Wan, W. Factors influencing fermentative hydrogen production: A review. Int. J. Hydrogen Energy 2009, 34, 799–811. [Google Scholar] [CrossRef]
- Guo, X.M.; Trably, E.; Latrille, E.; Carrère, H.; Steyer, J.-P. Hydrogen production from agricultural waste by dark fermentation: A review. Int. J. Hydrogen Energy 2010, 35, 10660–10673. [Google Scholar] [CrossRef]
- De Gioannis, G.; Muntoni, A.; Polettini, A.; Pomi, R. A review of dark fermentative hydrogen production from biodegradable municipal waste fractions. Waste Manag. 2013, 33, 1345–1361. [Google Scholar] [CrossRef]
- Kim, S.-H.; Han, S.-K.; Shin, H.-S. Feasibility of biohydrogen production by anaerobic co-digestion of food waste and sewage sludge. Int. J. Hydrogen Energy 2004, 29, 1607–1616. [Google Scholar] [CrossRef]
- Li, C.; Fang, H.H.P. Fermentative Hydrogen Production from Wastewater and Solid Wastes by Mixed Cultures. Crit. Rev. Environ. Sci. Technol. 2007, 37, 1–39. [Google Scholar] [CrossRef]
- Oh, S.-E.; Van Ginkel, S.; Logan, B.E. The Relative Effectiveness of pH Control and Heat Treatment for Enhancing Biohydrogen Gas Production. Environ. Sci. Technol. 2003, 37, 5186–5190. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Liu, J.; Wei, Y. Enhanced Biohydrogen Production from Sewage Sludge with Alkaline Pretreatment. Environ. Sci. Technol. 2004, 38, 3195–3202. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Chen, S. Pretreatment of methanogenic granules for immobilized hydrogen fermentation. Int. J. Hydrogen Energy 2007, 32, 3266–3273. [Google Scholar] [CrossRef]
- Zhu, H.; Béland, M. Evaluation of alternative methods of preparing hydrogen producing seeds from digested wastewater sludge. Int. J. Hydrogen Energy 2006, 31, 1980–1988. [Google Scholar] [CrossRef]
- Mu, Y.; Yu, H.-Q.; Wang, G. Evaluation of three methods for enriching H2-producing cultures from anaerobic sludge. Enzym. Microb. Technol. 2007, 40, 947–953. [Google Scholar] [CrossRef]
- Mohan, S.V.; Babu, V.L.; Sarma, P. Effect of various pretreatment methods on anaerobic mixed microflora to enhance biohydrogen production utilizing dairy wastewater as substrate. Bioresour. Technol. 2008, 99, 59–67. [Google Scholar] [CrossRef]
- Cheong, D.-Y.; Hansen, C.L. Bacterial stress enrichment enhances anaerobic hydrogen production in cattle manure sludge. Appl. Microbiol. Biotechnol. 2006, 72, 635–643. [Google Scholar] [CrossRef]
- Chaganti, S.R.; Kim, D.-H.; Lalman, J.A. Dark fermentative hydrogen production by mixed anaerobic cultures: Effect of inoculum treatment methods on hydrogen yield. Renew. Energy 2012, 48, 117–121. [Google Scholar] [CrossRef]
- Cai, J.; Wang, G.; Li, Y.; Zhu, D.; Pan, G. Enrichment and hydrogen production by marine anaerobic hydrogen-producing microflora. Chin. Sci. Bull. 2009, 54, 2656–2661. [Google Scholar] [CrossRef]
- Liu, H.; Wang, G.; Zhu, D.; Pan, G. Enrichment of the hydrogen-producing microbial community from marine intertidal sludge by different pretreatment methods. Int. J. Hydrogen Energy 2009, 34, 9696–9701. [Google Scholar] [CrossRef]
- Pendyala, B.; Chaganti, S.R.; Lalman, J.A.; Shanmugam, S.R.; Heath, D.D.; Lau, P.C. Pretreating mixed anaerobic communities from different sources: Correlating the hydrogen yield with hydrogenase activity and microbial diversity. Int. J. Hydrogen Energy 2012, 37, 12175–12186. [Google Scholar] [CrossRef]
- Argun, H.; Kargi, F. Effects of sludge pre-treatment method on bio-hydrogen production by dark fermentation of waste ground wheat. Int. J. Hydrogen Energy 2009, 34, 8543–8548. [Google Scholar] [CrossRef]
- Wang, J.; Wan, W. Comparison of different pretreatment methods for enriching hydrogen-producing bacteria from digested sludge. Int. J. Hydrogen Energy 2008, 33, 2934–2941. [Google Scholar] [CrossRef]
- Okoro-Shekwaga, C.K. Improving the Biomethane Yield and Biogas Quality of Food Waste during Anaerobic Digestion by Sequential Process Optimisation and Biomethanation; University of Leeds: Leeds, UK, 2019. [Google Scholar]
- Darcy, M. Water Heating Calculator. 2022. Available online: https://www.omnicalculator.com/physics/water-heating (accessed on 24 March 2022).
- Molloy, P. Run on Less with Hydrogen Fuel Cells. 2019. Available online: https://www.act-news.com/news/fcevs-run-on-less/ (accessed on 24 March 2022).
- Luo, G.; Karakashev, D.; Xie, L.; Zhou, Q.; Angelidaki, I. Long-term effect of inoculum pretreatment on fermentative hydrogen production by repeated batch cultivations: Homoacetogenesis and methanogenesis as competitors to hydrogen production. Biotechnol. Bioeng. 2011, 108, 1816–1827. [Google Scholar] [CrossRef]
- Chen, C.-C.; Lin, C.-Y.; Lin, M.-C. Acid–base enrichment enhances anaerobic hydrogen production process. Appl. Microbiol. Biotechnol. 2002, 58, 224–228. [Google Scholar] [CrossRef]
- Akutsu, Y.; Li, Y.-Y.; Harada, H.; Yu, H.-Q. Effects of temperature and substrate concentration on biological hydrogen production from starch. Int. J. Hydrogen Energy 2009, 34, 2558–2566. [Google Scholar] [CrossRef]
- Zhao, Y.; Liang, X.; Mu, H.; Zhang, X. Biohydrogen Production from Sewage Sludge by Sequential Dark and Photo Fermentation. J. Biobased Mater. Bioenergy 2015, 9, 95–100. [Google Scholar] [CrossRef]
- Valdez-Vazquez, I.; Ponce-Noyola, M.T.; Poggi-Varaldo, H.M. Nutrients related to spore germination improve H2 production from heat-shock-treated consortia. Int. J. Hydrogen Energy 2009, 34, 4291–4295. [Google Scholar] [CrossRef]
- Patel, S.K.; Kumar, P.; Kalia, V.C. Enhancing biological hydrogen production through complementary microbial metabolisms. Int. J. Hydrogen Energy 2012, 37, 10590–10603. [Google Scholar] [CrossRef]
- Hawkes, F.R.; Hussy, I.; Kyazze, G.; Dinsdale, R.; Hawkes, D.L. Continuous dark fermentative hydrogen production by mesophilic microflora: Principles and progress. Int. J. Hydrogen Energy 2007, 32, 172–184. [Google Scholar] [CrossRef]
- Pakarinen, O.; Lehtomäki, A.; Rintala, J. Batch dark fermentative hydrogen production from grass silage: The effect of inoculum, pH, temperature and VS ratio. Int. J. Hydrogen Energy 2008, 33, 594–601. [Google Scholar] [CrossRef]
- Yang, G.; Wang, J. Fermentative hydrogen production from sewage sludge. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1219–1281. [Google Scholar] [CrossRef]
- Temudo, M.F.; Muyzer, G.; Kleerebezem, R.; van Loosdrecht, M. Diversity of microbial communities in open mixed culture fermentations: Impact of the pH and carbon source. Appl. Microbiol. Biotechnol. 2008, 80, 1121–1130. [Google Scholar] [CrossRef]
- Mtui, G.Y. Recent advances in pretreatment of lignocellulosic wastes and production of value added products. Afr. J. Biotechnol. 2009, 8, 1398–1415. [Google Scholar]
- Bina, B.; Amin, M.M.; Pourzamani, H.; Fatehizadeh, A.; Ghasemian, M.; Mahdavi, M.; Taheri, E. Biohydrogen production from alkaline wastewater: The stoichiometric reactions, modeling, and electron equivalent. Methodsx 2019, 6, 1496–1505. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, G.; Shen, J. Hydrogen production in batch culture of mixed bacteria with sucrose under different iron concentrations. Int. J. Hydrogen Energy 2005, 30, 855–860. [Google Scholar] [CrossRef]
- Pagliaccia, P.; Gallipoli, A.; Gianico, A.; Montecchio, D.; Braguglia, C. Single stage anaerobic bioconversion of food waste in mono and co-digestion with olive husks: Impact of thermal pretreatment on hydrogen and methane production. Int. J. Hydrogen Energy 2016, 41, 905–915. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; Van ’T Riet, K. Modeling of the Bacterial Growth Curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Control | Test I | Test II | Test III |
|---|---|---|---|---|---|
| Inoculum | - | Untreated digestate | Heat shock treated digestate (HST) | Acid shock treated digestate (AST) | Basic shock treated digestate (BST) |
| Substrate | - | Glucose | Glucose | Glucose | Glucose |
| pH | - | 5.5 | 5.5 | 5.5 | 5.5 |
| Alkalinity | mg CaCO3/L | 287 (±3) | 249 (±6) | 200 (±13) | 255 (±9) |
| TS | % | 1.23 (±0.09) | 1.40 (±0.10) | 1.12 (±0.28) | 1.15 (±0.11) |
| VS | % | 0.88 (±0.04) | 1.00 (±0.06) | 0.80 (±0.31) | 0.82 (±0.11) |
| Parameter | Unit | Control | Test I | Test II | Test III |
|---|---|---|---|---|---|
| Inoculum | - | Untreated digestate | Heat shock treated digestate (HST) | Acid shock treated digestate (AST) | Basic shock treated digestate (BST) |
| Substrate | - | Glucose | Glucose | Glucose | Glucose |
| Total biogas a | mL | 207.7 | 203.3 | 192.7 | 198.7 |
| H2 | mL (%) | 59.7 (28.7) | 62.4 (30.7) | 52.8 (27.4) | 54.9 (27.6) |
| CO2 | mL (%) | 70.8 (34.1) | 61.7 (30.3) | 50.2 (26.1) | 60.6 (30.5) |
| N2 | mL (%) | 77.1 (37.1) | 79.3 (39.0) | 89.7 (46.5) | 83.2 (41.9) |
| CH4 | mL (%) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) |
| O2 | mL (%) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) |
| Test conditions | Pre-Treatment | Initial pH | Hydrogen Yield (mL-H2/g Glucose Added) | Reference |
|---|---|---|---|---|
| Inoculum: Digested sewage sludge Substrate: Glucose Temperature: Mesophilic (35–37 °C) Operational conditions: Batch reactor | Untreated | 5.5 | 153 | a a |
| 7 | 157 | |||
| 7 | 66 | c | ||
| 5.5 | 171 (27.5) | This study | ||
| HST | 5.5 | 155 | a a | |
| 7 | 141 | |||
| 6.2 | 120 | b | ||
| 7 | 221 | c | ||
| 5.5 | 192 (1.4) | This study | ||
| BST | 7 | 136 | c | |
| 5.5 | 154 (4.9) | This study | ||
| AST | 5.5 | 154 | a a | |
| 7 | 145 | |||
| 7 | 99 | c | ||
| 5.5 | 157 (0.8) | This study |
| Energy Consumption (Ec) | Energy Production (Ep) | Net Energy (Ep–Ec) |
|---|---|---|
| Kwh/Kg inoculum.year | Kwh/Kg substrate.year | Kwh |
| 23.7 a | 42.3 b | +18.64 |
| Pre-Treatment | Acetic Acid Concentration (mg/L) | Maximum Hydrogen Yield | Hydrogen Reduction (%) at Day 4 | ||
|---|---|---|---|---|---|
| Day 3 | Day 4 | Increment a | (mL-H2/g Glucose Added) | ||
| Untreated | 189 (38.2) | 348 (43.9) | 159 | 171 (27.5) | 38% |
| HST | 222 (51.6) | 251 (7.1) | 29 | 192 (1.4) | 8% |
| AST | 178 (15.5) | 157 (17.4) | -21 | 154 (0.9) | 6% |
| BST | 155 (22.7) | 246 (83.7) | 91 | 157 (4.9) | 13% |
| Conversion Pathway Type | Pre-Treatment | Theoretical Hydrogen Yield Potential (BHPth) | Experimental Hydrogen Yield Potential (BHPexp) | (BHPexp/BHPth) |
|---|---|---|---|---|
| Mole-H2/Mole Glucose Added | % | |||
| Butyrate | Untreated | 2.00 | 1.37 (0.22) | 68.5 |
| HST | 1.54 (0.01) | 77.0 | ||
| AST | 1.26 (0.01) | 63.0 | ||
| BST | 1.24 (0.00) | 62.0 | ||
| Pre-Treatment | P | Rm | R2 | |
|---|---|---|---|---|
| (mL) | (mL/h) | (h) | ||
| untreated | 178.8 | 4.31 | 19.54 | 0.997 |
| HST | 197.2 | 4.90 | 6.64 | 0.999 |
| AST | 195.5 | 2.60 | 4.25 | 0.987 |
| BST | 159.8 | 3.53 | 5.16 | 0.998 |
| Pre-Treatment | Correlation (Hydrogen-Acetate) | Correlation (Hydrogen-Butyrate) | n |
|---|---|---|---|
| Untreated | 0.594 * | 0.939 ** | 14 |
| HST | 0.936 ** | 0.996 ** | 15 |
| AST | 0.821 ** | 0.991 ** | 13 |
| BST | 0.417 | 0.973 ** | 13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Haddad, S.; Okoro-Shekwaga, C.K.; Fletcher, L.; Ross, A.; Camargo-Valero, M.A. Assessing Different Inoculum Treatments for Improved Production of Hydrogen through Dark Fermentation. Energies 2023, 16, 1233. https://doi.org/10.3390/en16031233
Al-Haddad S, Okoro-Shekwaga CK, Fletcher L, Ross A, Camargo-Valero MA. Assessing Different Inoculum Treatments for Improved Production of Hydrogen through Dark Fermentation. Energies. 2023; 16(3):1233. https://doi.org/10.3390/en16031233
Chicago/Turabian StyleAl-Haddad, Saleh, Cynthia Kusin Okoro-Shekwaga, Louise Fletcher, Andrew Ross, and Miller Alonso Camargo-Valero. 2023. "Assessing Different Inoculum Treatments for Improved Production of Hydrogen through Dark Fermentation" Energies 16, no. 3: 1233. https://doi.org/10.3390/en16031233
APA StyleAl-Haddad, S., Okoro-Shekwaga, C. K., Fletcher, L., Ross, A., & Camargo-Valero, M. A. (2023). Assessing Different Inoculum Treatments for Improved Production of Hydrogen through Dark Fermentation. Energies, 16(3), 1233. https://doi.org/10.3390/en16031233