
The Soil Respiration of Coal Mine Heaps’ Novel Ecosystems in Relation to Biomass and Biotic Parameters





Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Vegetation Sample Collection
2.3. Vegetation Diversity Analysis
2.4. Soil Sample Collection
2.5. Soil Biotic Parameters Analysis
Soil Fauna Samples
2.6. Enzyme Activity Measurement
2.7. Biomass Samples (Cover) Collection
2.8. Respiration Measurement—CO2 Gas Analyzer
2.9. Data Analysis
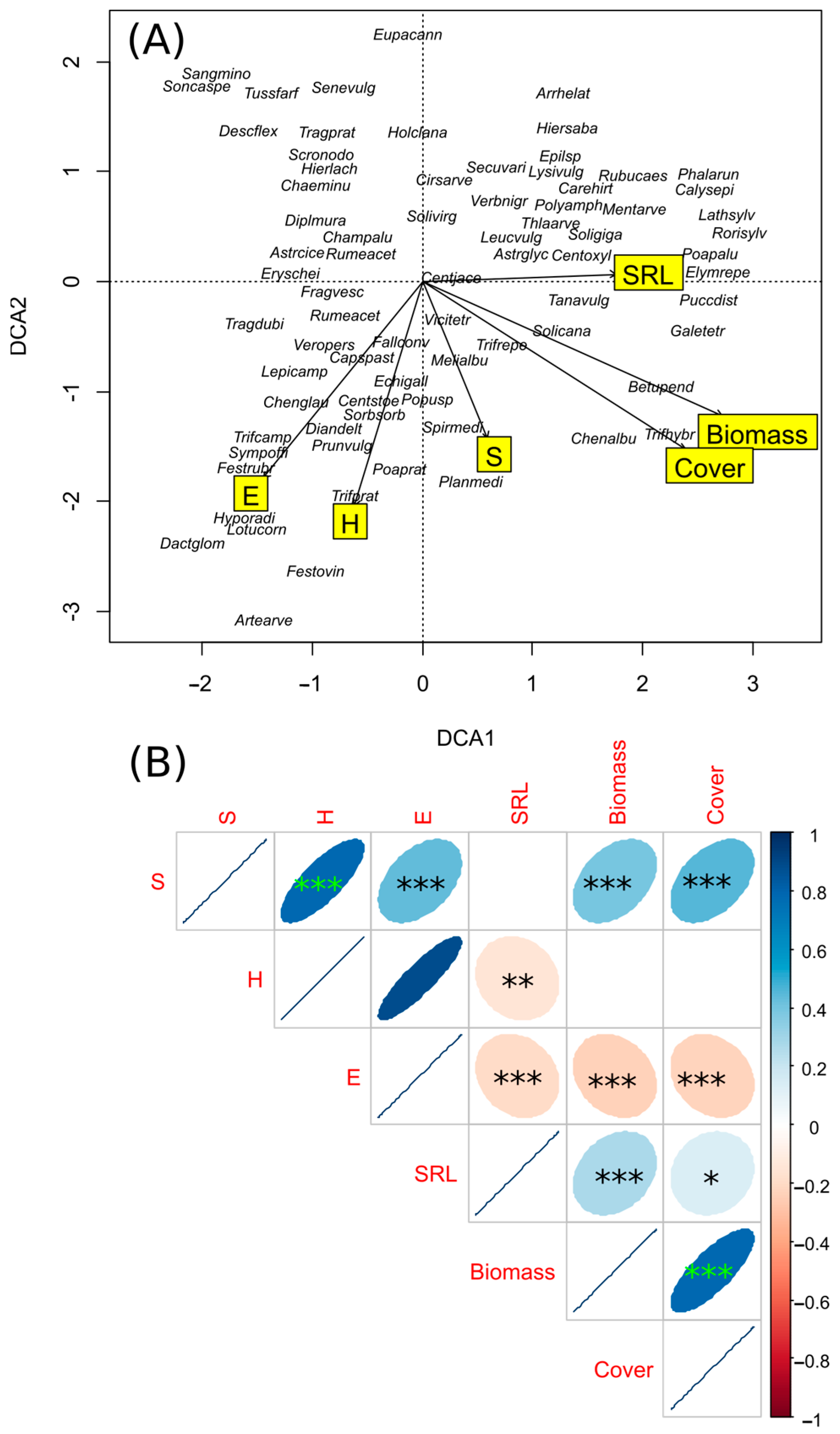
3. Results
4. Discussion
4.1. The Unusual Conditions of Novel Ecosystems
4.2. Diversity of Plant Species Composition and Soil Respiration
4.3. Soil Enzyme Activity and Respiration in the Studied Vegetation Types
4.4. Soil Organic Matter and Respiration in the Studied Vegetation Types
4.5. The Amount of Biomass and Respiration in the Studied Vegetation Types
4.6. Soil Fauna and Soil Respiration in the Studied Vegetation Types
4.7. The Environmental Novelty of the Coal Mine Heaps—Why Understanding the Novel Ecosystems Processes Is Important
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bardgett, R.D.; Van Der Putten, W.H. Belowground Biodiversity and Ecosystem Functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Błaszkowski, J.; Niezgoda, P.; Piątek, M.; Magurno, F.; Malicka, M.; Zubek, S.; Mleczko, P.; Yorou, N.S.; Jobim, K.; Vista, X.M.; et al. Rhizoglomus dalpeae, R. maiae, and R. silesianum, new species. Mycologia 2019, 111, 965–980. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.A.; Klironomos, J. Mechanisms of Plant–Soil Feedback: Interactions among Biotic and Abiotic Drivers. New Phytol. 2019, 222, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, J.; Weigelt, A.; Van Der Plas, F.; Laughlin, D.C.; Kuyper, T.W.; Guerrero-Ramirez, N.; Valverde-Barrantes, O.J.; Bruelheide, H.; Fresche, G.T.; Iversen, C.M.; et al. The Fungal Collaboration Gradient Dominates the Root Economics Space in Plants. Sci. Adv. 2020, 6, 3756. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Freeman, C.; Ostle, N.J. Microbial Contributions to Climate Change through Carbon Cycle Feedbacks. ISME J. 2008, 2, 805–814. [Google Scholar] [CrossRef]
- Allison, S.D.; Wallenstein, M.D.; Bradford, M.A. Soil-Carbon Response to Warming Dependent on Microbial Physiology. Nat. Geosci. 2010, 3, 336–340. [Google Scholar] [CrossRef]
- Woźniak, G.; Malicka, M.; Kasztowski, J.; Radosz, Ł.; Czarnecka, J.; Vangronsveld, J.; Prostański, D. How Important Are the Relations between Vegetation Diversity and Bacterial Functional Diversity for the Functioning of Novel Ecosystems? Sustainability 2023, 15, 678. [Google Scholar] [CrossRef]
- Keith, S.A.; Newton, A.C.; Herbert, R.J.H.; Morecroft, M.D.; Bealey, C.E. Non-Analogous Community Formation in Response to Climate Change. J. Nat. Conserv. 2009, 17, 228–235. [Google Scholar] [CrossRef]
- Woźniak, G. Diversity of Vegetation on Coal-Mine Heaps of the Upper Silesia; Polish Academy of Sciences: Poland, Kraków, 2010. [Google Scholar]
- Błońska, E.; Lasota, J.; Tullus, A.; Lutter, R.; Ostonen, I. Impact of Deadwood Decomposition on Soil Organic Carbon Sequestration in Estonian and Polish Forests. Ann. Sci. 2019, 76, 102. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Higgs, E.S.; Hall, C.M. Novel Ecosystems: Intervening in the New Ecological World Order; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 1–368. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Reich, P.B. Forest Productivity Increases with Evenness, Species Richness and Trait Variation: A Global Meta-Analysis. J. Ecol. 2012, 100, 742–749. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.Y.H. Global Effects of Plant Litter Alterations on Soil CO2 to the Atmosphere. Glob. Chang. Biol. 2018, 24, 3462–3471. [Google Scholar] [CrossRef] [PubMed]
- Chapman, S.K.; Newman, G.S. Biodiversity at the Plant-Soil Interface: Microbial Abundance and Community Structure Respond to Litter Mixing. Oecologia 2010, 162, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Eisenhauer, N.; Beßler, H.; Engels, C.; Gleixner, G.; Habekost, M.; Milcu, A.; Partsch, S.; Sabais, A.C.W.; Scherber, C.; Steinbeiss, S.; et al. Plant Diversity Effects on Soil Microorganisms Support the Singular Hypothesis. Ecology 2010, 91, 485–496. [Google Scholar] [CrossRef]
- Handa, I.T.; Aerts, R.; Berendse, F.; Berg, M.P.; Bruder, A.; Butenschoen, O.; Chauvet, E.; Gessner, M.O.; Jabiol, J.; Makkonen, M.; et al. Consequences of Biodiversity Loss for Litter Decomposition across Biomes. Nature 2014, 509, 218–221. [Google Scholar] [CrossRef]
- Sinsabaugh, R.; Antibus, R.K. An Enzymic Approach to the Analysis of Microbial Activity during Plant Litter Decomposition Drought-Induced Piñon Mortality View Project Climate Change Effects on Soil Invertebrate Communities View Project. Agric. Ecosyst. Environ. 1991, 34, 43–54. [Google Scholar] [CrossRef]
- Bradford, M.A.; Watts, B.W.; Davies, C.A. Thermal Adaptation of Heterotrophic Soil Respiration in Laboratory Microcosms. Glob. Chang. Biol. 2010, 16, 1576–1588. [Google Scholar] [CrossRef]
- Smulczak, L.; Tracz, H. Metabolizm Oddechowy Gleb w Roznych Wariantach Sposobu Przygotowania Gleby Oraz Udzialu Domieszek Na Zalesionych Gruntach Porolnych. Sylwan 2008, 152, 63–71. [Google Scholar]
- Kasprzak, K.; Państwowe Wydawnictwo Naukowe (1951–1992); Polska Akademia Nauk; Instytut Zoologii. Ènhitreidy (Enchytraeidae); Enchytraeids (Enchytraeidae, Oligochaeta). Fragm. Faun. 1981, 26, 65–76. [Google Scholar] [CrossRef]
- Buchmann, N. Biotic and Abiotic Factors Controlling Soil Respiration Rates in Picea Abies Stands. Soil Biol. Biochem. 2000, 32, 1625–1635. [Google Scholar] [CrossRef]
- Schinner, F. Introduction. In Methods Soil Biology; Springer: Berlin/Heidelberg, Germany, 1996; pp. 3–6. [Google Scholar] [CrossRef]
- Burmeier, H. Bioremediation of Soil. In Methods in Applied Soil Microbiology and Biochemistry; Academic Press: Cambridge, MA, USA, 1995; ISBN 9780125138406. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving Indicator Species Analysis by Combining Groups of Sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- Schimel, D.S.; Emanuel, W.; Rizzo, B.; Smith, T.; Woodward, F.I.; Fisher, H.; Kittel, T.G.F.; Mckeown, R.; Painter, T.; Rosenbloom, N.; et al. Continental Scale Variability in Ecosystem Processes: Models, Data, and the Role of Disturbance. Ecol. Monogr. 1997, 67, 251–271. [Google Scholar] [CrossRef]
- Kałucka, I.L.; Jagodziński, A.M. Successional Traits of Ectomycorrhizal Fungi in Forest Reclamation after Surface Mining and Agricultural Disturbances: A Review Environmental and Genetic Factors Affecting Productivity of Forest Ecosystems on Forest and Post-Industrial Habitats View Project. Dendrobiology 2016, 76, 91–104. [Google Scholar] [CrossRef]
- Woźniak, G.; Sierka, E.; Wheeler, A. Urban and Industrial Habitats: How Important They Are for Ecosystem Services. Ecosyst. Serv. Glob. Ecol. 2018, 19, 75723. [Google Scholar] [CrossRef]
- Prach, K.; Walker, L.R. Comparative Plant Succession among Terrestrial Biomes of the World; Cambridge University Press: Cambridge, UK, 2019; p. 400. [Google Scholar]
- Tropek, R.; Kadlec, T.; Hejda, M.; Kocarek, P.; Skuhrovec, J.; Malenovsky, I.; Vodka, S.; Spitzer, L.; Banar, P.; Konvicka, M. Technical Reclamations Are Wasting the Conservation Potential of Post-Mining Sites. A Case Study of Black Coal Spoil Dumps. Ecol. Eng. 2012, 43, 13–18. [Google Scholar] [CrossRef]
- Frouz, J.; Jílková, V. The Effect of Ants on Soil Properties and Processes (Hymenoptera: Formicidae). Myrmecol. News 2008, 11, 191–199. [Google Scholar]
- Řehounková, K.; Čížek, L.; Řehounek, J.; Šebelíková, L.; Tropek, R.; Lencová, K.; Bogusch, P.; Marhoul, P.; Máca, J. Additional Disturbances as a Beneficial Tool for Restoration of Post-Mining Sites: A Multi-Taxa Approach. Environ. Sci. Pollut. Res. 2016, 23, 13745–13753. [Google Scholar] [CrossRef]
- Rotherham, I.D. Recombinant Ecology—A Hybrid Future? Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar] [CrossRef]
- Zhang, M.; Sayer, E.J.; Zhang, W.; Ye, J.; Yuan, Z.; Lin, F.; Hao, Z.; Fang, S.; Mao, Z.; Ren, J.; et al. Seasonal Influence of Biodiversity on Soil Respiration in a Temperate Forest. Plants 2022, 11, 3391. [Google Scholar] [CrossRef]
- Vargas, R.; Allen, M.F. Environmental Controls and the Influence of Vegetation Type, Fine Roots and Rhizomorphs on Diel and Seasonal Variation in Soil Respiration. New Phytol. 2008, 179, 460–471. [Google Scholar] [CrossRef]
- Woźniak, G.; Chmura, D.; Małkowski, E.; Zieleźnik-Rusinowska, P.; Sitko, K.; Ziemer, B.; Błońska, A. Is the Age of Novel Ecosystem the Factor Driving Arbuscular Mycorrhizal Colonization in Poa compressa and Calamagrostis epigejos? Plants 2021, 10, 949. [Google Scholar] [CrossRef] [PubMed]
- He, X.Y.; Wang, K.L.; Zhang, W.; Chen, Z.H.; Zhu, Y.G.; Chen, H.S. Positive correlation between soil bacterial metabolic and plant species diversity and bacterial and fungal diversity in a vegetation succession on Karst. Plant Soil 2008, 307, 123–134. [Google Scholar] [CrossRef]
- Jiang, L.; Pu, Z.; Nemergut, D.R. On the Importance of the Negative Selection Effect for the Relationship between Biodiversity and Ecosystem Functioning. Oikos 2008, 117, 488–493. [Google Scholar] [CrossRef]
- Frouz, J.; Špaldoňová, A.; Fričová, K.; Bartuška, M. The Effect of Earthworms (Lumbricus rubellus) and Simulated Tillage on Soil Organic Carbon in a Long-Term Microcosm Experiment. Soil Biol. Biochem. 2014, 78, 58–64. [Google Scholar] [CrossRef]
- Błońska, A.; Chmura, D.; Hutniczak, A.; Wilczek, Z.; Jarosz, J.; Besenyei, L.; Woźniak, G. The Plant Species Composition of an Abandoned Meadow as an Element of an Ecosystem Mosaic within an Urban-Industrial Landscape. Sustainability 2022, 14, 11851. [Google Scholar] [CrossRef]
- Chmura, D.; Jagodziński, A.M.; Hutniczak, A.; Dyczko, A.; Woźniak, G. Novel Ecosystems in the Urban-Industrial Landscape–Interesting Aspects of Environmental Knowledge Requiring Broadening: A Review. Sustainability 2022, 14, 10829. [Google Scholar] [CrossRef]
- Khlifa, R.; Paquette, A.; Messier, C.; Reich, P.B.; Munson, A.D. Do Temperate Tree Species Diversity and Identity Influence Soil Microbial Community Function and Composition? Ecol. Evol. 2017, 7, 7965–7974. [Google Scholar] [CrossRef]
- Mestre, L.; Toro-Manríquez, M.; Soler, R.; Huertas-Herrera, A.; Martínez-Pastur, G.; Lencinas, M.V. The Influence of Canopy-Layer Composition on Understory Plant Diversity in Southern Temperate Forests. For. Ecosyst. 2017, 4, 6. [Google Scholar] [CrossRef]
- Stell, E.; Warner, D.; Jian, J.; Bond-Lamberty, B.; Vargas, R. Spatial Biases of Information Influence Global Estimates of Soil Respiration: How can we Improve Global Predictions? Glob. Chang. Biol. 2021, 27, 3923–3938. [Google Scholar] [CrossRef]
- Murphy, M.; Balser, T.; Buchmann, N.; Hahn, V.; Potvin, C. Linking Tree Biodiversity to Belowground Process in a Young Tropical Plantation: Impacts on Soil CO2 Flux. Ecol. Manag. 2008, 255, 2577–2588. [Google Scholar] [CrossRef]
- Allen, B.; Willner, D.; Oechel, W.C.; Lipson, D. Top-down Control of Microbial Activity and Biomass in an Arctic Soil Ecosystem. Environ. Microbiol. 2010, 12, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Waldrop, M.P.; Firestone, M.K. Seasonal Dynamics of Microbial Community Composition and Function in Oak Canopy and Open Grassland Soils. Microb. Ecol. 2006, 52, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Giasson, M.A.; Ellison, A.M.; Bowden, R.D.; Crill, P.M.; Davidson, E.A.; Drake, J.E.; Frey, S.D.; Hadley, J.L.; Lavine, M.; Melillo, J.M.; et al. Soil Respiration in a Northeastern US Temperate Forest: A 22-Year Synthesis. Ecosphere 2013, 4, 1–28. [Google Scholar] [CrossRef]
- Estiarte, M.; Peñuelas, J. Alteration of the Phenology of Leaf Senescence and Fall in Winter Deciduous Species by Climate Change: Effects on Nutrient Proficiency. Glob. Chang. Biol. 2015, 21, 1005–1017. [Google Scholar] [CrossRef]
- Kittredge, H.A.; Cannone, T.; Funk, J.; Chapman, S.K. Soil Respiration and Extracellular Enzyme Production Respond Differently across Seasons to Elevated Temperatures. Plant Soil 2018, 425, 351–361. [Google Scholar] [CrossRef]
- Fierer, N.; Craine, J.M.; Mclauchlan, K.; Schimel, J.P. Litter Quality and the Temperature Sensitivity of Decomposition. Ecology 2005, 86, 320–326. [Google Scholar] [CrossRef]
- Wallenstein, M.; Allison, S.D.; Ernakovich, J.; Steinweg, J.M.; Sinsabaugh, R. Controls on the Temperature Sensitivity of Soil Enzymes: A Key Driver of In Situ Enzyme Activity Rates. Soil Enzymol. 2010, 22, 245–258. [Google Scholar] [CrossRef]
- German, D.P.; Marcelo, K.R.B.; Stone, M.M.; Allison, S.D. The Michaelis–Menten Kinetics of Soil Extracellular Enzymes in Response to Temperature: A Cross-Latitudinal Study. Glob. Chang. Biol. 2012, 18, 1468–1479. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil Enzymes in a Changing Environment: Current Knowledge and Future Directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Hartley, I.P.; Heinemeyer, A.; Ineson, P. Effects of Three Years of Soil Warming and Shading on the Rate of Soil Respiration: Substrate Availability and Not Thermal Acclimation Mediates Observed Response. Glob. Chang. Biol. 2007, 13, 1761–1770. [Google Scholar] [CrossRef]
- Dorrepaal, E.; Toet, S.; Van Logtestijn, R.S.P.; Swart, E.; Van De Weg, M.J.; Callaghan, T.V.; Aerts, R. Carbon Respiration from Subsurface Peat Accelerated by Climate Warming in the Subarctic. Nature 2009, 460, 616–619. [Google Scholar] [CrossRef]
- Frey, S.D.; Lee, J.; Melillo, J.M.; Six, J. The Temperature Response of Soil Microbial Efficiency and Its Feedback to Climate. Nat. Clim. Chang. 2013, 3, 395–398. [Google Scholar] [CrossRef]
- Melillo, J.M.; Frey, S.D.; DeAngelis, K.M.; Werner, W.J.; Bernard, M.J.; Bowles, F.P.; Pold, G.; Knorr, M.A.; Grandy, A.S. Long-Term Pattern and Magnitude of Soil Carbon Feedback to the Climate System in a Warming World. Science 2017, 358, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kaur, S.; Parkash Choudhary, O.; Singh, M.; Al-Huqail, A.A.; Ali, H.M.; Kumar, R.; Siddiqui, M.H. Tillage, Green Manure and Residue Retention Improves Aggregate-Associated Phosphorus Fractions under Rice–Wheat Cropping. Sci. Rep. 2022, 12, 11106. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature Sensitivity of Soil Carbon Decomposition and Feedbacks to Climate Change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.A.; Davies, C.A.; Frey, S.D.; Maddox, T.R.; Melillo, J.M.; Mohan, J.E.; Reynolds, J.F.; Treseder, K.K.; Wallenstein, M.D. Thermal Adaptation of Soil Microbial Respiration to Elevated Temperature. Ecol. Lett. 2008, 11, 1316–1327. [Google Scholar] [CrossRef]
- Allison, S.D.; Vitousek, P.M. Responses of Extracellular Enzymes to Simple and Complex Nutrient Inputs. Soil Biol. Biochem. 2005, 37, 937–944. [Google Scholar] [CrossRef]
- Allison, S.D.; Chacon, S.S.; German, D.P. Substrate Concentration Constraints on Microbial Decomposition. Soil Biol. Biochem. 2014, 79, 43–49. [Google Scholar] [CrossRef]
- Karhu, K.; Auffret, M.D.; Dungait, J.A.J.; Hopkins, D.W.; Prosser, J.I.; Singh, B.K.; Subke, J.A.; Wookey, P.A.; Agren, G.I.; Sebastià, M.T.; et al. Temperature Sensitivity of Soil Respiration Rates Enhanced by Microbial Community Response. Nature 2014, 513, 81–84. [Google Scholar] [CrossRef]
- Bölscher, T.; Paterson, E.; Freitag, T.; Thornton, B.; Herrmann, A.M. Temperature Sensitivity of Substrate-Use Efficiency Can Result from Altered Microbial Physiology without Change to Community Composition. Soil Biol. Biochem. 2017, 109, 59–69. [Google Scholar] [CrossRef]
- Wu, L.; Yang, Y.; Wang, S.; Yue, H.; Lin, Q.; Hu, Y.; He, Z.; Van Nostrand, J.D.; Hale, L.; Li, X.; et al. Alpine Soil Carbon Is Vulnerable to Rapid Microbial Decomposition under Climate Cooling. ISME J. 2017, 11, 2102–2111. [Google Scholar] [CrossRef]
- DeAngelis, K.M.; Pold, G.; Topçuoglu, B.D.; van Diepen, L.T.A.; Varney, R.M.; Blanchard, J.L.; Melillo, J.; Frey, S.D. Long-Term Forest Soil Warming Alters Microbial Communities in Temperate Forest Soils. Front. Microbiol. 2015, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Chodak, M.; Niklińska, M. The Effect of Different Tree Species on the Chemical and Microbial Properties of Reclaimed Mine Soils. Biol. Fertil. Soils 2010, 46, 555–566. [Google Scholar] [CrossRef]
- Ciarkowska, K.; Sołek-Podwika, K.; Wieczorek, J. Enzyme Activity as an Indicator of Soil-Rehabilitation Processes at a Zinc and Lead Ore Mining and Processing Area. J. Environ. Manag. 2014, 132, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P.; Valášková, V. Degradation of Cellulose by Basidiomycetous Fungi. FEMS Microbiol. Rev. 2008, 32, 501–521. [Google Scholar] [CrossRef]
- Kompała-Bąba, A.; Bierza, W.; Sierka, E.; Błońska, A.; Besenyei, L.; Woźniak, G. The Role of Plants and Soil Properties in the Enzyme Activities of Substrates on Hard Coal Mine Spoil Heaps. Sci. Rep. 2021, 11, 5155. [Google Scholar] [CrossRef]
- Rodríguez-Loinaz, G.; Onaindia, M.; Amezaga, I.; Mijangos, I.; Garbisu, C. Relationship between Vegetation Diversity and Soil Functional Diversity in Native Mixed-Oak Forests. Soil Biol. Biochem. 2008, 40, 49–60. [Google Scholar] [CrossRef]
- Šantrůčková, H.; Vrba, J.; Picek, T.; Kopáček, J. Soil Biochemical Activity and Phosphorus Transformations and Losses from Acidified Forest Soils. Soil Biol. Biochem. 2004, 36, 1569–1576. [Google Scholar] [CrossRef]
- Abakumov, E.V.; Cajthaml, T.; Brus, J.; Frouz, J. Humus Accumulation, Humification, and Humic Acid Composition in Soils of Two Post-Mining Chronosequences after Coal Mining. J. Soils Sediments 2013, 13, 491–500. [Google Scholar] [CrossRef]
- Markowicz, A.; Woźniak, G.; Borymski, S.; Piotrowska-Seget, Z.; Chmura, D. Links in the Functional Diversity between Soil Microorganisms and Plant Communities during Natural Succession in Coal Mine Spoil Heaps. Ecol. Res. 2015, 30, 1005–1014. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Kapusta, P.; Błońska, A.; Kompała-Baba, A.; Woźniak, G. Effects of Calamagrostis Epigejos, Chamaenerion Palustre and Tussilago Farfara on Nutrient Availability and Microbial Activity in the Surface Layer of Spoil Heaps after Hard Coal Mining. Ecol. Eng. 2015, 83, 328–337. [Google Scholar] [CrossRef]
- Wozniak, G.; Markowicz, A.; Borymski, S.; Piotrowska-Seget, Z.; Chmura, D.; Besenyei, L. The Relationship between Successional Vascular Plant Assemblages and Associated Microbial Communities on Coal Mine Spoil Heaps. Community Ecol. 2015, 16, 23–32. [Google Scholar] [CrossRef]
- Acosta-Martínez, V.; Cruz, L.; Sotomayor-Ramírez, D.; Pérez-Alegría, L. Enzyme activities as affected by soil properties and land use in a tropical watershed. Appl. Soil Ecol. 2007, 35, 35–45. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; Mcmahon, S.K.; Schimel, J.P. Seasonal Variation in Enzyme Activities and Temperature Sensitivities in Arctic Tundra Soils. Glob. Chang. Biol. 2009, 15, 1631–1639. [Google Scholar] [CrossRef]
- Baldrian, P.; Šnajdr, J.; Merhautová, V.; Dobiášová, P.; Cajthaml, T.; Valášková, V. Responses of the Extracellular Enzyme Activities in Hardwood Forest to Soil Temperature and Seasonality and the Potential Effects of Climate Change. Soil Biol. Biochem. 2013, 56, 60–68. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G. Effects of Exotic Plant Invasions on Soil Nutrient Cycling Processes. Ecosystems 2003, 6, 503–523. [Google Scholar] [CrossRef]
- Ashton, I.W.; Hyatt, L.A.; Howe, K.M.; Gurevitch, J.; Lerdau, M.T. Invasive Species Accelerate Decomposition and Litter Nitrogen Loss in a Mixed Deciduous Forest. Ecol. Appl. 2005, 15, 1263–1272. [Google Scholar] [CrossRef]
- Chapman, S.K.; Devine, K.A.; Curran, C.; Jones, R.O.; Gilliam, F.S. Impacts of Soil Nitrogen and Carbon Additions on Forest Understory Communities with a Long Nitrogen Deposition History. Ecosystems 2016, 19, 142–154. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Hockenberry, A.W.; Adams, M.B.; Gilliam, F.S.; Hockenberry, A.W. Effects of Atmospheric Nitrogen Deposition on the Herbaceous Layer of a Central Appalachian Hardwood Forest. J. Torrey Bot. Soc. 2006, 133, 240–254. [Google Scholar] [CrossRef]
- Milewska-Hendel, A.; Chmura, D.; Wyrwał, K.; Kurczyńska, E.U.; Kompała-Bąba, A.; Jagodziński, A.M.; Woźniak, G. Cell Wall Epitopes in Grasses of Different Novel Ecosystem Habitats on Post-Industrial Sites. Land Degrad. Dev. 2021, 32, 1680–1694. [Google Scholar] [CrossRef]
- Almeida, T.S.; Arantes, M.R.; Lopes Neto, J.J.; Souza, T.M.; Pessoa, I.P.; Medeiros, J.L.; Tabosa, P.M.S.; Moreira, T.B.; Farias, D.F.; Carvalho, A.F.U. Evaluation of Seeds Ethanolic Extracts of Triplaris Gardneriana Wedd. Using in Vitro and in Vivo Toxicological Methods. J. Toxicol. Environ. Health A 2020, 83, 135–152. [Google Scholar] [CrossRef]
- Boone, R.D.; Nadelhoffer, K.J.; Canary, J.D.; Kaye, J.P. Roots Exert a Strong Influence on the Temperature Sensitivity of Soil Respiration. Nature 1998, 396, 570–572. [Google Scholar] [CrossRef]
- Gansert, D. Root Respiration and Its Importance for the Carbon Balance of Beech Saplings (Fagus sylvatica L.) in a Montane Beech Forest. Plant Soil 1994, 167, 109–119. [Google Scholar] [CrossRef]
- Burton, A.J.; Pregitzer, K.S.; Zogg, G.P.; Zak, D.R. Drought Reduces Root Respiration in Sugar Maple Forests. Ecol. Appl. 1998, 8, 771. [Google Scholar] [CrossRef]
- Högberg, P.; Ekblad, A. Substrate-Induced Respiration Measured in Situ in a C3-Plant Ecosystem Using Additions of C4-Sucrose. Soil Biol. Biochem. 1996, 28, 1131–1138. [Google Scholar] [CrossRef]
- Nakane, K.; Kohno, T.; Horikoshi, T. Root Respiration Rate before and Just after Clear-Felling in a Mature, Deciduous, Broad-Leaved Forest. Ecol. Res. 1996, 11, 111–119. [Google Scholar] [CrossRef]
- Bowden, R.D.; Nadelhoffer, K.J.; Boone, R.D.; Melillo, J.M.; Garrison, J.B. Contributions of Aboveground Litter, Belowground Litter, and Root Respiration to Total Soil Respiration in a Temperate Mixed Hardwood Forest. Can. J. For. Res. 2011, 23, 1402–1407. [Google Scholar] [CrossRef]
- Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. Separating Root and Soil Microbial Contributions to Soil Respiration: A Review of Methods and Observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Chapin, F.S.; Woodwell, G.M.; Randerson, J.T.; Rastetter, E.B.; Lovett, G.M.; Baldocchi, D.D.; Clark, D.A.; Harmon, M.E.; Schimel, D.S.; Valentini, R.; et al. Reconciling Carbon-Cycle Concepts, Terminology, and Methods. Ecosystems 2006, 9, 1041–1050. [Google Scholar] [CrossRef]
- Collalti, A.; Tjoelker, M.G.; Hoch, G.; Mäkelä, A.; Guidolotti, G.; Heskel, M.; Petit, G.; Ryan, M.G.; Battipaglia, G.; Matteucci, G.; et al. Plant Respiration: Controlled by Photosynthesis or Biomass? Glob. Chang. Biol. 2020, 26, 1739–1753. [Google Scholar] [CrossRef]
- West, G.B.; Brown, J.H.; Enquist, B.J. A General Model for the Structure and Allometry of Plant Vascular Systems. Nature 1999, 400, 664–667. [Google Scholar] [CrossRef]
- Reich, P.B.; Tjoelker, M.G.; Machado, J.L.; Oleksyn, J. Universal Scaling of Respiratory Metabolism, Size and Nitrogen in Plants. Nature 2006, 439, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Tjoelker, M.G.; Pregitzer, K.S.; Wright, I.J.; Oleksyn, J.; Machado, J.L. Scaling of Respiration to Nitrogen in Leaves, Stems and Roots of Higher Land Plants. Ecol. Lett. 2008, 11, 793–801. [Google Scholar] [CrossRef]
- Kira, T.; Shidei, T. Primary Production and Turnover of Organic Matter in Different Forest Ecosystems of the Western Pacific. Jpn. J. Ecol. 1967, 17, 70–87. [Google Scholar] [CrossRef]
- Odum, E.P. The Strategy of Ecosystem Development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef]
- O’Connor, M.P.; Kemp, S.J.; Agosta, S.J.; Hansen, F.; Sieg, A.E.; Wallace, B.P.; McNair, J.N.; Dunham, A.E. Reconsidering the Mechanistic Basis of the Metabolic Theory of Ecology. Oikos 2007, 116, 1058–1072. [Google Scholar] [CrossRef]
- Raich, J.W.; Schlesinger, W.H. The Global Carbon Dioxide Flux in Soil Respiration and Its Relationship to Vegetation and Climate. Tellus B 1992, 44, 81–99. [Google Scholar] [CrossRef]
- Lavigne, M.B.; Ryan, M.G.; Anderson, D.E.; Baldocchi, D.D.; Crill, P.M.; Fitzjarrald, D.R.; Goulden, M.L.; Gower, S.T.; Massheder, J.M.; McCaughey, J.H.; et al. Comparing Nocturnal Eddy Covariance Measurements to Estimates of Ecosystem Respiration Made by Scaling Chamber Measurements at Six Coniferous Boreal Sites. J. Geophys. Res. Atmos. 1997, 102, 28977–28985. [Google Scholar] [CrossRef]
- Lindroth, A.; Grelle, A.; Morén, A.S. Long-term Measurements of Boreal Forest Carbon Balance Reveal Large Temperature Sensitivity. Glob. Chang. Biol. 1998, 4, 443–450. [Google Scholar] [CrossRef]
- Raich, J.W.; Potter, C.S. Global Patterns of Carbon Dioxide Emissions from Soils. Glob. Biogeochem. Cycles 1995, 9, 23–36. [Google Scholar] [CrossRef]
- García-Palacios, P.; Maestre, F.T.; Kattge, J.; Wall, D.H. Climate and Litter Quality Differently Modulate the Effects of Soil Fauna on Litter Decomposition across Biomes. Ecol. Lett. 2013, 16, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Soong, J.L.; Vandegehuchte, M.L.; Horton, A.J.; Nielsen, U.N.; Denef, K.; Shaw, E.A.; de Tomasel, C.M.; Parton, W.; Wall, D.H.; Cotrufo, M.F. Soil Microarthropods Support Ecosystem Productivity and Soil C Accrual: Evidence from a Litter Decomposition Study in the Tallgrass Prairie. Soil Biol. Biochem. 2016, 92, 230–238. [Google Scholar] [CrossRef]
- Wall, D.H.; Bradford, M.A.; St. John, M.G.; Trofymow, J.A.; Behan-Pelletier, V.; Bignell, D.E.; Dangerfield, J.M.; Parton, W.J.; Rusek, J.; Voigt, W.; et al. Global Decomposition Experiment Shows Soil Animal Impacts on Decomposition Are Climate-Dependent. Glob. Chang. Biol. 2008, 14, 2661–2677. [Google Scholar] [CrossRef]
- Nielsen, U.N.; Ayres, E.; Wall, D.H.; Li, G.; Bardgett, R.D.; Wu, T.; Garey, J.R. Global-Scale Patterns of Assemblage Structure of Soil Nematodes in Relation to Climate and Ecosystem Properties. Glob. Ecol. Biogeogr. 2014, 23, 968–978. [Google Scholar] [CrossRef]
- Frouz, J. Effects of Soil Macro- and Mesofauna on Litter Decomposition and Soil Organic Matter Stabilization. Geoderma 2018, 332, 161–172. [Google Scholar] [CrossRef]
- Hicks Pries, C.E.; Castanha, C.; Porras, R.C.; Torn, M.S. The Whole-Soil Carbon Flux in Response to Warming. Science 2017, 355, 1420–1423. [Google Scholar] [CrossRef]
- Lavelle, P. Diversity of Soil Fauna and Ecosystem Function. Biol. Int. 1996, 33, 16. [Google Scholar]
- Heděnec, P.; Zheng, H.; Siqueira, D.P.; Peng, Y.; Schmidt, I.K.; Frøslev, T.G.; Kjøller, R.; Li, H.; Frouz, J.; Vesterdal, L. Litter Chemistry of Common European Tree Species Drives the Feeding Preference and Consumption Rate of Soil Invertebrates, and Shapes the Diversity and Structure of Gut and Faecal Microbiomes. Soil Biol. Biochem. 2022, 177, 108918. [Google Scholar] [CrossRef]
- Norman, J.M.; Kucharik, C.J.; Gower, S.T.; Baldocchi, D.D.; Crill, P.M.; Rayment, M.; Savage, K.; Striegl, R.G. A Comparison of Six Methods for Measuring Soil-Surface Carbon Dioxide Fluxes. J. Geophys. Res. Atmos. 1997, 102, 28771–28777. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Laskowski, M.J.; Burton, A.J.; Lessard, V.C.; Zak, D.R. Variation in Sugar Maple Root Respiration with Root Diameter and Soil Depth. Tree Physiol. 1998, 18, 665–670. [Google Scholar] [CrossRef]
- Morse, N.; Mcdowell, W.H.; Morse, N.B.; Pellissier, P.A.; Cianciola, E.N.; Brereton, R.L.; Sullivan, M.M.; Shonka, N.K.; Wheeler, T.B.; Mcdowell, W.H.; et al. Novel Ecosystems in the Anthropocene: A Revision of the Novel Ecosystem Concept for Pragmatic Applications. Artic. Ecol. Soc. 2014, 19, 10. [Google Scholar] [CrossRef]
- Evers, C.R.; Wardropper, C.B.; Branoff, B.; Granek, E.F.; Hirsch, S.L.; Link, T.E.; Olivero-Lora, S.; Wilson, C. The Ecosystem Services and Biodiversity of Novel Ecosystems: A Literature Review. Glob. Ecol. Conserv. 2018, 13, 362. [Google Scholar] [CrossRef]
- Golik, V.I.; Klyuev, R.V.; Martyushev, N.V.; Zyukin, D.A.; Karlina, A.I. Technology for Nonwaste Recovery of Tailings of the Mizur Mining and Processing Plant. Metallurgist 2023, 66, 1476–1480. [Google Scholar] [CrossRef]
- Ren, H.; Xiao, W.; Zhao, Y. Examining the effect of spontaneous combustion on vegetation restoration at coal waste dumps after reclamation: Taking Medicago sativa L. (Alfalfa) as an indicator. Sci. Total Environ. 2023, 901, 165668. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | IndVal Value | p-Value |
|---|---|---|
| Group 2 Phragmites australis | 0.994 | 0.001 |
| Bidens frondosa | 0.378 | 0.001 |
| Lathyrus sylvestris | 0.378 | 0.002 |
| Puccinellia distans | 0.365 | 0.006 |
| Plantago major | 0.363 | 0.006 |
| Phalaris arundinacea | 0.339 | 0.009 |
| Urtica dioica | 0.33 | 0.011 |
| Chelidonium majus | 0.267 | 0.041 |
| Eleocharis palustris | 0.267 | 0.033 |
| Polygonum hydropiper | 0.267 | 0.033 |
| Rorippa sylvestris | 0.267 | 0.033 |
| Group 3 Tussilago farfara | 0.969 | 0.001 |
| Group 4 Hieracium pilosella | 0.35 | 0.03 |
| Group 5 Solidago gigantea | 0.901 | 0.001 |
| Tanacetum vulgare | 0.661 | 0.001 |
| Hypericum perforatum | 0.637 | 0.001 |
| Melilotus albus | 0.512 | 0.015 |
| Vicia hirsuta | 0.48 | 0.001 |
| Carex spicata | 0.471 | 0.001 |
| Carex hirta | 0.457 | 0.002 |
| Deschampsia caespitosa | 0.437 | 0.002 |
| Astragalus glycyphyllos | 0.426 | 0.003 |
| Melandrium album | 0.399 | 0.002 |
| Lathyrus pratensis | 0.333 | 0.005 |
| Linaria vulgaris | 0.329 | 0.011 |
| Rosa canina | 0.316 | 0.012 |
| Leucanthemum vulgare | 0.252 | 0.047 |
| Group | Stat | p-Value |
|---|---|---|
| Group | 1 + 4 | |
| Arenaria serpyllifolia | 0.509 | 0.004 |
| Group | 2 + 5 | |
| Poa palustris | 0.515 | 0.001 |
| Agrostis gigantea | 0.483 | 0.001 |
| Elymus repens | 0.423 | 0.001 |
| Group | 4 + 5 | |
| Calamagrostis epigejos | 0.923 | 0.001 |
| Erigeron annuus | 0.574 | 0.003 |
| Achillea millefolium | 0.564 | 0.001 |
| Cirsium arvense | 0.509 | 0.023 |
| Vicia tetrasperma | 0.458 | 0.006 |
| Centaurea jacea | 0.287 | 0.036 |
| Group | 1 + 2 + 4 | |
| Lotus corniculatus | 0.529 | 0.023 |
| Group | 1 + 3 + 4 | |
| Chamaenerion palustre | 0.619 | 0.008 |
| Echium vulgare | 0.585 | 0.021 |
| Group | 1 + 4 + 5 | |
| Poa compressa | 0.659 | 0.001 |
| Centaurea stoebe | 0.603 | 0.015 |
| Group | 2 + 3 + 4 | |
| Pinus sylvestris | 0.425 | 0.036 |
| Group | 2 + 4 + 5 | |
| Epilobium sp. | 0.356 | 0.024 |
| Group | 1 + 2 + 3 + 4 | |
| Daucus carota | 0.772 | 0.002 |
| Hieracium piloselloides | 0.728 | 0.001 |
| Group | 1 + 2 + 4 + 5 | |
| Medicago lupulina | 0.607 | 0.003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radosz, Ł.; Chmura, D.; Prostański, D.; Woźniak, G. The Soil Respiration of Coal Mine Heaps’ Novel Ecosystems in Relation to Biomass and Biotic Parameters. Energies 2023, 16, 7083. https://doi.org/10.3390/en16207083
Radosz Ł, Chmura D, Prostański D, Woźniak G. The Soil Respiration of Coal Mine Heaps’ Novel Ecosystems in Relation to Biomass and Biotic Parameters. Energies. 2023; 16(20):7083. https://doi.org/10.3390/en16207083
Chicago/Turabian StyleRadosz, Łukasz, Damian Chmura, Dariusz Prostański, and Gabriela Woźniak. 2023. "The Soil Respiration of Coal Mine Heaps’ Novel Ecosystems in Relation to Biomass and Biotic Parameters" Energies 16, no. 20: 7083. https://doi.org/10.3390/en16207083
APA StyleRadosz, Ł., Chmura, D., Prostański, D., & Woźniak, G. (2023). The Soil Respiration of Coal Mine Heaps’ Novel Ecosystems in Relation to Biomass and Biotic Parameters. Energies, 16(20), 7083. https://doi.org/10.3390/en16207083