
Development of Stable Mixed Microbiota for High Yield Power to Methane Conversion

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Fermentation System
2.2. Volatile Organic Acid Analysis
2.3. Gas Composition Analysis
- η = conversion efficiency of H2 to CH4 (%)
- = CH4 production of reactor A (mL·L−1·d−1)
- CH4 production of control reactor (mL·L−1·d−1)
- injected amount of H2 to reactor A (mL·L−1·d−1)
- residual amount of H2 in reactor A (mL·L−1·d−1)
2.4. Determination of Fermentation Parameters
2.5. Total DNA Isolation for Metagenomics
2.6. Shotgun Sequencing
2.7. Raw Sequence Filtering
2.8. Read-Based Metagenome Data Processing and Statistical Analysis
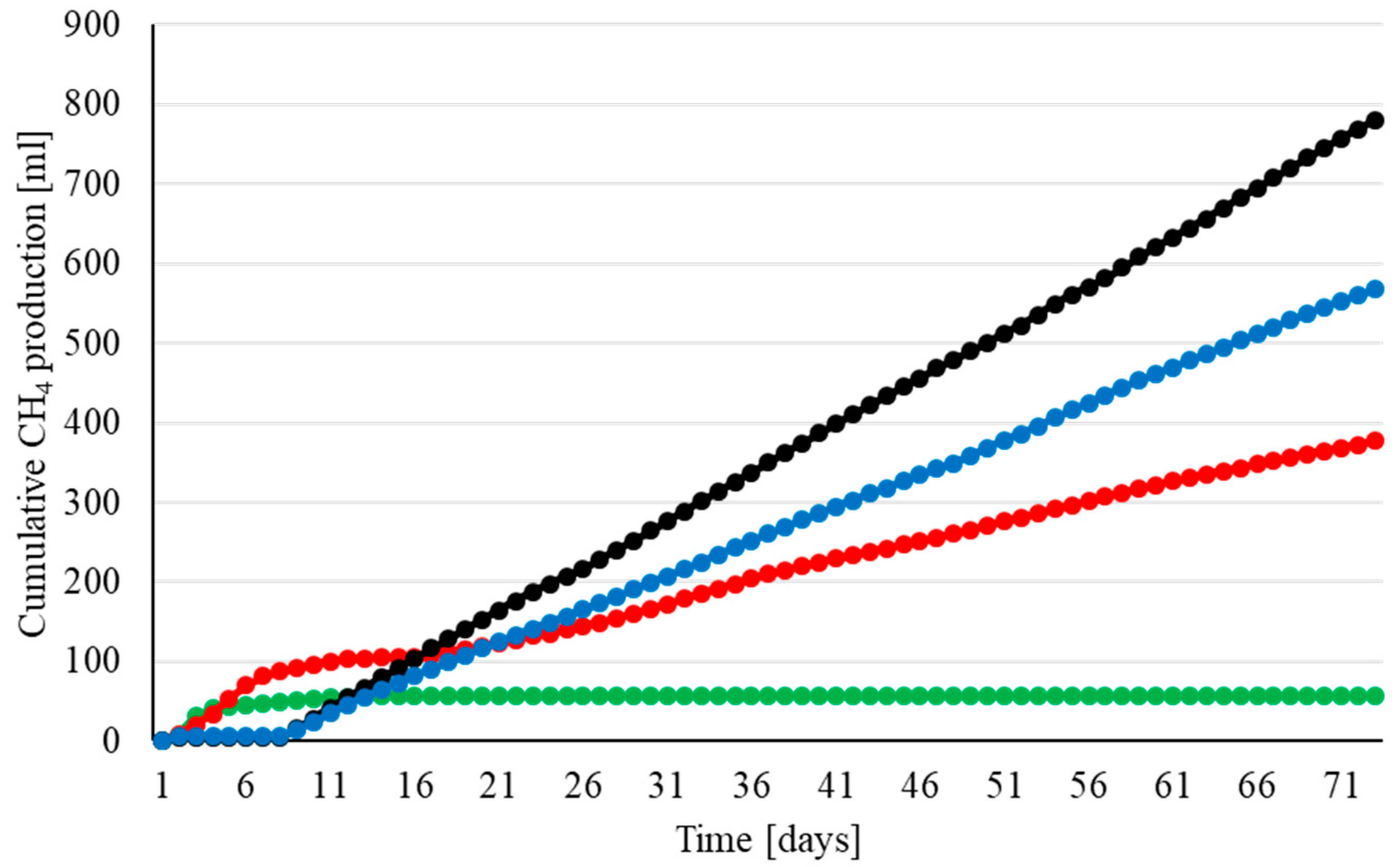
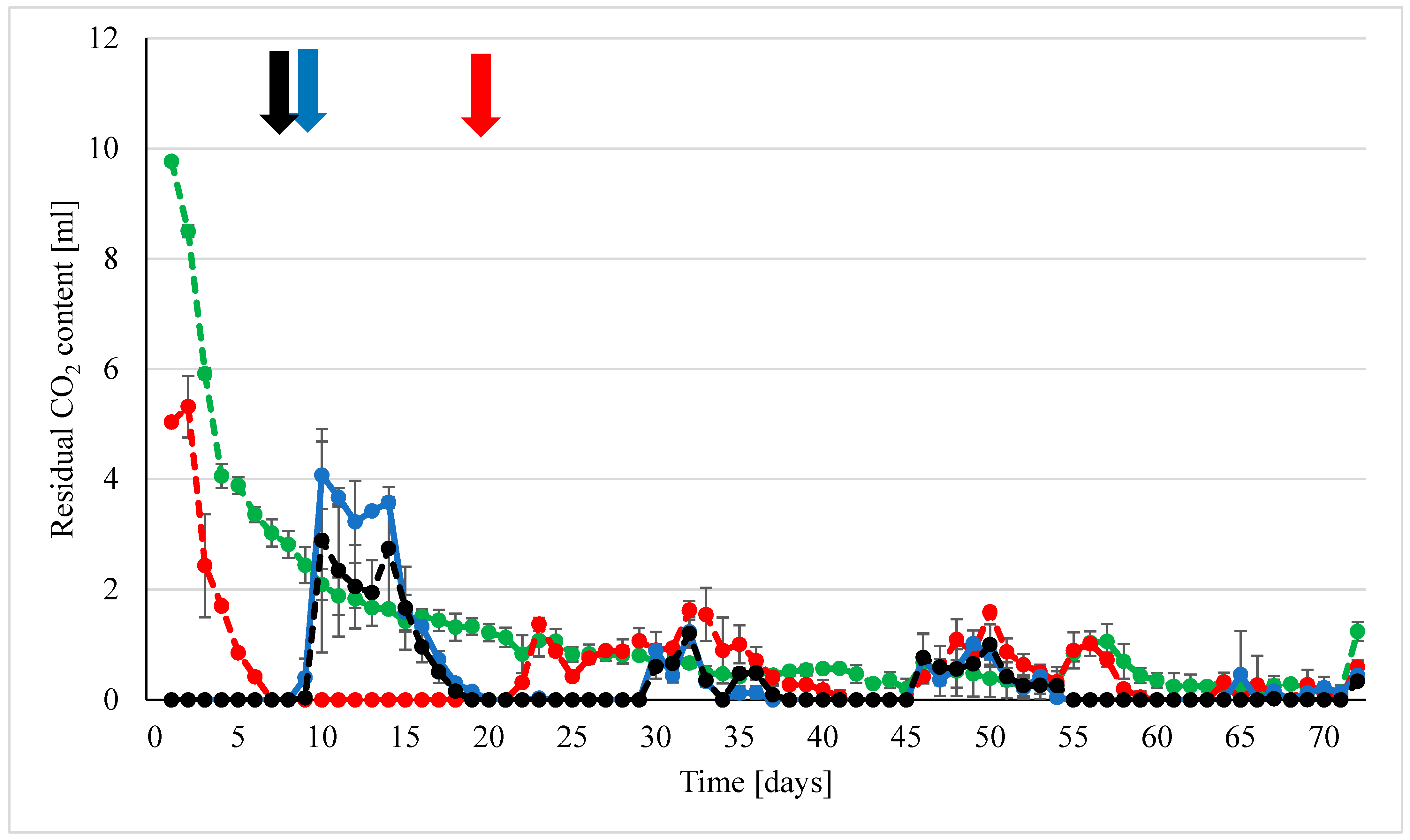
3. Results
3.1. Methanogenesis by H2 and H2 + CO2
3.2. Metagenomic Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- REN21. Renewables 2020 Global Status Report. 2020. Available online: https://www.ren21.net/reports/global-status-report/?gclid=CjwKCAjwoP6LBhBlEiwAvCcthM9n4BM6oHikcrY_nWbzC811LiRCEiGVVkKEgzPT2NcaGYXiUiOBkhoCLR4QAvD_BwE (accessed on 1 November 2021).
- Fawzy, S.; Osman, A.I.; Doran, J.; Rooney, D.W. Strategies for mitigation of climate change: A review. Environ. Chem. Lett. 2020, 18, 2069–2094. [Google Scholar] [CrossRef]
- Østergaard, P.A. Comparing electricity, heat and biogas storages’ impacts on renewable energy integration. Energy 2012, 37, 255–262. [Google Scholar] [CrossRef]
- Lund, P.D.; Lindgren, J.; Mikkola, J.; Salpakari, J. Review of energy system flexibility measures to enable high levels of variable renewable electricity. Renew. Sustain. Energy Rev. 2015, 45, 785–807. [Google Scholar] [CrossRef]
- Ketter, W.; Collins, J.; Saar-Tsechansky, M.; Marom, O. Information Systems for a Smart Electricity Grid. ACM Trans. Manag. Inf. Syst. 2018, 9, 1–22. [Google Scholar] [CrossRef]
- Holladay, J.D.; Hu, J.; King, D.L.; Wang, Y. An overview of hydrogen production technologies. Catal. Today 2009, 139, 244–260. [Google Scholar] [CrossRef]
- Thema, M.; Bauer, F.; Sterner, M. Power-to-Gas: Electrolysis and methanation status review. Renew. Sustain. Energy Rev. 2019, 112, 775–787. [Google Scholar] [CrossRef]
- Sharma, S.; Ghoshal, S.K. Hydrogen the future transportation fuel: From production to applications. Renew. Sustain. Energy Rev. 2015, 43, 1151–1158. [Google Scholar] [CrossRef]
- Herzog, A.; Tatsutani, M. A hydrogen future ? An Economic and Environmental Assessment of Hydrogen Production Pathways. Nat. Resour. Def. Counc. 2005, 23. [Google Scholar]
- Viswanathan, B. Hydrogen Storage. Energy Sources 2017, 185–212. [Google Scholar] [CrossRef]
- Andrei, H.; Badea, C.A.; Andrei, P.; Spertino, F. Energetic-Environmental-Economic Feasibility and Impact. Energies 2020, 14, 100. [Google Scholar] [CrossRef]
- Campana, P.E.; Mainardis, M.; Moretti, A.; Cottes, M. 100% renewable wastewater treatment plants: Techno-economic assessment using a modelling and optimization approach. Energy Convers. Manag. 2021, 239, 114214. [Google Scholar] [CrossRef]
- Luo, G.; Johansson, S.; Boe, K.; Xie, L.; Zhou, Q.; Angelidaki, I. Simultaneous hydrogen utilization and in situ biogas upgrading in an anaerobic reactor. Biotechnol. Bioeng. 2012, 109, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Global Methane Initiative. Global Methane Emissions and Mitigation Opportunities; Global Methane Initiative: Washington, DC, USA, 2010; Volume 2020, pp. 1–4. Available online: https://www.globalmethane.org/documents/gmi-mitigation-factsheet.pdf (accessed on 1 November 2021).
- Angelidaki, I.; Treu, L.; Tsapekos, P.; Luo, G.; Campanaro, S.; Wenzel, H.; Kougias, P.G. Biogas upgrading and utilization: Current status and perspectives. Biotechnol. Adv. 2018, 36, 452–466. [Google Scholar] [CrossRef]
- Piechota, G. Biogas/Biomethane Quality and Requirements for Combined Heat and Power (CHP) Units/Gas Grids with a Special Focus on Siloxanes-a Short Review. Sustain. Chem. Eng. 2022, 3, 1–10. [Google Scholar]
- Piechota, G. Multi-step biogas quality improving by adsorptive packed column system as application to biomethane upgrading. J. Environ. Chem. Eng. 2021, 9, 105944. [Google Scholar] [CrossRef]
- Angelidaki, I.; Karakashev, D.; Batstone, D.J.; Plugge, C.M.; Stams, A.J.M. Biomethanation and Its Potential, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2011. [Google Scholar] [CrossRef]
- Götz, M.; Lefebvre, J.; Mörs, F.; Koch, A.M.; Graf, F.; Bajohr, S.; Reimert, R.; Kolb, T. Renewable Power-to-Gas: A technological and economic review. Renew. Energy 2016, 85, 1371–1390. [Google Scholar] [CrossRef]
- Rafrafi, Y.; Laguillaumie, L.; Dumas, C. Biological Methanation of H2 and CO2 with Mixed Cultures: Current Advances, Hurdles and Challenges. Waste Biomass Valorization 2020, 12, 5259–5282. [Google Scholar] [CrossRef]
- Wahid, R.; Horn, S.J. Impact of operational conditions on methane yield and microbial community composition during biological methanation in in situ and hybrid reactor systems. Biotechnol. Biofuels 2021, 14, 170. [Google Scholar] [CrossRef]
- Adnan, A.I.; Ong, M.Y.; Nomanbhay, S.; Chew, K.W. Technologies for Biogas Upgrading to Biomethane: A Review. Bioengineering 2019, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Rusmanis, D.; O’Shea, R.; Wall, D.M.; Murphy, J.D. Biological hydrogen methanation systems–an overview of design and efficiency. Bioengineered 2019, 10, 604–634. [Google Scholar] [CrossRef]
- Rittmann, S.; Seifert, A.; Herwig, C. Essential prerequisites for successful bioprocess development of biological CH4 production from CO2 and H2. Crit. Rev. Biotechnol. 2015, 35, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Peillex, J.P.; Fardeau, M.L.; Boussand, R.; Navarro, J.M.; Belaich, J.P. Growth of Methanococcus thermolithotrophicus in batch and continuous culture on H2 and CO2: Influence of agitation. Appl. Microbiol. Biotechnol. 1988, 29, 560–564. [Google Scholar] [CrossRef]
- de Poorter, L.M.I.; Geerts, W.J.; Keltjens, J.T. Coupling of Methanothermobacter thermautotrophicus methane formation and growth in fed-batch and continuous cultures under different H2 gassing regimens. Appl. Environ. Microbiol. 2007, 73, 740–749. [Google Scholar] [CrossRef]
- Strevett, K.A.; Vieth, R.F.; Grasso, D. Chemo-autotrophic biogas purification for methane enrichment: Mechanism and kinetics. Chem. Eng. J. Biochem. Eng. J. 1995, 58, 71–79. [Google Scholar] [CrossRef]
- Szuhaj, M.; Ács, N.; Tengölics, R.; Bodor, A.; Rákhely, G.; Kovács, K.L.; Bagi, Z. Conversion of H2and CO2to CH4and acetate in fed-batch biogas reactors by mixed biogas community: A novel route for the power-to-gas concept. Biotechnol. Biofuels 2016, 9, 102. [Google Scholar] [CrossRef]
- Bassani, I.; Kougias, P.G.; Treu, L.; Angelidaki, I. Biogas Upgrading via Hydrogenotrophic Methanogenesis in Two-Stage Continuous Stirred Tank Reactors at Mesophilic and Thermophilic Conditions. Environ. Sci. Technol. 2015, 49, 12585–12593. [Google Scholar] [CrossRef]
- Bassani, I.; Kougias, P.G.; Treu, L.; Porté, H.; Campanaro, S.; Angelidaki, I. Optimization of hydrogen dispersion in thermophilic up-flow reactors for ex situ biogas upgrading. Bioresour. Technol. 2017, 234, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Agneessens, L.M.; Ottosen, L.D.M.; Voigt, N.V.; Nielsen, J.L.; de Jonge, N.; Fischer, C.H.; Kofoed, M.V.W. In-situ biogas upgrading with pulse H2additions: The relevance of methanogen adaption and inorganic carbon level. Bioresour. Technol. 2017, 233, 256–263. [Google Scholar] [CrossRef]
- Luo, G.; Angelidaki, I. Integrated biogas upgrading and hydrogen utilization in an anaerobic reactor containing enriched hydrogenotrophic methanogenic culture. Biotechnol. Bioeng. 2012, 109, 2729–2736. [Google Scholar] [CrossRef]
- Strübing, D.; Huber, B.; Lebuhn, M.; Drewes, J.E. High performance biological methanation in thermophilic anaerobic trickle bed reactors. Bioresour. Technol. 2017, 245, 1176–1183. [Google Scholar] [CrossRef] [PubMed]
- Rachbauer, L.; Beyer, R.; Bochmann, G.; Fuchs, W. Characteristics of adapted hydrogenotrophic community during biomethanation. Sci. Total Environ. 2017, 595, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Ács, N.; Szuhaj, M.; Wirth, R.; Bagi, Z.; Maróti, G.; Rákhely, G.; Kovács, K.L. Microbial Community Rearrangements in Power-to-Biomethane Reactors Employing Mesophilic Biogas Digestate. Front. Energy Res. 2019, 7, 132. [Google Scholar] [CrossRef]
- Bassani, I.; Kougias, P.G.; Angelidaki, I. In-situ biogas upgrading in thermophilic granular UASB reactor: Key factors affecting the hydrogen mass transfer rate. Bioresour. Technol. 2016, 221, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Afgan, E.; Baker, D.; van den Beek, M.; Blankenberg, D.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Eberhard, C.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2016 update. Nucleic Acids Res. 2016, 44, W3–W10. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef]
- Mende, D.R.; Letunic, I.; Huerta-Cepas, J.; Li, S.S.; Forslund, K.; Sunagawa, S.; Bork, P. ProGenomes: A resource for consistent functional and taxonomic annotations of prokaryotic genomes. Nucleic Acids Res. 2017, 45, D529–D534. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef]
- Siriwongrungson, V.; Zeng, R.J.; Angelidaki, I. Homoacetogenesis as the alternative pathway for H2 sink during thermophilic anaerobic degradation of butyrate under suppressed methanogenesis. Water Res. 2007, 41, 4204–4210. [Google Scholar] [CrossRef]
- Bouanane-Darenfed, A.; Hania, W.B.; Cayol, J.L.; Ollivier, B.; Fardeau, M.L. Reclassification of acetomicrobium faecale as caldicoprobacter faecalis comb. Nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 3286–3288. [Google Scholar] [CrossRef]
- Avdiyuk, K.V. Keratinolytic Enzymes: Producers, Physical and Chemical Properties. Application for Biotechnology. Biotechnol. Acta 2019, 12, 27–45. [Google Scholar] [CrossRef]
- Fortina, M.G.; Pukall, R.; Schumann, P.; Mora, D.; Parini, C.; Manachini, P.L.; Stackebrandt, E. Ureibacillus gen. nov., a new genus to accommodate Bacillus thermosphaericus (Andersson et al. 1995), emendation of Ureibacillus thermosphaericus and description of Ureibacillus terrenus sp. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 447–455. [Google Scholar] [CrossRef]
- Weon, H.Y.; Lee, S.Y.; Kim, B.Y.; Noh, H.J.; Schumann, P.; Kim, J.S.; Kwon, S.W. Ureibacillus composti sp. nov. and Ureibacillus thermophilus sp. nov., isolated from livestock-manure composts. Int. J. Syst. Evol. Microbiol. 2007, 57, 2908–2911. [Google Scholar] [CrossRef]
- Maus, I.; Koeck, D.E.; Cibis, K.G.; Hahnke, S.; Kim, Y.S.; Langer, T.; Kreubel, J.; Erhard, M.; Bremges, A.; Off, S.; et al. Unraveling the microbiome of a thermophilic biogas plant by metagenome and metatranscriptome analysis complemented by characterization of bacterial and archaeal isolates. Biotechnol. Biofuels 2016, 9, 1–28. [Google Scholar] [CrossRef]
- Shiratori, H.; Ohiwa, H.; Ikeno, H.; Ayame, S.; Kataoka, N.; Miya, A.; Beppu, T.; Ueda, K. Lutispora thermophila gen. nov., sp. nov., a thermophilic, spore-forming bacterium isolated from a thermophilic methanogenic bioreactor digesting municipal solid wastes. Int. J. Syst. Evol. Microbiol. 2008, 58, 964–969. [Google Scholar] [CrossRef]
- Logroño, W.; Popp, D.; Kleinsteuber, S.; Sträuber, H.; Harms, H.; Nikolausz, M. Microbial resource management for ex situ biomethanation of hydrogen at alkaline ph. Microorganisms 2020, 8, 614. [Google Scholar] [CrossRef] [PubMed]
- Wasserfallen, A.; Nölling, J.; Pfister, P.; Reeve, J.; de Macario, E.C. Phylogenetic analysis of 18 thermophilic Methanobacterium isolates supports the proposals to create a new genus, Methanothermobacter gen. nov., and to reclassify several isolates in three species, Methanothermobacter thermautotrophicus comb. nov., Methano. Int. J. Syst. Evol. Microbiol. 2000, 50, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, L.; Qian, Y.; Xu, Y.; Wu, H.; Zhang, J.; Huang, H.; Chang, Z. Contributions of thermotolerant bacteria to organic matter degradation under a hyperthermophilic pretreatment process during chicken manure composting. BioResources 2019, 14, 6747–6766. [Google Scholar] [CrossRef]
- Gorliczay, E.; Boczonádi, I.; Kiss, N.É.; Tóth, F.A.; Pabar, S.A.; Biró, B.; Kovács, L.R.; Tamás, J. Microbiological effectivity evaluation of new poultry farming organic waste recycling. Agric 2021, 11, 683. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Faria, C.; Nobre, M.F.; Schumann, P.; Nunes, O.C.; Manaia, C.M. Paenibacillus humicus sp. nov., isolated from poultry litter compost. Int. J. Syst. Evol. Microbiol. 2007, 57, 2267–2271. [Google Scholar] [CrossRef][Green Version]
- McInerney, M.J.; Bryant, M.P.; Hespell, R.B.; Costerton, J.W. Syntrophomonas wolfei gen. nov. sp. nov., an anaerobic, syntrophic, fatty acid-oxidizing bacterium. Appl. Environ. Microbiol. 1981, 41, 1029–1039. [Google Scholar] [CrossRef]
- Westerholm, M.; Roos, S.; Schnürer, A. Syntrophaceticus schinkiigen. nov., sp. nov., an anaerobic, syntrophic acetate-oxidizing bacterium isolated from a mesophilic anaerobic filter. FEMS Microbiol. Lett. 2010, 309, 100–104. [Google Scholar] [CrossRef]
- United Nations. Adoption of the Paris Agreement, Proposal by the President, Draft decision. In Proceedings of the Twenty-First Session of the Conference of the Parties (COP 21), Paris, France, 11 December 2015; Volume 32, p. 21932. Available online: http://unfccc.int/resource/docs/2015/cop21/eng/l09r01.pdf (accessed on 1 November 2021).
- Csedő, Z.; Zavarkó, M.; Vaszkun, B.; Koczkás, S. Hydrogen Economy Development Opportunities by Inter-Organizational Digital Knowledge Networks. Sustainability 2021, 13, 9194. [Google Scholar] [CrossRef]
- Kakuk, B.; Wirth, R.; Maróti, G.; Szuhaj, M.; Rakhely, G.; Laczi, K.; Kovács, K.L.; Bagi, Z. Early response of methanogenic archaea to H2 as evaluated by metagenomics and metatranscriptomics. Microb. Cell Fact. 2021, 20, 127. [Google Scholar] [CrossRef] [PubMed]
- Kleinsteuber, S. Metagenomics of Methanogenic Communities in Anaerobic Digesters. In Biogenesis of Hydrocarbons Handbook of Hydrocarbon and Lipid Microbiology; Springer: Cham, Switzerland, 2019; pp. 337–359. [Google Scholar] [CrossRef]
- Corbellini, V.; Kougias, P.G.; Treu, L.; Bassani, I.; Malpei, F.; Angelidaki, I. Hybrid biogas upgrading in a two-stage thermophilic reactor. Energy Convers. Manag. 2018, 168, 1–10. [Google Scholar] [CrossRef]
- Mulat, D.G.; Mosbæk, F.; Ward, A.J.; Polag, D.; Greule, M.; Keppler, F.; Nielsen, J.L.; Feilberg, A. Exogenous addition of H2for an in situ biogas upgrading through biological reduction of carbon dioxide into methane. Waste Manag. 2017, 68, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Bagi, Z.; Ács, N.; Böjti, T.; Kakuk, B.; Rákhely, G.; Strang, O.; Szuhaj, M.; Wirth, R.; Kovács, K.L. Biomethane: The energy storage, platform chemical and greenhouse gas mitigation target. Anaerobe 2017, 46, 13–22. [Google Scholar] [CrossRef]
- Agneessens, L.M.; Ottosen, L.D.M.; Andersen, M.; Olesen, C.B.; Feilberg, A.; Kofoed, M.V.W. Parameters affecting acetate concentrations during in-situ biological hydrogen methanation. Bioresour. Technol. 2018, 258, 33–40. [Google Scholar] [CrossRef]
- Waldron, C.R.; Becker-Vallone, C.A.; Eveleigh, D.E. Isolation and characterization of a cellulolytic actinomycete Microbispora bispora. Appl. Microbiol. Biotechnol. 1986, 24, 477–486. [Google Scholar] [CrossRef]
- Li, Y.; Chen, F.; Dong, K.; Wang, G. Actinotalea ferrariae sp. nov., isolated from an iron mine, and emended description of the genus Actinotalea. Int. J. Syst. Evol. Microbiol. 2013, 63, 3398–3403. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Kandler, O. Taxonomy of the genus Cellulomonas, based on phenotypic characters and deoxyribonucleic acid-deoxyribonucleic acid homology, and proposal of seven neotype strains. Int. J. Syst. Bacteriol. 1979, 29, 273–282. [Google Scholar] [CrossRef]
- Kushkevych, I.; Cejnar, J.; Vítězová, M.; Vítěz, T.; Dordević, D.; Bomble, Y. Occurrence of thermophilic microorganisms in different full scale biogas plants. Int. J. Mol. Sci. 2020, 21, 283. [Google Scholar] [CrossRef]
- Nouioui, I.; Carro, L.; García-López, M.; Meier-Kolthoff, J.P.; Woyke, T.; Kyrpides, N.C.; Pukall, R.; Klenk, H.P.; Goodfellow, M.; Göker, M. Genome-based taxonomic classification of the phylum actinobacteria. Front. Microbiol. 2018, 9, 1–119. [Google Scholar] [CrossRef]
- Hattori, S.; Kamagata, Y.; Hanada, S. A Strictly Anaerobic, Thermophilic, Syntrophic Acetate-Oxidizing Bacterium. Int. J. Syst. Evol. Microbiol. 2000, 50, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Zakaria, Z.; Abu, J.; Aziz, S.A.; Rojas-Ponce, G. Isolation of Mycobacterium avium and other nontuberculous mycobacteria in chickens and captive birds in peninsular Malaysia. BMC Vet. Res. 2021, 17, 13. [Google Scholar] [CrossRef] [PubMed]
- Dyksma, S.; Jansen, L.; Gallert, C. Syntrophic acetate oxidation replaces acetoclastic methanogenesis during thermophilic digestion of biowaste. Microbiome 2020, 8, 105. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, J.A.; Allen, E.; Wall, D.M.; Jackson, S.A.; Murphy, J.D.; Dobson, A.D.W. Methanosarcina play an important role in anaerobic co-digestion of the seaweed ulva lactuca: Taxonomy and predicted metabolism of functional microbial communities. PLoS ONE 2015, 10, e0142603. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.C.D.; Isha, A.; Chandra, R.; Vijay, V.K.; Subbarao, P.M.V.; Kumar, R.; Chaudhary, V.P.; Singh, H.; Khan, A.A.; Tyagi, V.K.; et al. Enhancing methane production in anaerobic digestion through hydrogen assisted pathways—A state-of-the-art review. Renew. Sustain. Energy Rev. 2021, 151, 111536. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | START | Control | 20 mL H2 Reactor | 40 mL H2 Reactor | 60 mL H2 Reactor |
|---|---|---|---|---|---|
| Methanosarcina | 21.69 | 22.80 | 15.24 | 13.70 | 9.35 |
| Caldicoprobacter | 19.89 | 7.12 | 12.32 | 9.90 | 4.20 |
| Ureibacillus | 9.10 | 0.05 | 0.19 | 0.12 | 0.05 |
| Methanothermobacter | 8.37 | 6.60 | 29.47 | 37.53 | 65.26 |
| Clostridium | 4.52 | 1.48 | 1.02 | 1.01 | 0.78 |
| Lutispora | 2.28 | 0.29 | 0.17 | 0.16 | 0.13 |
| Desulfotomaculum | 1.80 | 2.18 | 1.41 | 1.30 | 0.78 |
| Moorella | 1.53 | 1.26 | 0.85 | 0.73 | 0.43 |
| Herbinix | 1.35 | 0.50 | 0.27 | 0.28 | 0.22 |
| Lysinibacillus | 1.33 | 0.10 | 0.10 | 0.10 | 0.10 |
| Limnochorda | 1.31 | 1.09 | 0.63 | 0.50 | 0.23 |
| Acetomicrobium | 1.28 | 3.11 | 1.54 | 1.32 | 0.78 |
| Bacillus | 1.23 | 0.50 | 0.35 | 0.31 | 0.16 |
| Paenibacillus | 1.16 | 0.60 | 0.40 | 0.39 | 0.23 |
| Syntrophomonas | 1.03 | 0.36 | 0.22 | 0.21 | 0.13 |
| Tepidanaerobacter | 1.01 | 0.65 | 0.34 | 0.25 | 0.17 |
| Syntrophaceticus | 0.57 | 1.48 | 1.79 | 1.55 | 0.76 |
| Sphaerobacter | 0.57 | 1.60 | 0.67 | 0.76 | 0.36 |
| Thermacetogenium | 0.55 | 1.11 | 1.24 | 1.18 | 0.51 |
| Mycobacterium | 0.37 | 1.69 | 0.80 | 0.78 | 0.55 |
| Methanobacterium | 0.28 | 19.99 | 10.99 | 9.30 | 5.72 |
| Actinotalea | 0.10 | 5.09 | 5.32 | 5.39 | 2.10 |
| Cellulomonas | 0.10 | 2.44 | 2.65 | 2.66 | 1.05 |
| Bassani et al. (2015) | Corbellini et al. (2018) | Mulat et al. (2017) | Present Work | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | H2 Added | Control | CSTR | UASB | Control | H2 Added | Control | “A” Reactor | “B” Reactor | “C” Reactor | |
| Biogas composition (%) | After stabilization | After stabilization | After stabilization | ||||||||
| CH4 | 67.1 ± 0.8 | 85.1 ± 3.7 | 69.2 ± 1 | 86.4 ± 1.0 | 91.0 ± 2.0 | 65.2 ± 9.8 | 89.4 ± 0.4 | 38.57 ± 1.96 | 87.48 ± 5.86 | 95.48 ± 5.01 | 94.13 ± 4.90 |
| CO2 | 32.9 ± 0.9 | 6.6 ± 0.9 | 30.7 ± 1 | 10.7 ± 3.6 | 7.0 ± 1.0 | 34.8 ± 9.8 | 10.5 ± 0.4 | 61.43 ± 8.51 | 6.69 ± 2.31 | 1.56 ± 1.00 | 1.04 ± 0.56 |
| H2 | 0.0 | 8.3 ± 3.6 | 0.0 | 3.5 ± 1.5 | 2.0 ± 1.0 | 0.0 | 0.0 | 0.0 | 2.99 ± 1.61 | 1.90 ± 0.90 | 3.49 ± 2.50 |
| Gas production | |||||||||||
| (mL·L−1·d−1) | |||||||||||
| CH4 | 247 ± 27 | 359 ± 20 | n.d. | n.d. | 352.53 ± 53 | 224 | 267 | 19.8 ± 0.4 | 131.2 ± 14.4 | 197.3 ± 4.4 | 270.7 ± 7.4 |
| CH4 from H2 | 0 | 112.0 | 0.0 | n.d. | n.d. | 0.0 | 127.0 | 0.0 | 111.4 | 177.5 | 250.9 |
| CO2 | 121 ± 15 | 28 ± 5 | n.d. | n.d. | 28.03 ± 4.7 | 119.6 | 31.4 | 31.0 ± 3.3 | 16.1 ± 0.7 | 11.1 ± 0.8 | 8.5 ± 1.8 |
| H2 injection rate | 0.0 | 510 ± 32 | 0.0 | 550 | n.d. | 0.0 | 507 ± 30 | 0.0 | 540.3 ± 71.6 | 945 ± 51.3 | 1306 ± 45.5 |
| (mL·L−1·d−1) | |||||||||||
| H2 consumption | 0.0 | 470 ± 35 | 0.0 | n.d. | n.d. | 0.0 | 507 ± 30 | 0.0 | 509.00 ± 1.67 | 854.9 ± 3.48 | 1168.41 ± 4.98 |
| (mL·L−1·d−1) | |||||||||||
| H2 consumption | 0.0 | 92.20 | 0.0 | n.d. | n.d. | 0.0 | 100.00 | 0.0 | 99.05 | 99.40 | 98.90 |
| (%) | |||||||||||
| pH | 7.82 ± 0.16 | 8.49 ± 0.04 | n.d. | 8.6 ± 0.0 | 8.1 ± 0 | 7.49 ± 0.17 | 7.00-8.00 | 8.57 ± 0.19 | 8.75 ± 0.11 | 8.88 ± 0.09 | 8.52 ± 0.11 |
| Organic acids (g·L−1) | 1.18 ± 0.84 | 0.38 ± 0.07 | 0.2 | 2.7 | 0.1 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Acetate | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.00 | 0.06 | 0.04 | 0.04 |
| Propionate | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.05 | 0.05 | 0.03 | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szuhaj, M.; Wirth, R.; Bagi, Z.; Maróti, G.; Rákhely, G.; Kovács, K.L. Development of Stable Mixed Microbiota for High Yield Power to Methane Conversion. Energies 2021, 14, 7336. https://doi.org/10.3390/en14217336
Szuhaj M, Wirth R, Bagi Z, Maróti G, Rákhely G, Kovács KL. Development of Stable Mixed Microbiota for High Yield Power to Methane Conversion. Energies. 2021; 14(21):7336. https://doi.org/10.3390/en14217336
Chicago/Turabian StyleSzuhaj, Márk, Roland Wirth, Zoltán Bagi, Gergely Maróti, Gábor Rákhely, and Kornél L. Kovács. 2021. "Development of Stable Mixed Microbiota for High Yield Power to Methane Conversion" Energies 14, no. 21: 7336. https://doi.org/10.3390/en14217336
APA StyleSzuhaj, M., Wirth, R., Bagi, Z., Maróti, G., Rákhely, G., & Kovács, K. L. (2021). Development of Stable Mixed Microbiota for High Yield Power to Methane Conversion. Energies, 14(21), 7336. https://doi.org/10.3390/en14217336