Microsaccade Rate as a Measure of Drug Response
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Introduction
Methods
Participants
Eye tracking
Procedure
Data analysis
Results
Main Results
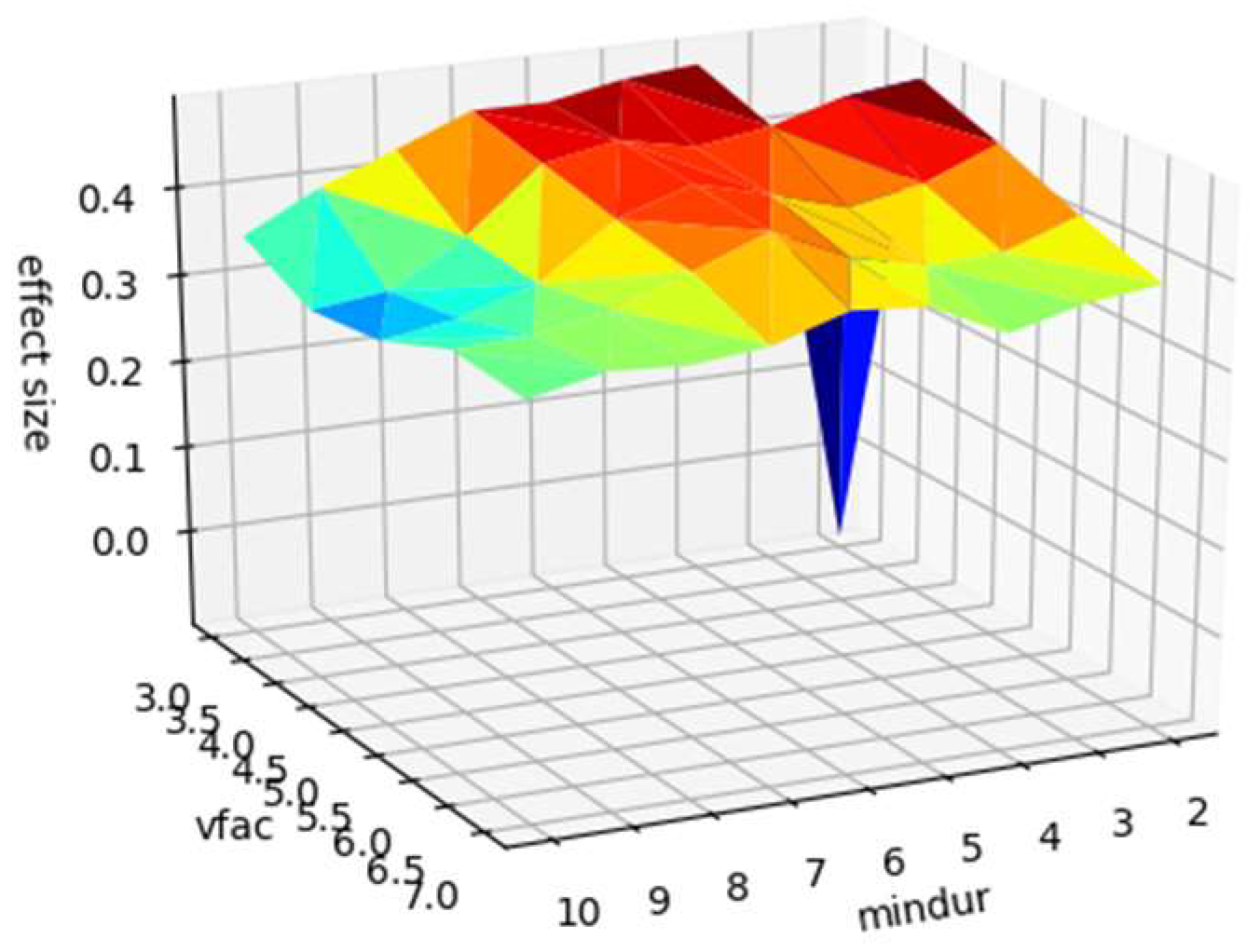
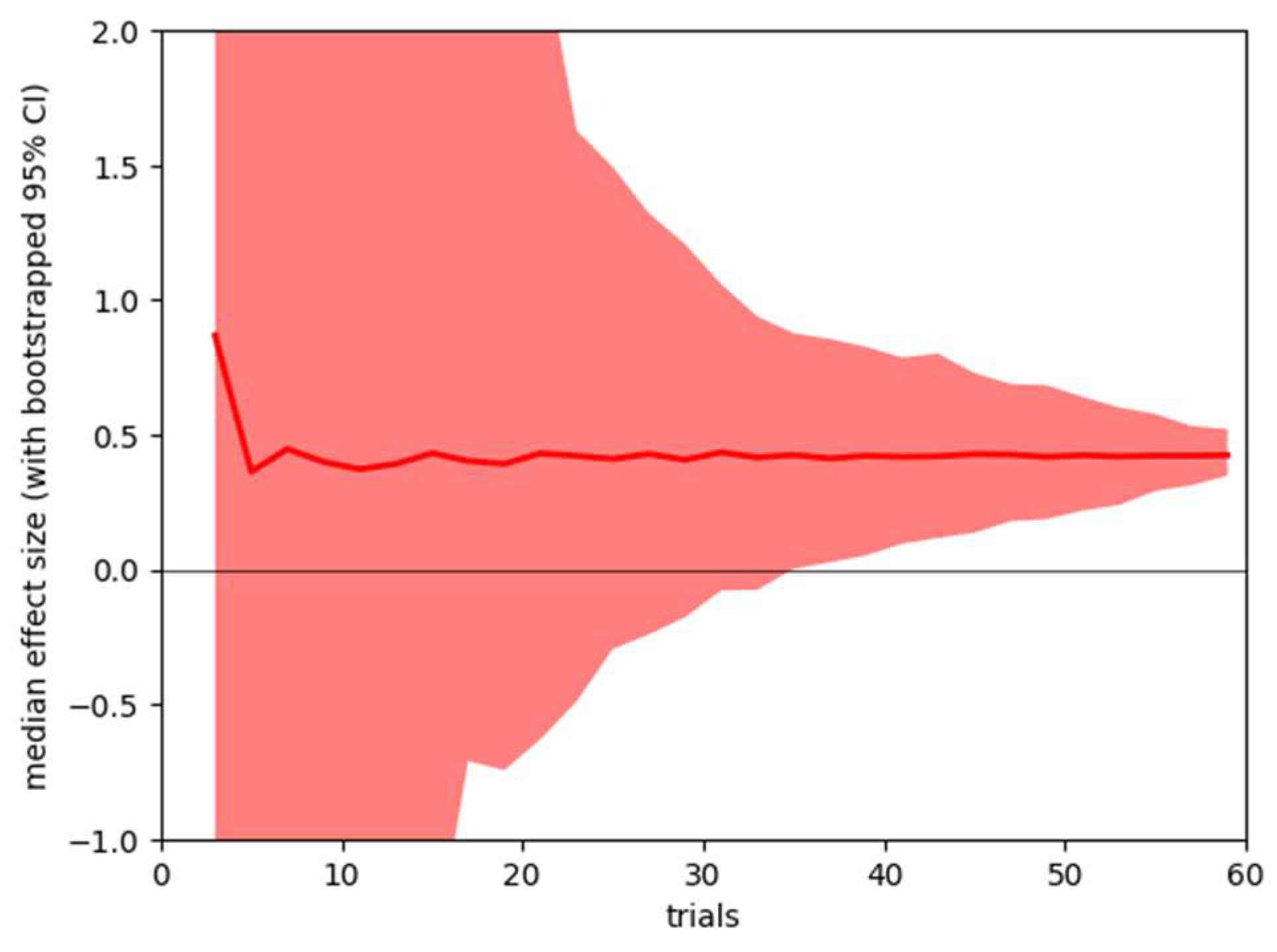
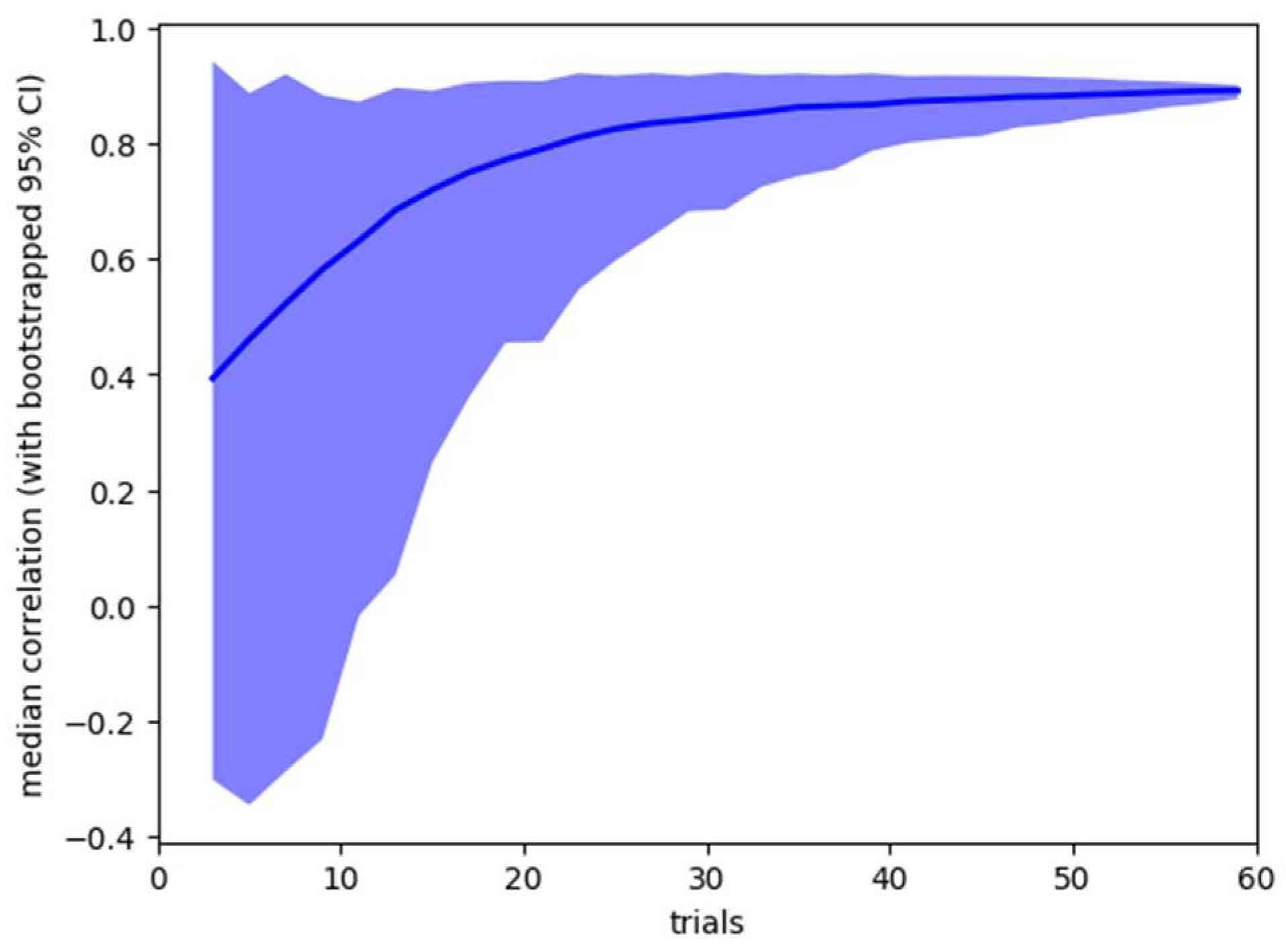
Sensitivity Analysis
Discussion
Acknowledgments
Conflicts of Interest
References
- Choe, K., R. Blake, and S. Lee. 2016. Pupil size dynamics during fixation impact the accuracy and precision of video-based gaze estimation. Vision Research 118: 48–59. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. 1992. A power primer. Psychological Bulletin 112, 1: 155–159. [Google Scholar] [CrossRef] [PubMed]
- Collins, N., M. Al-Kalbani, P. Kavanagh, A. Mahmud, G. Boyle, and D. Coakley. 2009. Effects of Caffeine on Fixational Eye Movements. Investigative Ophthalmology & Visual Science 50, 13: 2882. [Google Scholar]
- Dalmaijer, E., S. Mathôt, and S. Van der Stigchel. 2013. PyGaze: An open-source, cross-platform toolbox for minimal-effort pro-gramming of eyetracking experiments. Behavior Research Methods 46, 4: 913–921. [Google Scholar] [CrossRef]
- Dankner, Y., L. Shalev, M. Carrasco, and S. Yuval-Greenberg. 2017. Prestimulus inhibition of saccades in adults with and without attention-deficit/hyperactivity disorder as an index of temporal expectations. Psychological Science 28, 7: 835–850. [Google Scholar]
- Di Stasi, L. L., M. B. McCamy, A. Catena, S. L. Macknik, J. J. Canas, and S. Martinez-Conde. 2013. Microsaccade and drift dynamics reflect mental fatigue. European Journal of Neuroscience 38, 3: 2389–2398. [Google Scholar]
- Engbert, R., P. Sinn, K. Mergenthaler, and H. Trukenbrod. 2015. Microsaccade Toolbox for R. Available online: http://read.psych.uni-potsdam.de/index.php?option=com_content&view=article&id=140:engbert-et-al-2015-microsaccade-toolbox-for-r&catid=26:publications&Itemid=34.
- Fredholm, B., K. Bättig, J. Holmén, A. Nehlig, and E. Zvartau. 1999. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacological Reviews 51, 1: 83–133. [Google Scholar]
- Fried, M., E. Tsitsiashvili, Y. Bonneh, A. Sterkin, T. Wygnanski-Jaffe, T. Epstein, and U. Polat. 2014. ADHD subjects fail to suppress eye blinks and microsaccades while anticipating visual stimuli but recover with medication. Vision Research 101: 62–72. [Google Scholar] [CrossRef]
- Goffart, L., Z. Hafed, and R. Krauzlis. 2012. Visual Fixation as Equilibrium: Evidence from Superior Colliculus Inactivation. Journal Of Neuroscience 32, 31: 10627–10636. [Google Scholar] [CrossRef]
- Goodman, R., and S. Synder. 1982. Autoradiographic localization of adenosine receptors in rat brain using [3H]cyclohexyladenosine. The Journal Of Neuroscience 2, 9: 1230–1241. [Google Scholar] [CrossRef]
- Hafed, Z., L. Goffart, and R. Krauzlis. 2009. A Neural Mechanism for Microsaccade Generation in the Primate Superior Colliculus. Science 323, 5916: 940–943. [Google Scholar] [CrossRef] [PubMed]
- Hirai, H., and Y. Okada. 1995. Adenosine facilitates in vivo neurotransmission in the superior colliculus of the rat. Journal of Neurophysiology 74, 3: 950–960. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, S., K. Saijoh, and Y. Okada. 1997. Endogenous adenosine facilitates neurotrans-mission via A 2A adenosine receptors in the rat superior colliculus in vivo. Brain Research 757, 2: 268–275. [Google Scholar] [CrossRef] [PubMed]
- Johansson, B., and B. Fredholm. 1995. Further characterization of the binding of the adenosine receptor agonist [3H]CGS 21680 to rat brain using autoradiography. Neuropharmacology 34, 4: 393–403. [Google Scholar] [CrossRef]
- Kessler, R., L. Adler, M. Ames, O. Demler, S. Faraone, E. Hiripi, and et al. 2005. The World Health Organization adult ADHD self-report scale (ASRS): a short screening scale for use in the general population. Psychological Medicine 35, 2: 245–256. [Google Scholar] [CrossRef]
- Liguori, A., J. Hughes, and J. Grass. 1997. Absorption and Subjective Effects of Caffeine from Coffee, Cola and Capsules. Pharmacology Biochemistry And Behavior 58, 3: 721–726. [Google Scholar] [CrossRef]
- Magon, R., and U. Müller. 2012. ADHD with comorbid substance use disorder: review of treatment. Advances In Psychiatric Treatment 18, 6: 436–446. [Google Scholar] [CrossRef]
- McLellan, T., J. Caldwell, and H. Lieberman. 2016. A review of caffeine’s effects on cognitive, physical and occupational performance. Neuroscience & Biobehavioral Reviews 71: 294–312. [Google Scholar] [CrossRef]
- Mihali, A., A. G. Young, L. A. Adler, M. Halassa, and W. J. Ma. 2018. T186. Perceptual and executive behavioral deficits in ADHD and their differential correlation with microsaccade rate. Biological Psychiatry 83, 9: S200. [Google Scholar]
- Overton, P. 2008. Collicular dysfunction in attention deficit hyperactivity disorder. Medical Hypotheses 70, 6: 1121–1127. [Google Scholar] [CrossRef]
- Panagiotidi, M., P. Overton, and T. Stafford. 2017a. Attention-Deficit Hyperactivity Disorder-Like Traits and Distractibility in the Visual Periphery. Perception 46, 6: 665–678. [Google Scholar] [CrossRef] [PubMed]
- Panagiotidi, M., P. Overton, and T. Stafford. 2017b. Increased microsaccade rate in individuals with ADHD traits. Journal of Eye Movement Research 10, 1: 1–9. [Google Scholar] [CrossRef]
- Panagiotidi, M., P. Overton, and T. Stafford. 2017c. Multisensory integration and ADHD-like traits: Evidence for an abnormal temporal integration window in ADHD. Acta Psychologica 181: 10–17. [Google Scholar] [CrossRef] [PubMed]
- Panagiotidi, M., P. Overton, and T. Stafford. 2018. The relationship between ADHD traits and sensory sensitivity in the general population. Comprehensive Psychiatry 80: 179–185. [Google Scholar] [CrossRef]
- Roberts, M., B.K. Ashinoff, F.X. Castellanos, and M. Carrasco. 2018. When attention is intact in adults with ADHD. Psychonomic Bulletin & Review 25, 4: 1423–1434. [Google Scholar]
- Rosin, D., A. Robeva, R. Woodard, P. Guyenet, and J. Linden. 1998. Immunohistochemical localization of adenosine A2A receptors in the rat central nervous system. The Journal of Comparative Neurology 401, 2: 163–186. [Google Scholar] [CrossRef]
- Van Gisbergen, J., D. Robinson, and S. Gielen. 1981. A quantitative analysis of generation of saccadic eye movements by burst neurons. Journal Of Neurophysiology 45, 3: 417–442. [Google Scholar] [CrossRef]
- Volkow, N., G. Wang, J. Logan, D. Alexoff, J. Fowler, P. Thanos, and et al. 2015. Caffeine increases striatal dopamine D2/D3 receptor availability in the human brain. Translational Psychiatry 5, 4: e549. [Google Scholar] [CrossRef]
- Walker, L., A. Abraham, and K. Tercyak. 2010. Adolescent Caffeine Use, ADHD, and Cigarette Smoking. Children’s Health Care 39, 1: 73–90. [Google Scholar] [CrossRef]
- Xue, L., D. Huang, T. Wang, Q. Hu, X. Chai, L. Li, and Y. Chen. 2017. Dynamic modulation of the perceptual load on microsaccades during a selective spatial attention task. Scientific Reports 7, 1. [Google Scholar] [CrossRef]


Copyright © 2019. This article is licensed under a Creative Commons Attribution 4.0 International License.
Share and Cite
Hampsey, E.; Overton, P.G.; Stafford, T. Microsaccade Rate as a Measure of Drug Response. J. Eye Mov. Res. 2019, 12, 1-10. https://doi.org/10.16910/jemr.12.6.12
Hampsey E, Overton PG, Stafford T. Microsaccade Rate as a Measure of Drug Response. Journal of Eye Movement Research. 2019; 12(6):1-10. https://doi.org/10.16910/jemr.12.6.12
Chicago/Turabian StyleHampsey, Elliot, Paul G. Overton, and Tom Stafford. 2019. "Microsaccade Rate as a Measure of Drug Response" Journal of Eye Movement Research 12, no. 6: 1-10. https://doi.org/10.16910/jemr.12.6.12
APA StyleHampsey, E., Overton, P. G., & Stafford, T. (2019). Microsaccade Rate as a Measure of Drug Response. Journal of Eye Movement Research, 12(6), 1-10. https://doi.org/10.16910/jemr.12.6.12