Introduction
Over the past 10 years, transcranial direct current stimulation (tDCS) has been widely used to modulate cortical excitability to the benefit of cognitive and motor functions, in both healthy and clinical populations (
Stagg & Nitsche, 2011). However, studies investigating the effects of tDCS on control over eye movements have been scarce. Only recently evidence has emerged demon-strating that positively charged anodal tDCS applied over the frontal eye field (FEF) can be used to improve sac-cadic eye movement control in healthy young adults (Kanai, Muggleton, & Walsh, 2012). Given that healthy aging and a large number of age-related clinical condi-tions (e.g., mild cognitive impairment and Alzheimer’s and Parkinson’s disease) are associated with reduced control over the eye movement system, particularly when a high level of strategic control is required (Avila et al., 2015; Bos & Machado, 2013; Peltsch, Hemraj, Garcia, & Munoz, 2011, 2014), findings in
Kanai et al. (
2012) are exciting as they suggest that anodal tDCS may be a useful therapeutic tool for improving voluntary control over the oculomotor system in impaired populations.
Voluntary control over saccadic eye movements in-volves complex underlying neuromechanisms by which cortical oculomotor regions must be able to impose top-down regulation over subcortical oculomotor regions (
Johnston & Everling, 2008). The antisaccade paradigm (
Hallett, 1978) is a tool commonly used for behavioral measurement of voluntary control over saccadic eye movements. A successful antisaccade involves moving the eyes in the opposite direction when a stimulus sud-denly appears in the peripheral visual field. This capabil-ity involves two control processes: 1) suppressing an unwanted reflexive prosaccade toward the peripheral stimulus; 2) voluntarily generating an eye movement away from the peripheral stimulus to the mirror position (
Machado & Rafal, 2000;
Munoz & Everling, 2004). The frontal subregions most commonly posited to underpin accurate performance of antisaccades are the FEF and dorsolateral prefrontal cortex (DLPFC; Luna, Velanova, & Geier, 2008; Pierrot-Deseilligny, Muri, Nyffeler, & Milea, 2005). However, as reviewed in Chen and Macha-do (2016), the contributions of the FEF and DLPFC to the suppression of reflexive prosaccades and the genera-tion of correct antisaccades remain unclear. While the literature is in agreement that the FEF is the key region supporting generation of volitional eye movements, there is disagreement as to which frontal subregion supports suppression of reflexive eye movements. Specifically, some have reported evidence implicating the FEF as the key region supporting suppression of reflexive eye movements (
Machado & Rafal, 2004), while others have claimed that DLPFC (Brodmann’s area 46) plays the main role in suppressing reflexive eye movements, as reviewed in Pierrot-Deseilligny, Milea, and Muri (2004).
The one study (
Kanai et al., 2012) that assessed the influences of tDCS over cerebral cortex on oculomotor behavior found that in healthy young adults anodal tDCS over the FEF influenced subsequent antisaccade perfor-mance such that reflexive errors were reduced contrala-terally without any effect on correct antisaccade latencies, and in addition subsequent correct prosaccade latencies were shortened contralaterally. These findings indicate that while anodal tDCS over the FEF facilitates suppres-sion of unwanted contraversive reflexive eye movements, it also speeds the latencies of wanted contraversive re-flexive eye movements. These anodal tDCS benefits peaked 10 to 30 minutes post stimulation. In this seminal study, electrode positioning over the FEF was determined based on predefined standardized coordinates using struc-tural magnetic resonance imaging (MRI) of each individ-ual. In the current study, we tested whether the benefits reported in
Kanai et al. (
2012) can be induced using a more clinically practical protocol that does not entail expensive tools or time consuming procedures (e.g., MRI) to determine electrode positioning, as using such tools falls outside available resources in many clinical settings.
In addition to assessing anodal tDCS over the FEF, in the current study we assessed whether applying anodal tDCS over DLPFC might also benefit saccadic eye movement control, especially with respect to suppressing unwanted reflexive saccades, as one might predict based on human brain lesion studies (Pierrot-Deseilligny et al., 2003; Pierrot-Deseilligny, Rivaud, Gaymard, & Agid, 1991; Ploner, Gaymard, Rivaud-Pechoux, & Pierrot-Deseilligny, 2005). Furthermore, in the current study, we assessed whether saccadic eye movement control benefits extend to older adults. Ample evidence from non-oculomotor studies indicates tDCS confers more robust cognitive benefits in older adults (Hsu, Ku, Zanto, & Gazzaley, 2015), presumably due to far more room for improvement and thus greater potential for benefit. How-ever, no studies to date have assessed whether the same applies to saccadic eye movement control. In testing the efficacy of tDCS to improve saccadic eye movement control, we compared oculomotor behavior ipsilateral versus contralateral to the anodal electrode (as per Kanai et al., 2012) and we also included a sham control condi-tion (in contrast to Kanai et al., 2012). This enabled us to determine whether performance contralateral to the FEF and DLPFC electrodes was superior to ipsilateral perfor-mance, and also whether it was superior to performance contralateral to sham stimulation.
In sum, the purpose of the current study was three-fold: 1) determine whether benefits of anodal tDCS on saccadic eye movement behavior can be induced using a clinically practical protocol; 2) determine whether anodal tDCS over DLPFC also benefits saccadic eye movement control; 3) determine whether benefits extend to older adults.
Methods
Participants
Thirty adult males, 20 young (age range = 20-25 years, mean = 22.2,
SD = 1.0; education range = 15-18 years, mean = 16.2,
SD = 1.2) and 10 older (age range = 65-70 years, mean = 68.6,
SD = 1.1; education range = 10-31 years, mean = 15.1,
SD = 5.4) from the Dunedin community, New Zealand, participated and were reim-bursed NZ
$15 per session. Participants were all right-handed according to the Measurement of Handedness (
Chapman & Chapman, 1987). All participants reported having normal or corrected vision; no pace maker, im-planted electronic device or metal implants; no history of, and not currently taking any medications for neurological or psychiatric problems; no chronic skin conditions; and abstained from recreational drugs and alcohol in excess of three units during the 24 hours prior to their testing session. Participants also completed a depression invento-ry the Center for Epidemiologic Studies Depression Scale (CES-D; Radloff, 1977), which has a maximum score of 60. Of the young adults, 11 scored below 16, indicating they had no clinical symptoms of depression and nine scored between 16 and 22, indicating subthreshold de-pression symptoms. Of the older adults, five scored be-low 16, indicating they had no clinical symptoms of de-pression and five scored between 16 and 21, indicating subthreshold depression symptoms. Older adults were also screened for dementia using the Mini-Mental State Examination (MMSE; Folstein, Folstein, & McHugh, 1975); all scored at least 26 out of 30, which indicates none were demented. This study was approved by the University of Otago Human Ethics Committee (H13/123) and was performed in accordance with the relevant guide-lines and regulations. All participants gave informed consent prior to participation.
Design
The current study employed a randomized, single-blind, sham-controlled, crossover experimental design. As per
Kanai et al. (
2012), in the current study half of the participants in each age group were randomly assigned to have the anodal electrode positioned over the left hemi-sphere and half over the right. All participants completed three sessions of stimulation: active over each frontal subregion (FEF and DLPFC) and sham over an interme-diate frontal subregion, with the order of the stimulation conditions counterbalanced across participants within each age group, and each session separated by a mini-mum of 7 days. Each session lasted about 1 hr.
Electrodes Positioning
The 10-20 system for electroencephalography (EEG; Homan, Herman, & Purdy, 1987) was used to determine the placement of the anodal electrode over the assigned hemisphere. The anodal electrode was positioned for the FEF condition 1.5 cm anterior and 20% laterally from the vertex (Ro, Cheifet, Ingle, Shoup, & Rafal, 1999; Ro, Farne, & Chang, 2002), for the DLPFC condition 5 cm anterior and 20% laterally from the vertex (Pascual-Leone & Hallett, 1994; Pascual-Leone, Rubio, Pallardó, & Catalá, 1996), and for the sham condition 2.5 cm ante-rior and 20% laterally from the vertex (between the FEF and DLPFC positions). In all cases, the reference elec-trode (cathode) was positioned on the upper arm (just below the shoulder) ipsilateral to anodal electrode. Prior to proceeding, inspection of all sites of stimulation con-firmed there were no lesions or signs of skin irritation.
tDCS Protocol
A constant current 9 volt battery driven device (Acti-vaDose II) delivered 1 mA direct current through carbon rubber electrodes placed in sponge pockets soaked in saline solution. As per
Kanai et al. (
2012), the anodal electrode, which was 3 x 3 cm, delivered a current densi-ty of 0.11 mA/cm
2, and the reference electrode, which was 5 x 7 cm, delivered a current density of 0.03 mA/cm
2. The intensity of the current slowly ramped up to 1 mA over the initial 10 s of stimulation. During active stimulation current was delivered for 10 minutes, and during sham stimulation the device was turned off 30 s after the start of stimulation. At the end of each stimula-tion period, participants completed a questionnaire de-signed to monitor adverse effects. No adverse effects were reported, as might be expected given that the current density was 50 times lower than the previously studied safety threshold (
Liebetanz et al., 2009), and was also lower than the current densities used in many studies that were well tolerated and considered to be safe (Bikson, Datta, & Elwassif, 2009). When questioned at the end of their final session, no participants could differentiate between the active and sham conditions.
Eye Movement Testing
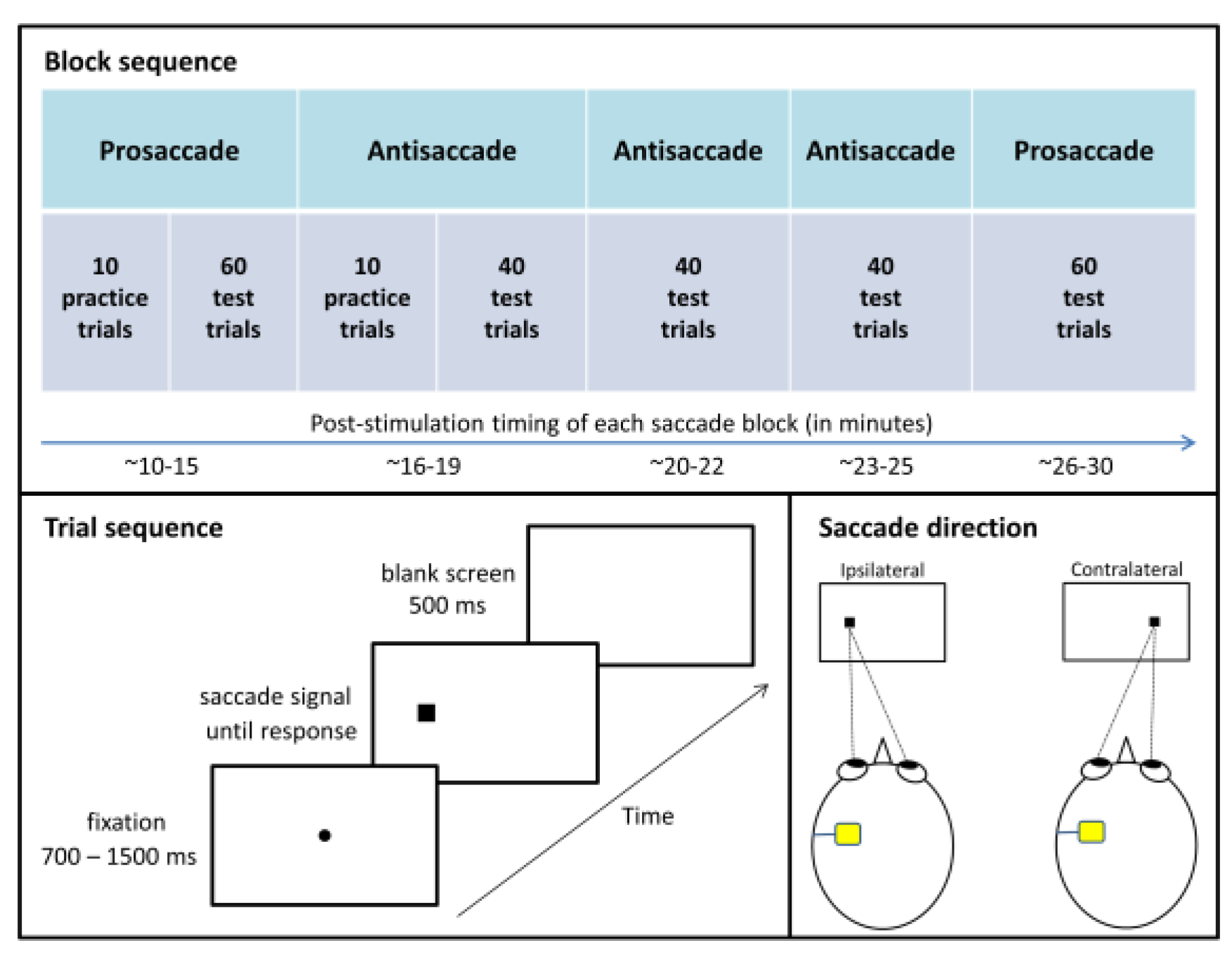
Figure 1 summarizes the eye movement testing pro-tocol, which was adapted from
Antoniades et al. (
2013). In order to target the post-stimulation time period that showed anodal tDCS benefits in
Kanai et al. (
2012), in the current study eye movement testing commenced 10 minutes post stimulation. Participants completed five eye movement blocks in this order: one block of prosaccades, three blocks of antisaccades, and then a second block of prosaccades. Between blocks, participants were provided with a 1 minute break. Between blocks of different types, the experimenter informed participants of the type of saccades required and instructed them in how to respond. Participants wore a head-mounted eye tracker (Model 310, Applied Science Laboratories, Massachusetts, USA) and sat 57 cm away from a computer screen in a dimly lit room, with distance maintained via a chinrest. The exper-imenter calibrated the eye-tracking system before each block. Stimuli were presented on a white background via MATLAB (The MathWorks, Natick, MA) and The Psy-chophysics Toolbox (
Brainard, 1997;
Pelli, 1997).
For prosaccade and antisaccade blocks, each trial commenced with the appearance of a black fixation dot extending 0.3° of visual angle and centered on the screen. After a variable interval (700, 900, 1100, 1300, or 1500 ms), the fixation dot disappeared and a black square sub-tending 1° appeared 8.5° to the left or right of center (measured to the center of the square). Fixation dot offset and peripheral square onset occurred simultaneously. Participants were instructed to respond to the appearance of the square as quickly as they could without compro-mising accuracy by looking at it during prosaccade blocks and by looking in the opposite direction during antisaccade blocks. During practice trials, a 900 Hz error tone sounded for 300 ms if participants made no re-sponse, responded in the wrong direction, or responded in less than 50 ms or more than 1000 ms after saccade sig-nal onset (i.e., the appearance of the peripheral square). The screen went blank for 500 ms between trials. Saccade signal position (left or right) and fixation duration (700, 900, 1100, 1300, or 1500 ms) were randomly selected for each trial with the constraint that each combination of conditions was equally likely to occur across the test trials. Each prosaccade test block had 60 trials and each antisaccade test block had 40 trials, and participants were given 10 practice trials at the beginning of the first block of each saccade type. Practice trials were repeated upon request by participants or if the experimenter identified the participant did not understand the instruction.
Horizontal position of the right eye was sampled at 1100 Hz. When the right eye exceeded the horizontal velocity of 50°/s with at least 1° amplitude, the move-ment was defined as a saccade. The program then record-ed the latency of saccade onset (by backtracking until the velocity dropped below 10°/s) and the direction of movement. During the trials, the experimenter manually rejected responses contaminated by blinking or other factors such as sneezing or coughing. In addition, trials were excluded from analysis if eye position at the time of saccade signal onset deviated from center by more than 3°, or if the latency was shorter than 50 ms or longer than 1000 ms.
Statistical Analyses
For each participant, the measured variables of inter-est were correct median reaction times (RTs) and per-centage of reflexive errors during antisaccade blocks as a function of stimulation condition (FEF, DLPFC, or sham) and saccade direction (ipsilateral or contralateral to the anodal electrode). Shapiro-Wilk test was used to deter-mine the normality of each data set. When assumptions of parametric tests were violated, non-parametric tests con-firmed the parametric results. In cases where sphericity was violated (
p < .05), a Greenhouse-Geisser correction was applied when Epsilon ranged from .70 to .90, other-wise a multivariate test (Pilai’s Trace) was applied. The alpha level was
p < .05. The sample size was chosen based on
Kanai et al. (
2012), which reported significant results for contralateral versus ipsilateral performance in a group of 16 young adults; a power analysis computed using G*Power 3.1.9.2 (Faul, Erdfelder, Lang, & Buch-ner, 2007) indicated our study had 87% power to detect a similar effect size (
dz = 0.5875), and thus beta was 0.13. Note that stimulation was always applied unilaterally, and the results were coded based on whether the saccade was directed ipsilaterally or contralaterally to the stimulated hemisphere (see
Figure 1, lower right panel, for exam-ples).
Results
To determine if performance varied across the blocks, initial repeated-measures analyses of variance (ANOVAs), with stimulation condition, saccade direction, and saccade block as factors, were performed for each of the measured variables of interest (prosaccade latencies, antisaccade latencies, and reflexive error rates during antisaccades). The results revealed no main effect of block for the latency variables, but there was a main effect of block for reflexive error rates during antisaccades,
F(2, 58) = 9.398,
p < .001,
r = .495, reflecting increasing reflexive error rates across blocks, presumably due to fatigue. However, since saccade block did not interact with stimulation condition or saccade direction for any of the measured variables of interest (all
ps > .200), the data were collapsed across blocks in the mixed ANOVAs reported below, all of which included age group as a between-participant factor, and stimulation condition and saccade direction as within-participant factors. Regardless of the ANOVA results, paired samples
t tests assessed hemispheric asymmetries in the active conditions and differences against sham stimulation. In addition, in light of individual differences in responsiveness to brain stimulation (Wiethoff, Hamada, & Rothwell, 2014), to determine whether a subset of the participants benefitted from active stimulation, each individual’s data was checked for any apparent asymmetries in the active stimulation conditions consistent with superior performance contralateral versus ipsilateral, and if so contralateral active versus sham; paired-samples
t tests tested whether any of the differences reached significance.
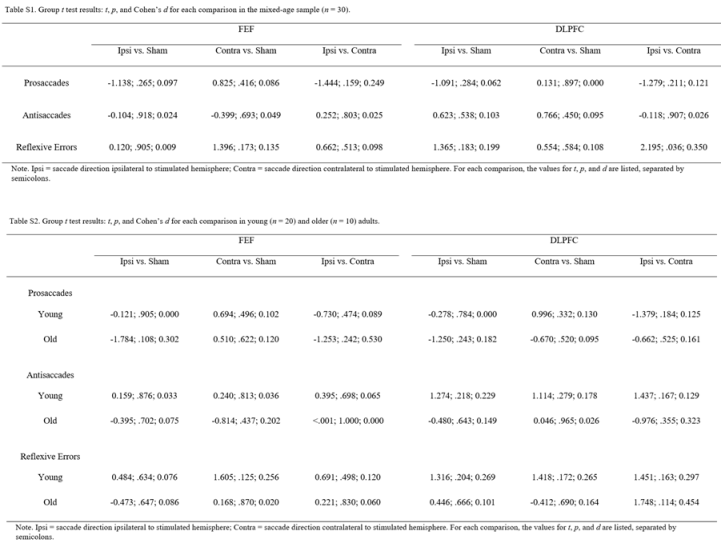
Table 1 summarizes the mean of the median correct response latencies and reflexive error rates during antisaccades for each stimulation condition in each age group. Tables S1 and S2 (in Appendix) detail the results of each group-level
t test.
Prosaccade Latencies
Figure 2 summarizes the latency data for the prosac-cade blocks. The mixed ANOVA revealed a significant main effect of age group,
F(1, 28) = 16.102,
p < .001,
r = .604, reflecting longer latencies in older than young adults. The expected two-way interaction between stimu-lation condition and saccade direction approached signif-icance,
F(2, 56) = 2.455,
p = .095,
r = .285; however, in contrast to the expected shortening of contralateral rela-tive to ipsilateral prosaccade latencies, contralateral la-tencies tended to be longer particularly in the FEF stimu-lation condition, although a paired-samples
t test showed that this trend for an asymmetry in the FEF stimulation condition did not reach significance (
p = .159). The three way interaction did not approach significance,
F(2, 56) = 1.684,
p = .195,
r = .239, and as can be seen in
Figure 2 neither age group exhibited the predicted pattern of faster contralateral prosaccades. No other main effects or inter-actions approached significance (all
ps > .100). Paired-samples
t tests computed for the full age-mixed sample confirmed no ipsilateral versus contralateral latency dif-ferences in either active stimulation condition (FEF or DLPFC) and no differences relative to the sham stimula-tion condition (all
ps > .100; see
Table S1 for details). Similarly,
t tests computed for each age group confirmed no ipsilateral versus contralateral latency differences in either active stimulation condition and no differences relative to the sham stimulation condition (all
ps > .100; see
Table S2 for details).
Consideration of each individual’s data also indicated a lack of benefits. In the FEF stimulation condition, only three of the 20 young adults and none of the older adults showed significantly faster contralateral relative to ipsi-lateral latencies, consistent with the pattern reported in
Kanai et al. (
2012), and only one of these three reached significance when compared with contralateral latencies in the sham stimulation condition,
t(57) = 2.552,
p = .013,
Cohen’s d = 0.676. In the DLPFC stimulation condition, only one of the 20 young adults and none of the older adults showed this asymmetry pattern, and the compari-son with the sham stimulation condition did not reach significance (
p > .050).
Antisaccade Latencies
Figure 3 summarizes the latency data for the anti-saccade blocks. The mixed ANOVA revealed a signifi-cant main effect of age group,
F(1, 28) = 26.643,
p < .001,
r = .699, again reflecting longer latencies in older than young adults. Of specific relevance here, the interac-tion between stimulation condition and saccade direction did not approach significance,
F(2, 56) = 0.555,
p = .577,
r = .138, which indicates that the different stimulation conditions did not differentially influence contralateral versus ipsilateral latencies. Furthermore, stimulation condition and saccade direction did not significantly interact with age group,
F(2, 56) = 1.905,
p = .158,
r = .253. As can be seen in
Figure 3, neither age group showed stimulation effects (i.e., asymmetries specific to active stimulation). No other main effects or interactions approached significance (all
ps > .100). Paired-samples
t tests confirmed no ipsilateral versus contralateral latency differences in either active stimulation condition (FEF or DLPFC) and no differences relative to the sham stimula-tion condition (all
ps > .400; see
Table S1 for details). Similarly,
t tests computed for each age group confirmed no ipsilateral versus contralateral latency differences in either active stimulation condition and no differences relative to the sham stimulation condition (all
ps > .100; see
Table S2 for details).
Consideration of each individual’s data also indicated a lack of benefits. In the FEF stimulation condition, only one of the 20 young adults and one of the 10 older adults showed significantly faster contralateral relative to ipsi-lateral latencies, and only the young participant reached significance when compared with contralateral latencies in the sham stimulation condition, t(38) = 3.017, p = .005, Cohen’s d = 0.979. In the DLPFC stimulation condition, one of the young adults and none of the older adults showed this asymmetry pattern, and the comparison with the sham stimulation condition did not reach significance (p > .050).
Reflexive Error Rates During Antisaccade Blocks
Figure 4 summarizes the reflexive error rates during the antisaccade blocks. The mixed ANOVA revealed that, although the data showed the expected trend for higher reflexive error rates in older compared to young adults, the main effect of age group did not approach significance,
F(1, 28) = 1.252,
p = .273,
r = .207, which could be due to the small sample size in the older age group (
n = 10). Of specific relevance here, the expected two-way interaction between stimulation condition and saccade direction did not approach significance,
F(2, 56) = 1.731,
p = .194,
r = .241, which indicates that the dif-ferent stimulation conditions did not differentially influ-ence contralateral versus ipsilateral reflexive errors. In addition, stimulation condition and saccade direction did not interact with age group,
F(2, 56) = 0.381,
p = .685,
r = .114, which suggests that the lack of stimulation effects applies to both age groups. As shown in
Figure 4, the pattern of reduced reflexive error rates contralaterally relative to ipsilaterally emerged in all three stimulation conditions, including sham, in both age groups. No other main effects or interactions approached significance (all
ps > .100). Paired-samples
t tests confirmed no ipsilateral versus contralateral performance differences in the active FEF stimulation condition and no differences in either active stimulation condition (FEF or DLPFC) relative to the sham stimulation condition (all
ps > .100); however, fewer reflexive errors were made toward contralateral than ipsilateral saccade signals in the active DLPFC stimulation condition (
p = .036; see
Table S1 for details), but this asymmetry is unlikely to reflect the tDCS given that performance in the active DLPFC stimulation condi-tion did not differ from performance in the sham condi-tion and moreover that contralateral reflexive error rates were higher in the active DLPFC condition (10.2%) than in the sham condition (9.2%). Separate consideration of each age group showed no ipsilateral versus contralateral latency differences in either active stimulation condition and no differences relative to the sham stimulation condi-tion (all
ps > .100; see
Table S2 for details).
Discussion
Using a more clinically practical protocol, the current study tested whether anodal tDCS over the FEF can in-duce oculomotor benefits similar to those reported in young adults in
Kanai et al. (
2012), and in addition as-sessed whether applying anodal tDCS over DLPFC might also benefit oculomotor behavior and whether these bene-fits extend to older adults, who are known to have sac-cadic eye movement control deficits (
Chen & Machado, 2016). Overall the results revealed no evidence of oculo-motor benefits following anodal tDCS, despite the sample size in the current study exceeding that used in
Kanai et al. (
2012). Specifically, group analyses showed no differ-ences in the active stimulation conditions relative to sham stimulation, and an asymmetry in saccadic eye movement behavior arose only in the active DLPFC stimulation condition (for reflexive errors in the full mixed-age sam-ple), but this did not reflect better performance relative to sham performance, and the sham condition showed a similar pattern. Analyses of individual participants backed up the null results at the group level, with signifi-cant effects relative to sham stimulation occurring in less than 5% of the participants (which is consistent with chance levels, as alpha was set to .05). These results indicate that neither active stimulation site (FEF or DLPFC) afforded better saccadic eye movement control. The absence of oculomotor benefits arose in both age groups, despite the older adults exhibiting the expected saccadic eye movement control deficits that indicate ample room for improvement. These negative outcomes indicate that the clinically practical protocol utilized in the current study was ineffective.
One of the main findings reported in
Kanai et al. (
2012) was that anodal tDCS over the FEF reduced re-flexive error rates toward contralateral relative to ipsilat-eral antisaccade signals. This pattern was also demon-strated in the current study, although the asymmetry did not reach significance. However, as shown in
Figure 4, the same pattern also occurred in the sham stimulation condition. Given that
Kanai et al. (
2012) did not include a sham stimulation condition, it is not possible to determine whether the lower rate of contralateral versus ipsilateral reflexive errors occurred as a result of the tDCS. To de-termine this, one would need to replicate the protocol used in
Kanai et al. (
2012) with the addition of a sham stimulation condition. The fact that the current study showed similar asymmetric patterns in the active and sham conditions highlights the need for a sham control comparison condition to confirm whether any observed asymmetries are specifically attributable to tDCS. The other main finding reported in
Kanai et al. (
2012) is that anodal tDCS over the FEF shortened prosaccade latencies contralateral versus ipsilateral to the stimulated hemi-sphere. This pattern was not demonstrated in the current study in either age group (see
Figure 2). Furthermore, none of the older adults and only three of the 20 young adults showed this pattern, and only one of these three reached significance when compared with sham stimula-tion, which was not assessed in
Kanai et al. (
2012).
A number of factors could potentially explain the dis-crepant outcomes. One of the main differences in the design of the current study relative to
Kanai et al. (
2012) was the lack of precise localization of the FEF. To speed application to better suit clinical environments, in the current study we simplified the tDCS protocol by using basic EEG-based measurements to position the FEF elec-trode, in accordance with
Ro et al. (
1999) and
Ro et al. (
2002). However, there were several other design differ-ences that may have influenced the results. For example, the saccade paradigm used in the current study (adapted from Antoniades et al., 2013) differed from that used in
Kanai et al. (
2012), in that in their study permanent boxes marked the possible saccade signal locations (where as the saccade signal locations were unmarked in the current study), the fixation dot overlapped with the saccade sig-nal (where as the fixation dot disappeared when the sac-cade signal appeared in the current study), the fixation duration varied from 300-700 ms (700-1500 ms in the current study), the response period varied from 50-400 ms (50-1500 ms in the current study), the saccade veloci-ty threshold was 28.6°/s (50°/s in the current study), and eye position was sampled at 250 Hz (1100 Hz in the current study). Also, the reference electrode was placed on the shoulder (deltoid muscle) in
Kanai et al. (
2012) but on the upper arm in the current study. Although these design differences may have influenced the results, none of these design differences should affect performance asymmetrically, and thus they cannot explain asymme-tries present in
Kanai et al. (
2012) but not in the current study. Hence, the use of basic measurements to position the electrodes seems the most likely factor underpinning the discrepant results.
The lack of benefits in older adults came as a particu-lar surprise given that they have far more room for im-provement and past research indicates that tDCS can confer greater benefits in older adults (
Hsu et al., 2015). One factor that may have contributed to the failure to induce improvements in saccadic eye movement control in the older adults pertains to age-related increases in cerebral spinal fluid (
Good et al., 2001), which can atten-uate electric field strength (Laakso, Tanaka, Koyama, De Santis, & Hirata, 2015). Another factor that may have reduced the chances of inducing benefits in the older adults is that the tDCS protocol used may not suit older adults due to age-related changes in brain activation pat-terns (Bierre, Lucas, Guiney, Cotter, & Machado, 2017). As reviewed in Dayan, Censor, Buch, Sandrini, and Co-hen (2013), small electrodes stimulate more focally, which can be beneficial in some circumstances. However, given that older adults normally show widespread pre-frontal activation not seen in young adults especially when engaged in higher level cognitive processing (see Chen & Machado, 2016, for a review), focal stimulation may not be optimal to induce pervasive physiological changes necessary to enhance saccadic eye movement control in older adults.
Another factor that may have contributed more gener-ally to the lack of tDCS effects pertains to the spatial distribution of the induced electric field. As demonstrated in Moliadze, Antal, and Paulus (2010), the reference electrode positioning determines the direction of current flow whilst the distance between the electrodes deter-mines where the peak electric field is focused. Given that current passes between the two electrodes, an anode placed over the frontal region and a cathode (i.e., refer-ence electrode) placed over the deltoid muscle or upper arm leads to the current flowing in from the anodal elec-trode site, passing through the brainstem and the spinal cord, and diffusing at the site of the reference electrode (Im, Park, Shim, Chang, & Kim, 2012). This tDCS mon-tage, used in the current study and in
Kanai et al. (
2012), should have resulted in the electric field concentration (i.e., the “hotspot”) being distributed outside of prefrontal regions, roughly around the neck region. Thus, the elec-trode positions used here and in
Kanai et al. (
2012) may not be optimal for inducing physiological changes in prefrontal regions.
With respect to developing a tDCS protocol that is more likely to induce physiological changes required to improve functioning, especially in older adults, future studies should take into consideration using a contrala-teral encephalic reference electrode (e.g., over the fore-head or cheek), which should optimize the electric field in prefrontal regions (Jones, Stephens, Alam, Bikson, & Berryhill, 2015). This arrangement, usually involving a large active electrode over prefrontal cortex combined with a contralateral encephalic reference electrode, has shown promise in a large number of studies that reported improvements in non-oculomotor cognitive functions in older adults (Prehn & Flöel, 2015; Teixeira-Santos, Nafee, Sampaio, Leite, & Carvalho, 2015). This more typical montage may be worthy of assessment in relation to oculomotor functions as well.

{kind=link}
{kind=link}
{kind=link}
{kind=link}