
Antiproliferative Effect of Clitoria ternatea Ethanolic Extract against Colorectal, Breast, and Medullary Thyroid Cancer Cell Lines

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
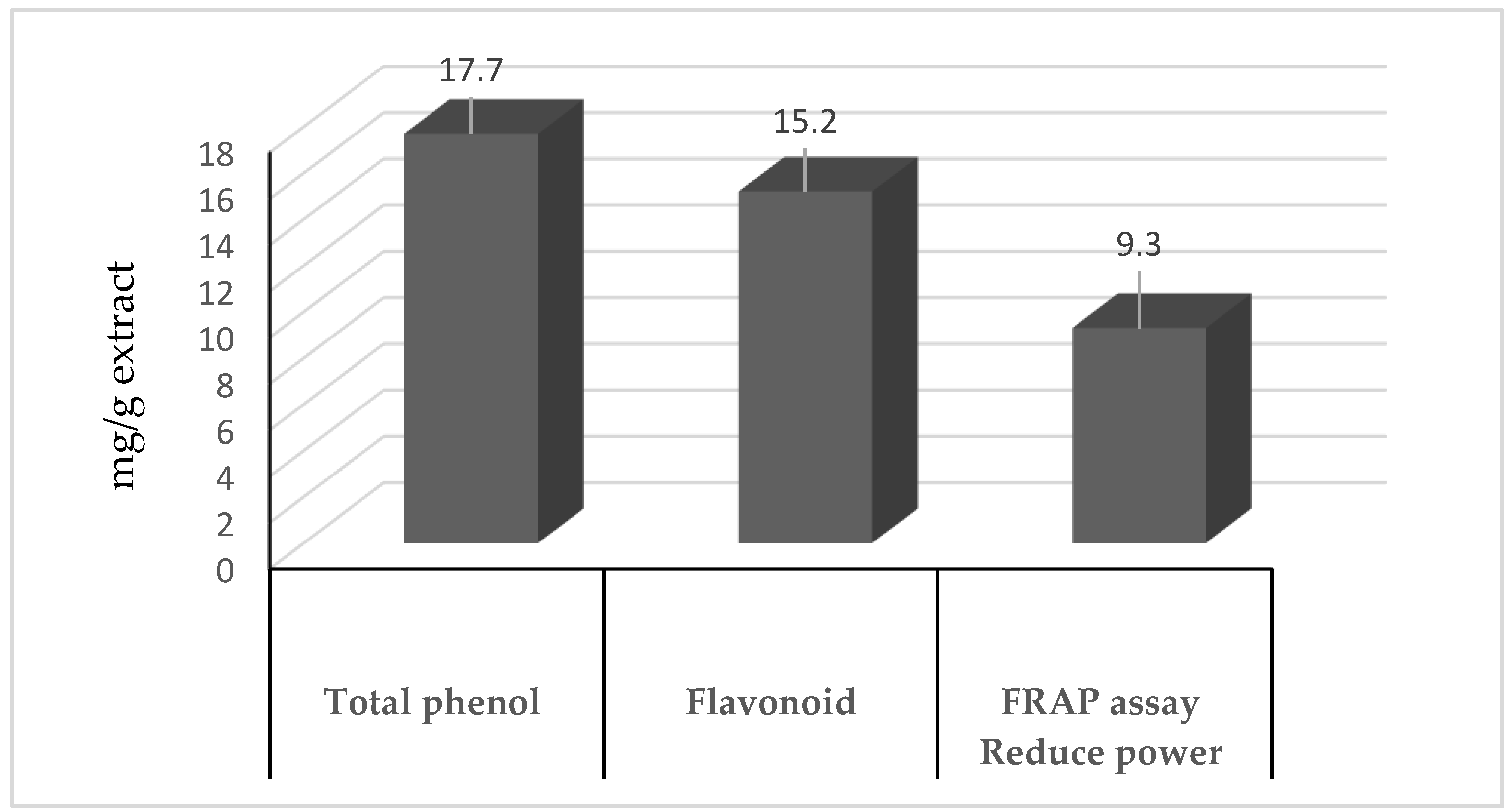
2.1. Antioxidants, Flavonoids, and Phenolics
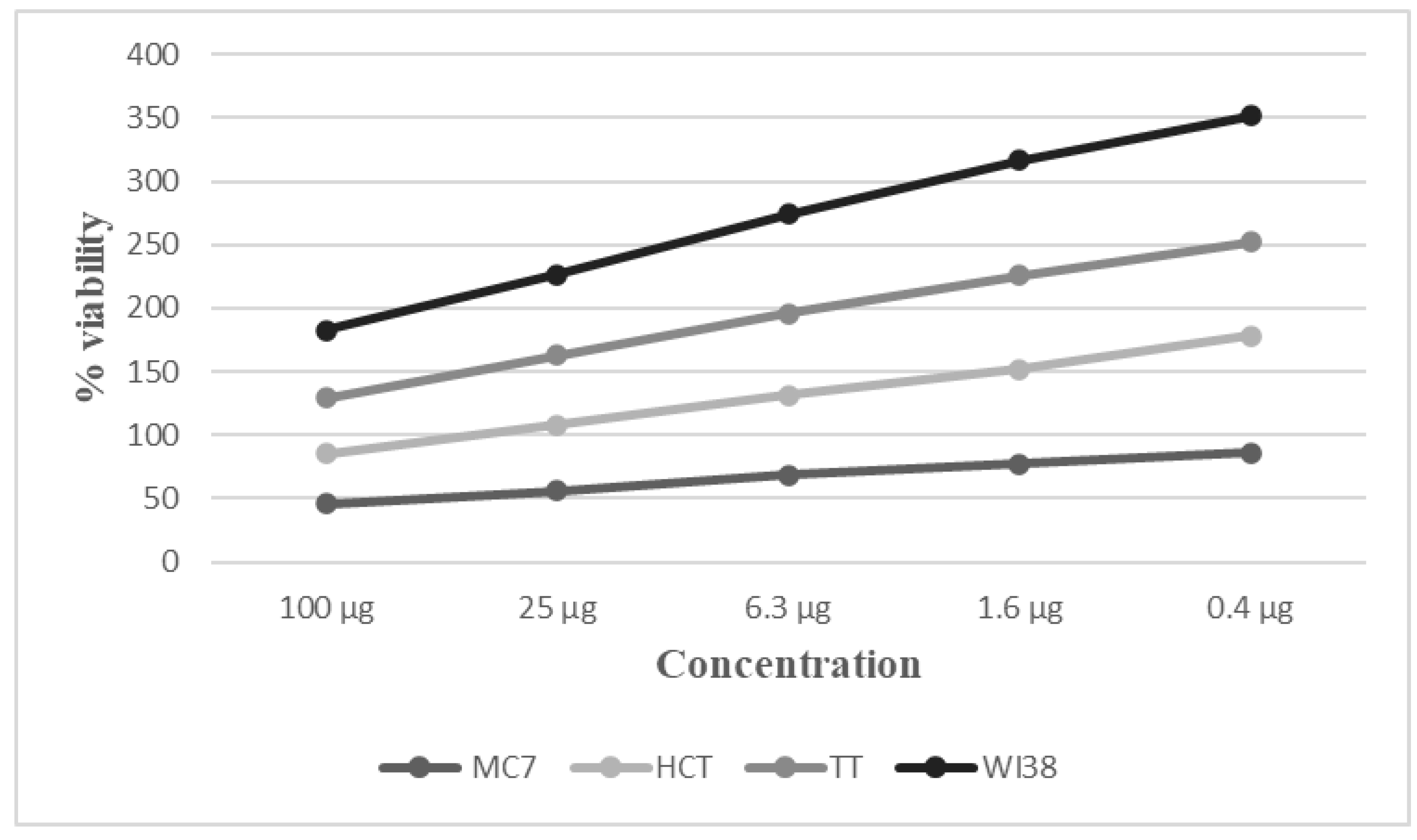
2.2. Cytotoxic Effects
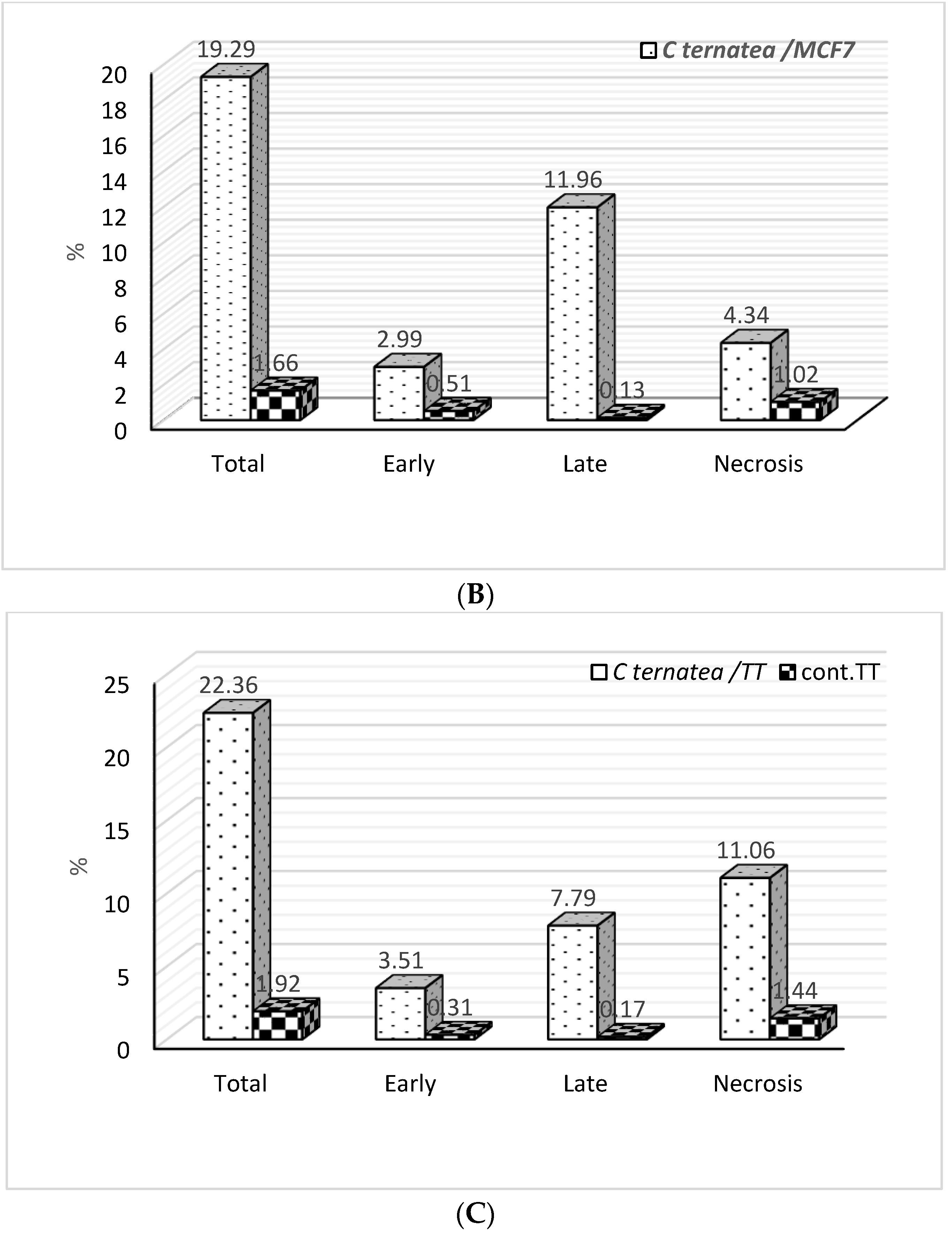
2.3. Apoptosis Detection
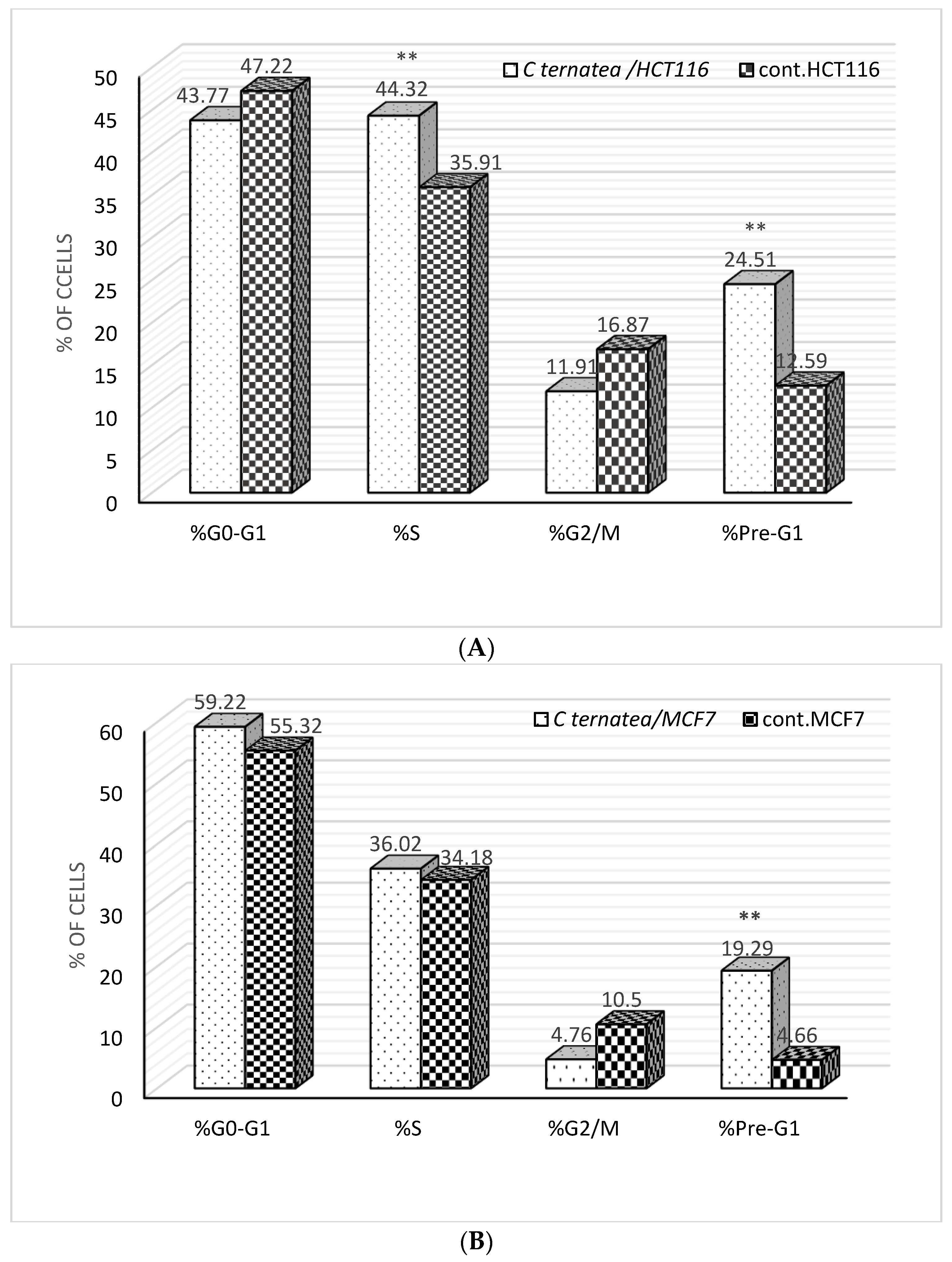
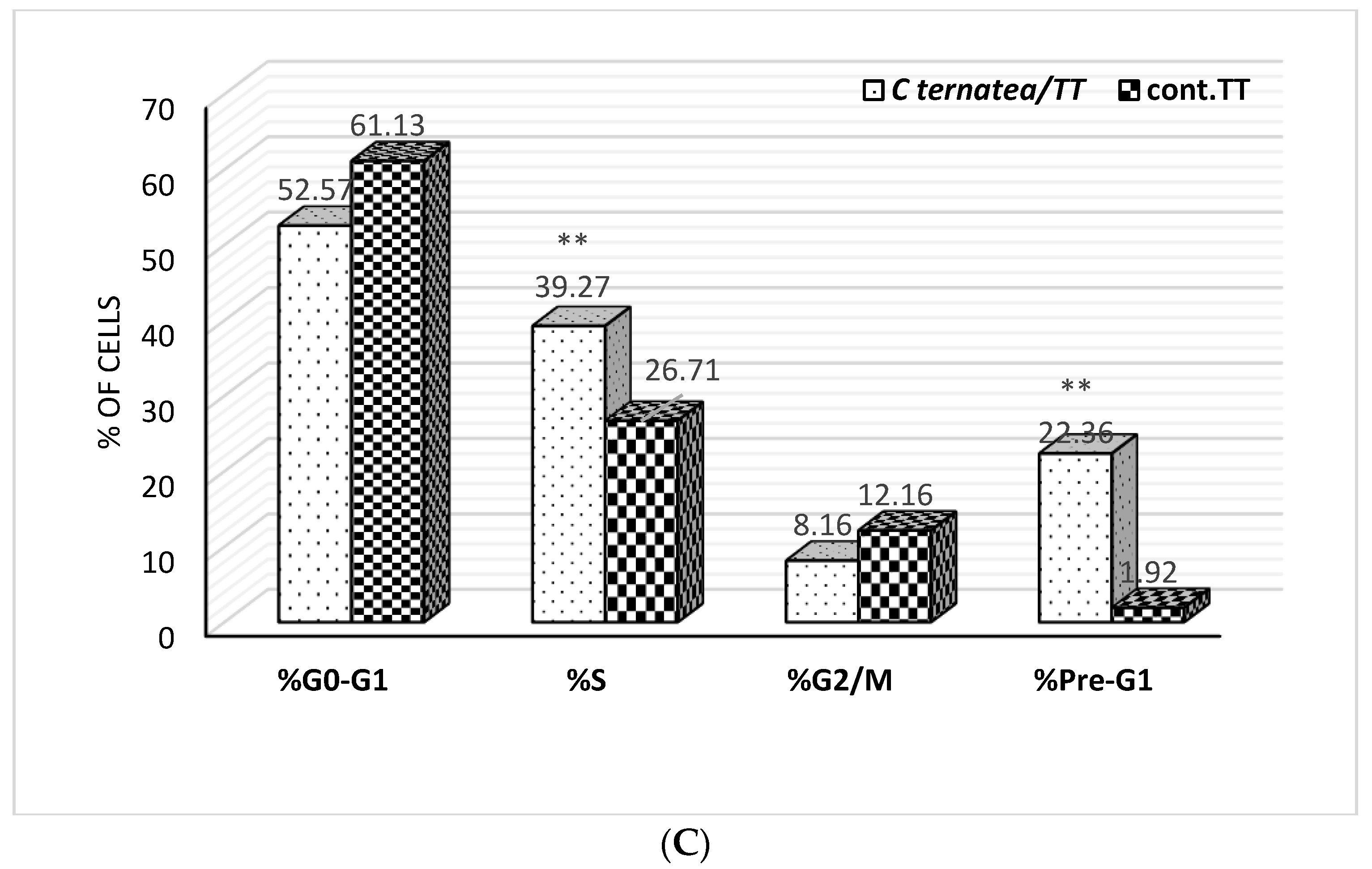
2.4. Cell Cycle Analysis
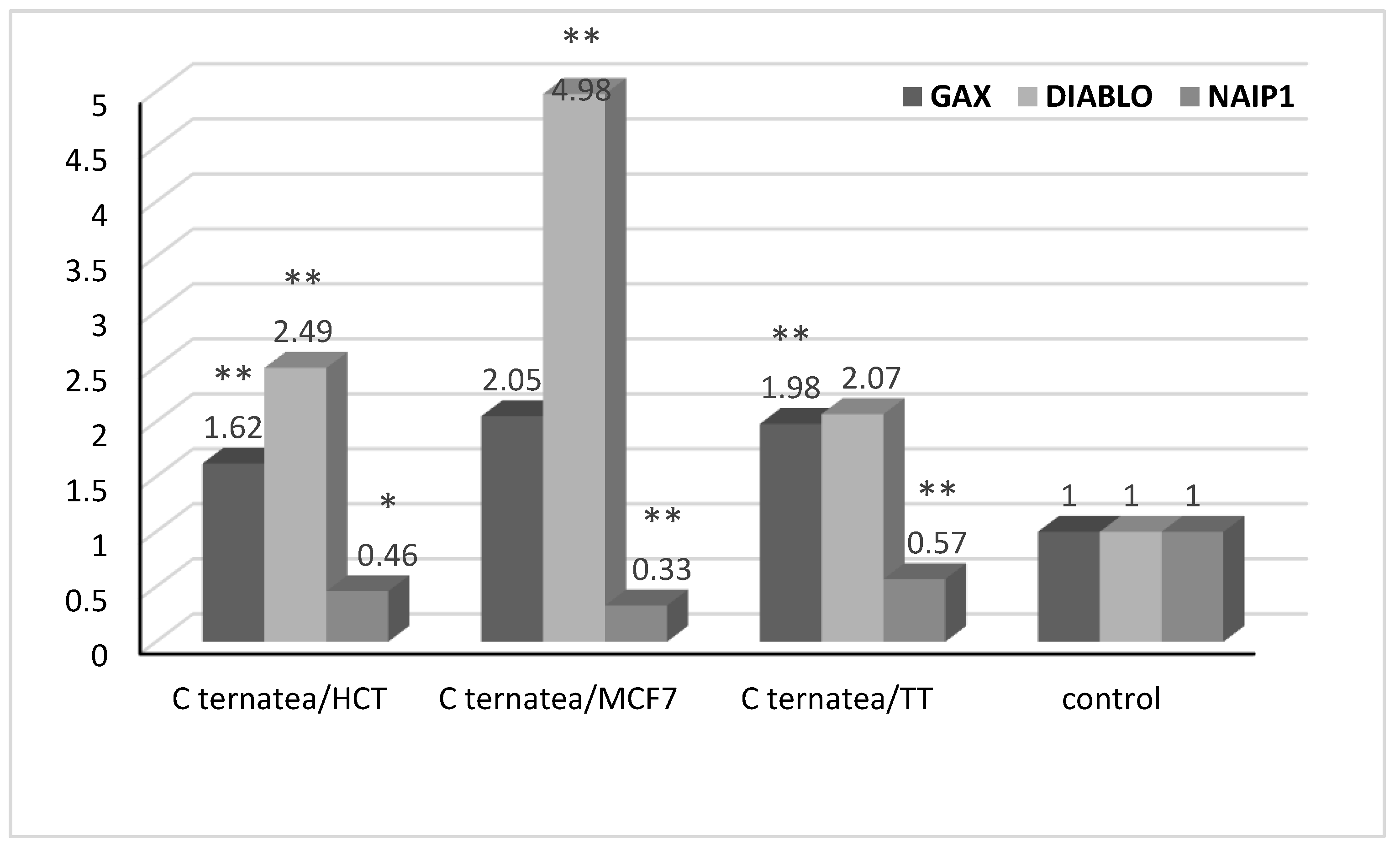
2.5. Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Research Design and Statistical Analysis
4.3. Ethanolic Extraction
4.4. Antioxidant, Phenolic, and Flavonoid Influences of the Plant Extract
4.5. Cell Cultures and Lines
4.6. In Vitro Cytotoxicity Examination
4.7. Apoptosis Detection
4.8. Cell Cycle Analysis
4.9. Expression Level Detection of GAX, DIABLO, and NAIP1 Genes
4.10. Real-Time Quantitative PCR (qPCR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The Ever-Increasing Importance of Cancer as a Leading Cause of Premature Death Worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- GBD 2019 Adolescent Young Adult Cancer Collaborators. The Global Burden of Adolescent and Young Adult Cancer in 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet Oncol. 2022, 23, 27–52. [Google Scholar] [CrossRef]
- GBD 2019 Stroke Collaborators. Global, Regional, and National Burden of Stroke and Its Risk Factors, 1990-2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet Neurol. 2021, 20, 795–820. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Global Health Estimates: Leading Causes of Death. Available online: https://www.who.int/data/gho/data/themes/mortality-and-global-health-estimates/ghe-leading-causes-of-death (accessed on 22 June 2022).
- Afifi, A.M.; Saad, A.M.; Al-Husseini, M.J.; Elmehrath, A.O.; Northfelt, D.W.; Sonbol, M.B. Causes of Death after Breast Cancer Diagnosis: A US Population-Based Analysis. Cancer 2020, 126, 1559–1567. [Google Scholar] [CrossRef]
- Rahib, L.; Wehner, M.R.; Matrisian, L.M.; Nead, K.T. Estimated Projection of US Cancer Incidence and Death to 2040. JAMA Netw. Open 2021, 4, e214708. [Google Scholar] [CrossRef]
- Kaspers, G.J.L. 20 Years of Expert Review of Anticancer Therapy. Expert Rev. Anticancer Ther. 2022, 22, 1–2. [Google Scholar] [CrossRef]
- Renzi, S.; Michaeli, O.; Ramaswamy, V.; Huang, A.; Stephens, D.; Maguire, B.; Tabori, U.; Bouffet, E.; Bartels, U. Causes of Death in Pediatric Neuro-Oncology: The Sickkids Experience from 2000 to 2017. J. Neurooncol. 2020, 149, 181–189. [Google Scholar] [CrossRef]
- van Niekerk, C.C.; Groenewoud, H.M.M.; Verbeek, A.L.M. Trends and Projections in Cutaneous Melanoma Death in the Netherlands from 1950 to 2045. Medicine 2021, 100, e27784. [Google Scholar] [CrossRef]
- Chen, X.; Zeh, H.J.; Kang, R.; Kroemer, G.; Tang, D. Cell Death in Pancreatic Cancer: From Pathogenesis to Therapy. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 804–823. [Google Scholar] [CrossRef]
- Wu, G.X.; Ituarte, P.H.G.; Ferrell, B.; Sun, V.; Erhunmwunsee, L.; Raz, D.J.; Kim, J.Y. Causes of Death and Hospitalization in Long-Term Lung Cancer Survivors: A Population-Based Appraisal. Clin. Lung Cancer 2020, 21, 204–213. [Google Scholar] [CrossRef]
- Neibart, S.S.; Manne, S.L.; Jabbour, S.K. Quality of Life After Radiotherapy for Rectal and Anal Cancer. Curr. Color. Cancer Rep. 2020, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Polanski, J.; Jankowska-Polanska, B.; Rosinczuk, J.; Chabowski, M.; Szymanska-Chabowska, A. Quality of Life of Patients with Lung Cancer. OncoTargets Ther. 2016, 9, 1023–1028. [Google Scholar] [CrossRef] [Green Version]
- Suarez-Almazor, M.; Pinnix, C.; Bhoo-Pathy, N.; Lu, Q.; Sedhom, R.; Parikh, R.B. Quality of Life in Cancer Care. Med 2021, 2, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.; Faithfull, S. “They’re Survivors Physically but We Want Them to Survive Mentally as Well”: Health Care Professionals’ Views on Providing Potential Late Effect Information. Support. Care Cancer 2013, 21, 2491–2497. [Google Scholar] [CrossRef] [Green Version]
- McKenna, N.P.; Bews, K.A.; Yost, K.J.; Cima, R.R.; Habermann, E.B. Bowel Dysfunction after Low Anterior Resection for Colorectal Cancer: A Frequent Late Effect of Surgery Infrequently Treated. J. Am. Coll. Surg. 2022, 234, 529–537. [Google Scholar] [CrossRef]
- GBD 2013 Mortality and Causes of Death Collaborators. Global, Regional, and National Age-Sex Specific All-Cause and Cause-Specific Mortality for 240 Causes of Death, 1990-2013: A Systematic Analysis for the Global Burden of Disease Study 2013. Lancet 2015, 385, 117–171. [Google Scholar] [CrossRef] [Green Version]
- Walli-Attaei, M.; Joseph, P.; Rosengren, A.; Chow, C.K.; Rangarajan, S.; Lear, S.A.; AlHabib, K.F.; Davletov, K.; Dans, A.; Lanas, F.; et al. Variations between Women and Men in Risk Factors, Treatments, Cardiovascular Disease Incidence, and Death in 27 High-Income, Middle-Income, and Low-Income Countries (PURE): A Prospective Cohort Study. Lancet 2020, 396, 97–109. [Google Scholar] [CrossRef]
- Mandlik Ingawale, D.S.; Namdeo, A.G. Pharmacological Evaluation of Ashwagandha Highlighting Its Healthcare Claims, Safety, and Toxicity Aspects. J. Diet. Suppl. 2021, 18, 183–226. [Google Scholar] [CrossRef]
- Wang, K.; Chen, Q.; Shao, Y.; Yin, S.; Liu, C.; Liu, Y.; Wang, R.; Wang, T.; Qiu, Y.; Yu, H. Anticancer Activities of TCM and Their Active Components against Tumor Metastasis. Biomed. Pharm. 2021, 133, 111044. [Google Scholar] [CrossRef]
- Buyel, J.F. Plants as Sources of Natural and Recombinant Anti-Cancer Agents. Biotechnol. Adv. 2018, 36, 506–520. [Google Scholar] [CrossRef] [PubMed]
- Illam, S.P.; Kandiyil, S.P.; Raghavamenon, A.C. Targeting Histone Onco- Modifications Using Plant-Derived Products. Curr. Drug Targets 2021, 22, 1317–1331. [Google Scholar] [CrossRef] [PubMed]
- Raees, M.A.; Hussain, H.; Al-Rawahi, A.; Csuk, R.; Muhammad, S.A.; Khan, H.Y.; Rehman, N.U.; Abbas, G.; Al-Broumi, M.A.; Green, I.R.; et al. Anti-Proliferative and Computational Studies of Two New Pregnane Glycosides from Desmidorchis Flava. Bioorg. Chem. 2016, 67, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Schnekenburger, M.; Dicato, M.; Diederich, M. Plant-Derived Epigenetic Modulators for Cancer Treatment and Prevention. Biotechnol. Adv. 2014, 32, 1123–1132. [Google Scholar] [CrossRef]
- Oguis, G.K.; Gilding, E.K.; Jackson, M.A.; Craik, D.J. Butterfly Pea (Clitoria ternatea), a Cyclotide-Bearing Plant With Applications in Agriculture and Medicine. Front. Plant Sci. 2019, 10, 645. [Google Scholar] [CrossRef] [Green Version]
- Oguis, G.K.; Gilding, E.K.; Huang, Y.-H.; Poth, A.G.; Jackson, M.A.; Craik, D.J. Insecticidal Diversity of Butterfly Pea (Clitoria ternatea) Accessions. Ind. Crop. Prod. 2020, 147, 112214. [Google Scholar] [CrossRef]
- Devi, B.P.; Boominathan, R.; Mandal, S.C. Anti-Inflammatory, Analgesic and Antipyretic Properties of Clitoria ternatea Root. Fitoterapia 2003, 74, 345–349. [Google Scholar] [CrossRef]
- Singh, N.K.; Garabadu, D.; Sharma, P.; Shrivastava, S.K.; Mishra, P. Anti-Allergy and Anti-Tussive Activity of Clitoria ternatea L. in Experimental Animals. J. Ethnopharmacol. 2018, 224, 15–26. [Google Scholar] [CrossRef]
- Sreekala, S.; Muraleedharan, U.D. Cationic Clitoria ternatea Seed Peptide as a Potential Novel Bioactive Molecule. Protein Pept. Lett. 2021, 28, 1259–1271. [Google Scholar] [CrossRef]
- Thell, K.; Hellinger, R.; Sahin, E.; Michenthaler, P.; Gold-Binder, M.; Haider, T.; Kuttke, M.; Liutkevičiūtė, Z.; Göransson, U.; Gründemann, C.; et al. Oral Activity of a Nature-Derived Cyclic Peptide for the Treatment of Multiple Sclerosis. Proc. Natl. Acad. Sci. USA 2016, 113, 3960–3965. [Google Scholar] [CrossRef]
- Harada, G.; Onoue, S.; Inoue, C.; Hanada, S.; Katakura, Y. Delphinidin-3-Glucoside Suppresses Lipid Accumulation in HepG2 Cells. Cytotechnology 2018, 70, 1707–1712. [Google Scholar] [CrossRef] [PubMed]
- Jacob, L.; Latha, M. Anticancer Activity of Clitoria ternatea Linn. against Dalton’s Lymphoma. Int. J. Pharmacogn. Phytochem. Res. 2012, 4, 207–212. [Google Scholar]
- Neda, G.D.; Mohd Salleh, R.; Ong, M.T. Chemical Composition and Anti-Proliferative Properties of Flowers of Clitoria ternatea. Int. Food Res. J. 2013, 20, 1229–1234. [Google Scholar]
- Escher, G.B.; Marques, M.B.; do Carmo, M.A.V.; Azevedo, L.; Furtado, M.M.; Sant’Ana, A.S.; da Silva, M.C.; Genovese, M.I.; Wen, M.; Zhang, L.; et al. Clitoria ternatea L. Petal Bioactive Compounds Display Antioxidant, Antihemolytic and Antihypertensive Effects, Inhibit α-Amylase and α-Glucosidase Activities and Reduce Human LDL Cholesterol and DNA Induced Oxidation. Food Res. Int. 2020, 128, 108763. [Google Scholar] [CrossRef]
- Fu, X.; Wu, Q.; Wang, J.; Chen, Y.; Zhu, G.; Zhu, Z. Spectral Characteristic, Storage Stability and Antioxidant Properties of Anthocyanin Extracts from Flowers of Butterfly Pea (Clitoria ternatea L.). Molecules 2021, 26, 7000. [Google Scholar] [CrossRef]
- López Prado, A.S.; Shen, Y.; Ardoin, R.; Osorio, L.F.; Cardona, J.; Xu, Z.; Prinyawiwatkul, W. Effects of Different Solvents on Total Phenolic and Total Anthocyanin Contents of Clitoria ternatea L. Petal and Their Anti-Cholesterol Oxidation Capabilities. Int. J. Food Sci. Technol. 2019, 54, 424–431. [Google Scholar] [CrossRef]
- Mus, A.A.; Goh, L.P.W.; Marbawi, H.; Gansau, J.A. The Biosynthesis and Medicinal Properties of Taraxerol. Biomedicines 2022, 10, 807. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Shi, H.; Ji, G.; Xie, J. Effects of taraxerol and taraxerol acetate on cell cycle and apoptosis of human gastric epithelial cell line AGS. Zhong Xi Yi Jie He Xue Bao 2011, 9, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Csupor-Löffler, B.; Hajdú, Z.; Zupkó, I.; Molnár, J.; Forgo, P.; Vasas, A.; Kele, Z.; Hohmann, J. Antiproliferative Constituents of the Roots of Conyza Canadensis. Planta Med. 2011, 77, 1183–1188. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.S.; Rout, K.K.; Chand, P.K. Production of Triterpenoid Anti-Cancer Compound Taraxerol in Agrobacterium-Transformed Root Cultures of Butterfly Pea (Clitoria ternatea L.). Appl. Biochem. Biotechnol. 2012, 168, 487–503. [Google Scholar] [CrossRef]
- Lakshan, S.A.T.; Jayanath, N.Y.; Abeysekera, W.P.K.M.; Abeysekera, W.K.S.M. A Commercial Potential Blue Pea (Clitoria ternatea L.) Flower Extract Incorporated Beverage Having Functional Properties. Evid. Based Complement. Altern. Med. 2019, 2019, 2916914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaafar, N.F.; Ramli, M.E.; Mohd Salleh, R. Optimum Extraction Condition of Clitorea ternatea Flower on Antioxidant Activities, Total Phenolic, Total Flavonoid and Total Anthocyanin Contents. Trop Life Sci. Res. 2020, 31, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Akter, R.; Uddin, S.J.; Grice, I.D.; Tiralongo, E. Cytotoxic Activity Screening of Bangladeshi Medicinal Plant Extracts. J. Nat. Med. 2014, 68, 246–252. [Google Scholar] [CrossRef]
- Deepika, S.; Selvaraj, C.I.; Anbalagan, M. Phytochemical Characterization and Cancer Cell Line Cytotoxicity of Clitoria ternatea. Bangladesh J. Pharmacol. 2018, 13, 349–352. [Google Scholar] [CrossRef]
- Zakaria, N.N.A.; Okello, E.J.; Howes, M.-J.; Birch-Machin, M.A.; Bowman, A. In Vitro Protective Effects of an Aqueous Extract of Clitoria ternatea L. Flower against Hydrogen Peroxide-Induced Cytotoxicity and UV-Induced MtDNA Damage in Human Keratinocytes. Phytother. Res. 2018, 32, 1064–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Silva, A.; Tavares-Carreón, F.; Figueroa, M.; De la Torre-Zavala, S.; Gastelum-Arellanez, A.; Rodríguez-García, A.; Galán-Wong, L.J.; Avilés-Arnaut, H. Anticancer Potential of Thevetia Peruviana Fruit Methanolic Extract. BMC Complement. Altern. Med. 2017, 17, 241. [Google Scholar] [CrossRef] [Green Version]
- Mat Zian, N.F.A.; Swain, P.; Mohd Faudzi, S.M.; Zakaria, N.; Wan Ibrahim, W.N.; Abu Bakar, N.; Shaari, K.; Stanslas, J.; Choi, T.-I.; Kim, C.-H. Mapping Molecular Networks within Clitoria ternatea Linn. against LPS-Induced Neuroinflammation in Microglial Cells, with Molecular Docking and In Vivo Toxicity Assessment in Zebrafish. Pharmaceuticals 2022, 15, 467. [Google Scholar] [CrossRef]
- Shen, Y.; Du, L.; Zeng, H.; Zhang, X.; Prinyawiwatkul, W.; Alonso-Marenco, J.; Xu, Z. Butterfly Pea (Clitoria ternatea) Seed and Petal Extracts Decreased HEp-2 Carcinoma Cell Viability. Int. J. Food Sci. Technol. 2016, 51, 1860–1868. [Google Scholar] [CrossRef]
- Asysyifa, A.; Agustiningtyas, A.; Nurgina, A.I. Butterfly Pea (Clitoria ternatea Linn.) Flower Extract Prevents MCF-7 HER2-Positive Breast Cancer Cell Metastasis in-Vitro. Ann. Oncol. 2020, 31, S1266. [Google Scholar] [CrossRef]
- Weber, G. Down-Regulation of Increased Signal Transduction Capacity in Human Cancer Cells. Adv. Enzym. Regul. 2005, 45, 37–51. [Google Scholar] [CrossRef]
- Lee, J.-H.; Jeong, Y.-J.; Lee, S.-W.; Kim, D.; Oh, S.-J.; Lim, H.-S.; Oh, H.-K.; Kim, S.-H.; Kim, W.-J.; Jung, J.-Y. EGCG Induces Apoptosis in Human Laryngeal Epidermoid Carcinoma Hep2 Cells via Mitochondria with the Release of Apoptosis-Inducing Factor and Endonuclease G. Cancer Lett. 2010, 290, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Blayney, A.; Liu, X.; Gandy, L.; Jin, W.; Yan, L.; Ha, J.-H.; Canning, A.J.; Connelly, M.; Yang, C.; et al. EGCG Binds Intrinsically Disordered N-Terminal Domain of P53 and Disrupts P53-MDM2 Interaction. Nat. Commun. 2021, 12, 986. [Google Scholar] [CrossRef] [PubMed]
- Hazafa, A.; Rehman, K.-U.-; Jahan, N.; Jabeen, Z. The Role of Polyphenol (Flavonoids) Compounds in the Treatment of Cancer Cells. Nutr. Cancer 2020, 72, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Chayaratanasin, P.; Caobi, A.; Suparpprom, C.; Saenset, S.; Pasukamonset, P.; Suanpairintr, N.; Barbieri, M.A.; Adisakwattana, S. Clitoria ternatea Flower Petal Extract Inhibits Adipogenesis and Lipid Accumulation in 3T3-L1 Preadipocytes by Downregulating Adipogenic Gene Expression. Molecules 2019, 24, 1894. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Zhang, C.; Feng, J.B.; Zhao, Y.X.; Wang, X.P.; Yang, J.M.; Zhang, M.X.; Wang, X.L.; Zhang, Y. Cross Talk among Smad, MAPK, and Integrin Signaling Pathways Enhances Adventitial Fibroblast Functions Activated by Transforming Growth Factor-Beta1 and Inhibited by Gax. Arter. Thromb. Vasc. Biol. 2008, 28, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Hong, G.; Wang, S.; Gao, W.; Wang, P. Tumor-Derived Exosomal MiRNA-141 Promote Angiogenesis and Malignant Progression of Lung Cancer by Targeting Growth Arrest-Specific Homeobox Gene (GAX). Bioengineered 2021, 12, 821–831. [Google Scholar] [CrossRef]
- Anguiano-Hernandez, Y.M.; Chartier, A.; Huerta, S. Smac/DIABLO and Colon Cancer. Anticancer Agents Med. Chem. 2007, 7, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Zhou, J.; Zhu, G.; Zhao, D.; He, F.; Zhang, J.; Lu, Y.; Yu, T.; Liu, L.; Wang, Y. Sublytic C5b-9 Triggers Glomerular Mesangial Cell Apoptosis via XAF1 Gene Activation Mediated by P300-Dependent IRF-1 Acetylation. Cell Death Dis. 2014, 5, e1176. [Google Scholar] [CrossRef] [Green Version]
- Üyetürk, U.; Ocak, Z.; Acar, M.; Özbey, I.; Tekelioğlu, V. The Effects of Salidrosid on DIABLO and XAF1 Gene Expression in PC3 Prostat Cancer Cells. Acta Med. Anatolia 2014, 2, 133. [Google Scholar] [CrossRef]
- Espinosa, M.; Lizárraga, F.; Vázquez-Santillán, K.; Hidalgo-Miranda, A.; Piña-Sánchez, P.; Torres, J.; García-Ramírez, R.A.; Maldonado, V.; Melendez-Zajgla, J.; Ceballos-Cancino, G. Coexpression of Smac/DIABLO and Estrogen Receptor in Breast Cancer. Cancer Biomark. 2021, 30, 429–446. [Google Scholar] [CrossRef]
- Xu, X.; Wu, X.; Jiang, Q.; Sun, Y.; Liu, H.; Chen, R.; Wu, S. Downregulation of MicroRNA-1 and MicroRNA-145 Contributes Synergistically to the Development of Colon Cancer. Int. J. Mol. Med. 2015, 36, 1630–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, C.; Wang, R.; Kirkby, M.; Man, S.M. Molecular Mechanisms Activating the NAIP-NLRC4 Inflammasome: Implications in Infectious Disease, Autoinflammation, and Cancer. Immunol. Rev. 2020, 297, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.C.; Nayak, A.K.; Dhara, A.K. Herbal Biomolecules in Healthcare Applications; Academic Press: London, UK, 2022; ISBN 978-0-323-90080-5. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of Total Flavonoid Content in Propolis by Two Complementary Colometric Methods. J. Food Drug Anal. 2020, 10, 3. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on Products of Browning Reaction Prepared from Glucose Amine Products Derived from Bees. J. Pharm. Biomed. Anal. 1986, 41, 1220–1234. [Google Scholar]
- Puranik, S.I.; Hiremath, M.; Nerli, R.B.; Ghagane, S.C. Evaluation of in Vitro Antioxidant and Anticancer Activity of Tabernaemontana divaricata Leaf Extracts Against T-24 Human Bladder Cancer Cell Lines. Int. J. Cancer Res. 2018, 14, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Ansari, A.A.; Khan, A.A.; Abdulla, M.; Al-Obaid, O.; Ahmad, R. In Vitro Evaluation of Cytotoxicity, Possible Alteration of Apoptotic Regulatory Proteins, and Antibacterial Activity of Synthesized Copper Oxide Nanoparticles. Colloids Surf. B Biointerfaces 2017, 153, 320–326. [Google Scholar] [CrossRef]
- Morón-Calvente, V.; Romero-Pinedo, S.; Toribio-Castelló, S.; Plaza-Díaz, J.; Abadía-Molina, A.C.; Rojas-Barros, D.I.; Beug, S.T.; LaCasse, E.C.; MacKenzie, A.; Korneluk, R.; et al. Inhibitor of Apoptosis Proteins, NAIP, CIAP1 and CIAP2 Expression during Macrophage Differentiation and M1/M2 Polarization. PLoS ONE 2018, 13, e0193643. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ser | Cytotoxicity IC50 µg/mL | ||||
|---|---|---|---|---|---|
| Code | MCF-7 | HCT116 | TT | WI38 | |
| 1 | BTL | 61.6 ± 3.32 | 29.2 ± 1.57 | 55.7 ± 3 | 124 ± 6.67 |
| 2 | Staurosporine | 12.7 ± 0.68 | 6.99 ± 0.38 | 7.39 ± 0.4 | 30.4 ± 1.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
ALshamrani, S.M.; Safhi, F.A.; Mobasher, M.A.; Saleem, R.M.; Alharthi, A.; Alshaya, D.S.; Awad, N.S. Antiproliferative Effect of Clitoria ternatea Ethanolic Extract against Colorectal, Breast, and Medullary Thyroid Cancer Cell Lines. Separations 2022, 9, 331. https://doi.org/10.3390/separations9110331
ALshamrani SM, Safhi FA, Mobasher MA, Saleem RM, Alharthi A, Alshaya DS, Awad NS. Antiproliferative Effect of Clitoria ternatea Ethanolic Extract against Colorectal, Breast, and Medullary Thyroid Cancer Cell Lines. Separations. 2022; 9(11):331. https://doi.org/10.3390/separations9110331
Chicago/Turabian StyleALshamrani, Salha M., Fatmah A. Safhi, Maysa A. Mobasher, Rasha M. Saleem, Afaf Alharthi, Dalal Sulaiman Alshaya, and Nabil S. Awad. 2022. "Antiproliferative Effect of Clitoria ternatea Ethanolic Extract against Colorectal, Breast, and Medullary Thyroid Cancer Cell Lines" Separations 9, no. 11: 331. https://doi.org/10.3390/separations9110331
APA StyleALshamrani, S. M., Safhi, F. A., Mobasher, M. A., Saleem, R. M., Alharthi, A., Alshaya, D. S., & Awad, N. S. (2022). Antiproliferative Effect of Clitoria ternatea Ethanolic Extract against Colorectal, Breast, and Medullary Thyroid Cancer Cell Lines. Separations, 9(11), 331. https://doi.org/10.3390/separations9110331