


Unravelling the In Vitro and In Vivo Anti-Helicobacter pylori Effect of Delphinidin-3-O-Glucoside Rich Extract from Pomegranate Exocarp: Enhancing Autophagy and Downregulating TNF-α and COX2

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Extraction and Characterization
2.1.1. Plant Material and Extraction
2.1.2. Characterization of the Contents of PEAME Extract
Determination of Total Polyphenolic Content
Determination of Anthocyanin Content
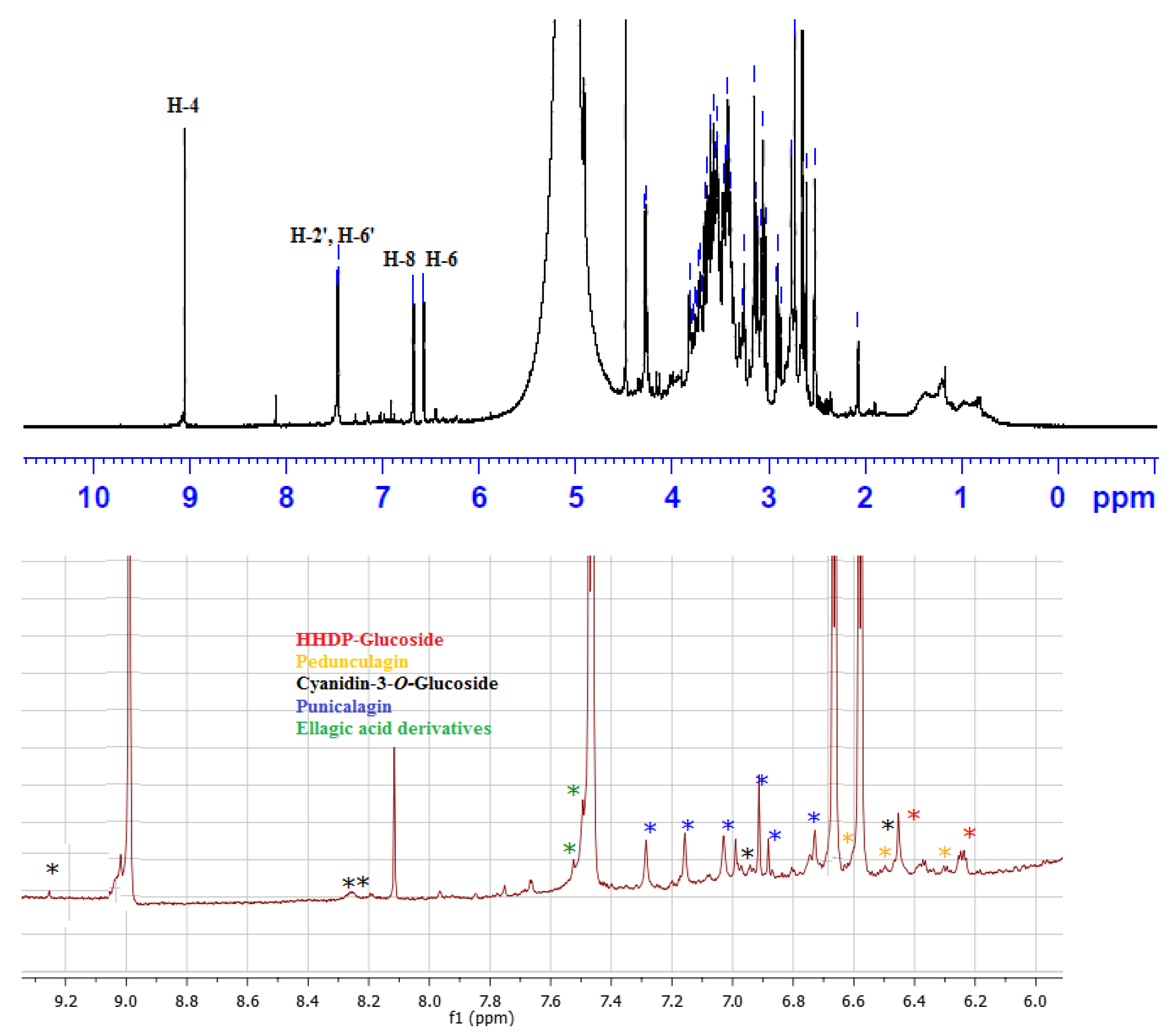
Characterization of Pomegranate Exocarp Anthocyanin Methanol Extract (PEAME) by UPLC-PDA-MS/MS
1H NMR Fingerprint of PEAME
2.2. Ethical Statement
2.3. In Vitro Anti-H. pylori Effect of PEAME
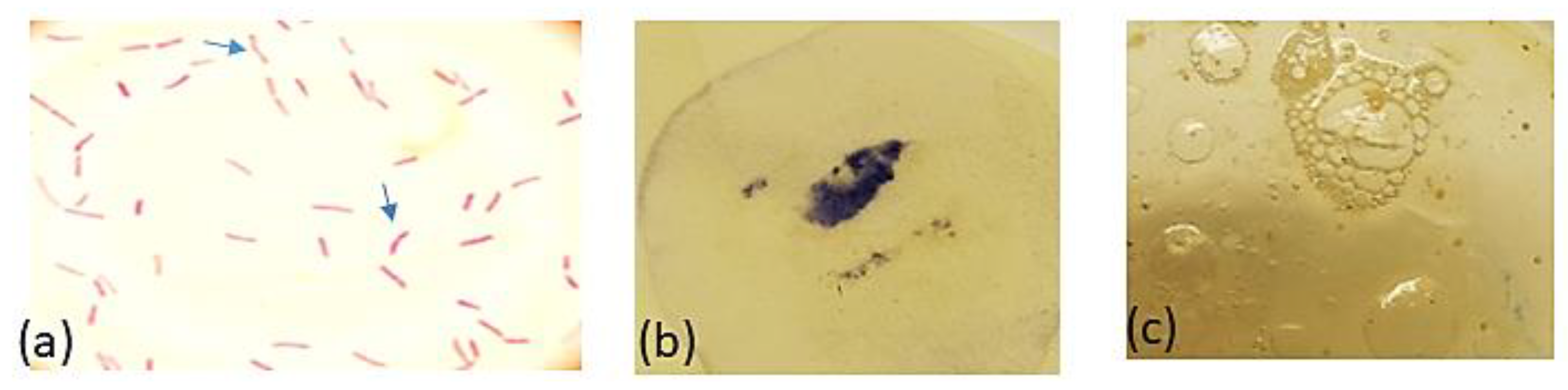
2.3.1. Collection of Gastric Biopsies and Isolation and Identification of the Test Pathogens
2.3.2. Susceptibility Test
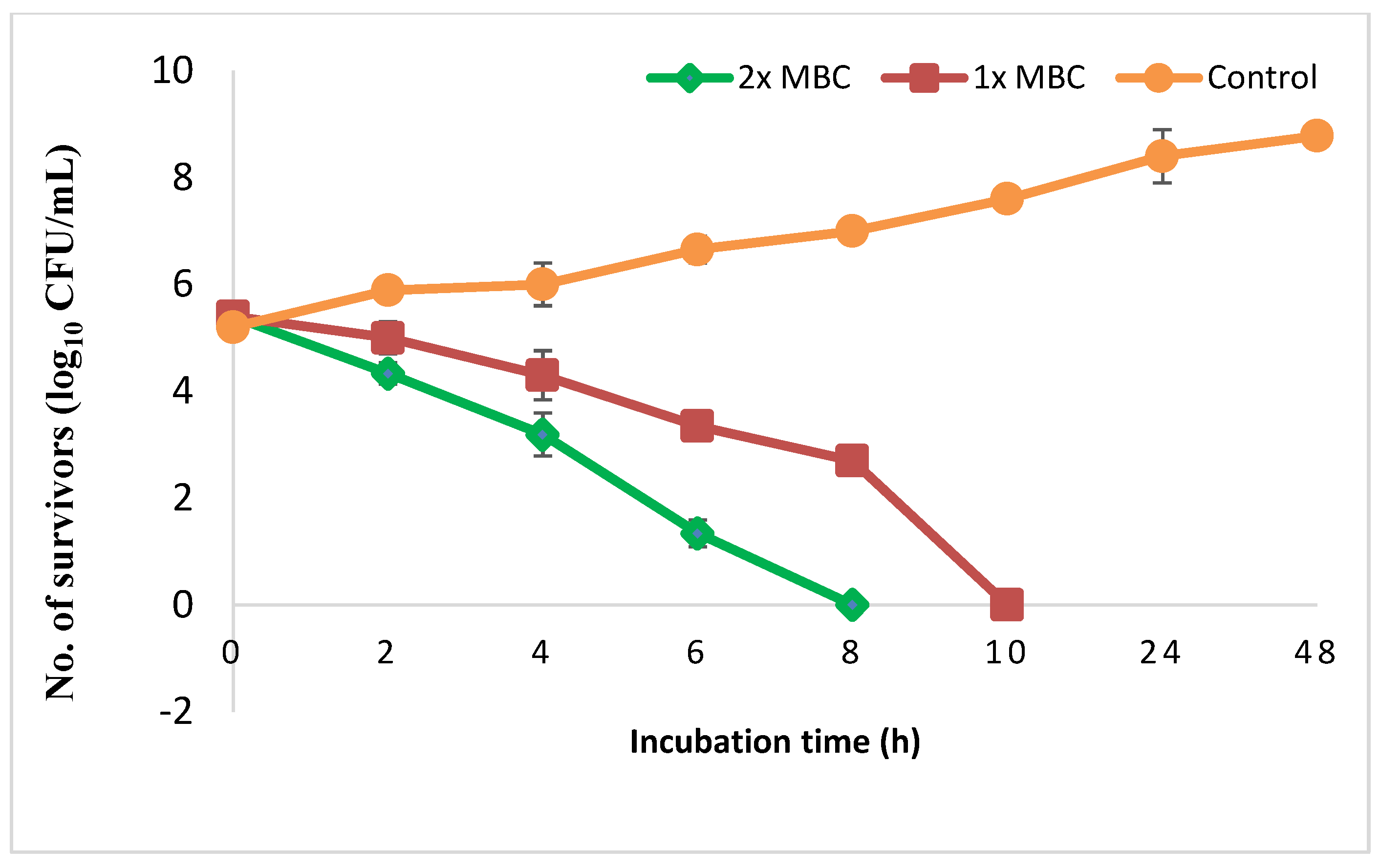
2.3.3. Time-Kill Assay
2.4. Studying the Effect of PEAME on H. pylori Infection In Vivo
2.4.1. Animals
2.4.2. Determination of Oxidative Stress and Antioxidant Markers
Markers of Lipid Peroxidation
Catalase Activity
2.4.3. Assessing the Expression of TNF-α and Autophagy Genes Using Quantitative Real Time-Polymerase Chain Reaction (RT-qPCR)
2.4.4. Histopathological Evaluation
2.4.5. Immunohistochemical Staining of Cyclooxygenase 2 (COX2)
2.4.6. Bacteriological Analysis
2.4.7. Statistical Analysis
3. Results
3.1. Characterization of Polyphenolics in PEAME
3.2. In Vitro Anti-H. pylori Effect of PEAME
3.3. In Vivo Anti-H. pylori Effect of PEAME
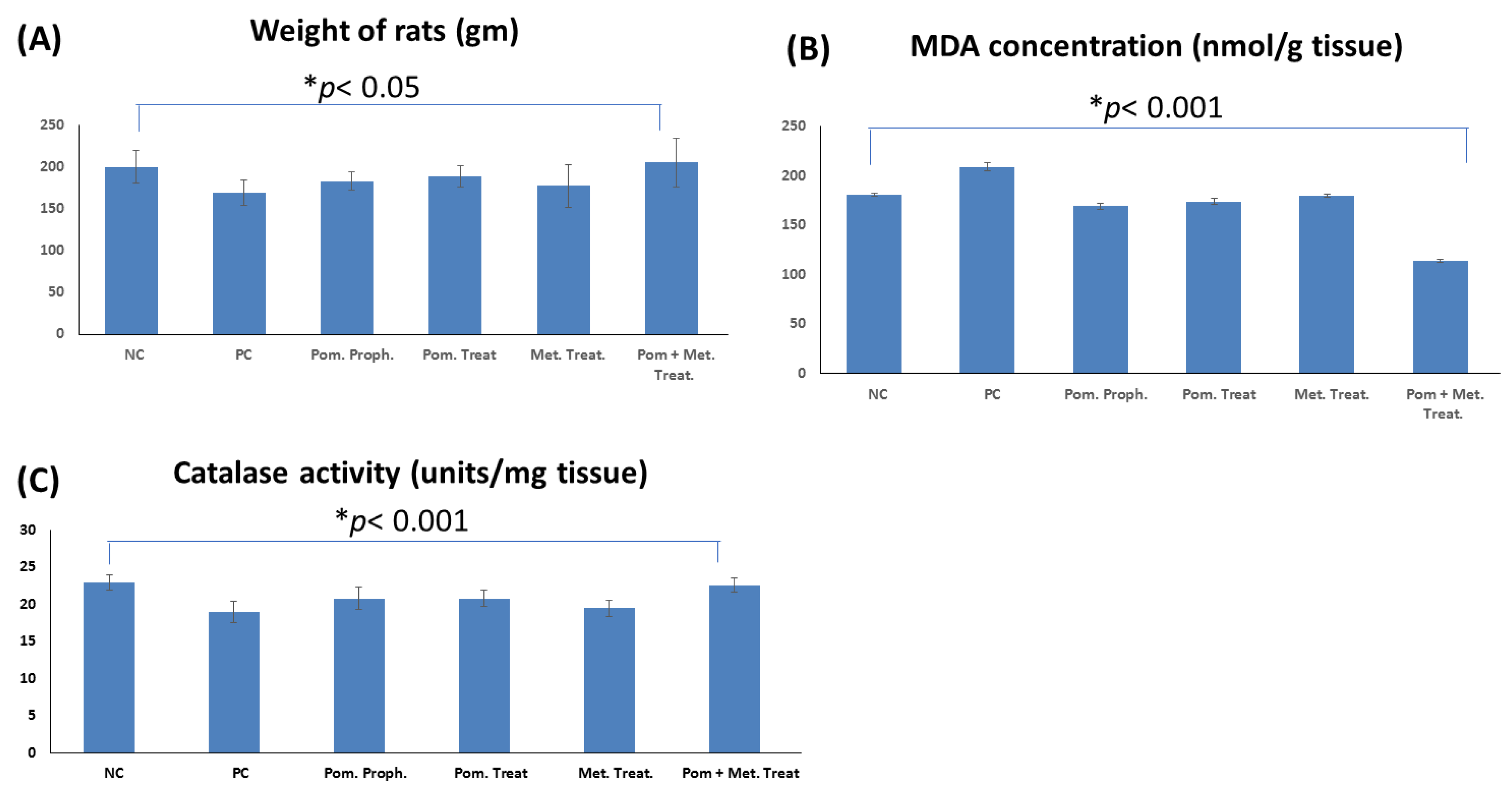
3.3.1. Effect on the Weight of the Rats
3.3.2. Effect on the Colonization of H. pylori in the Stomach of the Infected Rats
3.3.3. Effect on the Lipid Peroxidation of the Stomach Tissue in the Rats Infected with H. pylori
3.3.4. Effect on the Catalase Activity in the Stomach Tissue of the Rats Infected with H. pylori
3.3.5. Effect on the Gene Expression of TNF-α in the Stomach Tissue of the Rats Infected with H. pylori
3.3.6. Effect on the Expression of Autophagy Genes in the Stomach Tissue of the Rats Infected with H. pylori
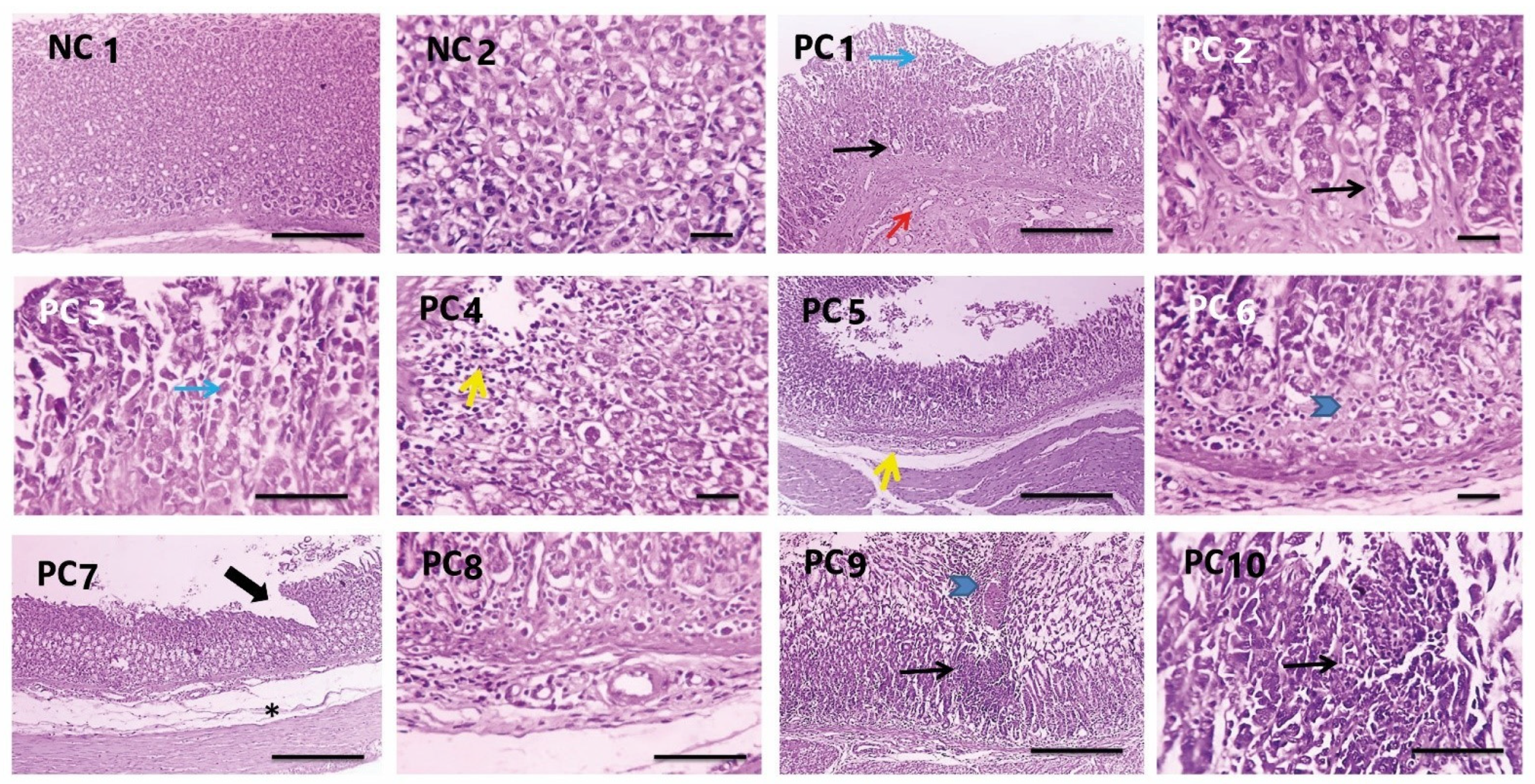
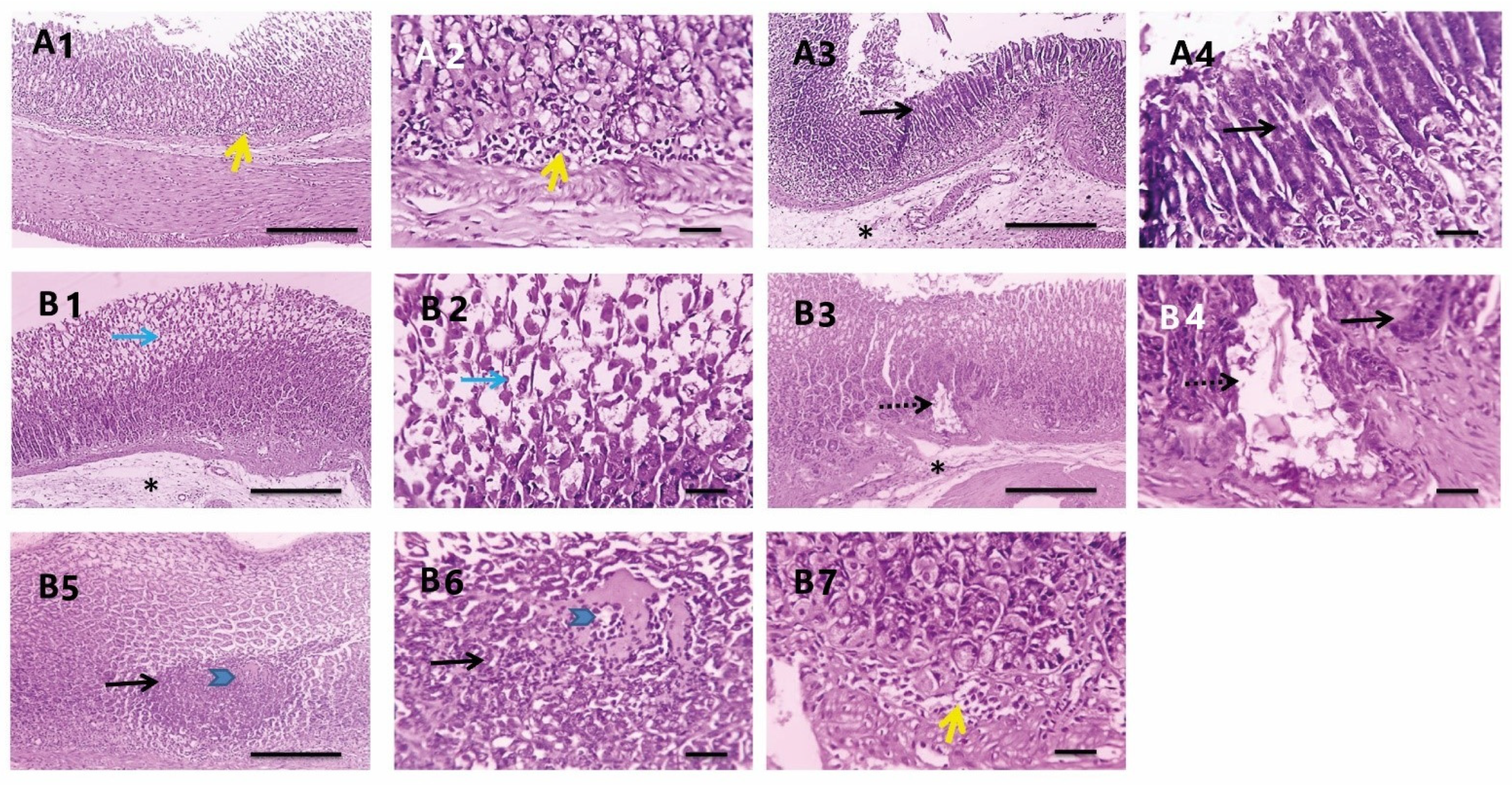
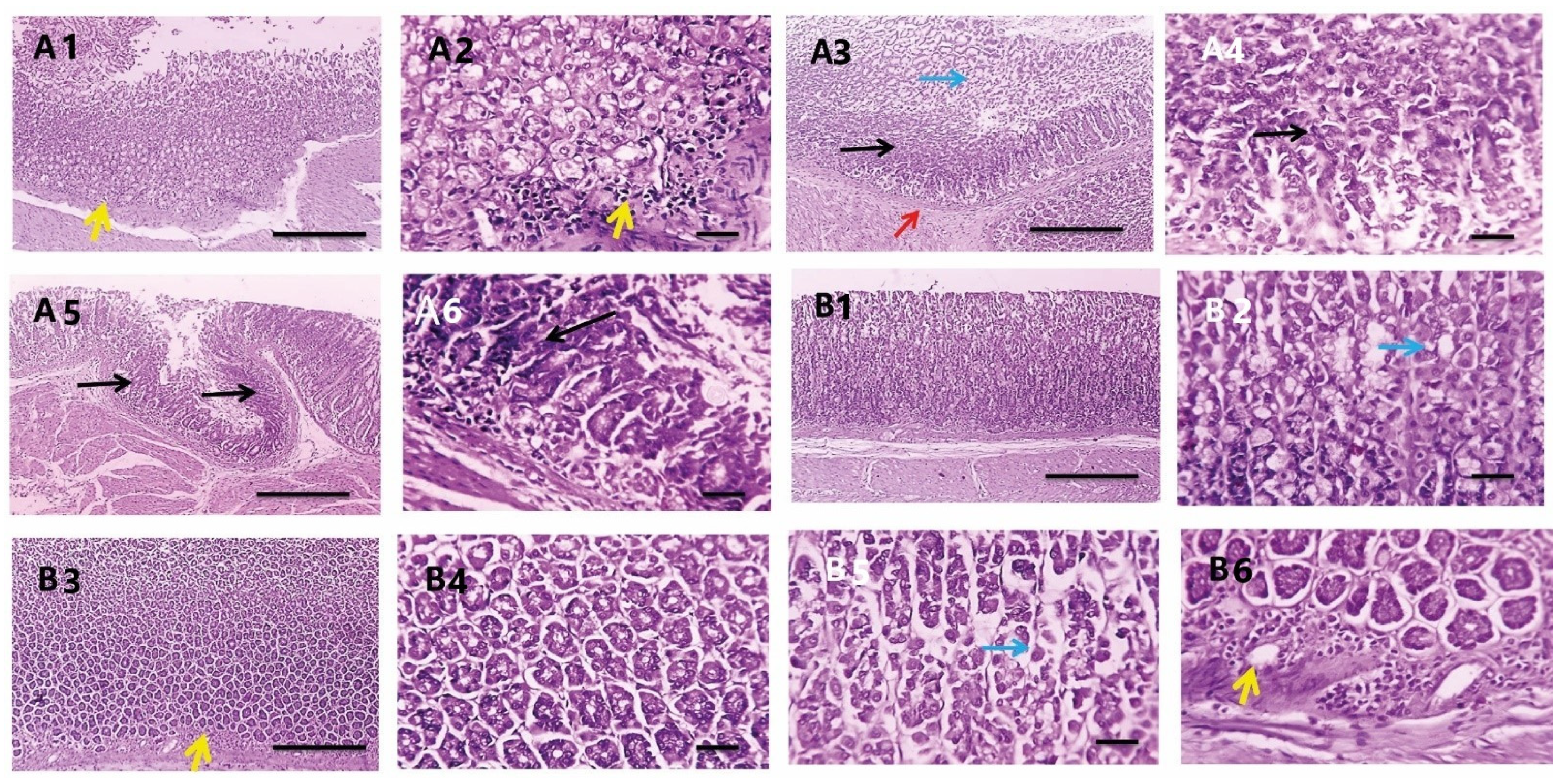
3.3.7. Histopathological Investigations
3.3.8. Effect on the COX2 Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, R.H. Health-promoting components of fruits and vegetables in the diet. Adv. Nutr. 2013, 4, 384S–392S. [Google Scholar] [CrossRef] [PubMed]
- Suriyaprom, S.; Mosoni, P.; Leroy, S.; Kaewkod, T.; Desvaux, M.; Tragoolpua, Y. Antioxidants of fruit extracts as antimicrobial agents against pathogenic bacteria. Antioxidants 2022, 11, 602. [Google Scholar] [CrossRef] [PubMed]
- De Francescoa, V.; Zullob, A.; Mantac, R.; Satrianod, A.; Fiorinie, G.; Pavonie, M.; Saracinoe, I.M.; Giostraf, F.; Montif, G.; Vairae, D. Culture-based antibiotic susceptibility testing for Helicobacter pylori infection: A systematic review. Ann. Gastroenterol. 2022, 35, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Ogata, S.K.; Gales, A.C.; Kawakam, E. Antimicrobial susceptibility testing for Helicobacter pylori isolates from Brazilian children and adolescents: Comparing agar dilution, E-test, and disk diffusion. Braz. J. Microbiol. 2014, 45, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Matos, I.A.; Oliva, S.E.D.; Escobedo, A.A.; Jiménez, O.M.V.; Villaurrutia, Y.D.C.V. Helicobacter pylori infection in children. BMJ Paediatr. Open 2020, 4, e000679. [Google Scholar]
- El-Deeb, N.; Al-Madboly, L. Probiotics, the natural microbiota in living organisms fundamentals and applications. In Probiotics and GIT Diseases/Stomach Ulcer, 1st ed.; Taylor & Francis Group, LLC: Abingdon, UK, 2021. [Google Scholar]
- Corthesy, B.; Boris, S.; Isler, P.; Grangette, C.; Mercenier, A. Oral immunization of mice with lactic acid bacteria producing Helicobacter pylori urease B subunit partially protects against challenge with Helicobacter felis. J. Infect. Dis. 2005, 192, 1441–1449. [Google Scholar] [CrossRef]
- Chung, C.; Seo, W.; Silwal, P.; Jo, E.K. Crosstalks between inflammasome and autophagy in cancer. J. Hematol. Oncol. 2020, 13, 100. [Google Scholar] [CrossRef]
- Tanaka, S.; Nagashima, H.; Uotani, T.; Graham, D.Y.; Yamaoka, Y. Autophagy-related genes in Helicobacter pylori infection. Helicobacter 2017, 22, e12376. [Google Scholar] [CrossRef]
- Alam, M.S.; Alam, M.A.; Ahmad, S.; Najmi, A.K.; Asif, M.; Jahangir, T. Protective effects of Punica granatum in experimentally-induced gastric ulcers. Toxicol. Mech. Methods 2010, 20, 572–578. [Google Scholar]
- Moghaddam, G.; Sharifzadeh, M.; Hassanzadeh, G.; Khanavi, M.; Hajimahmoodi, M. Anti-ulcerogenic activity of the pomegranate peel (Punica granatum) methanol extract. Food Nutr. Sci. 2013, 4, 43. [Google Scholar]
- Nuraddin, S.M.; Amin, Z.A.; Sofi, S.H.; Osman, S. Antibacterial and anti-ulcerogenic effects of Punica granatum peel extract against ethanol-induced acute gastric lesion in rats. Zanco J. Med. Sci. 2019, 23, 308–314. [Google Scholar] [CrossRef]
- Lansky, E.P.; Newman, R.A. Punica granatum (pomegranate) and its potential for prevention and treatment of inflammation and cancer. J. Ethnopharmacol. 2007, 109, 177–206. [Google Scholar] [PubMed]
- Subkorn, P.; Norkaew, C.; Deesrisak, K.; Tanyong, D. Punicalagin, a pomegranate compound, induces apoptosis and autophagy in acute leukemia. PeerJ 2021, 9, e12303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Halemahebai, G.; Tian, L.; Dong, H.; Aisker, G. Urolithin A, a pomegranate metabolite, protects pancreatic β cells from apoptosis by activating autophagy. J. Ethnopharmacol. 2021, 272, 113628. [Google Scholar] [CrossRef] [PubMed]
- Mayyas, A.; Abu-Sini, M.; Amr, R.; Akasheh, R.T.; Zalloum, W.; Khdair, A.; Hamad, I.; Aburjai, T.; Darwish, R.M.; Abu-Qatouseh, L. Novel in vitro and in vivo anti-Helicobacter pylori effects of pomegranate peel ethanol extract. Vet. World 2021, 14, 120–128. [Google Scholar] [CrossRef]
- Attard, E.A. rapid microtitre plate Folin-Ciocalteu method for the assessment of polyphenols. Cent. Eur. J. Biol. 2013, 8, 48–53. [Google Scholar]
- Lee, J.; Durst, R.W.; Wrolstad, R.E.; Collaborators: Eisele, T.; Giusti, M.M.; Hach, J.; Hofsommer, H.; Koswig, S.; Krueger, D.A.; Kupina, S.; et al. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: Collaborative study. J. AOAC Int. 2005, 88, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Assar, D.H.; Mokhbatly, A.A.A.; Ghazy, E.W.; Ragab, A.E.; Abou Asa, S.; Abdo, W.; Elbialy, Z.I.; Mohamed, N.E.; El-Far, A.H. Ameliorative effects of Aspergillus awamori against the initiation of hepatocarcinogenesis induced by diethylnitrosamine in a rat model: Regulation of Cyp19 and p53 gene expression. Antioxidants 2021, 10, 922. [Google Scholar] [CrossRef] [PubMed]
- Assar, D.H.; Elhabashi, N.; Mokhbatly, A.A.A.; Ragab, A.E.; Elbialy, Z.I.; Rizk, S.A.; Albalawi, A.E.; Althobaiti, N.A.; Al Jaouni, S.; Atiba, A. Wound healing potential of licorice extract in rat model: Antioxidants, histopathological, immunohistochemical and gene expression evidences. Biomed. Pharmacother. 2021, 143, 112151. [Google Scholar] [CrossRef]
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Family II Helicobacteraceae fam. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Springer: New York, NY, USA, 2005; Volume 2. [Google Scholar]
- Clinical and Laboratory Standards Institute CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Approved standard M7-A5; Informational supplement M100-S10; NCCLS: Wayne, PA, USA, 2000. [Google Scholar]
- Ko, J.K.; Ching, C.K.; Chow, J.Y.; Zhang, S.T.; Lam, S.K.; Cho, C.H. The vascular and glandular organoprotective properties of metronidazole in the rodent stomach. Aliment. Pharmacol. Ther. 1997, 11, 811–819. [Google Scholar] [CrossRef]
- Ganapaty, S.; Chandrashekhar, V.M.; Lakshmi, N.M.; Raghavendra, H.L. Antioxidant activity of natural products against aluminium fluoride induced oxidative stress. Sci. Technol. Arts Res. J. 2012, 1, 26–37. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Aebi, H.E. Catalase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie: Weinhem, Germany, 1983; pp. 273–286. [Google Scholar]
- Li, Q.; Cai, M.; Wang, J.; Gao, Q.; Guo, X.; Jia, X.; Xu, S.; Zhu, H. Decreased ovarian function and autophagy gene methylation in aging rats. J. Ovarian Res. 2020, 13, 1–9. [Google Scholar] [CrossRef]
- Khan, H.A.; Abdelhalim, M.A.; Alhomida, A.S.; Al Ayed, M.S. Transient increase in IL-1β, IL-6 and TNF-α gene expression in rat liver exposed to gold nanoparticles. Genet. Mol. Res. 2013, 12, 5851–5857. [Google Scholar] [CrossRef] [PubMed]
- Heid, C.A.; Stevens, J.; Livak, K.J.; Williams, P.M. Real time quantitative PCR. Genome Res. 1996, 6, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Sozzi, M.M.D.; Valentini, M.M.D.; Figura, N.M.D.; De Paoli, P.M.D.; Tedeschi, R.M.S.C.D.; Gloghini, A.S.C.D.; Serraino, D.M.D.; Poletti, M.M.D.; Carbone, A.M.D. Atrophic gastritis and intestinal metaplasia in Helicobacter pylori infection: The role of CagA status. Am. J. Gastroenterol. 1998, 93, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.V.; Feakins, R.; Farthing, M.J.; Ballinger, A. Cyclooxygenase 2, p53, beta-catenin, and APC protein expression in gastric adenomatous polyps. Am. J. Clin. Pathol. 2005, 123, 415–420. [Google Scholar] [CrossRef]
- Ku, N.S.; Lee, S.-H.; Lim, Y.-S.; Choi, H.; Ahn, J.Y.; Jeong, S.J.; Shin, S.J.; Choi, J.Y.; Choi, Y.H.; Yeom, J.-S.; et al. In vivo efficacy of combination of colistin with fosfomycin or minocycline in a mouse model of multidrug-resistant Acinetobacter baumannii pneumonia. Sci. Rep. 2019, 9, 17127. [Google Scholar] [CrossRef]
- Field, A. Discovering Statistics Using IBM SPSS Statistics; SAGE: Thousand Oaks, CA, USA, 2013. [Google Scholar]
- Abid, M.; Yaich, H.; Cheikhrouhou, S.; Khemakhem, I.; Bouaziz, M.; Attia, H.; Ayadi, M.A. Antioxidant properties and phenolic profile characterization by LC–MS/MS of selected Tunisian pomegranate peels. J. Food Sci. Technol. 2017, 54, 2890–2901. [Google Scholar] [CrossRef]
- Yisimayili, Z.; Abdulla, R.; Tian, Q.; Wang, Y.; Chen, M.; Sun, Z.; Li, Z.; Liu, F.; Aisa, H.A.; Huang, C. A comprehensive study of pomegranate flowers polyphenols and metabolites in rat biological samples by high-performance liquid chromatography quadrupole time-of-flight mass spectrometry. J. Chromatogr. A 2019, 1604, 460472. [Google Scholar] [CrossRef]
- García, P.; Fredes, C.; Cea, I.; Lozano-Sánchez, J.; Leyva-Jiménez, F.J.; Robert, P.; Vergara, C.; Jimenez, P. Recovery of bioactive compounds from pomegranate (Punica granatum L.) peel using pressurized liquid extraction. Foods 2021, 10, 203. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.L.; Liao, X.J.; Wang, Y.H.; Si, X.; Shu, C.; Gong, E.S.; Xie, X.; Ran, X.L.; Li, B. Identification of cyanidin-3-arabinoside extracted from blueberry as a selective protein tyrosine phosphatase 1B inhibitor. J. Agric. Food Chem. 2019, 67, 13624–13634. [Google Scholar] [CrossRef] [PubMed]
- Gabr, S.K.; Bakr, R.O.; Mostafa, E.S.; El-Fishawy, A.M.; El-Alfy, T.S. Antioxidant activity and molecular docking study of Erytherina × neillii polyphenolics. S. Afr. J. Bot. 2019, 121, 470–477. [Google Scholar] [CrossRef]
- Ahmed, S.R.; El-Sherei, M.M.; El-Dine, R.S.; El-Toumy, S.A. Phytoconstituents, hepatoprotective, and antioxidant activities of Euphorbia cooperi N. E. Br. Egypt. J. Chem. 2019, 62, 831–840. [Google Scholar]
- Hasanpour, M.; Saberi, S.; Iranshahi, M. Metabolic profiling and untargeted 1H-NMR -based metabolomics study of different Iranian pomegranate (Punica granatum) ecotypes. Planta Med. 2020, 86, 212–216. [Google Scholar] [CrossRef]
- Khanbabaee, K.; Großer, M. An efficient total synthesis of pedunculagin using a twofold intramolecular double esterification strategy. Eur. J. Org. Chem. 2003, 2003, 2128–2131. [Google Scholar] [CrossRef]
- Mohammad, M.A.; Hussein, L.; Coward, A.; Jackson, S.J. Prevalence of Helicobacter pylori infection among Egyptian children: Impact of social background and effect on growth. Public Health Nutr. 2008, 11, 230–236. [Google Scholar] [CrossRef]
- El-Sharouny, E.; El-Shazli, H.; Olama, Z. Detection of Helicobacter pylori DNA in some Egyptian water systems and its incidence of transmission to individuals. Iran. J. Public Health 2015, 44, 203–210. [Google Scholar]
- Ogata, S.K.; Godoy, A.P.O.; da Silva, P.F.R.; Kawakami, E. High Helicobacter pylori resistance to metronidazole and clarithromycin in Brazilian children and adolescents. J. Pediatr. Gastroenterol. Nutr. 2013, 56, 645–648. [Google Scholar] [CrossRef]
- Zhao, X.; Yuan, Z. Anthocyanins from pomegranate (Punica granatum L.) and their role in antioxidant capacities in vitro. Chem. Biodivers. 2021, 18, e2100399. [Google Scholar] [CrossRef]
- Burger, O.; Ofek, I.; Tabak, M.; Weiss, E.I.; Sharon, N.; Neeman, I. A high molecular mass constituent of cranberry juice inhibits Helicobacter pylori adhesion to human gastric mucus. FEMS Immunol. Med. Microbiol. 2000, 29, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Burger, O.; Weiss, E.; Sharon, N.; Tabak, M.; Neeman, I.; Ofek, I. Inhibition of Helicobacter pylori adhesion to human gastric mucus by a high-molecular-weight constituent of cranberry juice. Crit. Rev. Food Sci. Nutr. 2002, 42, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Park, M.; Woo, H.; Tharmalingam, N.; Lee, G.; Rhee, K.J.; Eom, Y.B.; Han, S.I.; Seo, W.D.; Kim, J.B. Inhibitory effects of anthocyanins on secretion of Helicobacter pylori CagA and VacA toxins. Int. J. Med. Sci. 2012, 9, 838. [Google Scholar] [CrossRef][Green Version]
- Parseh, H.; Shahablavasani, A. Comparing total anthocyanins, total phenolics and antioxidant activities of extracts (aqueous, organic and anthocyanin) obtained from pomegranate (peel, juice, and seed) and antimicrobial activity of peel extracts on the four pathogenic bacteria. J. Food Bioprocess Eng. 2019, 2, 7–12. [Google Scholar]
- Rodriguez-Saona, L.E.; Wrolstad, R.E. Extraction, isolation, and purification of anthocyanins. Curr. Protoc. Food Anal. Chem. 2001, 1, F1.1.1–F1.1.11. [Google Scholar] [CrossRef]
- Zhao, X.; Yuan, Z.; Fang, Y.; Yin, Y.; Feng, L. Characterization and evaluation of major anthocyanins in pomegranate (Punica granatum L.) peel of different cultivars and their development phases. Eur. Food Res. Technol. 2013, 236, 109–117. [Google Scholar] [CrossRef]
- Zhu, F.; Yuan, Z.; Zhao, X.; Yin, Y.; Feng, L. Composition and contents of anthocyanins in different pomegranate cultivars. Acta Hortic. 2015, 1089, 35–41. [Google Scholar]
- Calvino, F.M.; Parra, C.T. H. pylori and mitochondrial changes in epithelial cells. The role of oxidative stress. Rev. Esp. Enferm. Dig. 2010, 102, 41–50. [Google Scholar]
- Sayed, S.; Alotaibi, S.S.; El-Shehawi, A.M.; Hassan, M.M.; Shukry, M.; Alkafafy, M.; Soliman, M.M. The anti-Inflammatory, anti-Apoptotic, and antioxidant effects of a pomegranate-peel extract against acrylamide-induced hepatotoxicity in rats. Life 2022, 12, 224. [Google Scholar] [CrossRef]
- Szentmihályi, K.; Süle, K.; Egresi, A.; Blázovics, A.; May, Z. Metronidazole does not show direct antioxidant activity in in vitro global systems. Heliyon 2021, 7, e06902. [Google Scholar] [CrossRef]
- Zelová, H.; Hošek, J. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Jia, J. Natural COX-2 inhibitors as promising anti-inflammatory agents: An Update. Curr. Med. Chem. 2021, 28, 3622–3646. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Chan, H.; Lu, L.; Wong, K.T.; Wong, S.H.; Li, M.X.; Xiao, Z.G.; Cho, C.H.; Gin, T.; Chan, M.T.V.; et al. Autophagy in intracellular bacterial infection. Semin. Cell Dev. Biol. 2020, 101, 41–50. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | Rt min | [M-H]− m/z | MS2 Ions m/z | Identification |
|---|---|---|---|---|
| 1 | 4.25 | 783 | 481, 301, 275 | Pedunculagin I |
| 2 | 5.76 7.27 | 541 * 1083 | 541, 302 1083, 781, 603, 601, 575, 541, 302 | Punicalagin (α, β) |
| 3 | 6.75 | 783 | 765, 481, 301, 275 | Pedunculagin II |
| 4 | 7.84 | 801 | 649, 348, 347, 301 | Punigluconin |
| 5 | 8.12 | 463 | 302 | Delphenidin-3-O-glucoside |
| 6 | 8.43 | 469 | 425, 301, 169, 125 | Valoneic acid dilactone |
| 7 | 8.87 | 447 | 345, 259, 219, 160, 113 | Cyanidin-3-O-glucoside |
| 8 | 9.29 | 433 | 301 | Ellagic acid-O-pentoside |
| 9 | 11.06 | 301 | 301, 229, 185 | Ellagic acid |
| 10 | 12.50 | 481 | 301 | HHDP-glucoside |
| Test Parameter | Observation |
|---|---|
| Colony morphology | Round, small, translucent, 2–3 mm |
| Bacterial cell shape | Spiral, helical or curved with blunt ends |
| Gram staining reaction | Negative |
| Urease | Positive |
| Catalase | Positive |
| oxidase | Positive |
| Motility | Positive |
| Nitrate reduction | Negative |
| Glycine utilization | Negative |
| Growth on blood agar | Positive |
| Antimicrobial Agents | Resistance (%) | MIC Range (µg/mL) * |
|---|---|---|
| Metronidazole | 53 | 0.5–64 |
| Clarithromycin | 41 | 0.5–16 |
| Amoxicillin | 38 | 0.25–64 |
| Group | Test Groups | Log No. of Survivors ± SD |
|---|---|---|
| 1 | Negative control | 0 ± 0 * |
| 2 | Positive control | 4.11 ± 0.22 |
| 3 | Pomegranate Prophylaxis | 3.71 ± 0.23 |
| 4 | Treatment with pomegranate | 3.62 ± 0.14 |
| 5 | Treatment with metronidazole | 3.54 ± 0.09 |
| 6 | Treatment combination of pomegranate and metronidazole | 1.7 ± 0.12 |
| Scores 0–3 | NC | PC | PEAME Proph. | PEAME Treat. | Met Treat. | PEAME + Met Treat |
|---|---|---|---|---|---|---|
| Inflammation | 0 | 3 | 1 | 1 | 2 | 0 |
| 0 | 3 | 2 | 0 | 1 | 1 | |
| 0 | 2 | 1 | 1 | 2 | 1 | |
| 0 | 2 | 0 | 2 | 1 | 0 | |
| 0 | 2 | 1 | 2 | 1 | 1 | |
| 0 | 3 | 2 | 2 | 1 | 0 | |
| Metaplasia | 0 | 1 | 1 | 1 | 3 | 0 |
| 0 | 2 | 2 | 2 | 1 | 0 | |
| 0 | 2 | 2 | 1 | 2 | 0 | |
| 0 | 2 | 0 | 1 | 2 | 0 | |
| 0 | 2 | 1 | 0 | 0 | 0 | |
| 0 | 1 | 0 | 0 | 0 | 0 | |
| COX2 | 0 | 3 | 2 | 2 | 2 | 1 |
| 0 | 3 | 2 | 1 | 1 | 1 | |
| 0 | 3 | 2 | 2 | 3 | 1 | |
| 0 | 3 | 3 | 1 | 2 | 2 | |
| 0 | 3 | 2 | 2 | 2 | 2 | |
| 0 | 3 | 2 | 3 | 3 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragab, A.E.; Al-Madboly, L.A.; Al-Ashmawy, G.M.; Saber-Ayad, M.; Abo-Saif, M.A. Unravelling the In Vitro and In Vivo Anti-Helicobacter pylori Effect of Delphinidin-3-O-Glucoside Rich Extract from Pomegranate Exocarp: Enhancing Autophagy and Downregulating TNF-α and COX2. Antioxidants 2022, 11, 1752. https://doi.org/10.3390/antiox11091752
Ragab AE, Al-Madboly LA, Al-Ashmawy GM, Saber-Ayad M, Abo-Saif MA. Unravelling the In Vitro and In Vivo Anti-Helicobacter pylori Effect of Delphinidin-3-O-Glucoside Rich Extract from Pomegranate Exocarp: Enhancing Autophagy and Downregulating TNF-α and COX2. Antioxidants. 2022; 11(9):1752. https://doi.org/10.3390/antiox11091752
Chicago/Turabian StyleRagab, Amany E., Lamiaa A. Al-Madboly, Ghada M. Al-Ashmawy, Maha Saber-Ayad, and Mariam A. Abo-Saif. 2022. "Unravelling the In Vitro and In Vivo Anti-Helicobacter pylori Effect of Delphinidin-3-O-Glucoside Rich Extract from Pomegranate Exocarp: Enhancing Autophagy and Downregulating TNF-α and COX2" Antioxidants 11, no. 9: 1752. https://doi.org/10.3390/antiox11091752
APA StyleRagab, A. E., Al-Madboly, L. A., Al-Ashmawy, G. M., Saber-Ayad, M., & Abo-Saif, M. A. (2022). Unravelling the In Vitro and In Vivo Anti-Helicobacter pylori Effect of Delphinidin-3-O-Glucoside Rich Extract from Pomegranate Exocarp: Enhancing Autophagy and Downregulating TNF-α and COX2. Antioxidants, 11(9), 1752. https://doi.org/10.3390/antiox11091752