


Effects of Phloem-Feeding Pest, Dalbulus maidis on Morphological Expression of Drought-Tolerant Traits in Maize


Abstract
:1. Introduction
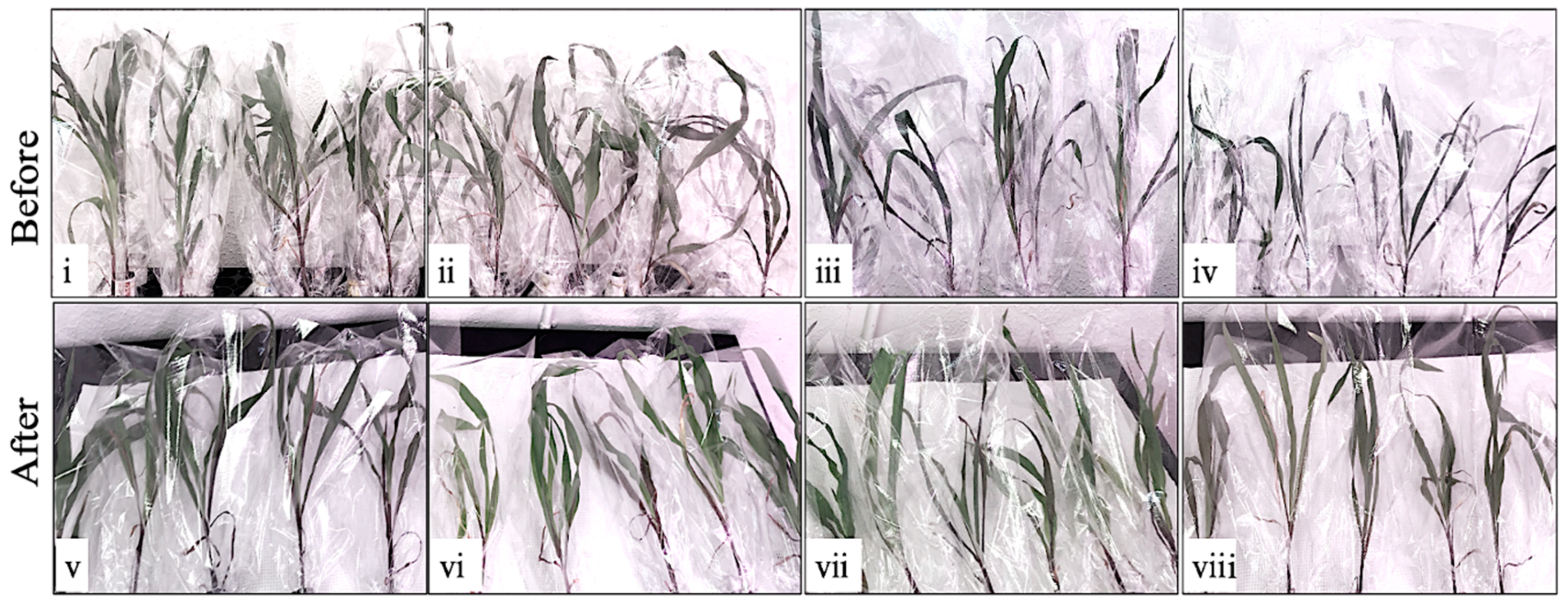
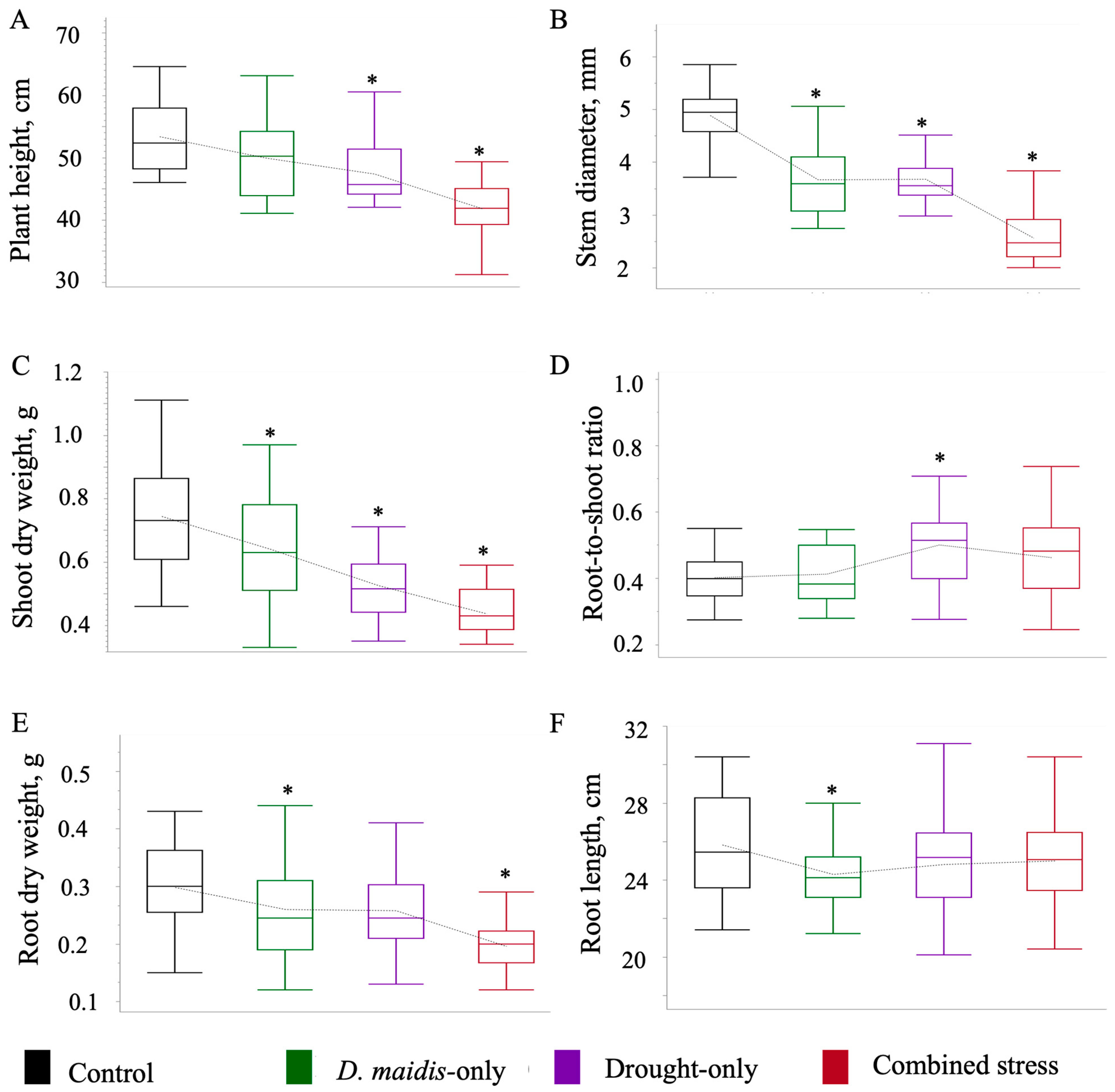
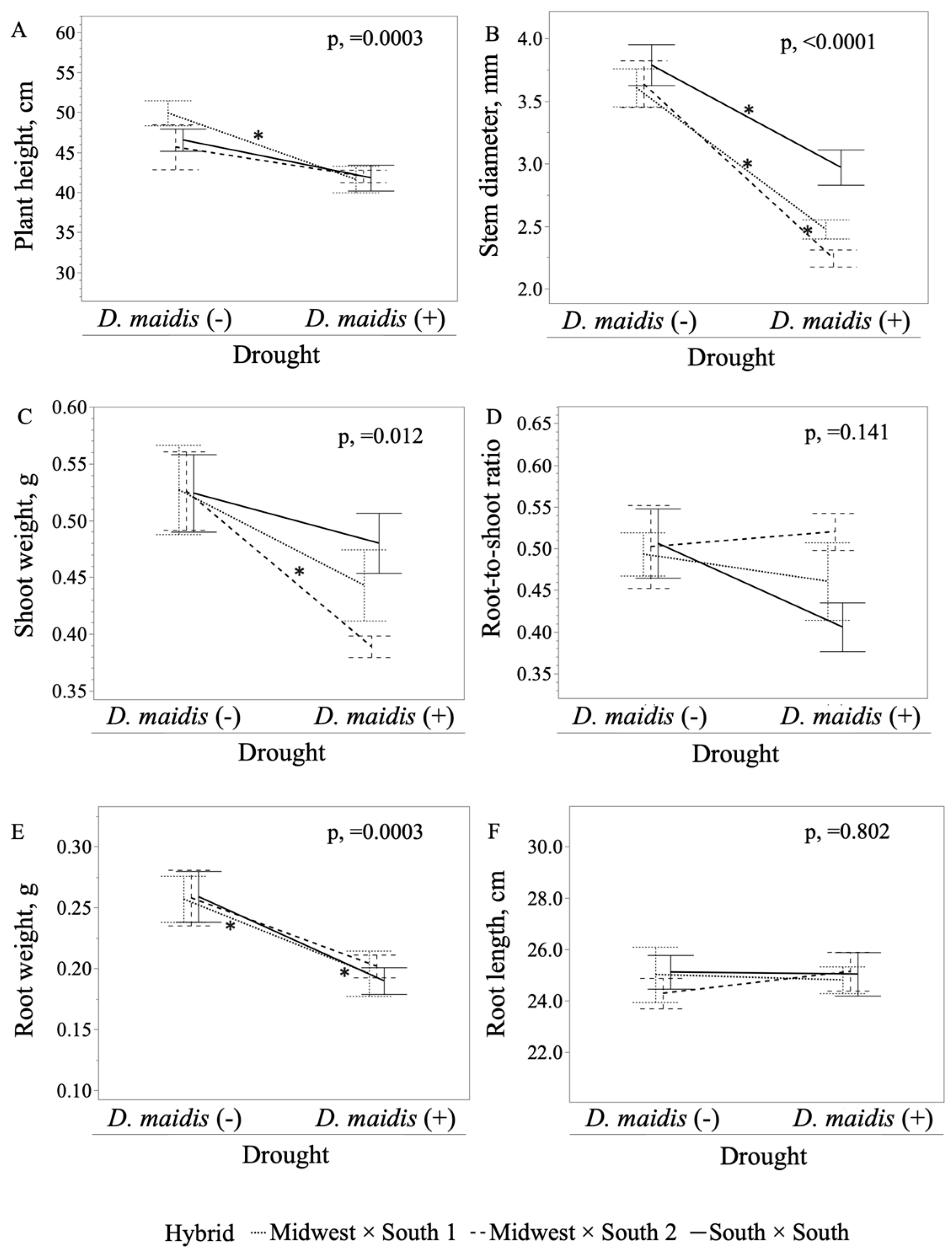
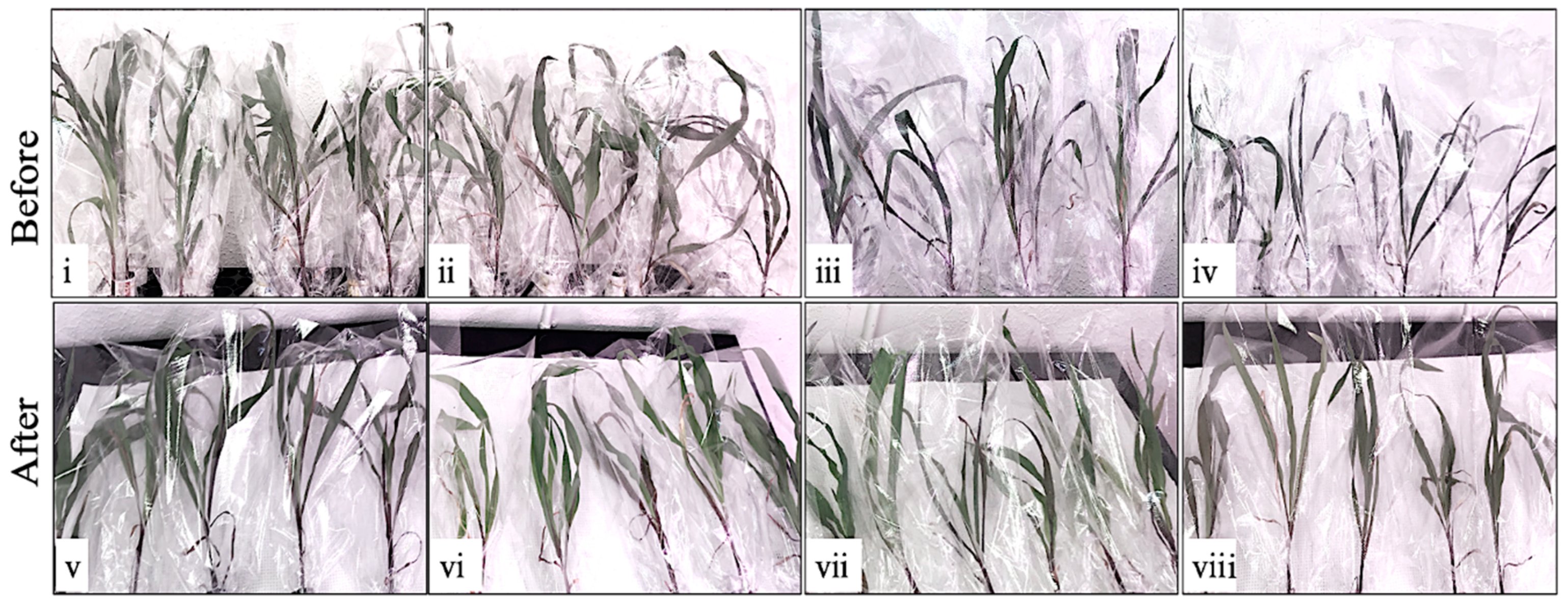
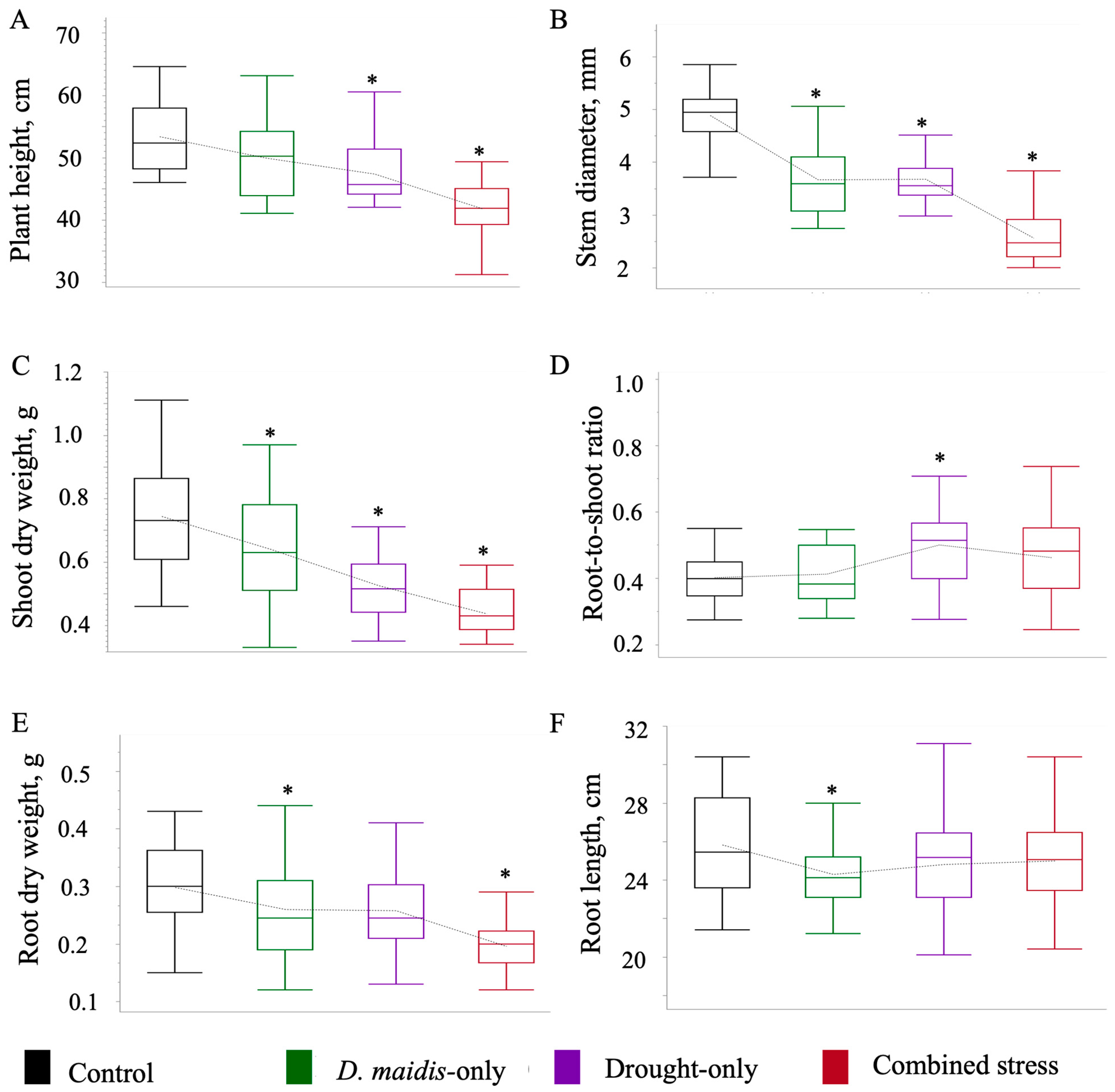
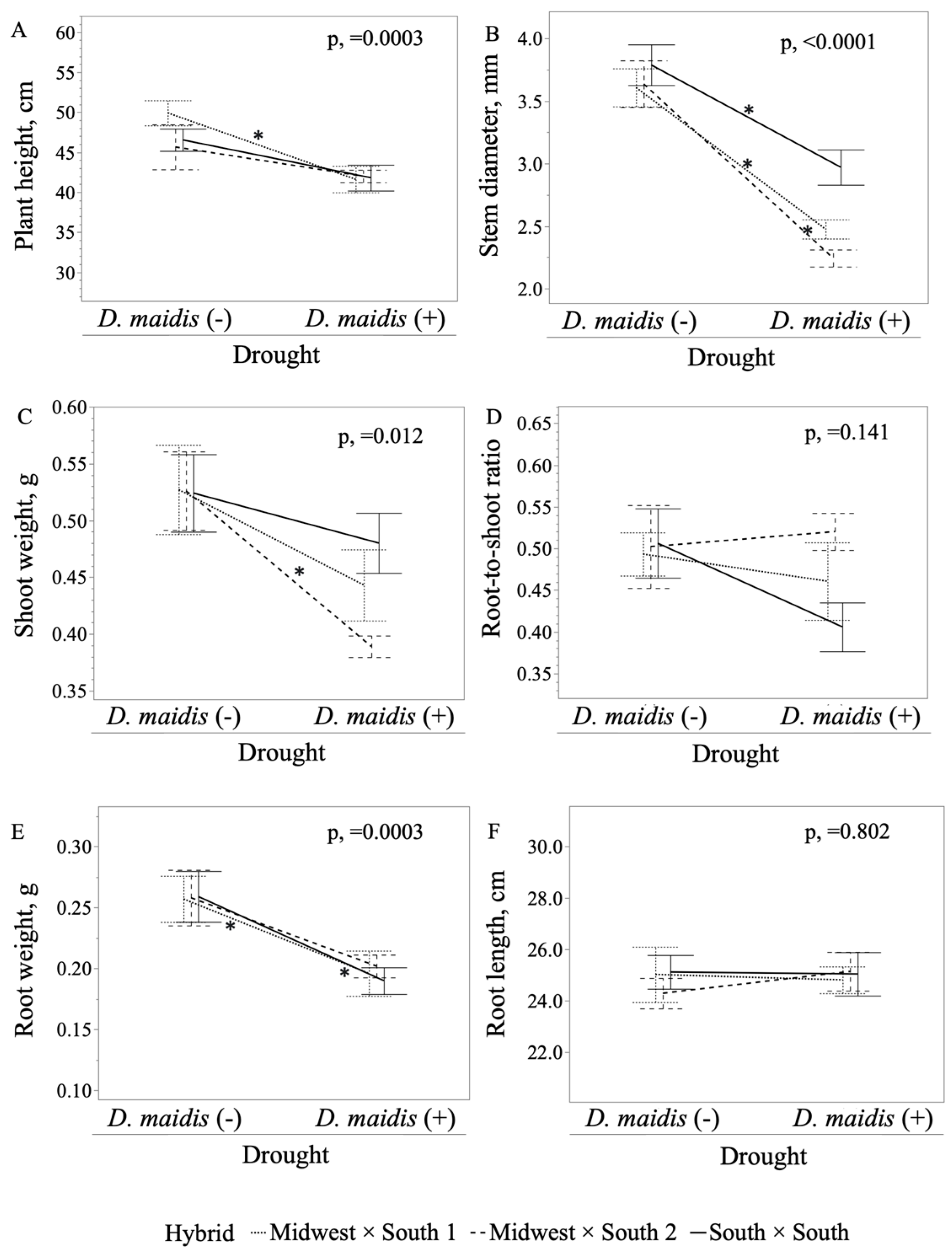
2. Results
3. Discussion
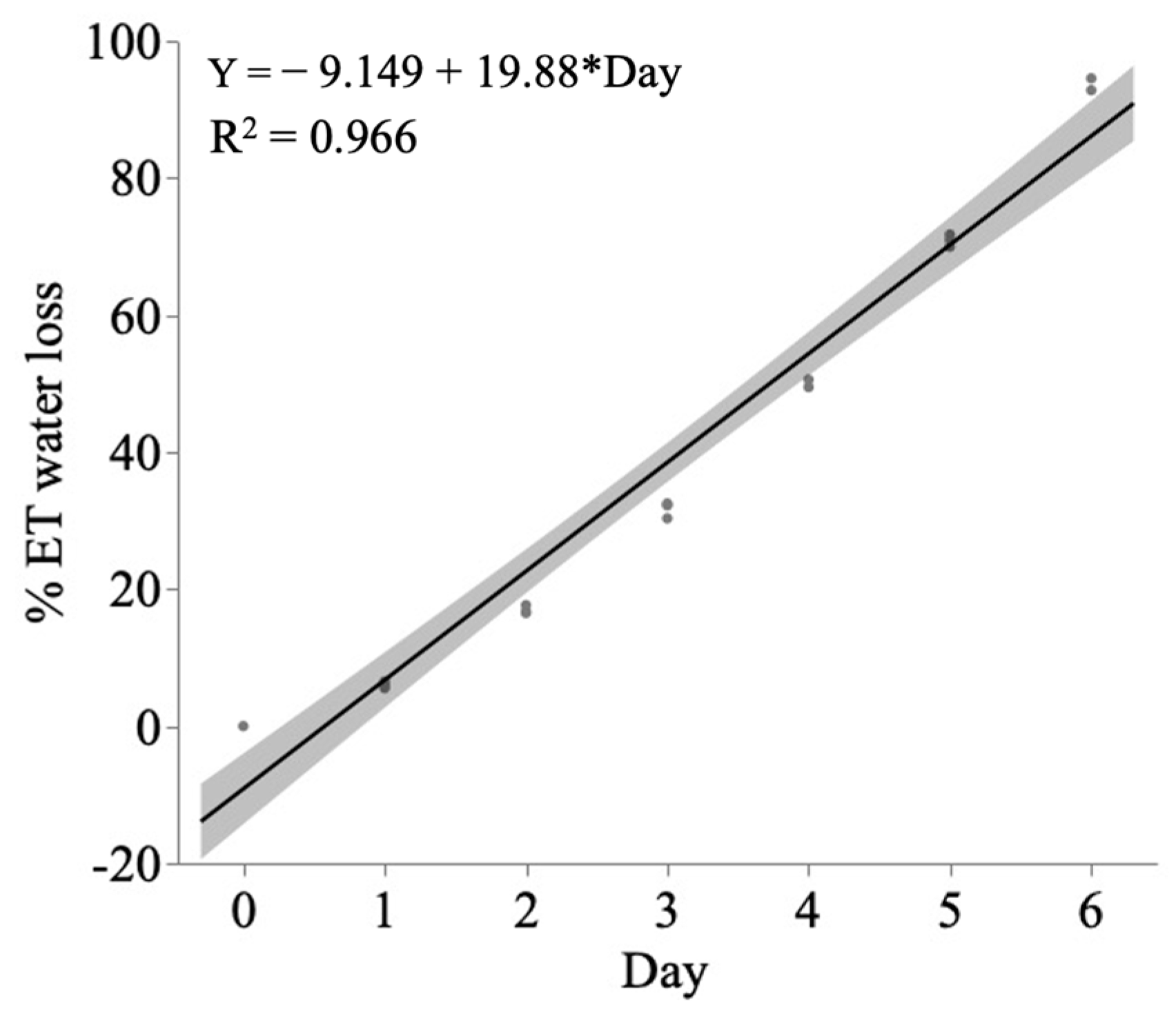
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Dalbulus maidis Rearing
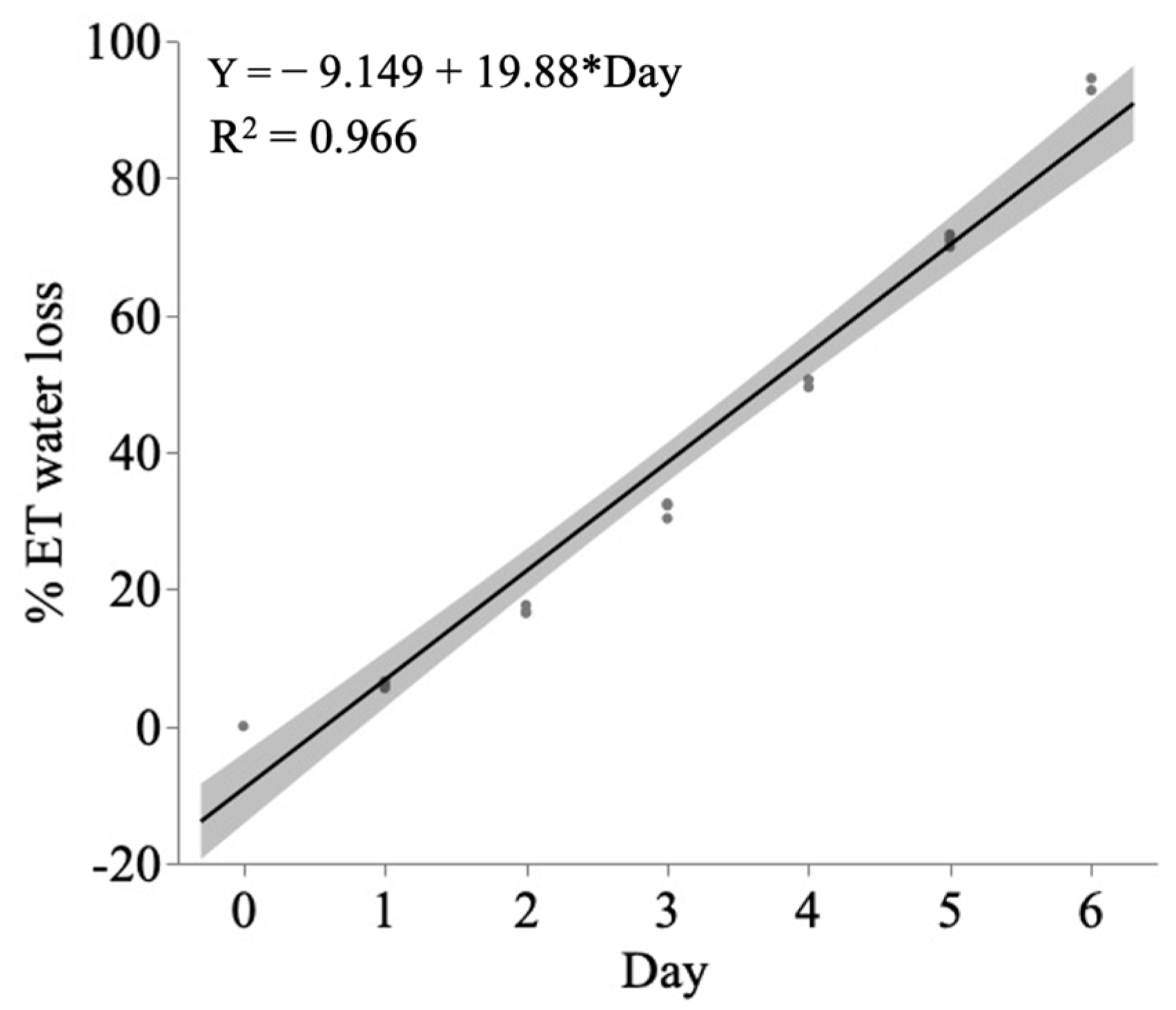
4.3. Drought Induction and Water Regime
4.4. Treatments
4.5. Effects of Dalbulus maidis and Drought on Drought-Tolerant Maize Hybrids
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rafique, S. Drought responses on physiological attributes of Zea mays in relation to nitrogen and source-sink relationships. In Abiotic Stress in Plants; IntechOpen: London, UK, 2020. [Google Scholar]
- Bhusal, B.; Poudel, M.R.; Rishav, P.; Regmi, R.; Neupane, P.; Bhattarai, K.; Maharjan, B.; Bigyan, K.C.; Acharya, S. A Review on Abiotic Stress Resistance in Maize (Zea mays L.): Effects, Resistance Mechanisms and Management. J. Biol. Today’s World 2021, 10, 1–3. [Google Scholar]
- Daryanto, S.; Wang, L.; Jacinthe, P.-A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.C.; Smith, R.G.; Schipanski, M.E.; Atwood, L.W.; Mortensen, D.A. Agriculture in 2050: Recalibrating targets for sustainable intensification. Bioscience 2017, 67, 386–391. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed]
- Maazou, A.-R.S.; Tu, J.; Qiu, J.; Liu, Z. Breeding for drought tolerance in maize (Zea mays L.). Am. J. Plant Sci. 2016, 7, 1858. [Google Scholar] [CrossRef]
- Tigchelaar, M.; Battisti, D.S.; Naylor, R.L.; Ray, D.K. Future warming increases probability of globally synchronized maize production shocks. Proc. Natl. Acad. Sci. USA 2018, 115, 6644. [Google Scholar] [CrossRef]
- Gurian-Sherman, D. High and Dry: Why Genetic Engineering Is Not Solving Agriculture’s Drought Problem in a Thirsty World; Union of Concerned Scientists: Cambridge, MA, USA, 2012. [Google Scholar]
- IPCC. Global Warming of 1.5 °C: Summary of Policy Makers; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2018.
- Boote, K.J.; Ibrahim, A.M.H.; Lafitte, R.; McCulley, R.; Messina, C.; Murray, S.C.; Specht, J.E.; Taylor, S.; Westgate, M.E.; Glasener, K. Position statement on crop adaptation to climate change. Crop Sci. 2011, 51, 2337–2343. [Google Scholar] [CrossRef]
- Ziyomo, C.; Bernardo, R. Drought tolerance in maize: Indirect selection through secondary traits versus genomewide selection. Crop Sci. 2013, 53, 1269–1275. [Google Scholar] [CrossRef]
- Edmeades, G.O. Progress in Achieving and Delivering Drought Tolerance in Maize-an Update; ISAAA: Ithaca, NY, USA, 2013; p. 130. [Google Scholar]
- Ribaut, J.-M.; Betran, J.; Monneveux, P.; Setter, T. Drought tolerance in maize. In Handbook of Maize: Its Biology; Springer: Berlin/Heidelberg, Germany, 2009; pp. 311–344. [Google Scholar]
- Araus, J.L.; Serret, M.D.; Edmeades, G. Phenotyping maize for adaptation to drought. Front. Physiol. 2012, 3, 305. [Google Scholar] [CrossRef]
- Tardieu, F. Any trait or trait-related allele can confer drought tolerance: Just design the right drought scenario. J. Exp. Bot. 2012, 63, 25–31. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar]
- Meeks, M.; Murray, S.C.; Hague, S.; Hays, D. Measuring Maize Seedling Drought Response in Search of Tolerant Germplasm. Agronomy 2013, 3, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Hasibuzzaman, A.S.M.; Akter, F.; Bagum, S.A.; Hossain, N.; Akter, T.; Uddin, M.S. Morpho-Physiological Mechanisms of Maize for Drought Tolerance. In Plant Stress Physiology; IntechOpen: London, UK, 2021; p. 229. [Google Scholar]
- Jenks, M.A.; Hasegawa, P.M.; Jain, S.M. Advances in Molecular Breeding toward Drought and Salt Tolerant Crops; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Hsiao, T.C.; Xu, L.K. Sensitivity of growth of roots versus leaves to water stress: Biophysical analysis and relation to water transport. J. Exp. Bot. 2000, 51, 1595–1616. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant. 2015, 37, 9. [Google Scholar] [CrossRef]
- Nejad, T.S. Effect of drought stress on shoot/root ratio. World Acad. Sci. Eng. Technol. 2011, 5, 539–541. [Google Scholar]
- Khalil, A.M.; Murchie, E.H.; Mooney, S.J. Quantifying the influence of water deficit on root and shoot growth in wheat using X-ray Computed Tomography. AoB Plants 2020, 12, plaa036. [Google Scholar] [CrossRef]
- Comas, L.; Becker, S.; Cruz, V.M.V.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef]
- Shin, S.; Kim, S.G.; Lee, J.S.; Go, T.-H.; Shon, J.; Kang, S.; Lee, J.-S.; Bae, H.H.; Son, B.-Y.; Shim, K.-B.; et al. Impact of the consecutive days of visible wilting on growth and yield during tassel initiation in maize (Zea Mays L.). J. Crop Sci. Biotechnol. 2015, 18, 219–229. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, H.; Wu, X.; Wang, W. Identification of drought tolerant mechanisms in a drought-tolerant maize mutant based on physiological, biochemical and transcriptomic analyses. BMC Plant Biol. 2020, 20, 315. [Google Scholar] [CrossRef]
- Ali, F.; Ahsan, M.; Ali, Q.; Kanwal, N. Phenotypic Stability of Zea mays Grain Yield and Its Attributing Traits under Drought Stress. Front. Plant Sci. 2017, 8, 1397. [Google Scholar] [CrossRef]
- Mousavi, S.M.N.; Nagy, J. Evaluation of plant characteristics related to grain yield of FAO410 and FAO340 hybrids using regression models. Cereal Res. Commun. 2021, 49, 161–169. [Google Scholar] [CrossRef]
- Slewinski, T.L. Non-structural carbohydrate partitioning in grass stems: A target to increase yield stability, stress tolerance, and biofuel production. J. Exp. Bot. 2012, 63, 4647–4670. [Google Scholar] [CrossRef] [PubMed]
- Kaya, Y.; Evci, G.; Durak, S.; Pekcan, V.; Gücer, T. Determining the relationships between yield and yield attributes in sunflower. Turk. J. Agric. For. 2007, 31, 237–244. [Google Scholar]
- Liang, X.-L.; Abu, L.-L.; Feng, G.J. Yield Performance of Maize Hybrids and Analysis of Correlation between Yield and Agronomic Characteristics. Maize Sci. 2001, 9, 1. [Google Scholar]
- Pedersen, I.F.; Christensen, J.T.; Sørensen, P.; Christensen, B.T.; Holton Rubæk, G. Early plant height: A defining factor for yields of silage maize with contrasting phosphorus supply. Soil Use Manag. 2021, 38, 537–548. [Google Scholar] [CrossRef]
- Aman, J.; Bantte, K.; Alamerew, S.; Sbhatu, D.B. Correlation and path coefficient analysis of yield and yield components of quality protein maize (Zea mays L.) hybrids at Jimma, western Ethiopia. Int. J. Agron. 2020, 2020, 6998871. [Google Scholar] [CrossRef]
- Culy, M.D. Yield loss of field corn from insects. In Biotic Stress and Yield Loss; CRC Press: Boca Raton, FL, USA, 2000; pp. 57–86. [Google Scholar]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef]
- Virla, E.G.; Araoz, M.V.C.; Albarracin, E.L. Estimation of direct damage to maize seedlings by the corn leafhopper, Dalbulus maidis (Hemiptera: Cicadellidae), under different watering regimes. Bull. Entomol. Res. 2021, 111, 438–444. [Google Scholar] [CrossRef]
- Chander, S.; Husain, M.; Pal, V. Insect Pest Management in Climate Change; Studera Press: Delhi, India, 2016. [Google Scholar]
- Konvicka, M.; Maradova, M.; Benes, J.; Fric, Z.; Kepka, P. Uphill shifts in distribution of butterflies in the Czech Republic: Effects of changing climate detected on a regional scale. Glob. Ecol. Biogeogr. 2003, 12, 403–410. [Google Scholar] [CrossRef]
- Panizzi, A.R. Suboptimal nutrition and feeding behavior of hemipterans on less preferred plant food sources. An. Soc. Entomol. Bras. 2000, 29, 1–12. [Google Scholar] [CrossRef]
- Waquil, J.M. Sampling and abundance of leafhoppers and damage by Dalbulus maidis (DeLong&Wolcott) (Homoptera: Cicadellidae) to maize seedling. An. Soc. Entomol. Bras. 1997, 26, 27–33. [Google Scholar]
- Santana Jr, P.A.; Kumar, L.; Da Silva, R.S.; Pereira, J.L.; Picanço, M.C. Assessing the impact of climate change on the worldwide distribution of Dalbulus maidis (DeLong) using MaxEnt. Pest Manag. Sci. 2019, 75, 2706–2715. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Cabral, N.Y.Z.; Kumar, L.; Shabani, F. Global alterations in areas of suitability for maize production from climate change and using a mechanistic species distribution model (CLIMEX). Sci. Rep. 2017, 7, 5910. [Google Scholar] [CrossRef]
- Nault, L.R.; Delong, D.M. Evidence for co-evolution of leafhoppers in the genus Dalbulus (Cicadellidae: Homoptera) with maize and its ancestors. Ann. Entomol. Soc. Am. 1980, 73, 349–353. [Google Scholar] [CrossRef]
- Medina, R.F.; Reyna, S.M.; Bernal, J.S. Population genetic structure of a specialist leafhopper on Zea: Likely anthropogenic and ecological determinants of gene flow. Entomol. Exp. Appl. 2012, 142, 223–235. [Google Scholar] [CrossRef]
- Bellota, E.; Dávila-Flores, A.; Bernal, J.S. A Bird in the Hand Versus Two in the Bush? The Specialist Leafhopper Dalbulus maidis (Hemiptera: Cicadellidae) Does Not Discriminate Against Sub-optimal Host Plants (Zea spp.). Neotrop. Entomol. 2017, 47, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Arias, C.C.; Ligarreto-Moreno, G.A.; Ramírez-Godoy, A.; Restrepo-Díaz, H. Maize Responses Challenged by Drought, Elevated Daytime Temperature and Arthropod Herbivory Stresses: A Physiological, Biochemical and Molecular View. Front. Plant Sci. 2021, 12, 1512. [Google Scholar] [CrossRef]
- Park, Y.-S.; Bae, D.-W.; Ryu, C.-M. Aboveground whitefly infestation modulates transcriptional levels of anthocyanin biosynthesis and jasmonic acid signaling-related genes and augments the cope with drought stress of maize. PLoS ONE 2015, 10, e0143879. [Google Scholar] [CrossRef]
- Strauss, S.Y.; Agrawal, A.A. The ecology and evolution of plant tolerance to herbivory. Trends Ecol. Evol. 1999, 14, 179–185. [Google Scholar] [CrossRef]
- Müller-Schärer, H.; Schaffner, U.; Steinger, T. Evolution in invasive plants: Implications for biological control. Trends Ecol. Evol. 2004, 19, 417–422. [Google Scholar] [CrossRef]
- Chinchilla-Ramírez, M.; Borrego, E.J.; DeWitt, T.J.; Kolomiets, M.V.; Bernal, J.S. Maize seedling morphology and defence hormone profiles, but not herbivory tolerance, were mediated by domestication and modern breeding. Ann. Appl. Biol. 2017, 170, 315–332. [Google Scholar] [CrossRef]
- White, T.C.R. A hypothesis to explain outbreaks of looper caterpillars, with special reference to populations of Selidosema suavis in a plantation of Pinus radiata in New Zealand. Oecologia 1974, 16, 279–301. [Google Scholar] [CrossRef] [PubMed]
- Dale, A.G.; Frank, S.D. Warming and drought combine to increase pest insect fitness on urban trees. PLoS ONE 2017, 12, e0173844. [Google Scholar] [CrossRef] [PubMed]
- Herms, D.A.; Mattson, W.J. The dilemma of plants: To grow or defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef]
- Koricheva, J.; Larsson, S.; Haukioja, E. Insect performance on experimentally stressed woody plants: A meta-analysis. Annu. Rev. Entomol. 1998, 43, 195–216. [Google Scholar] [CrossRef] [PubMed]
- Diezel, C.; von Dahl, C.C.; Gaquerel, E.; Baldwin, I.T. Different lepidopteran elicitors account for cross-talk in herbivory-induced phytohormone signaling. Plant Physiol. 2009, 150, 1576–1586. [Google Scholar] [CrossRef] [Green Version]
- Huberty, A.; Denno, R. Plant water stress and its consequences for herbivorous insects: A new synthesis. Ecology 2004, 85, 1383–1398. [Google Scholar] [CrossRef]
- Beetge, L.; Krüger, K. Drought and heat waves associated with climate change affect performance of the potato aphid Macrosiphum euphorbiae. Sci. Rep. 2019, 9, 3645. [Google Scholar] [CrossRef]
- Xie, H.; Shi, J.; Shi, F.; Wang, X.; Xu, H.; He, K.; Wang, Z. Aphid fecundity and defenses in wheat exposed to a combination of heat and drought stress. J. Exp. Bot. 2020, 71, 2713–2722. [Google Scholar] [CrossRef]
- Copolovici, L.; Kännaste, A.; Remmel, T.; Niinemets, Ü. Volatile organic compound emissions from Alnus glutinosa under interacting drought and herbivory stresses. Environ. Exp. Bot. 2014, 100, 55–63. [Google Scholar] [CrossRef]
- Cooper, M.; Gho, C.; Leafgren, R.; Tang, T.; Messina, C. Breeding drought-tolerant maize hybrids for the US corn-belt: Discovery to product. J. Exp. Bot. 2014, 65, 6191–6204. [Google Scholar] [CrossRef]
- Sawilowsky, S.S. New effect size rules of thumb. J. Mod. Appl. Stat. Methods 2009, 8, 26. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge: London, UK, 2013. [Google Scholar]
- Ashraf, M.; Nawazish, S.; Athar, H.-U.-R. Are chlorophyll fluorescence and photosynthetic capacity potential physiological determinants of drought tolerance in maize (Zea mays L.). Pak. J. Bot. 2007, 39, 1123–1131. [Google Scholar]
- Rahbarian, R.; Khavari-Nejad, R.; Ganjeali, A.; Bagheri, A.; Najafi, F. Drought stress effects on photosynthesis, chlorophyll fluorescence and water relations in tolerant and susceptible chickpea (Cicer arietinum L.) genotypes. Acta Biol. Crac. Ser. Bot. 2011, 53, 47–56. [Google Scholar] [CrossRef]
- Jin, Z.; Xue, Q.-W.; Jessup, K.E.; Hou, X.-B.; Hao, B.-Z.; Marek, T.H.; Xu, W.-W.; Evett, S.R.; O’Shaughnessy, S.A.; Brauer, D.K. Shoot and root traits in drought tolerant maize (Zea mays L.) hybrids. J. Integr. Agric. 2018, 17, 1093–1105. [Google Scholar]
- Aslam, M.; Maqbool, M.A.; Cengiz, R. Effects of drought on maize. In Drought Stress in Maize (Zea mays L.); Springer: Berlin/Heidelberg, Germany, 2015; pp. 5–17. [Google Scholar]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Gilbert, M.E.; Medina, V. Drought Adaptation Mechanisms Should Guide Experimental Design. Trends Plant Sci. 2016, 21, 639–647. [Google Scholar] [CrossRef]
- Kränzlein, M.; Geilfus, C.-M.; Franzisky, B.L.; Zhang, X.; Wimmer, M.A.; Zörb, C. Physiological Responses of Contrasting Maize (Zea mays L.) Hybrids to Repeated Drought. J. Plant Growth Regul. 2021. [Google Scholar] [CrossRef]
- Avramova, V.; Nagel, K.A.; AbdElgawad, H.; Bustos, D.; DuPlessis, M.; Fiorani, F.; Beemster, G.T.S. Screening for drought tolerance of maize hybrids by multi-scale analysis of root and shoot traits at the seedling stage. J. Exp. Bot. 2016, 67, 2453–2466. [Google Scholar] [CrossRef]
- Li, X.-J.; Yang, M.-F.; Chen, H.; Qu, L.-Q.; Chen, F.; Shen, S.-H. Abscisic acid pretreatment enhances salt tolerance of rice seedlings: Proteomic evidence. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2010, 1804, 929–940. [Google Scholar] [CrossRef]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 2016, 4, 162–176. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [PubMed]
- Bostock, R.M.; Pye, M.F.; Roubtsova, T.V. Predisposition in plant disease: Exploiting the nexus in abiotic and biotic stress perception and response. Annu. Rev. Phytopathol. 2014, 52, 517–549. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Dommel, M.; Mou, Z. Abscisic acid promotes proteasome-mediated degradation of the transcription coactivator NPR 1 in Arabidopsis thaliana. Plant J. 2016, 86, 20–34. [Google Scholar] [CrossRef]
- Berens, M.L.; Wolinska, K.W.; Spaepen, S.; Ziegler, J.; Nobori, T.; Nair, A.; Krüler, V.; Winkelmüller, T.M.; Wang, Y.; Mine, A.; et al. Balancing trade-offs between biotic and abiotic stress responses through leaf age-dependent variation in stress hormone cross-talk. Proc. Natl. Acad. Sci. USA 2019, 116, 2364. [Google Scholar] [CrossRef] [PubMed]
- Moeder, W.; Ung, H.; Mosher, S.; Yoshioka, K. SA-ABA antagonism in defense responses. Plant Signal. Behav. 2010, 5, 1231–1233. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.C.; Mayfield, K.; Pekar, J.; Brown, P.; Lorenz, A.; Isakeit, T.; Odvody, G.; Xu, W.; Betran, J. Tx741, Tx777, Tx779, Tx780, and Tx782 Inbred Maize Lines for Yield and Southern United States Stress Adaptation. J. Plant Regist. 2019, 13, 258–269. [Google Scholar] [CrossRef]
- Llorente, C.F.; Betrán, F.J.; Bockholt, A.; Fojt Iii, F. Registration of Tx772 Maize. Crop Sci. 2004, 44, 1036–1037. [Google Scholar] [CrossRef]
- Foley, T.J. Inbred Corn Line LH195. U.S. Patant 5059745A, 22 October 1991. [Google Scholar]
- Abdi, H. Bonferroni and Šidák corrections for multiple comparisons. Encycl. Meas. Stat. 2007, 3, 103–107. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Level | -Level | Difference ± SE | p-Value | Effect Size |
|---|---|---|---|---|---|
| Plant height | |||||
| insect | control | −3.386 ± 1.504 | 0.069 | 0.592 | |
| drought | control | −5.999 ± 1.476 | 0.0003 | 1.049 | |
| combine | control | −11.555 ± 1.477 | <0.0001 | 2.021 | |
| Stem diameter | |||||
| insect | control | −1.256 ± 0.144 | <0.0001 | 2.290 | |
| drought | control | −1.210 ± 0.142 | <0.0001 | 2.207 | |
| combine | control | −2.330 ± 0.142 | <0.0001 | 4.250 | |
| Shoot weight | |||||
| insect | control | −0.115 ± 0.036 | 0.005 | 0.844 | |
| drought | control | −0.219 ± 0.035 | <0.0001 | 1.604 | |
| combine | control | −0.309 ± 0.035 | <0.0001 | 2.263 | |
| Root/shoot ratio | |||||
| insect | control | −0.003 ± 0.027 | 0.999 | 0.026 | |
| drought | control | −0.100 ± 0.027 | 0.001 | 0.954 | |
| combine | control | 0.059 ± 0.027 | 0.075 | 0.572 | |
| Root weight | |||||
| insect | control | −0.048 ± 0.017 | 0.019 | 0.722 | |
| drought | control | −0.041 ± 0.017 | 0.048 | 0.621 | |
| combine | control | −0.104 ± 0.017 | <0.0001 | 1.581 | |
| Root length | |||||
| insect | control | −1.800 ± 0.607 | 0.010 | 13.187 | |
| drought | control | −1.031 ± 0.596 | 0.207 | 7.553 | |
| combine | control | −0.881 ± 0.596 | 0.323 | 6.456 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, T.-K.L.; Medina, R.F.; Bernal, J.S. Effects of Phloem-Feeding Pest, Dalbulus maidis on Morphological Expression of Drought-Tolerant Traits in Maize. Stresses 2022, 2, 322-335. https://doi.org/10.3390/stresses2030023
Jones T-KL, Medina RF, Bernal JS. Effects of Phloem-Feeding Pest, Dalbulus maidis on Morphological Expression of Drought-Tolerant Traits in Maize. Stresses. 2022; 2(3):322-335. https://doi.org/10.3390/stresses2030023
Chicago/Turabian StyleJones, Tara-Kay L., Raul F. Medina, and Julio S. Bernal. 2022. "Effects of Phloem-Feeding Pest, Dalbulus maidis on Morphological Expression of Drought-Tolerant Traits in Maize" Stresses 2, no. 3: 322-335. https://doi.org/10.3390/stresses2030023
APA StyleJones, T.-K. L., Medina, R. F., & Bernal, J. S. (2022). Effects of Phloem-Feeding Pest, Dalbulus maidis on Morphological Expression of Drought-Tolerant Traits in Maize. Stresses, 2(3), 322-335. https://doi.org/10.3390/stresses2030023