
Comparative Study of Trehalose and Trehalose 6-Phosphate to Improve Antioxidant Defense Mechanisms in Wheat and Mustard Seedlings under Salt and Water Deficit Stresses

 ,
, 

Abstract
:1. Introduction
2. Results
2.1. Growth of Wheat and Mustard Seedlings Was Improved by Trehalose and T6P Application under Salt and Water Deficit Stress
2.2. Trehalose and T6P Improved Photosynthetic Pigments under Salt and Water Deficit Stress
2.3. Trehalose Contents Increased Due to the Application of Trehalose and T6P under Salt and Water Deficit Stress
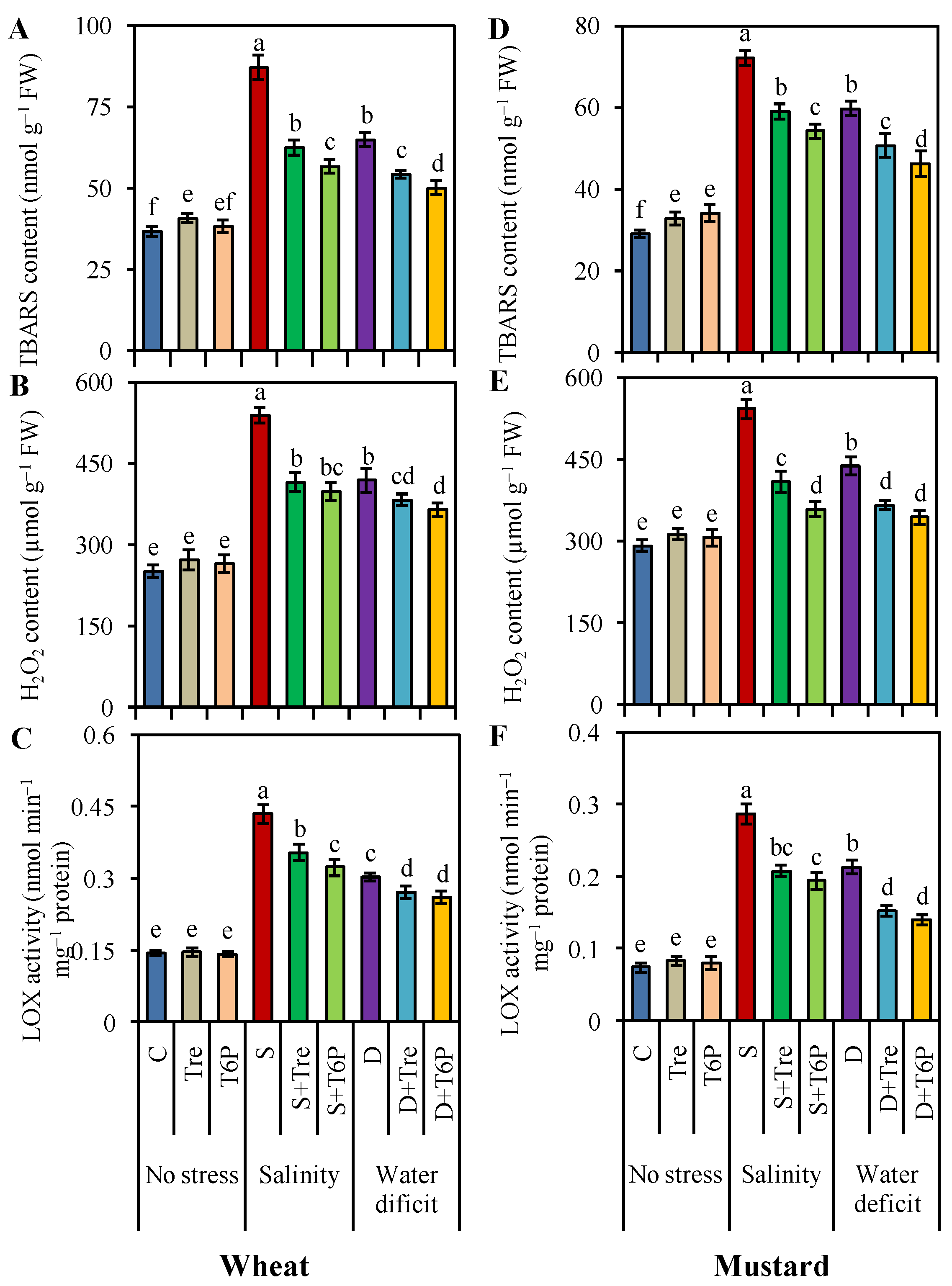
2.4. Trehalose and T6P Reduced Oxidative Stress Indicators
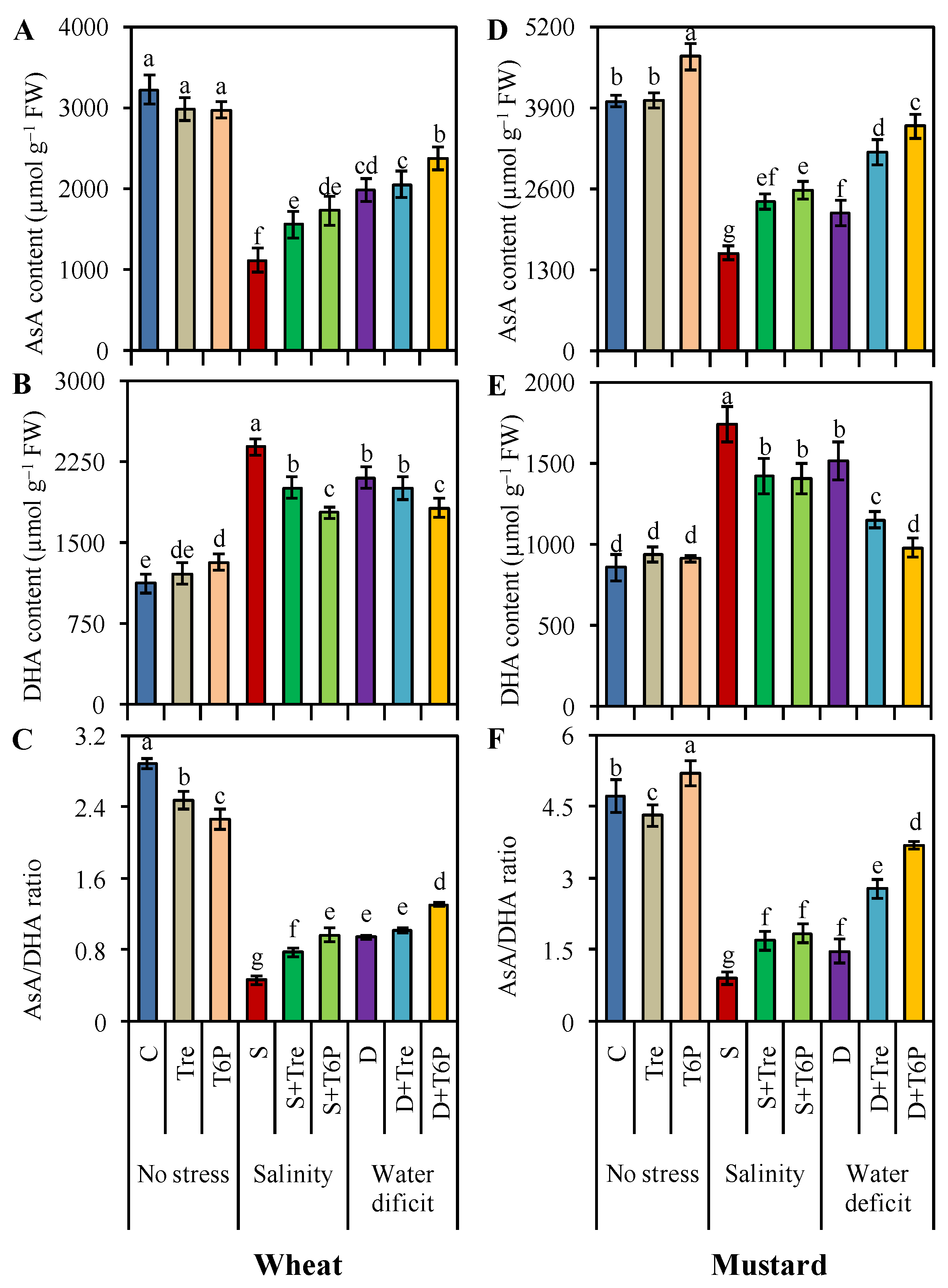
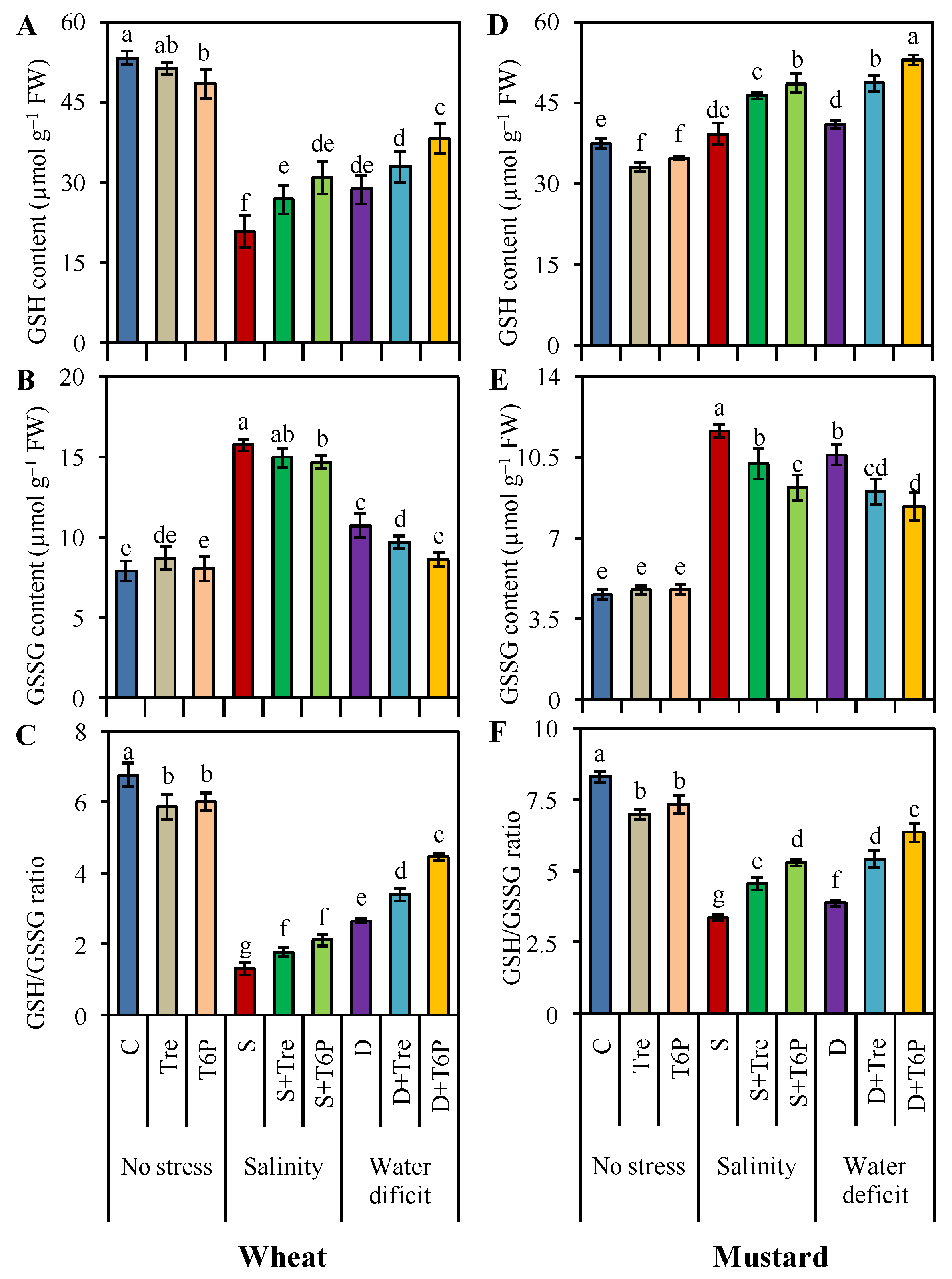
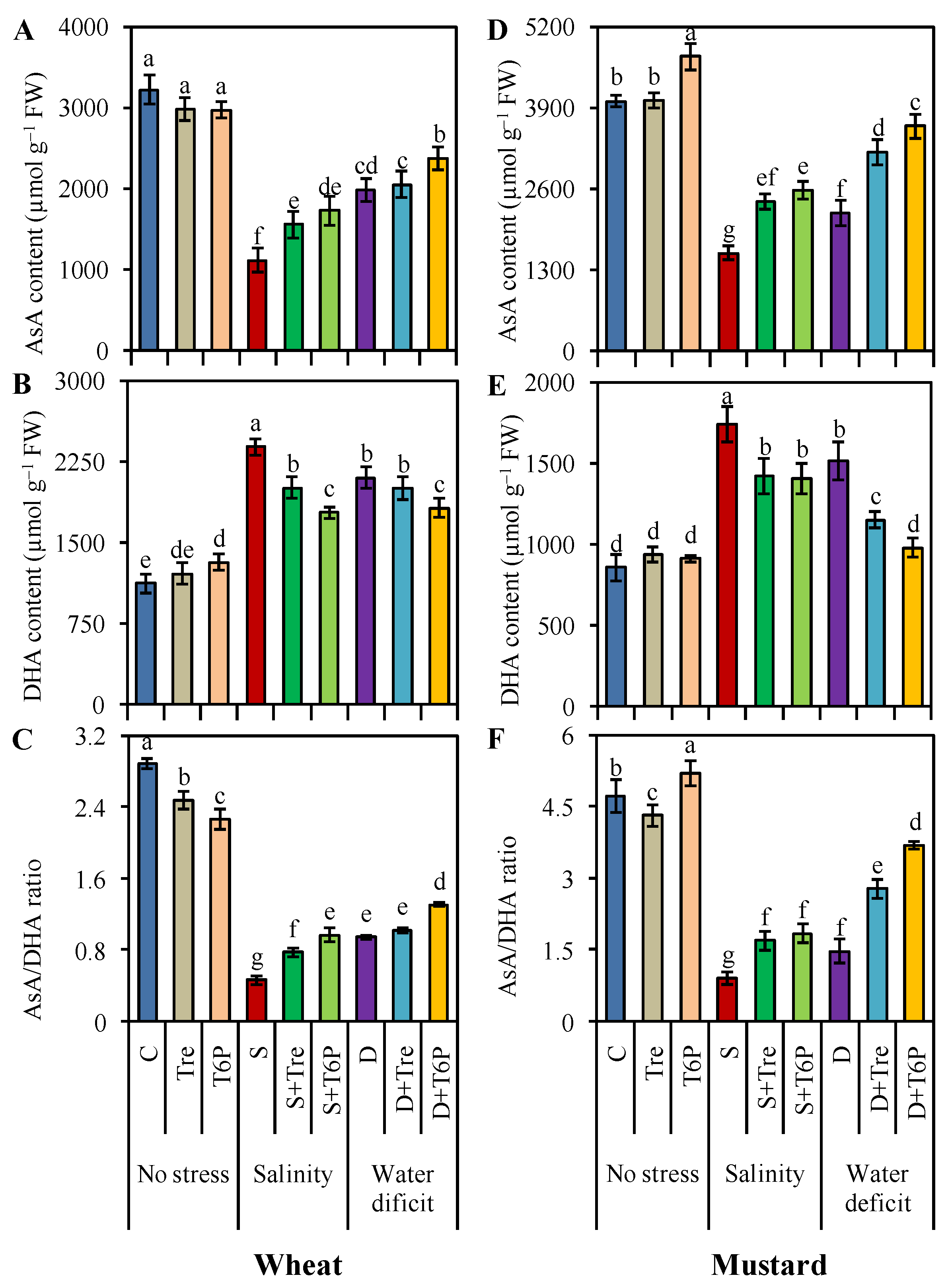
2.5. AsA-GSH Pool Is Enhanced by Trehalose and T6P under Salt and Water Deficit Stress
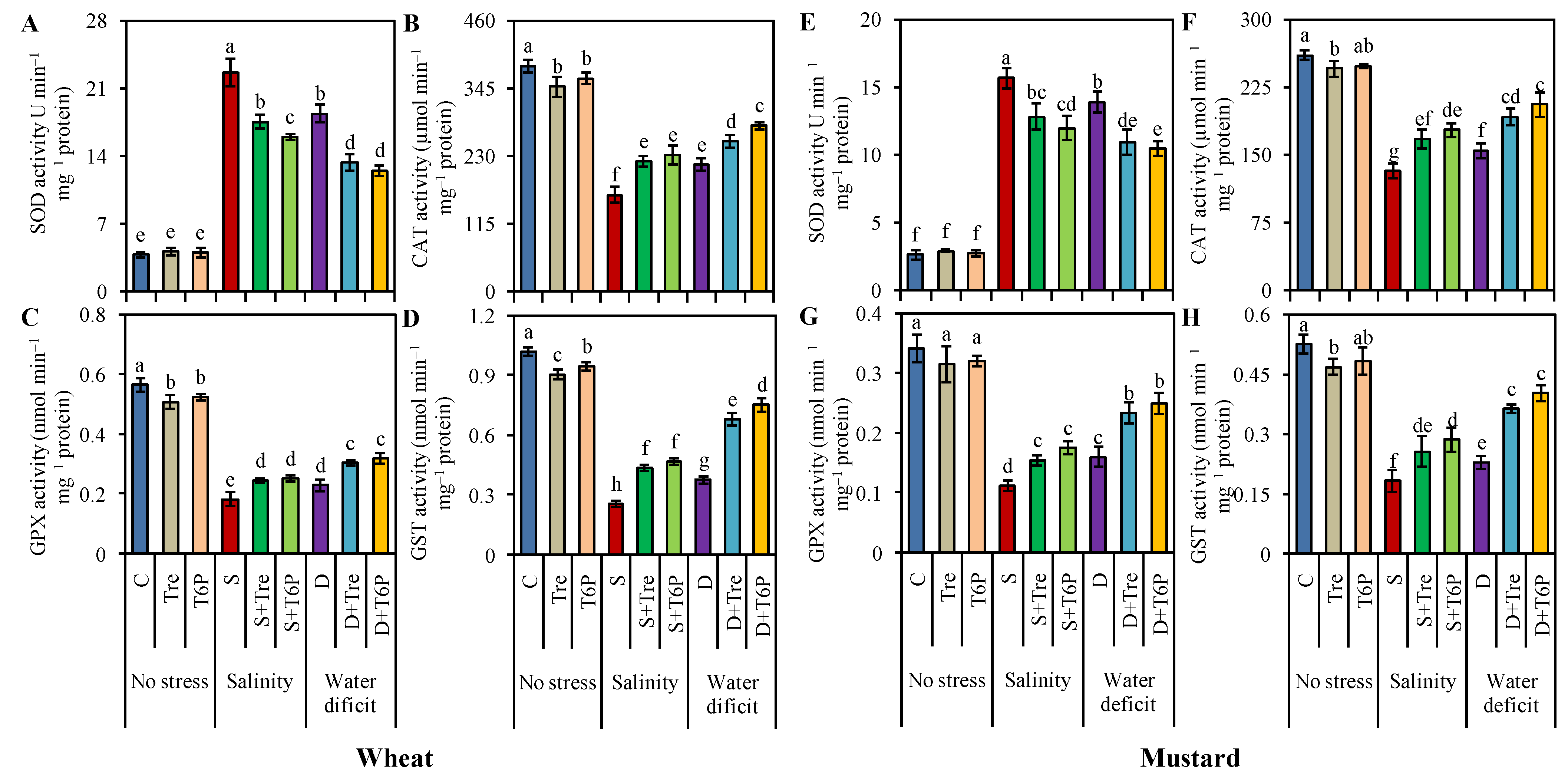
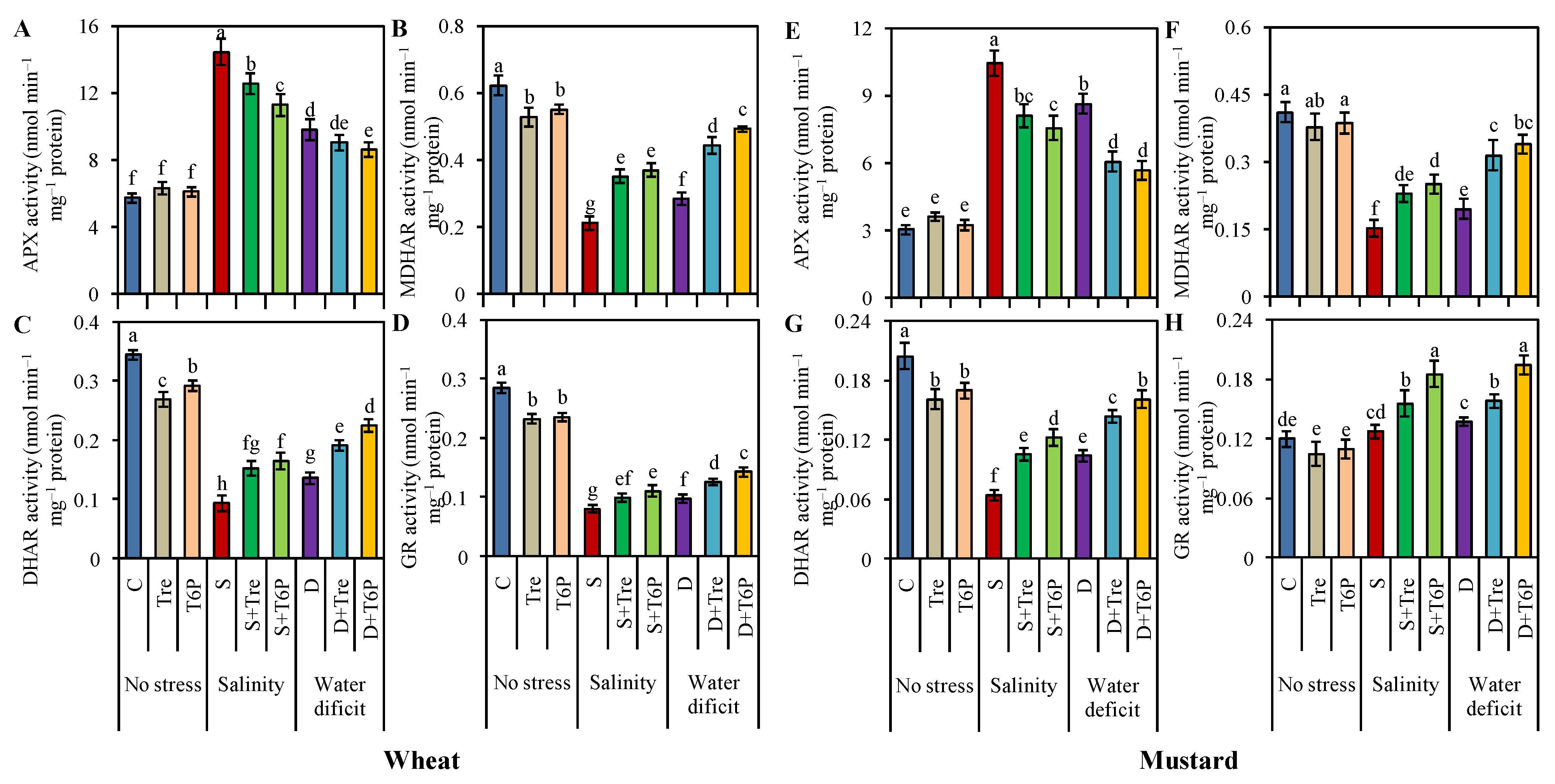
2.6. Up-Regulation of Antioxidant Enzyme Activities Due to Trehalose and T6P Application under Salt and Water Deficit Stress
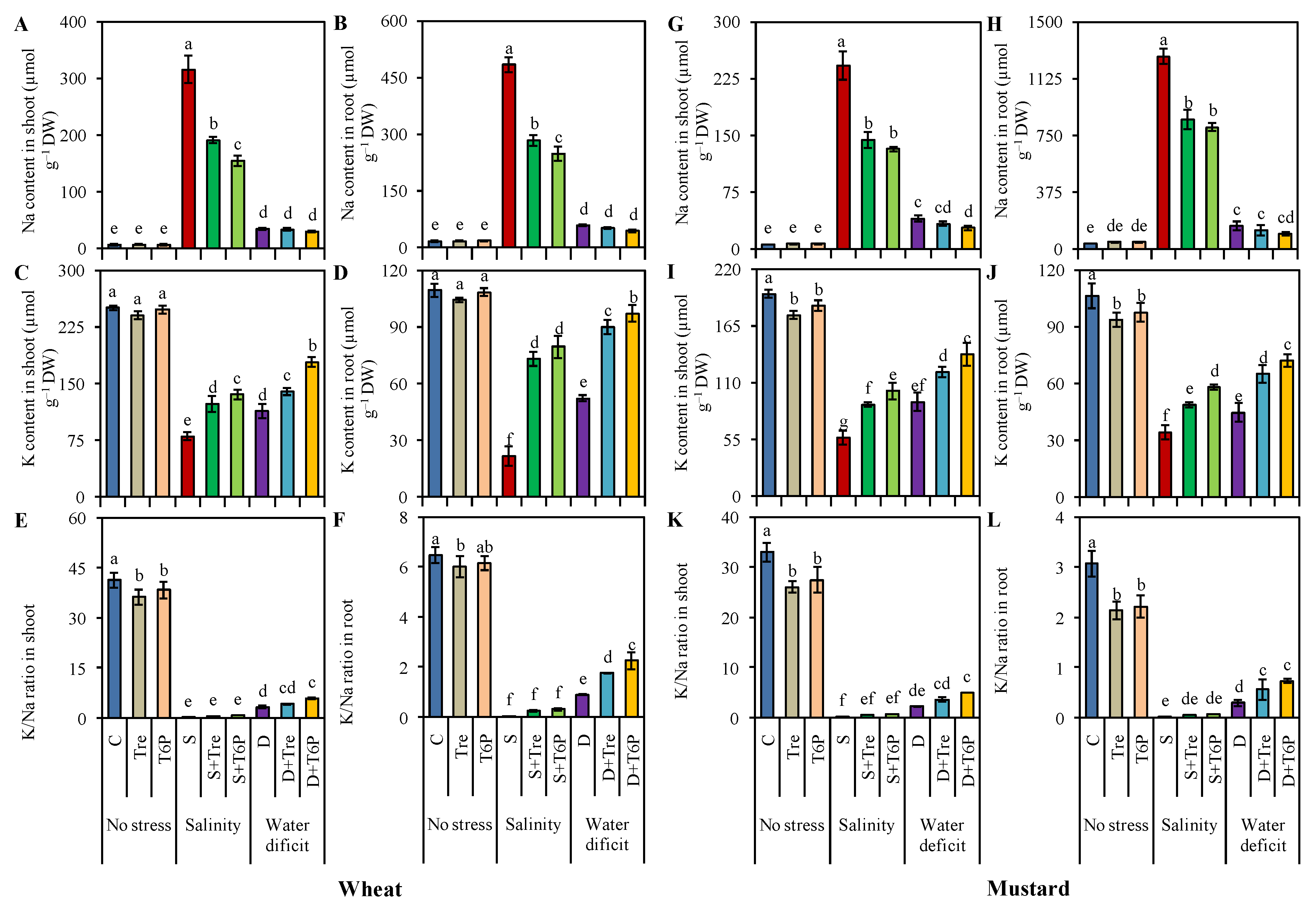
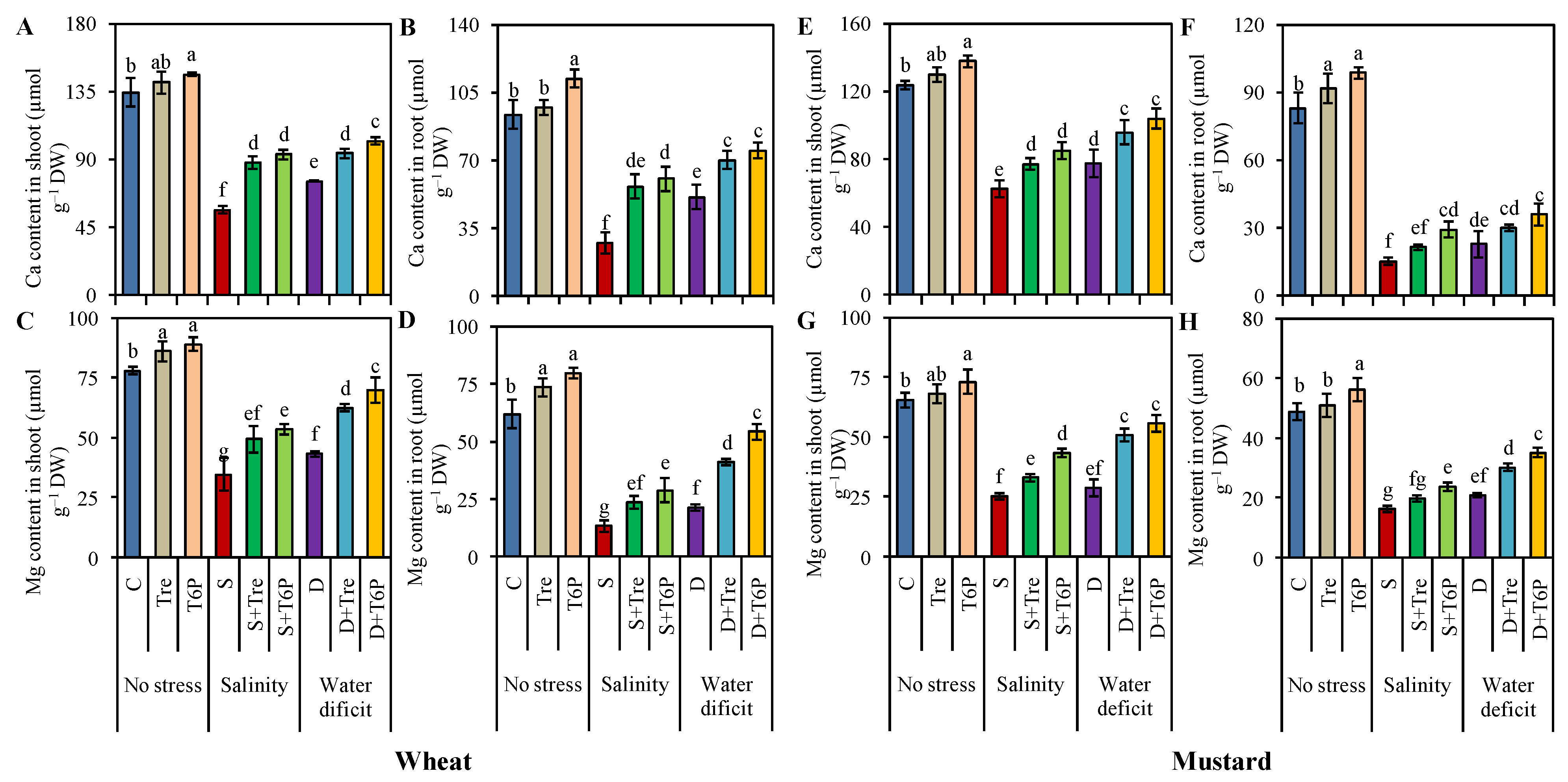
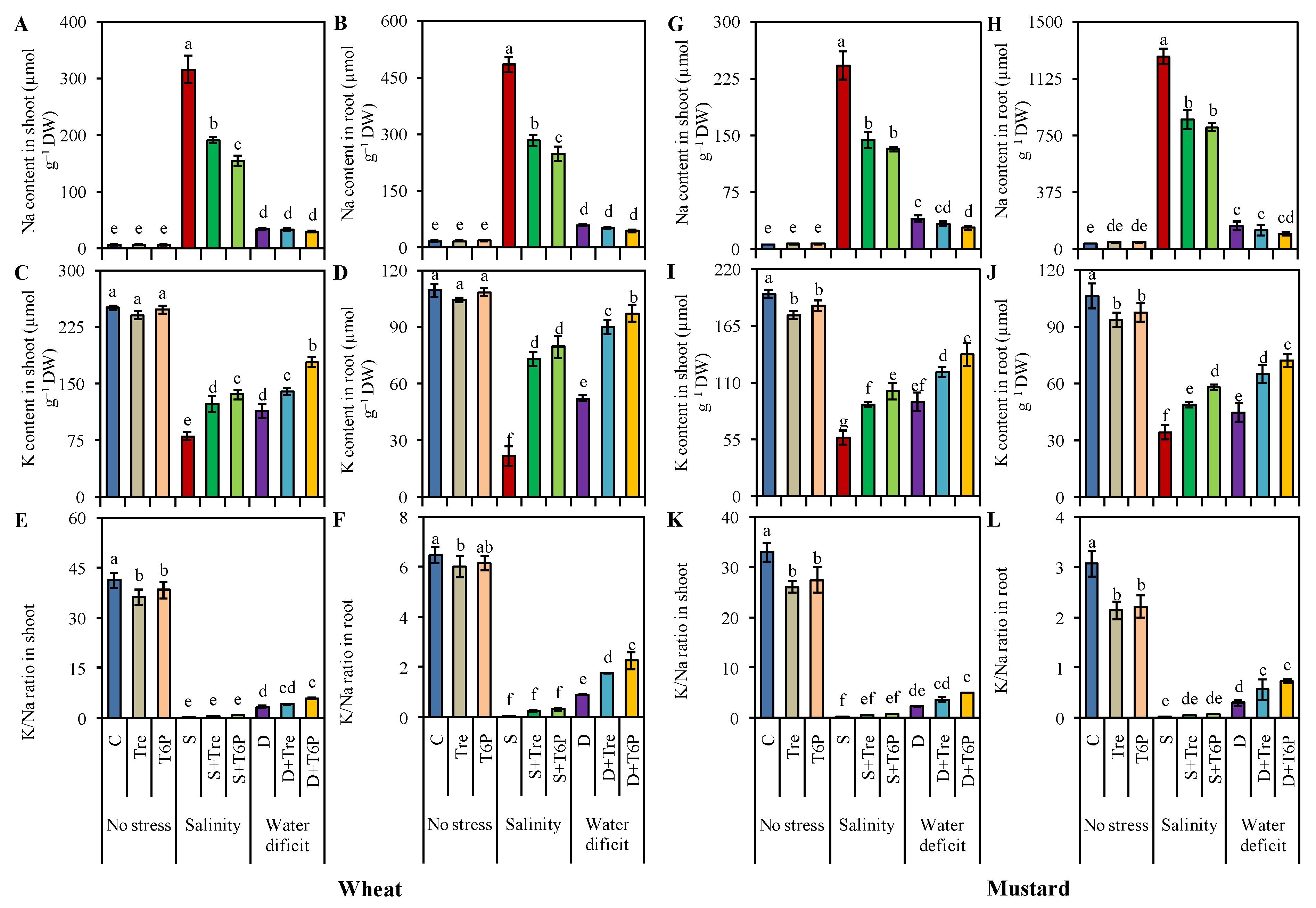
2.7. Trehalose and T6P Reduced Na Accumulation but Increased K, Ca, and Mg Uptake
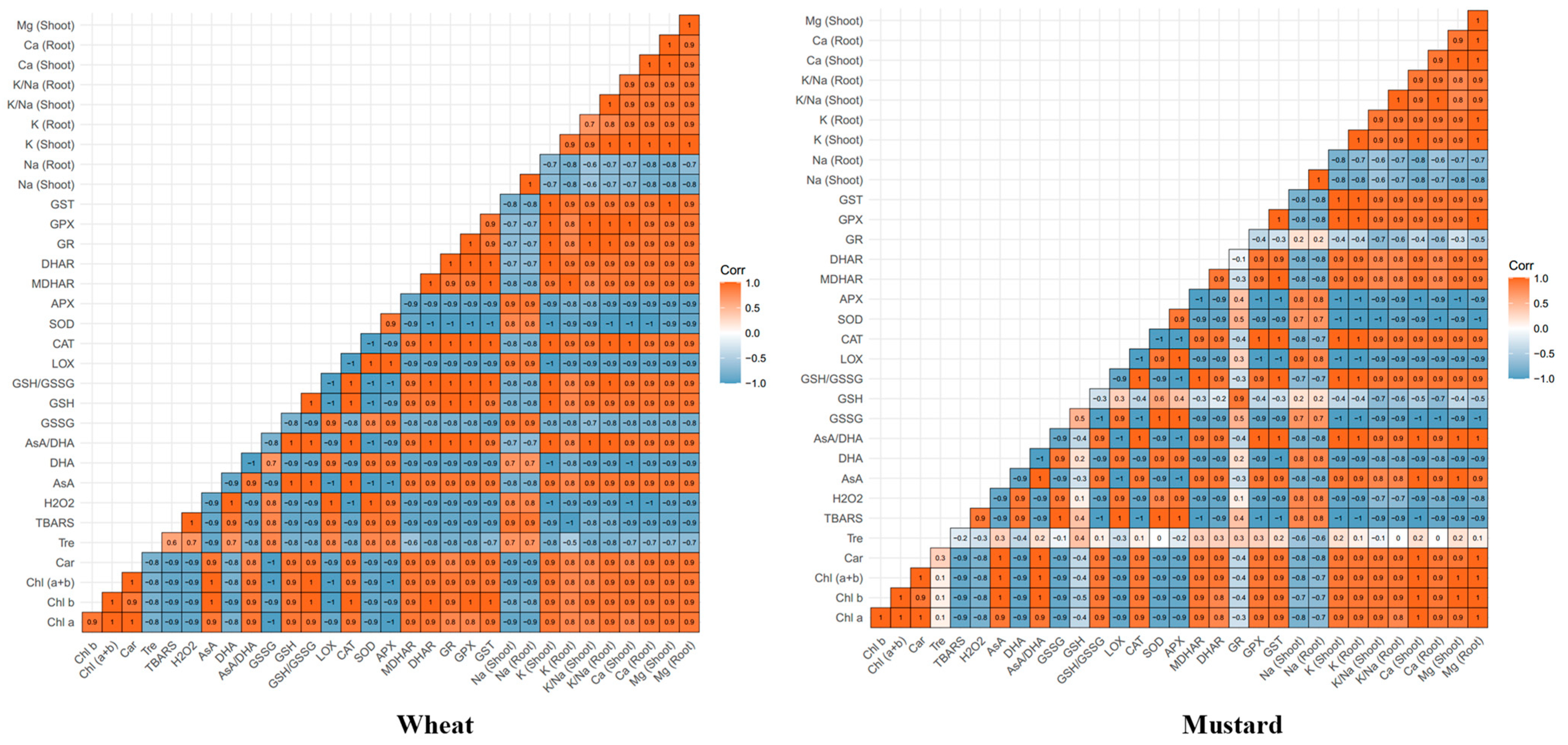
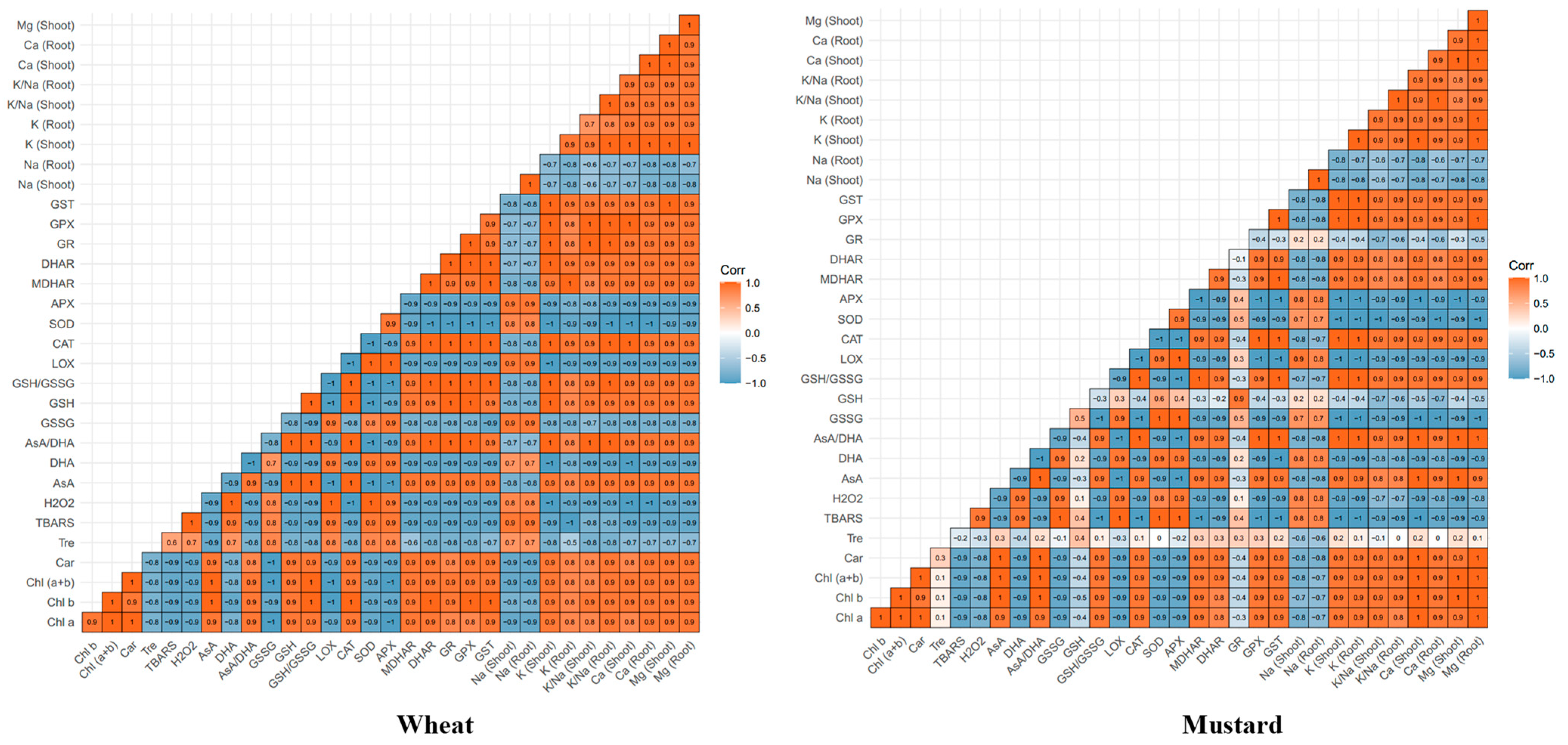
2.8. Relationship between Various Physio-Biochemical Parameters
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material and Growth Conditions
4.3. Measurement of Chlorophyll and Carotenoid Contents
4.4. Determination of Trehalose Content
4.5. Measurement of Thiobarbituric Acid-Reactive Substances Content
4.6. Hydrogen Peroxide Content Determination
4.7. Measurement of Ascorbate and Glutathione Contents
4.8. Soluble Protein and Antioxidant Enzyme Activity Measurement
4.9. Measurement of Mineral Contents
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muchate, N.S.; Rajurkar, N.S.; Suprasanna, P.; Nikam, T.D. NaCl induced salt adaptive changes and enhanced accumulation of 20-hydroxyecdysone in the in vitro shoot cultures of Spinacia oleracea (L.). Sci. Rep. 2019, 9, 12522. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, H.; Jamil, M.; Haq, A.; Ali, S.; Ahmad, R.; Malik, Z.; Parveen, Z. Salt stress manifestation on plants, mechanism of salt tolerance and potassium role in alleviating it: A review. Zemdirb. Agric. 2016, 103, 229–238. [Google Scholar] [CrossRef]
- Mohsin, S.M.; Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Parvin, K.; Fujita, M. Exogenous tebuconazole and trifloxystrobin regulates reactive oxygen species metabolism toward mitigating salt-induced damages in cucumber seedling. Plants 2019, 8, 428. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, S.M.; Hasanuzzaman, M.; Nahar, K.; Hossain, M.S.; Bhuyan, M.H.M.B.; Parvin, K.; Fujita, M. Tebuconazole and trifloxystrobin regulate the physiology, antioxidant defense and methylglyoxal detoxification systems in conferring salt stress tolerance in Triticum aestivum L. Physiol. Mol. Biol. Plants 2020, 26, 1139–1154. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.H.M.B.; Mahmud, J.A.; Baluska, F.; Fujita, M. Nitricoxide-induced salt stress tolerance in plants: ROS metabolism, signaling, and molecular interactions. Plant Biotechnol. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Mohsin, S.M.; Hasanuzzaman, M.; Parvin, K.; Fujita, M. Pretreatment of wheat(Triticum aestivum L.) seedlings with 2,4-D improves tolerance to salinity-induced oxidative stress and methylglyoxal toxicity by modulating ion homeostasis, antioxidant defenses, and glyoxalase systems. Plant Physiol. Biochem. 2020, 152, 221–231. [Google Scholar] [CrossRef]
- Shafiq, S.; Akram, N.A.; Ashraf, M.; Arshad, A. Synergistic effects of drought and ascorbic acid ongrowth, mineral nutrients and oxidative defense system in canola (Brassica napus L.) plants. Acta Physiol. Plant. 2014, 36, 1539–1553. [Google Scholar] [CrossRef]
- Samarah, N.; Mullen, R.; Cianzio, S. Size distribution and mineral nutrients of soybean seeds in response to drought stress. J. Plant Nutr. 2004, 27, 815–835. [Google Scholar] [CrossRef]
- Mahesh, K.; Balaraju, P.; Ramakrishna, B.; Rao, S.S.R. Effect of brassinosteroids on germination and seedling growth of radish (Raphanus sativus L.) under PEG-6000 induced water stress. Am. J. Plant Sci. 2013, 4, 2305–2313. [Google Scholar] [CrossRef]
- Ashraf, M.; Akram, N.A.; Al-Qurainy, F.; Foolad, M.R. Drought tolerance: Roles of organic osmolytes, growth regulators, and mineral nutrients. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Burlington, ON, Canada, 2011; Volume 111, pp. 249–296. [Google Scholar]
- Türkan, I.; Demiral, T. Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 2009, 67, 2–9. [Google Scholar] [CrossRef]
- Min, C.; Zhen, Y.; Jing, L.; Tingting, Z.; Xiaocen, W.; Hai, F.; Baoshan, W. Adaptation mechanism of salt excluders under saline conditions and its applications. Int. J. Mol. Sci. 2018, 19, 3668. [Google Scholar] [CrossRef]
- Liu, Q.Q.; Liu, R.R.; Ma, Y.C.; Song, J. Physiological and molecular evidence for Na+ and Cl− exclusion in the roots of two Suaeda salsa populations. Aquat. Bot. 2018, 146, 1–7. [Google Scholar] [CrossRef]
- Asaf, S.; Khan, A.L.; Khan, M.A.; Imran, Q.M.; Yun, B.W.; Lee, I.J. Osmoprotective functions conferred to soybean plants via inoculation with Sphingomona sp. LK11 and exogenous trehalose. Microbiol. Res. 2017, 205, 135–145. [Google Scholar] [CrossRef]
- Trouvelot, S.; Heloir, M.C.; Poinssot, B.; Gauthier, A.; Paris, F.; Guillier, C.; Combier, M.; Trda, L.; Daire, X.; Adrian, M. Carbohydrates in plant immunity and plant protection: Roles and potential application as foliar sprays. Front. Plant. Sci. 2014, 5, 592. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Kosar, F.; Akram, N.A.; Sadiq, M.; Al-Qurainy, F.; Ashraf, M. Trehalose: A key organic osmolyte effectively involved in plant abiotic stress tolerance. J. Plant Growth. Regul. 2019, 38, 606–618. [Google Scholar] [CrossRef]
- Fatemeh, K.; Nasrin, M.; Khosro, P. Increment of abiotic stress tolerance by metabolic engineering of compatible solutes. Int. J. Agric. Crop Sci. 2012, 4, 1856–1866. [Google Scholar]
- Paul, M. Trehalose 6-phosphate. Curr. Opin. Plant Biol. 2007, 10, 303–309. [Google Scholar] [CrossRef]
- Ponnu, J.; Wahl, V.; Schmid, M. Trehalose-6-phosphate: Connecting plant metabolism and development. Front. Plant Sci. 2011, 2, 70. [Google Scholar] [CrossRef]
- Nezhadahmadi, A.; Prodhan, Z.; Faruq, G. Drought tolerance in wheat. Sci. World J. 2013, 13, 1–12. [Google Scholar] [CrossRef]
- Ashraf, M.; Akram, N.A. Improving salinity tolerance of plants through conventional breeding and genetic engineering: Ananalytical comparison. Biotechnol. Adv. 2009, 27, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Shahbaz, M.; Abid, A.; Masood, A.; Waraich, E.A. Foliar-applied trehalose modulates growth, mineral nutrition, photosyntheticability, and oxidative defense system of rice (Oryza sativa L.) under saline stress. J. Plant Nutr. 2017, 40, 584–599. [Google Scholar] [CrossRef]
- Rohman, M.; Islam, M.; Monsur, M.B.; Amiruzzaman, M.; Fujita, M.; Hasanuzzaman, M. Trehalose protects maize plants from salt stress and phosphorus deficiency. Plants 2019, 8, 568. [Google Scholar] [CrossRef] [PubMed]
- Mittal, S.; Kumari, N.; Sharma, V. Differential response of salt stress on Brassica juncea: Photosynthetic performance, pigment, proline, D1 and antioxidant enzymes. Plant Physiol. Biochem. 2012, 54, 17–26. [Google Scholar] [CrossRef]
- Mahmud, J.A.; Hasanuzzaman, M.; Khan, M.I.R.; Nahar, K.; Fujita, M. β-Amino butyric acid pretreatment confers salt stress tolerance in Brassica napus L. by modulating reactive oxygen species metabolism and methylglyoxal detoxification. Plants 2020, 9, 241. [Google Scholar] [CrossRef]
- Alam, M.M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Trehalose-induced drought stress tolerance: A comparative study among different Brassica species. Plant Omics 2014, 7, 271–283. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Inafuku, M.; Oku, H.; Fujita, M. Exogenous nitric oxide donor and arginine provide protection against short-term drought stress in wheat seedlings. Physiol. Mol. Biol. Plants 2018, 24, 993–1004. [Google Scholar] [CrossRef]
- Sadak, M.S. Physiological role of trehalose on enhancing salinity tolerance of wheat plant. Bull. Natl. Res. Cent. 2019, 43, 53. [Google Scholar] [CrossRef]
- Krasensky, J.; Broyart, C.; Rabanal, F.A.; Jonak, C. The redox-sensitive chloroplast trehalose-6-phosphate phosphatase AtTPPD regulates salt stress tolerance. Antioxid. Redox Signal. 2014, 21, 1289–1304. [Google Scholar] [CrossRef]
- Ozfidan-Konakci, C.; Yildiztugay, E.; Kucukoduk, M. Upregulation of antioxidant enzymes by exogenous gallic acid contributes to the amelioration in Oryza sativa roots exposed to salt and osmotic stress. Environ. Sci. Poll. Res. 2015, 22, 1487–1498. [Google Scholar] [CrossRef]
- Ahmad, P.; Jhon, R.; Sarwat, M.; Umar, S. Responses of proline, lipid peroxidation and antioxidative enzymes in two varieties of Pisum sativum L. under salt stress. Int. J. Plant Prod. 2008, 2, 353–366. [Google Scholar]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Yadav, S.; Hasanuzzaman, M.; Fujita, M.; Mishra, P.; Sabat, S.C.; Tuteja, N. Superoxide dismutase—mentor of abiotic stress tolerance in crop plants. Environ. Sci. Pollut. Res. 2015, 22, 10375–10394. [Google Scholar] [CrossRef] [PubMed]
- Bhuyan, M.H.M.B.; Parvin, K.; Mohsin, S.M.; Mahmud, J.A.; Hasanuzzaman, M.; Fujita, M. Modulation of cadmium tolerance in rice: Insight into vanillic acid-induced upregulation of antioxidant defense and glyoxalase systems. Plants 2020, 9, 188. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.S.A.; Javed, F.; Ashraf, M. Iso-osmotic effect of NaCl and PEG on growth, cations and free proline accumulation in callus tissue of two indica rice(Oryza sativa L.) genotypes. Plant Growth Regul. 2007, 53, 53–63. [Google Scholar] [CrossRef]
- Mohsin, S.M.; Hasanuzzaman, M.; Parvin, K.; Hossain, M.S.; Fujita, M. Protective role of tebuconazole and trifloxystrobin in wheat (Triticum aestivum L.) under cadmium stress via enhancement of antioxidant defense and glyoxalase systems. Physiol. Mol. Biol. Plants 2021, 27, 1043–1057. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Packer, L., Douce, R., Eds.; Academic Press: Cambridge, UK, 1987; pp. 350–382. [Google Scholar]
- Li, Z.G.; Luo, L.J.; Zhu, L.P. Involvement of trehalose in hydrogen sulfide donor sodium hydrosulfide induced the acquisition of heat tolerance in maize (Zea mays L.) seedlings. Bot. Stud. 2014, 55, 20. [Google Scholar] [CrossRef]
- Taulavuori, E.; Hellström, E.K.; Taulavuori, K.; Laine, K. Comparison of two methods used to analyze lipid peroxidation from Vaccinium myrtillus (L.) during snow removal, reacclimation and cold acclimation. J. Exp. Bot. 2001, 52, 2375–2380. [Google Scholar] [CrossRef]
- Yu, C.W.; Murphy, T.M.; Lin, C.H. Hydrogen peroxide-induces chilling tolerance in mung beans mediated through ABA-independent glutathione accumulation. Funct. Plant Biol. 2003, 30, 955–963. [Google Scholar] [CrossRef]
- Lechno, S.; Zamski, E.; Tel-Or, E. Salt stress-induced responses in cucumber plants. J. Plant Physiol. 1997, 150, 206–211. [Google Scholar] [CrossRef]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. The effect of hydrogen peroxide and of paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Doderer, A.; Kokkelink, I.; VanderVeen, S.; Valk, B.; Schram, A.; Douma, A. Purification and characterization of two lipoygenase isoenzymes from germinating barley. Biochim. Biophys. Acta 1992, 112, 97–104. [Google Scholar] [CrossRef]
- El-Shabrawi, H.; Kumar, B.; Kaul, T.; Reddy, M.K.; Singla-Pareek, S.L.; Sopory, S.K. Redox homeostasis, antioxidant defense, and methylglyoxal detoxification as markers for salt tolerance in Pokkali rice. Protoplasma 2010, 245, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Alam, M.M.; Nahar, K.; Mohsin, S.M.; Bhuyan, M.H.M.B.; Parvin, K.; Hawrylak-Nowak, B.; Fujita, M. Silicon-induced antioxidant defense and methylglyoxal detoxification works coordinately in alleviating nickel toxicity in Oryza sativa L. Ecotoxicology 2019, 28, 261–276. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Mohsin, S.M.; Hasanuzzaman, M.; Parvin, K.; Fujita, M. Effect of tebuconazole and trifloxystrobin on Ceratocystis fimbriata to control black rot of sweet potato: Processes of reactive oxygen species generation and antioxidant defense responses. World J. Microbiol. Biotechnol. 2021, 37, 148. [Google Scholar] [CrossRef]
- Zasoski, R.J.; Burau, R.G. A rapid nitric-perchloric acid digestion method for multi-element tissue analysis. Commun. Soil. Sci. Plant Anal. 1977, 8, 425–436. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Chlorophyll a Content (mg g−1 FW) | Chlorophyll b Content (mg g−1 FW) | Chlorophyll (a + b) Content (mg g−1 FW) | Carotenoid Content (mg g−1 FW) | Trehalose Content (µmol g–1 FW) | |
|---|---|---|---|---|---|---|
| No stress | C | 9.21 ± 0.29 a | 4.99 ± 0.16 a | 14.20 ± 0.26 a | 6.07 ± 0.25 a | 10.99 ± 0.59 e |
| Tre | 8.13 ± 0.22 bc | 4.13 ± 0.43 c | 12.26 ± 0.23 c | 5.18 ± 0.22 bc | 11.82 ± 0.72 de | |
| T6P | 8.46 ± 0.19 b | 4.58 ± 0.19 b | 13.04 ± 0.03 b | 5.53 ± 0.08 b | 11.48 ± 0.49 e | |
| Salinity | S | 3.14 ± 0.41 g | 1.32 ± 0.19 f | 4.45 ± 0.51 i | 1.36 ± 0.15 g | 15.11 ± 0.51 b |
| S + Tre | 4.11 ± 0.26 f | 1.60 ± 0.08 f | 5.72 ± 0.26 h | 1.81 ± 0.19 f | 16.97 ± 0.71 a | |
| S + T6P | 4.52 ± 0.36 f | 2.06 ± 0.13 e | 6.58 ± 0.43 g | 2.07 ± 0.12 f | 15.93 ± 0.40 ab | |
| Water deficit | D | 6.34 ± 0.30 e | 2.32 ± 0.23 e | 8.67 ± 0.37 f | 3.38 ± 0.17 e | 12.97 ± 1.02 cd |
| D + Tre | 6.94 ± 0.43 d | 3.03 ± 0.19 d | 9.96 ± 0.40 e | 4.49 ± 0.32 d | 15.15 ± 0.67 b | |
| D + T6P | 7.79 ± 0.30 c | 3.29 ± 0.19 d | 11.08 ± 0.23 d | 5.02 ± 0.23 c | 13.70 ± 0.72 c | |
| Treatments | Chlorophyll a Content (mg g−1 FW) | Chlorophyll b Content (mg g−1FW) | Chlorophyll (a + b) Content (mg g−1 FW) | Carotenoid Content (mg g−1 FW) | Trehalose Content (µmol g–1 FW) | |
|---|---|---|---|---|---|---|
| No stress | C | 4.89 ± 0.18 b | 2.06 ± 0.08 b | 6.99 ± 0.20 b | 3.46 ± 0.16 b | 6.04 ± 0.44 de |
| Tre | 5.16 ± 0.15 b | 2.19 ± 0.10 b | 7.28 ± 0.15 b | 3.68 ± 0.10 ab | 6.64 ± 0.67 cd | |
| T6P | 5.80 ± 0.21 a | 2.53 ± 0.13 a | 8.26 ± 0.24 a | 3.83 ± 0.12 a | 6.33 ± 0.66 de | |
| Salinity | S | 2.47 ± 0.16 f | 1.10 ± 0.10 f | 3.61 ± 0.20 f | 1.93 ± 0.12 g | 4.90 ± 0.47 f |
| S + Tre | 2.66 ± 0.14 f | 1.27 ± 0.08 e | 4.13 ± 0.17 e | 2.12 ± 0.10 fg | 5.81 ± 0.34 de | |
| S + T6P | 3.69 ± 0.19 d | 1.44 ± 0.05 d | 5.13 ± 0.16 d | 2.29 ± 0.10 ef | 5.55 ± 0.50 ef | |
| Water deficit | D | 3.08 ± 0.15 e | 1.22 ± 0.11 ef | 4.23 ± 0.20 e | 2.47 ± 0.15 e | 7.41 ± 0.45 bc |
| D + Tre | 3.42 ± 0.19 d | 1.56 ± 0.07 d | 5.22 ± 0.17 d | 2.81 ± 0.11 d | 8.79 ± 0.33 a | |
| D + T6P | 4.44 ± 0.22 c | 1.76 ± 0.10 c | 6.23 ± 0.25 c | 3.23 ± 0.13 c | 8.07 ± 0.44 ab | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohsin, S.M.; Fardus, J.; Nagata, A.; Tamano, N.; Mitani, H.; Fujita, M. Comparative Study of Trehalose and Trehalose 6-Phosphate to Improve Antioxidant Defense Mechanisms in Wheat and Mustard Seedlings under Salt and Water Deficit Stresses. Stresses 2022, 2, 336-354. https://doi.org/10.3390/stresses2030024
Mohsin SM, Fardus J, Nagata A, Tamano N, Mitani H, Fujita M. Comparative Study of Trehalose and Trehalose 6-Phosphate to Improve Antioxidant Defense Mechanisms in Wheat and Mustard Seedlings under Salt and Water Deficit Stresses. Stresses. 2022; 2(3):336-354. https://doi.org/10.3390/stresses2030024
Chicago/Turabian StyleMohsin, Sayed Mohammad, Jannatul Fardus, Atsushi Nagata, Nobuhisa Tamano, Hirofumi Mitani, and Masayuki Fujita. 2022. "Comparative Study of Trehalose and Trehalose 6-Phosphate to Improve Antioxidant Defense Mechanisms in Wheat and Mustard Seedlings under Salt and Water Deficit Stresses" Stresses 2, no. 3: 336-354. https://doi.org/10.3390/stresses2030024
APA StyleMohsin, S. M., Fardus, J., Nagata, A., Tamano, N., Mitani, H., & Fujita, M. (2022). Comparative Study of Trehalose and Trehalose 6-Phosphate to Improve Antioxidant Defense Mechanisms in Wheat and Mustard Seedlings under Salt and Water Deficit Stresses. Stresses, 2(3), 336-354. https://doi.org/10.3390/stresses2030024