Plasma Cell-Free Tumor Methylome as a Biomarker in Solid Tumors: Biology and Applications


Abstract

1. Introduction
2. DNA Methylation in Cancer
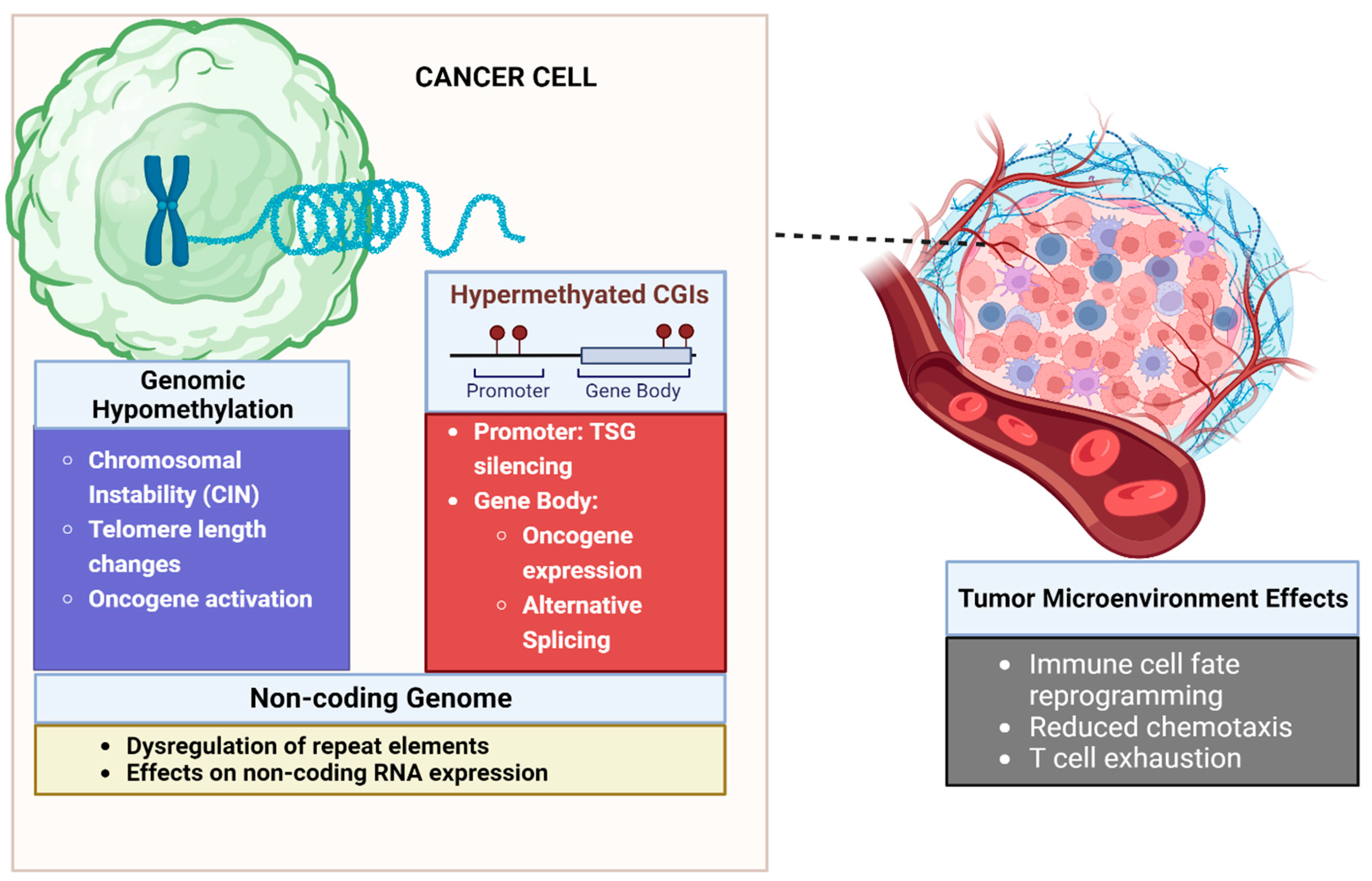
2.1. Biology of DNA Methylation in Malignancy
2.1.1. Aberrant DNA Methylation of CpG Islands Are Seen in Cancer
2.1.2. Gene Promoter Hypermethylation Leads to Silencing of Tumor Suppressor Genes
2.1.3. Methylation Can Lead to Epigenetic Activation of Oncogenic Pathways
2.1.4. Global DNA Hypomethylation Leads to Chromosomal Instability
2.1.5. DNA Methylation Influences Gene Expression beyond Promoter Silencing
2.1.6. Methylation Can Influence the Non-Coding Genome, Leading to Cancer
2.1.7. Role of DNA Methylation in the Tumor Microenvironment
2.2. Cell-Free Tumor Methylome Recapitulates the Cancer Epigenome
2.3. Applications of cfDNA Methylation as Biomarker in Cancer
2.3.1. Methylation as a Diagnostic Biomarker
2.3.2. Methylation as a Prognostic Biomarker of Patient Outcomes
2.3.3. Methylation as a Predictive Biomarker of Treatment Response
2.3.4. Minimal Residual Disease
3. Challenges and Future Directions
3.1. DNA Methylation in Oncology
3.2. Liquid Biopsy in Oncology
3.3. ctDNA Methylation as a Biomarker in Oncology
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Narrative Review Search Strategy
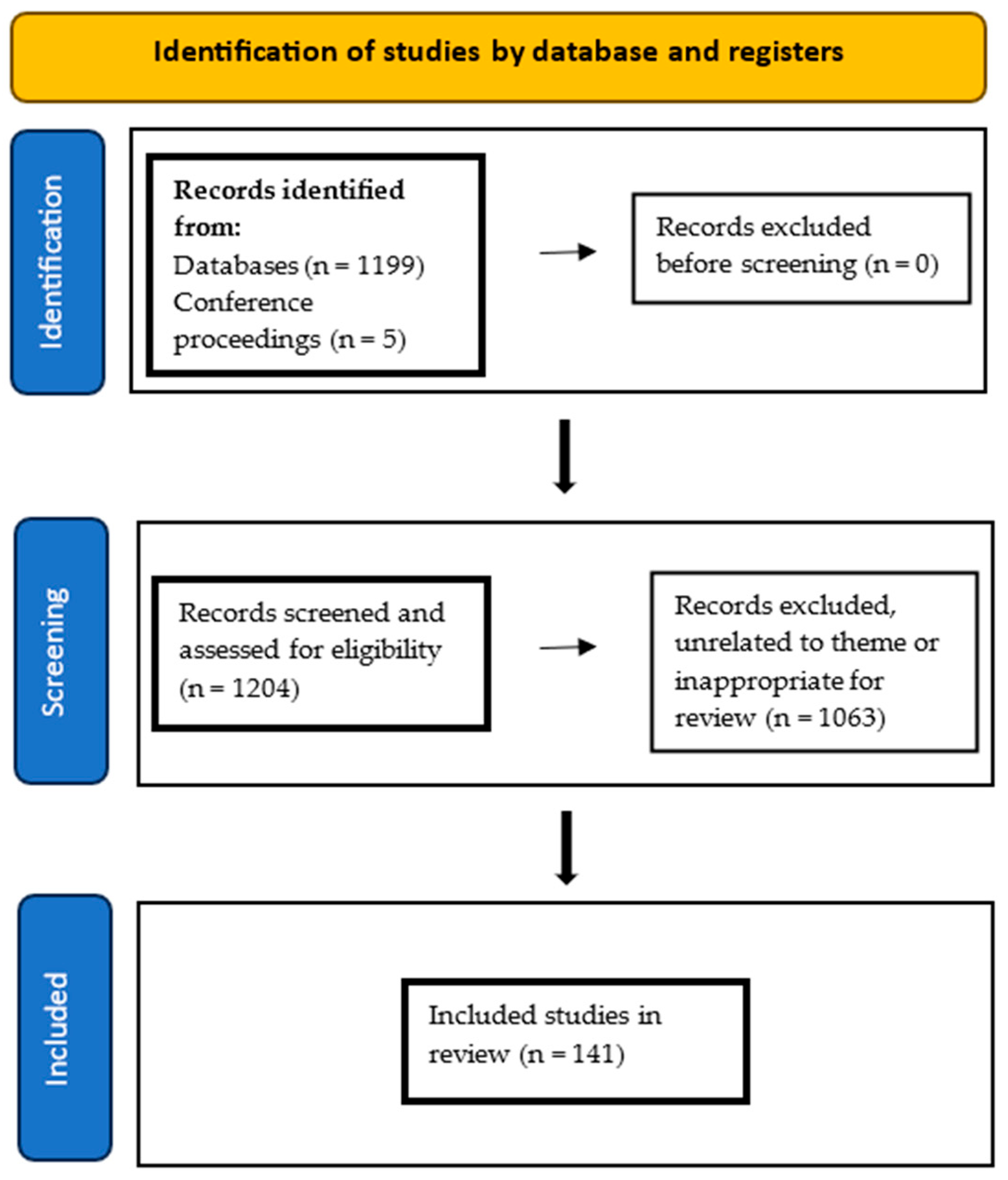
References
- Wang, Q.; Xiong, F.; Wu, G.; Liu, W.; Chen, J.; Wang, B.; Chen, Y. Gene Body Methylation in Cancer: Molecular Mechanisms and Clinical Applications. Clin. Epigenetics 2022, 14, 154. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Nakanishi, M. Navigating the DNA Methylation Landscape of Cancer. Trends Genet. 2021, 37, 1012–1027. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T.; Takehara, K.; Sugimoto, N.; Kaneko, K.; Fujimoto, E.; Okazawa-Sakai, M.; Okame, S.; Shiroyama, Y.; Yokoyama, T.; Teramoto, N.; et al. Lynch Syndrome-Associated Endometrial Carcinoma with MLH1 rmline Mutation and MLH1 Promoter Hypermethylation: A Case Report and Literature Review. BMC Cancer 2018, 18, 576. [Google Scholar] [CrossRef] [PubMed]
- Joo, J.E.; Mahmood, K.; Walker, R.; Georgeson, P.; Candiloro, I.; Clendenning, M.; Como, J.; Joseland, S.; Preston, S.; Graversen, L.; et al. Identifying Primary and Secondary MLH1 Epimutation Carriers Displaying Low-Level Constitutional MLH1 Methylation Using Droplet Digital PCR and Genome-Wide DNA Methylation Profiling of Colorectal Cancers. Clin. Epigenetics 2023, 15, 95. [Google Scholar] [CrossRef]
- Baylin, S.B.; Jones, P.A. Epigenetic Determinants of Cancer. Cold Spring Harb. Perspect. Biol. 2016, 8, a019505. [Google Scholar] [CrossRef]
- Yates, J.; Boeva, V. Deciphering the Etiology and Role in Oncogenic Transformation of the CpG Island Methylator Phenotype: A Pan-Cancer Analysis. Brief. Bioinform. 2022, 23, bbab610. [Google Scholar] [CrossRef]
- Koudonas, A.; Papaioannou, M.; Kampantais, S.; Anastasiadis, A.; Hatzimouratidis, K.; Dimitriadis, G. Methylation of PCDH17 and NEFH as Prognostic Biomarker for Nonmetastatic RCC: A Cohort Study. Medicine 2022, 101, e29599. [Google Scholar] [CrossRef]
- Li, J.; Xu, C.; Lee, H.J.; Ren, S.; Zi, X.; Zhang, Z.; Wang, H.; Yu, Y.; Yang, C.; Gao, X.; et al. A Genomic and Epigenomic Atlas of Prostate Cancer in Asian Populations. Nature 2020, 580, 93–99. [Google Scholar] [CrossRef]
- Suman, M.; Dugué, P.-A.; Wong, E.M.; Joo, J.E.; Hopper, J.L.; Nguyen-Dumont, T.; Giles, G.G.; Milne, R.L.; McLean, C.; Southey, M.C. Association of Variably Methylated Tumour DNA Regions with Overall Survival for Invasive Lobular Breast Cancer. Clin. Epigenetics 2021, 13, 11. [Google Scholar] [CrossRef]
- Shiu, B.-H.; Lu, W.-Y.; Tantoh, D.M.; Chou, M.-C.; Nfor, O.N.; Huang, C.-C.; Liaw, Y.-P. Interactive Association between Dietary Fat and Sex on CDH13 Cg02263260 Methylation. BMC Med. Genom. 2021, 14, 13. [Google Scholar] [CrossRef]
- Chen, S.; Petricca, J.; Ye, W.; Guan, J.; Zeng, Y.; Cheng, N.; Gong, L.; Shen, S.Y.; Hua, J.T.; Crumbaker, M.; et al. The Cell-Free DNA Methylome Captures Distinctions between Localized and Metastatic Prostate Tumors. Nat. Commun. 2022, 13, 6467. [Google Scholar] [CrossRef] [PubMed]
- Romero-Garcia, S.; Prado-Garcia, H.; Carlos-Reyes, A. Role of DNA Methylation in the Resistance to Therapy in Solid Tumors. Front. Oncol. 2020, 10, 1152. [Google Scholar] [CrossRef]
- McLachlan, T.; Matthews, W.C.; Jackson, E.R.; Staudt, D.E.; Douglas, A.M.; Findlay, I.J.; Persson, M.L.; Duchatel, R.J.; Mannan, A.; Germon, Z.P.; et al. B-Cell Lymphoma 6 (BCL6): From Master Regulator of Humoral Immunity to Oncogenic Driver in Pediatric Cancers. Mol. Cancer Res. 2022, 20, 1711–1723. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Sheng, Q.; Zhao, N.; Huang, S.; Zhao, Y. DNA Hypomethylation Mediates Immune Response in Pan-Cancer. Epigenetics 2023, 18, 2192894. [Google Scholar] [CrossRef] [PubMed]
- Ankill, J.; Aure, M.R.; Bjørklund, S.; Langberg, S.; Kristensen, V.N.; Vitelli, V.; Tekpli, X.; Fleischer, T. Epigenetic Alterations at Distal Enhancers Are Linked to Proliferation in Human Breast Cancer. NAR Cancer 2022, 4, zcac008. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.-W.; Shim, H.S.; Lee, C.Y.; Park, S.Y.; Hong, M.H.; Lee, I.; Kim, H.R. The Importance of Enhancer Methylation for Epigenetic Regulation of Tumorigenesis in Squamous Lung Cancer. Exp. Mol. Med. 2022, 54, 12–22. [Google Scholar] [CrossRef]
- Pongor, L.S.; Tlemsani, C.; Elloumi, F.; Arakawa, Y.; Jo, U.; Gross, J.M.; Mosavarpour, S.; Varma, S.; Kollipara, R.K.; Roper, N.; et al. Integrative Epigenomic Analyses of Small Cell Lung Cancer Cells Demonstrates the Clinical Translational Relevance of Gene Body Methylation. iScience 2022, 25, 105338. [Google Scholar] [CrossRef]
- Yang, X.; Han, H.; De Carvalho, D.D.; Lay, F.D.; Jones, P.A.; Liang, G. Gene Body Methylation Can Alter Gene Expression and Is a Therapeutic Target in Cancer. Cancer Cell 2014, 26, 577–590. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, T.; Wang, Y.; Xie, M.; Ji, X.; Luo, X.; Huang, W.; Xia, L. Homeobox Genes in Cancers: From Carcinogenesis to Recent Therapeutic Intervention. Front. Oncol. 2021, 11, 770428. [Google Scholar] [CrossRef]
- Su, J.; Huang, Y.-H.; Cui, X.; Wang, X.; Zhang, X.; Lei, Y.; Xu, J.; Lin, X.; Chen, K.; Lv, J.; et al. Homeobox Oncogene Activation by Pan-Cancer DNA Hypermethylation. Genome Biol. 2018, 19, 108. [Google Scholar] [CrossRef]
- Besselink, N.; Keijer, J.; Vermeulen, C.; Boymans, S.; de Ridder, J.; van Hoeck, A.; Cuppen, E.; Kuijk, E. The Genome-Wide Mutational Consequences of DNA Hypomethylation. Sci. Rep. 2023, 13, 6874. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Ishak, C.A.; De Carvalho, D.D. Endogenous Retroelements and the Viral Mimicry Response in Cancer Therapy and Cellular Homeostasis. Cancer Discov. 2021, 11, 2707–2725. [Google Scholar] [CrossRef] [PubMed]
- García-Muse, T.; Aguilera, A. R Loops: From Physiological to Pathological Roles. Cell 2019, 179, 604–618. [Google Scholar] [CrossRef] [PubMed]
- Tire, B.; Ozturk, S. Potential Effects of Assisted Reproductive Technology on Telomere Length and Telomerase Activity in Human Oocytes and Early Embryos. J. Ovarian Res. 2023, 16, 130. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Seiler, M.; Peng, S.; Agrawal, A.A.; Palacino, J.; Teng, T.; Zhu, P.; Smith, P.G.; Caesar-Johnson, S.J.; Demchok, J.A.; Felau, I.; et al. Somatic Mutational Landscape of Splicing Factor Genes and Their Functional Consequences across 33 Cancer Types. Cell Rep. 2018, 23, 282–296.e4. [Google Scholar] [CrossRef]
- Bonnal, S.C.; López-Oreja, I.; Valcárcel, J. Roles and Mechanisms of Alternative Splicing in Cancer—Implications for Care. Nat. Rev. Clin. Oncol. 2020, 17, 457–474. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Elnitski, L. Aberrant DNA Methylation Defines Isoform Usage in Cancer, with Functional Implications. PLoS Comput. Biol. 2019, 15, e1007095. [Google Scholar] [CrossRef]
- Jang, H.S.; Shah, N.M.; Du, A.Y.; Dailey, Z.Z.; Pehrsson, E.C.; Godoy, P.M.; Zhang, D.; Li, D.; Xing, X.; Kim, S.; et al. Transposable Elements Drive Widespread Expression of Oncogenes in Human Cancers. Nat. Genet. 2019, 51, 611–617. [Google Scholar] [CrossRef]
- Espinet, E.; Gu, Z.; Imbusch, C.D.; Giese, N.A.; Büscher, M.; Safavi, M.; Weisenburger, S.; Klein, C.; Vogel, V.; Falcone, M.; et al. Aggressive PDACs Show Hypomethylation of Repetitive Elements and the Execution of an Intrinsic IFN Program Linked to a Ductal Cell of Origin. Cancer Discov. 2021, 11, 638–659. [Google Scholar] [CrossRef]
- Tavora, B.; Mederer, T.; Wessel, K.J.; Ruffing, S.; Sadjadi, M.; Missmahl, M.; Ostendorf, B.N.; Liu, X.; Kim, J.-Y.; Olsen, O.; et al. Tumoural Activation of TLR3–SLIT2 Axis in Endothelium Drives Metastasis. Nature 2020, 586, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Dang, H.X.; Lim, D.A.; Feng, F.Y.; Maher, C.A. Long Noncoding RNAs in Cancer Metastasis. Nat. Rev. Cancer 2021, 21, 446–460. [Google Scholar] [CrossRef] [PubMed]
- Bunch, H. Gene Regulation of Mammalian Long Non-Coding RNA. Mol. Genet. Genom. 2018, 293, 1–15. [Google Scholar] [CrossRef]
- Yang, Z.; Xu, F.; Wang, H.; Teschendorff, A.E.; Xie, F.; He, Y. Pan-Cancer Characterization of Long Non-Coding RNA and DNA Methylation Mediated Transcriptional Dysregulation. EBioMedicine 2021, 68, 103399. [Google Scholar] [CrossRef]
- Li, S.; Hu, J.; Li, G.; Mai, H.; Gao, Y.; Liang, B.; Wu, H.; Guo, J.; Duan, Y. Epigenetic Regulation of LINC01270 in Breast Cancer Progression by Mediating LAMA2 Promoter Methylation and MAPK Signaling Pathway. Cell Biol. Toxicol. 2023, 39, 1359–1375. [Google Scholar] [CrossRef]
- Xu, X.; Yuan, X.; Ni, J.; Guo, J.; Gao, Y.; Yin, W.; Li, F.; Wei, L.; Zhang, J. MAGI2-AS3 Inhibits Breast Cancer by Downregulating DNA Methylation of MAGI2. J. Cell. Physiol. 2021, 236, 1116–1130. [Google Scholar] [CrossRef]
- Al-Imam, M.J.; Hussein, U.A.-R.; Sead, F.F.; Faqri, A.M.A.; Mekkey, S.M.; Khazel, A.J.; Almashhadani, H.A. The Interactions between DNA Methylation Machinery and Long Non-Coding RNAs in Tumor Progression and Drug Resistance. DNA Repair 2023, 128, 103526. [Google Scholar] [CrossRef] [PubMed]
- Sideris, N.; Dama, P.; Bayraktar, S.; Stiff, T.; Castellano, L. LncRNAs in Breast Cancer: A Link to Future Approaches. Cancer Gene Ther. 2022, 29, 1866–1877. [Google Scholar] [CrossRef]
- Shen, S.; Chen, J.; Li, H.; Jiang, Y.; Wei, Y.; Zhang, R.; Zhao, Y.; Chen, F. Large-Scale Integration of the Non-Coding RNAs with DNA Methylation in Human Cancers. Cell Rep. 2023, 42, 112261. [Google Scholar] [CrossRef]
- Zhong, F.; Lin, Y.; Zhao, L.; Yang, C.; Ye, Y.; Shen, Z. Reshaping the Tumour Immune Microenvironment in Solid Tumours via Tumour Cell and Immune Cell DNA Methylation: From Mechanisms to Therapeutics. Br. J. Cancer 2023, 129, 24–37. [Google Scholar] [CrossRef]
- Sun, L.; Su, Y.; Jiao, A.; Wang, X.; Zhang, B. T Cells in Health and Disease. Signal Transduct. Target. Ther. 2023, 8, 235. [Google Scholar] [CrossRef] [PubMed]
- Henning, A.N.; Roychoudhuri, R.; Restifo, N.P. Epigenetic Control of CD8+ T Cell Differentiation. Nat. Rev. Immunol. 2018, 18, 340–356. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ubreva, J.; Català-Moll, F.; Obermajer, N.; Álvarez-Errico, D.; Ramirez, R.N.; Company, C.; Vento-Tormo, R.; Moreno-Bueno, G.; Edwards, R.P.; Mortazavi, A.; et al. Prostaglandin E2 Leads to the Acquisition of DNMT3A-Dependent Tolerogenic Functions in Human Myeloid-Derived Suppressor Cells. Cell Rep. 2017, 21, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zhao, E.; Yu, H.; Yuan, C.; Abbas, M.N.; Cui, H. Methylation across the Central Dogma in Health and Diseases: New Therapeutic Strategies. Signal Transduct. Target. Ther. 2023, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cao, Q.; Yu, L.; Shi, H.; Xue, B.; Shi, H. Epigenetic Regulation of Macrophage Polarization and Inflammation by DNA Methylation in Obesity. JCI Insight 2016, 1, e87748. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, T.; Tang, Y.; Luo, G.; Liang, G.; He, W. Epigenetic Regulation of Macrophage Polarization in Wound Healing. Burns Trauma 2023, 11, tkac057. [Google Scholar] [CrossRef] [PubMed]
- Vadevoo, S.M.P.; Gunassekaran, G.R.; Yoo, J.D.; Kwon, T.-H.; Hur, K.; Chae, S.; Lee, B. Epigenetic Therapy Reprograms M2-Type Tumor-Associated Macrophages into an M1-like Phenotype by Upregulating MiR-7083-5p. Front. Immunol. 2022, 13, 976196. [Google Scholar] [CrossRef]
- Dangaj, D.; Bruand, M.; Grimm, A.J.; Ronet, C.; Barras, D.; Duttagupta, P.A.; Lanitis, E.; Duraiswamy, J.; Tanyi, J.L.; Benencia, F.; et al. Cooperation between Constitutive and Inducible Chemokines Enables T Cell Engraftment and Immune Attack in Solid Tumors. Cancer Cell 2019, 35, 885–900.e10. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, Z.; Wei, S.; Liu, Z.; Chen, G. Epigenetic Silencing of Chemokine CCL2 Represses Macrophage Infiltration to Potentiate Tumor Development in Small Cell Lung Cancer. Cancer Lett. 2021, 499, 148–163. [Google Scholar] [CrossRef]
- Khan, P.; Fatima, M.; Khan, M.A.; Batra, S.K.; Nasser, M.W. Emerging Role of Chemokines in Small Cell Lung Cancer: Road Signs for Metastasis, Heterogeneity, and Immune Response. Semin. Cancer Biol. 2022, 87, 117–126. [Google Scholar] [CrossRef]
- Chen, X.; Pan, X.; Zhang, W.; Guo, H.; Cheng, S.; He, Q.; Yang, B.; Ding, L. Epigenetic Strategies Synergize with PD-L1/PD-1 Targeted Cancer Immunotherapies to Enhance Antitumor Responses. Acta Pharm. Sin. B 2020, 10, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Prinzing, B.; Zebley, C.C.; Petersen, C.T.; Fan, Y.; Anido, A.A.; Yi, Z.; Nguyen, P.; Houke, H.; Bell, M.; Haydar, D.; et al. Deleting DNMT3A in CAR T Cells Prevents Exhaustion and Enhances Antitumor Activity. Sci. Transl. Med. 2023, 13, eabh0272. [Google Scholar] [CrossRef]
- Yang, R.; Cheng, S.; Luo, N.; Gao, R.; Yu, K.; Kang, B.; Wang, L.; Zhang, Q.; Fang, Q.; Zhang, L.; et al. Distinct Epigenetic Features of Tumor-Reactive CD8+ T Cells in Colorectal Cancer Patients Revealed by Genome-Wide DNA Methylation Analysis. Genome Biol. 2019, 21, 2. [Google Scholar] [CrossRef]
- Wan, J.C.M.; Massie, C.; Garcia-Corbacho, J.; Mouliere, F.; Brenton, J.D.; Caldas, C.; Pacey, S.; Baird, R.; Rosenfeld, N. Liquid Biopsies Come of Age: Towards Implementation of Circulating Tumour DNA. Nat. Rev. Cancer 2017, 17, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Gaitsch, H.; Franklin, R.J.M.; Reich, D.S. Cell-Free DNA-Based Liquid Biopsies in Neurology. Brain 2023, 146, 1758–1774. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, R.B.; Chabner, B.A. Application of Cell-Free DNA Analysis to Cancer Treatment. N. Engl. J. Med. 2018, 379, 1754–1765. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.T.; Almeda, A.; Schauer, M.; McNamara, M.; Bai, X.; Meng, Q.; Partha, M.; Grimes, S.M.; Lee, H.; Heestand, G.M.; et al. Single-Molecule Methylation Profiles of Cell-Free DNA in Cancer with Nanopore Sequencing. Genome Med. 2023, 15, 33. [Google Scholar] [CrossRef]
- Cescon, D.W.; Bratman, S.V.; Chan, S.M.; Siu, L.L. Circulating Tumor DNA and Liquid Biopsy in Oncology. Nat. Cancer 2020, 1, 276–290. [Google Scholar] [CrossRef]
- Igari, F.; Tanaka, H.; Giuliano, A.E. The Applications of Plasma Cell-Free DNA in Cancer Detection: Implications in the Management of Breast Cancer Patients. Crit. Rev. Oncol. Hematol. 2022, 175, 103725. [Google Scholar] [CrossRef]
- Smith, J.T.; Balar, A.; Lakhani, D.A.; Kluwe, C.; Zhao, Z.; Kopparapu, P.; Almodovar, K.; Muterspaugh, A.; Yan, Y.; York, S.; et al. Circulating Tumor DNA as a Biomarker of Radiographic Tumor Burden in SCLC. JTO Clin. Res. Rep. 2021, 2, 100110. [Google Scholar] [CrossRef]
- Mohan, S.; Foy, V.; Ayub, M.; Leong, H.S.; Schofield, P.; Sahoo, S.; Descamps, T.; Kilerci, B.; Smith, N.K.; Carter, M.; et al. Profiling of Circulating Free DNA Using Targeted and Genome-Wide Sequencing in Patients with SCLC. J. Thorac. Oncol. 2020, 15, 216–230. [Google Scholar] [CrossRef] [PubMed]
- Almodovar, K.; Iams, W.T.; Meador, C.B.; Zhao, Z.; York, S.; Horn, L.; Yan, Y.; Hernandez, J.; Chen, H.; Shyr, Y.; et al. Longitudinal Cell-Free DNA Analysis in Patients with Small Cell Lung Cancer Reveals Dynamic Insights into Treatment Efficacy and Disease Relapse. J. Thorac. Oncol. 2018, 13, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Tolmeijer, S.H.; Boerrigter, E.; Sumiyoshi, T.; Kwan, E.M.; Ng, S.W.S.; Annala, M.; Donnellan, G.; Herberts, C.; Benoist, G.E.; Hamberg, P.; et al. Early On-Treatment Changes in Circulating Tumor DNA Fraction and Response to Enzalutamide or Abiraterone in Metastatic Castration-Resistant Prostate Cancer. Clin. Cancer Res. 2023, 29, 2835–2844. [Google Scholar] [CrossRef]
- Semenkovich, N.P.; Szymanski, J.J.; Earland, N.; Chauhan, P.S.; Pellini, B.; Chaudhuri, A.A. Genomic Approaches to Cancer and Minimal Residual Disease Detection Using Circulating Tumor DNA. J. Immunother. Cancer 2023, 11, e006284. [Google Scholar] [CrossRef] [PubMed]
- Gatto, L.; Franceschi, E.; Di Nunno, V.; Tosoni, A.; Lodi, R.; Brandes, A.A. Liquid Biopsy in Glioblastoma Management: From Current Research to Future Perspectives. Oncologist 2021, 26, 865–878. [Google Scholar] [CrossRef]
- Mair, R.; Mouliere, F. Cell-Free DNA Technologies for the Analysis of Brain Cancer. Br. J. Cancer 2022, 126, 371–378. [Google Scholar] [CrossRef]
- Liu, L.; Toung, J.M.; Jassowicz, A.F.; Vijayaraghavan, R.; Kang, H.; Zhang, R.; Kruglyak, K.M.; Huang, H.J.; Hinoue, T.; Shen, H.; et al. Targeted Methylation Sequencing of Plasma Cell-Free DNA for Cancer Detection and Classification. Ann. Oncol. 2018, 29, 1445–1453. [Google Scholar] [CrossRef]
- Sadeh, R.; Sharkia, I.; Fialkoff, G.; Rahat, A.; Gutin, J.; Chappleboim, A.; Nitzan, M.; Fox-Fisher, I.; Neiman, D.; Meler, G.; et al. ChIP-Seq of Plasma Cell-Free Nucleosomes Identifies Gene Expression Programs of the Cells of Origin. Nat. Biotechnol. 2021, 39, 586–598. [Google Scholar] [CrossRef]
- Baca, S.C.; Seo, J.-H.; Davidsohn, M.P.; Fortunato, B.; Semaan, K.; Sotudian, S.; Lakshminarayanan, G.; Diossy, M.; Qiu, X.; El Zarif, T.; et al. Liquid Biopsy Epigenomic Profiling for Cancer Subtyping. Nat. Med. 2023, 29, 2737–2741. [Google Scholar] [CrossRef]
- Nassiri, F.; Chakravarthy, A.; Feng, S.; Shen, S.Y.; Nejad, R.; Zuccato, J.A.; Voisin, M.R.; Patil, V.; Horbinski, C.; Aldape, K.; et al. Detection and Discrimination of Intracranial Tumors Using Plasma Cell-Free DNA Methylomes. Nat. Med. 2020, 26, 1044–1047. [Google Scholar] [CrossRef]
- Burgener, J.M.; Zou, J.; Zhao, Z.; Zheng, Y.; Shen, S.Y.; Huang, S.H.; Keshavarzi, S.; Xu, W.; Liu, F.-F.; Liu, G.; et al. Tumor-Naïve Multimodal Profiling of Circulating Tumor DNA in Head and Neck Squamous Cell Carcinoma. Clin. Cancer Res. 2021, 27, 4230–4244. [Google Scholar] [CrossRef] [PubMed]
- Stackpole, M.L.; Zeng, W.; Li, S.; Liu, C.-C.; Zhou, Y.; He, S.; Yeh, A.; Wang, Z.; Sun, F.; Li, Q.; et al. Cost-Effective Methylome Sequencing of Cell-Free DNA for Accurately Detecting and Locating Cancer. Nat. Commun. 2022, 13, 5566. [Google Scholar] [CrossRef] [PubMed]
- Galardi, F.; De Luca, F.; Romagnoli, D.; Biagioni, C.; Moretti, E.; Biganzoli, L.; Di Leo, A.; Migliaccio, I.; Malorni, L.; Benelli, M. Cell-Free DNA-Methylation-Based Methods and Applications in Oncology. Biomolecules 2020, 10, 1677. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Debes, J.D.; Boonstra, A. DNA Methylation Markers in the Detection of Hepatocellular Carcinoma. Eur. J. Cancer 2023, 191, 112960. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wei, W.; Ye, Z.; Zheng, J.; Xu, R. Liquid Biopsy of Methylation Biomarkers in Cell-Free DNA. Trends Mol. Med. 2021, 27, 482–500. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.Y.; Burgener, J.M.; Bratman, S.V.; De Carvalho, D.D. Preparation of CfMeDIP-Seq Libraries for Methylome Profiling of Plasma Cell-Free DNA. Nat. Protoc. 2019, 14, 2749–2780. [Google Scholar] [CrossRef] [PubMed]
- deVos, T.; Tetzner, R.; Model, F.; Weiss, G.; Schuster, M.; Distler, J.; Steiger, K.V.; Grutzmann, R.; Pilarsky, C.; Habermann, J.K.; et al. Circulating Methylated SEPT9 DNA in Plasma Is a Biomarker for Colorectal Cancer. Clin. Chem. 2009, 55, 1337–1346. [Google Scholar] [CrossRef]
- Müller, D.; Győrffy, B. DNA Methylation-Based Diagnostic, Prognostic, and Predictive Biomarkers in Colorectal Cancer. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2022, 1877, 188722. [Google Scholar] [CrossRef]
- Weiss, G.; Schlegel, A.; Kottwitz, D.; König, T.; Tetzner, R. Validation of the SHOX2/PTGER4 DNA Methylation Marker Panel for Plasma-Based Discrimination between Patients with Malignant and Nonmalignant Lung Disease. J. Thorac. Oncol. 2017, 12, 77–84. [Google Scholar] [CrossRef]
- Oh, T.; Kim, N.; Moon, Y.; Kim, M.S.; Hoehn, B.D.; Park, C.H.; Kim, T.S.; Kim, N.K.; Chung, H.C.; An, S. Genome-Wide Identification and Validation of a Novel Methylation Biomarker, SDC2, for Blood-Based Detection of Colorectal Cancer. J. Mol. Diagn. 2013, 15, 498–507. [Google Scholar] [CrossRef]
- Klein, E.A.; Madhavan, S.; Beer, T.M.; Bettegowda, C.; Liu, M.C.; Hartman, A.-R.; Hackshaw, A. Dying to Find Out: The Cost of Time at the Dawn of the Multicancer Early Detection Era. Cancer Epidemiol. Biomark. Prev. 2023, 32, 1003–1010. [Google Scholar] [CrossRef]
- Liu, M.C.; Oxnard, G.R.; Klein, E.A.; Swanton, C.; Seiden, M.V.; Liu, M.C.; Oxnard, G.R.; Klein, E.A.; Smith, D.; Richards, D.; et al. Sensitive and Specific Multi-Cancer Detection and Localization Using Methylation Signatures in Cell-Free DNA. Ann. Oncol. 2020, 31, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, B.D.; Oke, J.; Virdee, P.S.; Harris, D.A.; O’Doherty, C.; Park, J.E.S.; Hamady, Z.; Sehgal, V.; Millar, A.; Medley, L.; et al. Multi-Cancer Early Detection Test in Symptomatic Patients Referred for Cancer Investigation in England and Wales (SYMPLIFY): A Large-Scale, Observational Cohort Study. Lancet Oncol. 2023, 24, 733–743. [Google Scholar] [CrossRef]
- Schrag, D.; Beer, T.M.; McDonnell, C.H.; Nadauld, L.; Dilaveri, C.A.; Reid, R.; Marinac, C.R.; Chung, K.C.; Lopatin, M.; Fung, E.T.; et al. Blood-Based Tests for Multicancer Early Detection (PATHFINDER): A Prospective Cohort Study. Lancet 2023, 402, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Nadauld, L.D.; McDonnell, C.H., III; Beer, T.M.; Liu, M.C.; Klein, E.A.; Hudnut, A.; Whittington, R.A.; Taylor, B.; Oxnard, G.R.; Lipson, J. The PATHFINDER Study: Assessment of the Implementation of an Investigational Multi-Cancer Early Detection Test into Clinical Practice. Cancers 2021, 13, 3501. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.; Robbins, H.A. PATHFINDER: Another Step on the Uncharted Path to Multicancer Screening. Lancet 2023, 402, 1213–1215. [Google Scholar] [CrossRef] [PubMed]
- Neal, R.D.; Johnson, P.; Clarke, C.A.; Hamilton, S.A.; Zhang, N.; Kumar, H.; Swanton, C.; Sasieni, P. Cell-Free DNA–Based Multi-Cancer Early Detection Test in an Asymptomatic Screening Population (NHS-Galleri): Design of a Pragmatic, Prospective Randomised Controlled Trial. Cancers 2022, 14, 4818. [Google Scholar] [CrossRef]
- Park, B.H.; Shen, S.Y.; Min, J.; Fleshner, N.; Knox, J.; May, T.; Ailles, L.; Newton, Y.; Zhang, J.; Singhania, R. Development of a Genome-Wide Methylome Enrichment Platform for Multi-Cancer Early Detection (MCED). Cancer Res. 2023, 83 (Suppl. S7), 1030. [Google Scholar] [CrossRef]
- Chemi, F.; Pearce, S.P.; Clipson, A.; Hill, S.M.; Conway, A.-M.; Richardson, S.A.; Kamieniecka, K.; Caeser, R.; White, D.J.; Mohan, S.; et al. CfDNA Methylome Profiling for Detection and Subtyping of Small Cell Lung Cancers. Nat. Cancer 2022, 3, 1260–1270. [Google Scholar] [CrossRef]
- Jeong, I.B.; Yoon, Y.S.; Park, S.Y.; Cha, E.J.; Na, M.J.; Kwon, S.J.; Kim, J.H.; Oh, T.J.; An, S.; Park, C.R. PCDHGA12 Methylation Biomarker in Bronchial Washing Specimens as an Adjunctive Diagnostic Tool to Bronchoscopy in Lung Cancer. Oncol. Lett. 2018, 16, 1039–1045. [Google Scholar] [CrossRef]
- Klein, E.A.; Richards, D.; Cohn, A.; Tummala, M.; Lapham, R.; Cosgrove, D.; Chung, G.; Clement, J.; Gao, J.; Hunkapiller, N.; et al. Clinical Validation of a Targeted Methylation-Based Multi-Cancer Early Detection Test Using an Independent Validation Set. Ann. Oncol. 2021, 32, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Parikh, A.R.; Van Seventer, E.E.; Siravegna, G.; Hartwig, A.V.; Jaimovich, A.; He, Y.; Kanter, K.; Fish, M.G.; Fosbenner, K.D.; Miao, B.; et al. Minimal Residual Disease Detection Using a Plasma-Only Circulating Tumor DNA Assay in Patients with Colorectal Cancer. Clin. Cancer Res. 2021, 27, 5586–5594. [Google Scholar] [CrossRef] [PubMed]
- Janni, W.; Huober, J.; Braun, T.; Müller, V.; Fink, A.; de Gregorio, A.; Rack, B.; Friedl, T.W.; Wiesmüller, L.; Pantel, K.; et al. Abstract 3403: Multiomic, Plasma-Only Circulating Tumor DNA (CtDNA) Assay Identifies Breast Cancer Patients with Minimal Residual Disease (MRD) and Predicts Distant Recurrence. Cancer Res. 2022, 82 (Suppl. S12), 3403. [Google Scholar] [CrossRef]
- Rini, B.I.; Zhang, J.; Hall, O.; Bergener, J.; Wang, Y.; Brown, B.; Min, J.; Shen, S.Y.; Fleshner, N.; Polio, A.; et al. 1910P Evaluation of a Genome-Wide Methylome Enrichment Platform for Circulating Tumor DNA Quantification and Prognostic Performance in Renal Cell Carcinoma (RCC). Ann. Oncol. 2023, 34, S1028. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, J.; Hall, O.; Bergener, J.; Wang, Y.; Brown, B.; Min, J.; Shen, S.Y.; Pienkowski, M.; Huang, S.H.; et al. 866P Prognostic Performance of a Genome-Wide Methylome Enrichment Platform in Head and Neck Cancer. Ann. Oncol. 2023, 34, S561. [Google Scholar] [CrossRef]
- Huang, M.; He, J.; Lai, W.; Liu, L.; Xu, H.; Zeng, Y.; Lan, Q.; Lin, X.; Chu, Z. Methylated Septin 9 Gene Is an Important Prognostic Marker in Stage II and Stage III Colorectal Cancer for Evaluating Local Recurrence or Distant Metastasis after Surgery. BMC Gastroenterol. 2022, 22, 87. [Google Scholar] [CrossRef]
- Hier, J.; Vachon, O.; Bernstein, A.; Ibrahim, I.; Mlynarek, A.; Hier, M.; Alaoui-Jamali, M.A.; Maschietto, M.; da Silva, S.D. Portrait of DNA Methylated Genes Predictive of Poor Prognosis in Head and Neck Cancer and the Implication for Targeted Therapy. Sci. Rep. 2021, 11, 10012. [Google Scholar] [CrossRef]
- Ko, K.; Kananazawa, Y.; Yamada, T.; Kakinuma, D.; Matsuno, K.; Ando, F.; Kuriyama, S.; Matsuda, A.; Yoshida, H. Methylation Status and Long-Fragment Cell-Free DNA Are Prognostic Biomarkers for Gastric Cancer. Cancer Med. 2021, 10, 2003–2012. [Google Scholar] [CrossRef]
- Bae, J.M.; Shin, S.-H.; Kwon, H.-J.; Park, S.-Y.; Kook, M.C.; Kim, Y.-W.; Cho, N.-Y.; Kim, N.; Kim, T.-Y.; Kim, D.; et al. ALU and LINE-1 Hypomethylations in Multistep Gastric Carcinogenesis and Their Prognostic Implications. Int. J. Cancer 2012, 131, 1323–1331. [Google Scholar] [CrossRef]
- Chen, J.; Huan, W.; Zuo, H.; Zhao, L.; Huang, C.; Liu, X.; Hou, S.; Qi, J.; Shi, W. Alu Methylation Serves as a Biomarker for Non-Invasive Diagnosis of Glioma. Oncotarget 2016, 7, 26099. [Google Scholar] [CrossRef]
- Ul Haq, S.; Schmid, S.; Aparnathi, M.K.; Hueniken, K.; Zhan, L.J.; Sacdalan, D.; Li, J.J.N.; Meti, N.; Patel, D.; Cheng, D.; et al. Cell-Free DNA Methylation-Defined Prognostic Subgroups in Small-Cell Lung Cancer Identified by Leukocyte Methylation Subtraction. iScience 2022, 25, 105487. [Google Scholar] [CrossRef] [PubMed]
- Majchrzak-Celińska, A.; Paluszczak, J.; Kleszcz, R.; Magiera, M.; Barciszewska, A.-M.; Nowak, S.; Baer-Dubowska, W. Detection of MGMT, RASSF1A, P15INK4B, and P14ARF Promoter Methylation in Circulating Tumor-Derived DNA of Central Nervous System Cancer Patients. J. Appl. Genet. 2013, 54, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Lok, B.H.; Gardner, E.E.; Schneeberger, V.E.; Ni, A.; Desmeules, P.; Rekhtman, N.; de Stanchina, E.; Teicher, B.A.; Riaz, N.; Powell, S.N.; et al. PARP Inhibitor Activity Correlates with SLFN11 Expression and Demonstrates Synergy with Temozolomide in Small Cell Lung Cancer. Clin. Cancer Res. 2017, 23, 523–535. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Zhang, M.; Zheng, R.; Zheng, S.; Linghu, E.; Herman, J.G.; Guo, M. Methylation of SLFN11 Is a Marker of Poor Prognosis and Cisplatin Resistance in Colorectal Cancer. Epigenomics 2017, 9, 849–862. [Google Scholar] [CrossRef]
- Stewart, C.A.; Tong, P.; Cardnell, R.J.; Sen, T.; Li, L.; Gay, C.M.; Masrorpour, F.; Fan, Y.; Bara, R.O.; Feng, Y. Dynamic Variations in Epithelial-to-Mesenchymal Transition (EMT), ATM, and SLFN11 Govern Response to PARP Inhibitors and Cisplatin in Small Cell Lung Cancer. Oncotarget 2017, 8, 28575. [Google Scholar] [CrossRef] [PubMed]
- Murai, J.; Feng, Y.; Guoying, K.Y.; Ru, Y.; Tang, S.-W.; Shen, Y.; Pommier, Y. Resistance to PARP Inhibitors by SLFN11 Inactivation Can Be Overcome by ATR Inhibition. Oncotarget 2016, 7, 76534. [Google Scholar] [CrossRef] [PubMed]
- Tserpeli, V.; Stergiopoulou, D.; Londra, D.; Giannopoulou, L.; Buderath, P.; Balgkouranidou, I.; Xenidis, N.; Grech, C.; Obermayr, E.; Zeillinger, R. Prognostic Significance of SLFN11 Methylation in Plasma Cell-Free DNA in Advanced High-Grade Serous Ovarian Cancer. Cancers 2021, 14, 4. [Google Scholar] [CrossRef]
- André, T.; Cohen, R.; Salem, M.E. Immune Checkpoint Blockade Therapy in Patients with Colorectal Cancer Harboring Microsatellite Instability/Mismatch Repair Deficiency in 2022. In American Society of Clinical Oncology Educational Book; American Society of Clinical Oncology: Alexandria, VA, USA, 2022; Volume 42, pp. 233–241. [Google Scholar] [CrossRef]
- Pasanen, A.; Loukovaara, M.; Bützow, R. Clinicopathological Significance of Deficient DNA Mismatch Repair and MLH1 Promoter Methylation in Endometrioid Endometrial Carcinoma. Mod. Pathol. 2020, 33, 1443–1452. [Google Scholar] [CrossRef]
- Wang, D.; O’Rourke, D.; Sanchez-Garcia, J.F.; Cai, T.; Scheuenpflug, J.; Feng, Z. Development of a Liquid Biopsy Based Purely Quantitative Digital Droplet PCR Assay for Detection of MLH1 Promoter Methylation in Colorectal Cancer Patients. BMC Cancer 2021, 21, 797. [Google Scholar] [CrossRef]
- Lu, Y.-T.; Plets, M.; Morrison, G.; Cunha, A.T.; Cen, S.Y.; Rhie, S.K.; Siegmund, K.D.; Daneshmand, S.; Quinn, D.I.; Meeks, J.J.; et al. Cell-Free DNA Methylation as a Predictive Biomarker of Response to Neoadjuvant Chemotherapy for Patients with Muscle-Invasive Bladder Cancer in SWOG S1314. Eur. Urol. Oncol. 2023, 6, 516–524. [Google Scholar] [CrossRef]
- Cindy Yang, S.Y.; Lien, S.C.; Wang, B.X.; Clouthier, D.L.; Hanna, Y.; Cirlan, I.; Zhu, K.; Bruce, J.P.; El Ghamrasni, S.; Iafolla, M.A.J.; et al. Pan-Cancer Analysis of Longitudinal Metastatic Tumors Reveals Genomic Alterations and Immune Landscape Dynamics Associated with Pembrolizumab Sensitivity. Nat. Commun. 2021, 12, 5137. [Google Scholar] [CrossRef] [PubMed]
- Sanz Garcia, E.; Zhao, E.Y.; Liu, Z.; Marsh, K.; Abdul Razak, A.R.; Spreafico, A.; Bedard, P.L.; Hansen, A.R.; Lheureux, S.; Torti, D.; et al. Methylated Circulating Tumor DNA (CfMeDIP) as a Predictive Biomarker of Clinical Outcome in Pan-Cancer Patients (Pts) Treated with Pembrolizumab (P). J. Clin. Oncol. 2022, 40 (Suppl. S16), 2550. [Google Scholar] [CrossRef]
- Kim, D.; Kim, J.; Lee, J.; Han, S.K.; Lee, K.; Kong, J.; Kim, Y.J.; Lee, W.Y.; Yun, S.H.; Kim, H.C.; et al. Deconvolution of Bulk Tumors into Distinct Immune Cell States Predicts Colorectal Cancer Recurrence. iScience 2022, 25, 105392. [Google Scholar] [CrossRef] [PubMed]
- Filipski, K.; Scherer, M.; Zeiner, K.N.; Bucher, A.; Kleemann, J.; Jurmeister, P.; Hartung, T.I.; Meissner, M.; Plate, K.H.; Fenton, T.R.; et al. DNA Methylation-Based Prediction of Response to Immune Checkpoint Inhibition in Metastatic Melanoma. J. Immunother. Cancer 2021, 9, e002226. [Google Scholar] [CrossRef]
- Grabovska, Y.; Mackay, A.; O’Hare, P.; Crosier, S.; Finetti, M.; Schwalbe, E.C.; Pickles, J.C.; Fairchild, A.R.; Avery, A.; Cockle, J.; et al. Pediatric Pan-Central Nervous System Tumor Analysis of Immune-Cell Infiltration Identifies Correlates of Antitumor Immunity. Nat. Commun. 2020, 11, 4324. [Google Scholar] [CrossRef]
- Chakravarthy, A.; Furness, A.; Joshi, K.; Ghorani, E.; Ford, K.; Ward, M.J.; King, E.V.; Lechner, M.; Marafioti, T.; Quezada, S.A.; et al. Pan-Cancer Deconvolution of Tumour Composition Using DNA Methylation. Nat. Commun. 2018, 9, 3220. [Google Scholar] [CrossRef] [PubMed]
- Safaei, S.; Mohme, M.; Niesen, J.; Schüller, U.; Bockmayr, M. DIMEimmune: Robust Estimation of Infiltrating Lymphocytes in CNS Tumors from DNA Methylation Profiles. Oncoimmunology 2021, 10, 1932365. [Google Scholar] [CrossRef]
- Liu, T.; Guo, L.; Liu, G.; Hu, X.; Li, X.; Zhang, J.; Dai, Z.; Yu, P.; Jiang, M.; Wang, J.; et al. Molecular Characterization of the Clinical and Tumor Immune Microenvironment Signature of 5-Methylcytosine-Related Regulators in Non-Small Cell Lung Cancer. Front. Cell Dev. Biol. 2021, 9, 779367. [Google Scholar] [CrossRef]
- He, R.; Feng, X.; Yang, K.; Zhou, X.; Li, W.; Zeng, J. Construction of a 5-Methylcytosine-Related Molecular Signature to Inform the Prognosis and Immunotherapy of Lung Squamous Cell Carcinoma. Expert Rev. Mol. Diagn. 2022, 22, 905–913. [Google Scholar] [CrossRef]
- Luskin, M.R.; Murakami, M.A.; Manalis, S.R.; Weinstock, D.M. Targeting Minimal Residual Disease: A Path to Cure? Nat. Rev. Cancer 2018, 18, 255–263. [Google Scholar] [CrossRef]
- Norton, L. Cancer Log-Kill Revisited. In American Society of Clinical Oncology Educational Book; American Society of Clinical Oncology: Alexandria, VA, USA, 2014; Volume 34, pp. 3–7. [Google Scholar] [CrossRef]
- Blackburn, E.H. Cancer Interception. Cancer Prev. Res. 2011, 4, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, J.; Ruan, W.; Huang, M.; Wang, C.; Wang, H.; Jiang, Z.; Wang, S.; Liu, Z.; Liu, C.; et al. Urine DNA Methylation Assay Enables Early Detection and Recurrence Monitoring for Bladder Cancer. J. Clin. Investig. 2020, 130, 6278–6289. [Google Scholar] [CrossRef] [PubMed]
- van Zogchel, L.M.J.; Lak, N.S.M.; Verhagen, O.J.H.M.; Tissoudali, A.; Gussmalla Nuru, M.; Gelineau, N.U.; Zappeij-Kannengieter, L.; Javadi, A.; Zijtregtop, E.A.M.; Merks, J.H.M.; et al. Novel Circulating Hypermethylated RASSF1A DdPCR for Liquid Biopsies in Patients with Pediatric Solid Tumors. JCO Precis. Oncol. 2021, 5, 1738–1748. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Wang, S.; Ni, K.; Zhan, Y.; Ma, H.; Liu, X.; Xin, R.; Zhou, X.; Liu, Z.; Zhao, X. Circulating Methylated SEPT9 DNA Analyses to Predict Recurrence Risk and Adjuvant Chemotherapy Benefit in Stage II to III Colorectal Cancer. Med. Sci. Monit. 2022, 28, e937757-1. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.; Ye, L.; Wang, D.; Han, L.; Zhou, S.; Wang, H.; Dai, W.; Wang, Y.; Luo, W.; Wang, R.; et al. Early Detection of Molecular Residual Disease and Risk Stratification for Stage I to III Colorectal Cancer via Circulating Tumor DNA Methylation. JAMA Oncol. 2023, 9, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Leon Arellano, M.; García-Arranz, M.; Guadalajara, H.; Olivera-Salazar, R.; Valdes-Sanchez, T.; García-Olmo, D. Analysis of Septin 9 Gene Hypermethylation as Follow-Up Biomarker of Colorectal Cancer Patients after Curative Surgery. Diagnostics 2022, 12, 993. [Google Scholar] [CrossRef]
- Chen, K.; Kang, G.; Zhang, Z.; Lizaso, A.; Beck, S.; Lyskjær, I.; Chervova, O.; Li, B.; Shen, H.; Wang, C.; et al. Individualized Dynamic Methylation-Based Analysis of Cell-Free DNA in Postoperative Monitoring of Lung Cancer. BMC Med. 2023, 21, 255. [Google Scholar] [CrossRef]
- Shen, S.Y.; Singhania, R.; Fehringer, G.; Chakravarthy, A.; Roehrl, M.H.A.; Chadwick, D.; Zuzarte, P.C.; Borgida, A.; Wang, T.T.; Li, T.; et al. Sensitive Tumour Detection and Classification Using Plasma Cell-Free DNA Methylomes. Nature 2018, 563, 579–583. [Google Scholar] [CrossRef]
- Papanicolau-Sengos, A.; Aldape, K. DNA Methylation Profiling: An Emerging Paradigm for Cancer Diagnosis. Annu. Rev. Pathol. Mech. Dis. 2022, 17, 295–321. [Google Scholar] [CrossRef]
- Cai, Z.; Poulos, R.C.; Liu, J.; Zhong, Q. Machine Learning for Multi-Omics Data Integration in Cancer. iScience 2022, 25, 103798. [Google Scholar] [CrossRef]
- Akhoundova, D.; Rubin, M.A. Clinical Application of Advanced Multi-Omics Tumor Profiling: Shaping Precision Oncology of the Future. Cancer Cell 2022, 40, 920–938. [Google Scholar] [CrossRef] [PubMed]
- Walls, G.M.; McConnell, L.; McAleese, J.; Murray, P.; Lynch, T.B.; Savage, K.; Hanna, G.G.; de Castro, D.G. Early Circulating Tumour DNA Kinetics Measured by Ultra-Deep next-Generation Sequencing during Radical Radiotherapy for Non-Small Cell Lung Cancer: A Feasibility Study. Radiat. Oncol. 2020, 15, 132. [Google Scholar] [CrossRef] [PubMed]
- Tivey, A.; Church, M.; Rothwell, D.; Dive, C.; Cook, N. Circulating Tumour DNA—Looking beyond the Blood. Nat. Rev. Clin. Oncol. 2022, 19, 600–612. [Google Scholar] [CrossRef]
- Dang, D.K.; Park, B.H. Circulating Tumor DNA: Current Challenges for Clinical Utility. J. Clin. Investig. 2022, 132, e154941. [Google Scholar] [CrossRef] [PubMed]
- Nagasaka, M.; Uddin, M.H.; Al-Hallak, M.N.; Rahman, S.; Balasubramanian, S.; Sukari, A.; Azmi, A.S. Liquid Biopsy for Therapy Monitoring in Early-Stage Non-Small Cell Lung Cancer. Mol. Cancer 2021, 20, 82. [Google Scholar] [CrossRef]
- Hayes, D.F. Defining Clinical Utility of Tumor Biomarker Tests: A Clinician’s Viewpoint. J. Clin. Oncol. 2020, 39, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Boscolo Bielo, L.; Trapani, D.; Repetto, M.; Crimini, E.; Valenza, C.; Belli, C.; Criscitiello, C.; Marra, A.; Subbiah, V.; Curigliano, G. Variant Allele Frequency: A Decision-Making Tool in Precision Oncology? Trends Cancer 2023, 9, 1058–1068. [Google Scholar] [CrossRef]
- Newman, A.M.; Bratman, S.V.; To, J.; Wynne, J.F.; Eclov, N.C.W.; Modlin, L.A.; Liu, C.L.; Neal, J.W.; Wakelee, H.A.; Merritt, R.E.; et al. An Ultrasensitive Method for Quantitating Circulating Tumor DNA with Broad Patient Coverage. Nat. Med. 2014, 20, 548–554. [Google Scholar] [CrossRef]
- An, Y.; Zhao, X.; Zhang, Z.; Xia, Z.; Yang, M.; Ma, L.; Zhao, Y.; Xu, G.; Du, S.; Wu, X.; et al. DNA Methylation Analysis Explores the Molecular Basis of Plasma Cell-Free DNA Fragmentation. Nat. Commun. 2023, 14, 287. [Google Scholar] [CrossRef]
- Dudley, J.C.; Schroers-Martin, J.; Lazzareschi, D.V.; Shi, W.Y.; Chen, S.B.; Esfahani, M.S.; Trivedi, D.; Chabon, J.J.; Chaudhuri, A.A.; Stehr, H.; et al. Detection and Surveillance of Bladder Cancer Using Urine Tumor DNA. Cancer Discov. 2019, 9, 500–509. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Application | Cancer | Test/Assay/Target | Study Size (N) | Sensitivity | Specificity | Comments | References |
|---|---|---|---|---|---|---|---|
| Diagnosis | Colon | Epi proColon®/methylation specific PCR/SEPT9 | 245 | 67% (95% CI: 56–77%) | 89% (95% CI: 83–93%) | Obtained FDA approval in 2016 | [77] |
| Diagnosis | Lung | Epi proLung®/methylation specific PCR/SHOX2 and PTGER4 | 172 | AUROC = 0.88; at specificity = 90%, sensitivity = 67%; at sensitivity = 90%, specificity = 73%. | Obtained CE-IVD mark in 2017 | [79] | |
| Diagnosis | Colon | EarlyTect® Colon/methylation-specific PCR/SCD2 | 256 | 87.0% (95% CI: 80.0–92.3%) | 95.2% (95% CI: 89.8–98.2%) | -- | [80] |
| Diagnosis | Lung | EarlyTect® Lung Cancer/methylation-specific PCR/PCDHGA12 | -- | 75.0% (95% CI: 61.8–81.8%) | 78.9% (95% CI: 62.2–89.8%) | In development for use with bronchial washings or blood | [90] |
| Diagnosis (MCED) | Pan-cancer | GRAIL(Galleri™)/NGS and custom classifier/ methylation panel covering 1.1 × 106 CpGs | 4077 | 51.5% (95% CI: 49.6–53.3%) | 99.5% (95% CI: 99.0–99.8%) | Data based on CCGA trial (NCT02889978) | [91] |
| MRD | Pan-cancer | Guardant Reveal™/NGS and custom classifier/500 genes and 4Mb of DMRs | CRC cohort = 84; Breast cancer cohort = 20 | CRC cohort: 55.6% (95% CI: 35.3–74.5) Breast cancer cohort: sensitivity = 85% 1 | CRC cohort: 100% (95% CI: 90.5–100) Breast cancer cohort: specificity = 100% 1 | -- | [92,93] |
| Diagnosis (MCED)/Prognosis/MRD | Pan-cancer | ADELA/cfMeDIP-seq/whole genome | MCED = 4322 Prognosis: HNSCC cohort = 93; RCC cohort = 151 | MCED: Multi-cancer cohort—Cancer cases discriminated from controls with an AUROC of 0.94 (95% CI: 0.93, 0.96). In low cfDNA shedding tumors, AUROC was 0.92 (95% CI: 0.91, 0.94). Prognosis: HNSCC—Likelihood of recurrence or progression (HR 3.51, 95% CI: 1.1–11.19, p = 0.034); RCC—Likelihood of recurrence or progression (HR 13.28, 95% CI: 5.47–32.26, p < 0.001) | MRD assay currently in development | [88,94,95] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sacdalan, D.B.; Ul Haq, S.; Lok, B.H. Plasma Cell-Free Tumor Methylome as a Biomarker in Solid Tumors: Biology and Applications. Curr. Oncol. 2024, 31, 482-500. https://doi.org/10.3390/curroncol31010033
Sacdalan DB, Ul Haq S, Lok BH. Plasma Cell-Free Tumor Methylome as a Biomarker in Solid Tumors: Biology and Applications. Current Oncology. 2024; 31(1):482-500. https://doi.org/10.3390/curroncol31010033
Chicago/Turabian StyleSacdalan, Danielle Benedict, Sami Ul Haq, and Benjamin H. Lok. 2024. "Plasma Cell-Free Tumor Methylome as a Biomarker in Solid Tumors: Biology and Applications" Current Oncology 31, no. 1: 482-500. https://doi.org/10.3390/curroncol31010033
APA StyleSacdalan, D. B., Ul Haq, S., & Lok, B. H. (2024). Plasma Cell-Free Tumor Methylome as a Biomarker in Solid Tumors: Biology and Applications. Current Oncology, 31(1), 482-500. https://doi.org/10.3390/curroncol31010033