Exercise-Induced Circulating Irisin Level Is Correlated with Improved Cardiac Function in Rats

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Exercise Protocols
2.3. Blood and Tissues Preparation
2.4. Biochemical Analysis
2.5. Echocardiography
2.6. Statistics
3. Results
3.1. Basic Characteristics between the Control and Exercise Groups after 12 Weeks of Exercise
3.2. Comparison of Lipid Profiles between the Control and Exercise Group after 12 Weeks of Exercise
3.3. Comparison of the Levels of Irisin and pro-Inflammatory Cytokines between the Control and Exercise Groups
3.4. Comparison of Echocardiography Variables between the Control and Exercise Groups
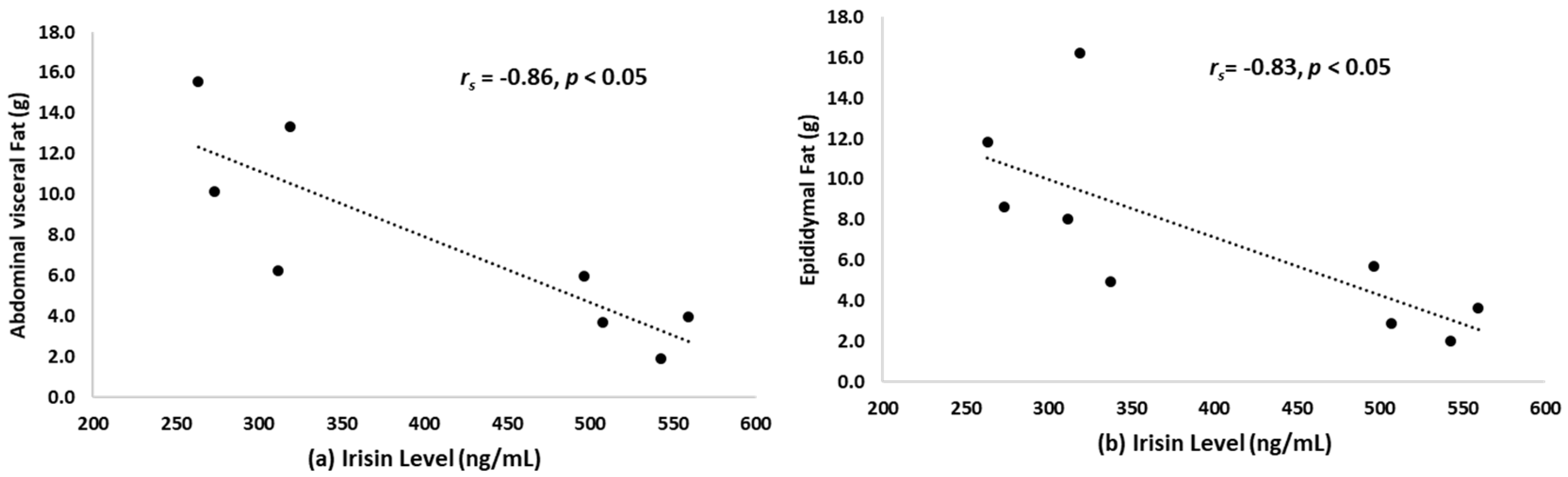
3.5. Correlation between Irisin Level and Abdominal Visceral Fat and Epididymal Fat
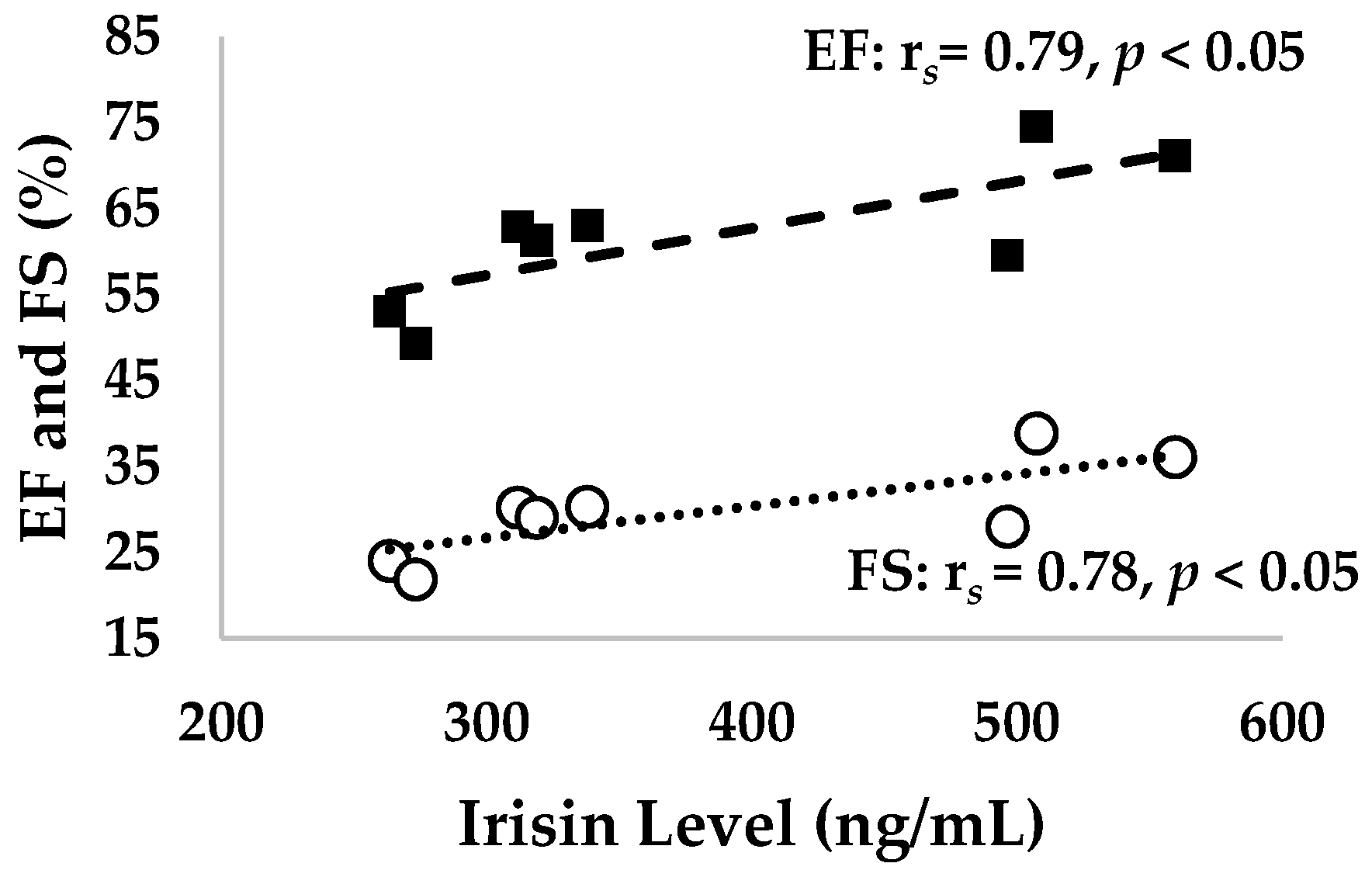
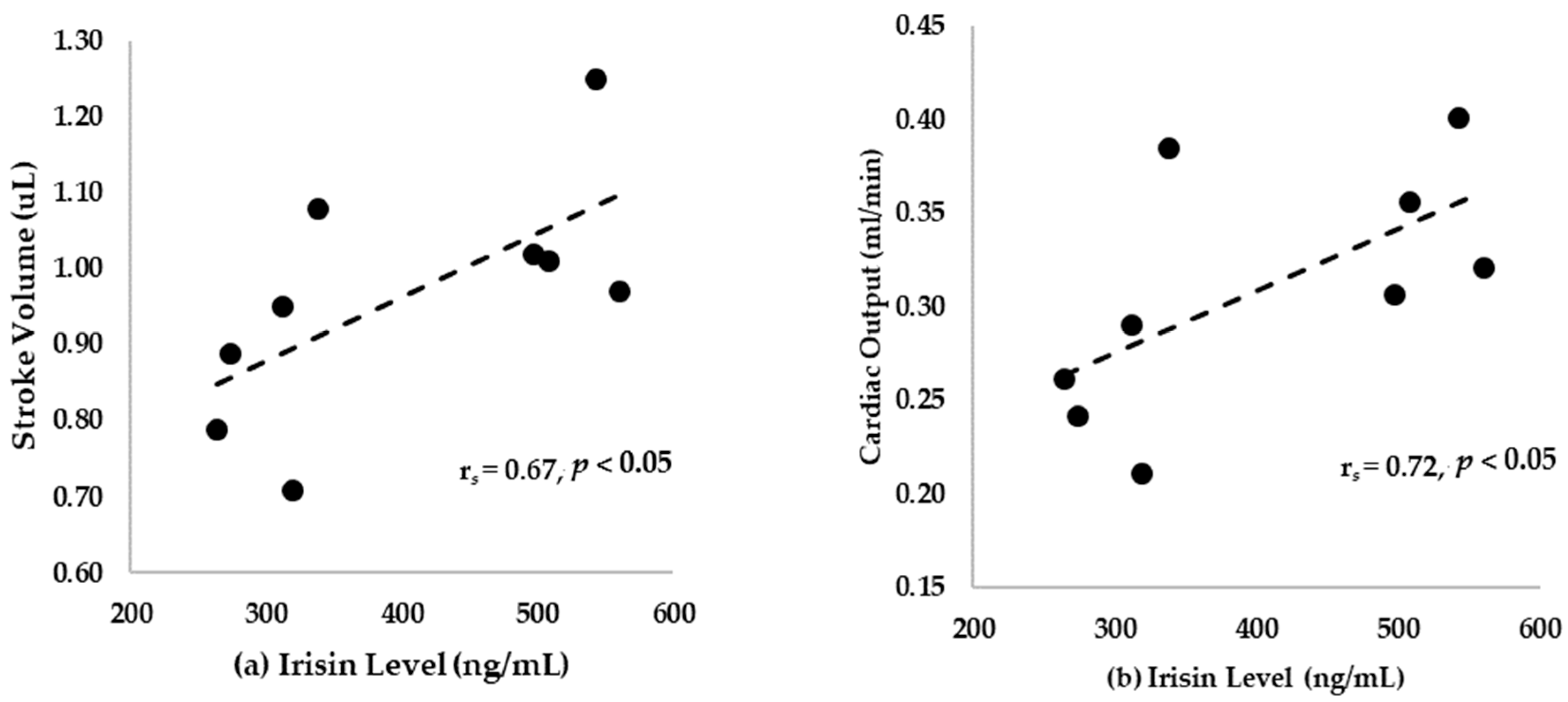
3.6. Correlation between Irisin Level and Ejection Fraction (EF), Fractional Shortening (FS), Stroke Volume, and Cardiac Output
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Booth, F.W.; Roberts, C.K.; Laye, M.J. Lack of exercise is a major cause of chronic diseases. Compr. Physiol. 2012, 2, 1143–1211. [Google Scholar] [CrossRef] [PubMed]
- Im, J.Y.; Bang, H.S.; Seo, D.Y. The effects of 12 weeks of a combined exercise program on physical function and hormonal status in elderly korean women. Int. J. Environ. Res. Public Health 2019, 16, 4196. [Google Scholar] [CrossRef] [PubMed]
- Arhire, L.I.; Mihalache, L.; Covasa, M. Irisin: A hope in understanding and managing obesity and metabolic syndrome. Front. Endocrinol. (Lausanne) 2019, 10, 524. [Google Scholar] [CrossRef] [PubMed]
- Leal, L.G.; Lopes, M.A.; Batista, M.L. Physical exercise-induced myokines and muscle-adipose tissue crosstalk: A review of current knowledge and the implications for health and metabolic diseases. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jun, H.S. Role of myokines in regulating skeletal muscle mass and function. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A pgc1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Q.; Huang, Y.Y.; Gusdon, A.M.; Qu, S. Irisin: A new molecular marker and target in metabolic disorder. Lipids in Health Dis. 2015, 14, 2. [Google Scholar] [CrossRef]
- Vaughan, R.A.; Gannon, N.P.; Barberena, M.A.; Garcia-Smith, R.; Bisoffi, M.; Mermier, C.M.; Conn, C.A.; Trujillo, K.A. Characterization of the metabolic effects of irisin on skeletal muscle in vitro. Diabetes Obes. Metab. 2014, 16, 711–718. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, R.; Meng, Y.; Li, S.; Donelan, W.; Zhao, Y.; Qi, L.; Zhang, M.; Wang, X.; Cui, T.; et al. Irisin stimulates browning of white adipocytes through mitogen-activated protein kinase p38 map kinase and erk map kinase signaling. Diabetes 2014, 63, 514–525. [Google Scholar] [CrossRef]
- Liu, J.J.; Wong, M.D.S.; Toy, W.C.; Tan, C.S.H.; Liu, S.; Ng, X.W.; Tavintharan, S.; Sum, C.F.; Lim, S.C. Lower circulating irisin is associated with type 2 diabetes mellitus. J. Diabetes Complicat. 2013, 27, 365–369. [Google Scholar] [CrossRef]
- Aronis, K.N.; Moreno, M.; Polyzos, S.A.; Moreno-Navarrete, J.M.; Ricart, W.; Delgado, E.; de la Hera, J.; Sahin-Efe, A.; Chamberland, J.P.; Berman, R.; et al. Circulating irisin levels and coronary heart disease: Association with future acute coronary syndrome and major adverse cardiovascular events. Int. J. Obes. (Lond.) 2015, 39, 156–161. [Google Scholar] [CrossRef]
- Wu, F.; Song, H.; Zhang, Y.; Zhang, Y.; Mu, Q.; Jiang, M.; Wang, F.; Zhang, W.; Li, L.; Li, H.; et al. Irisin induces angiogenesis in human umbilical vein endothelial cells in vitro and in zebrafish embryos in vivo via activation of the erk signaling pathway. PLoS ONE 2015, 10, e0134662. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wu, F.; Zhang, Y.; Zhang, Y.; Wang, F.; Jiang, M.; Wang, Z.; Zhang, M.; Li, S.; Yang, L.; et al. Irisin promotes human umbilical vein endothelial cell proliferation through the erk signaling pathway and partly suppresses high glucose-induced apoptosis. PLoS ONE 2014, 9, e110273. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Navarrete, J.M.; Ortega, F.; Serrano, M.; Guerra, E.; Pardo, G.; Tinahones, F.; Ricart, W.; Fernandez-Real, J.M. Irisin is expressed and produced by human muscle and adipose tissue in association with obesity and insulin resistance. J. Clin. Endocrinol. Metab. 2013, 98, E769–E778. [Google Scholar] [CrossRef] [PubMed]
- Roca-Rivada, A.; Castelao, C.; Senin, L.L.; Landrove, M.O.; Baltar, J.; Belen Crujeiras, A.; Seoane, L.M.; Casanueva, F.F.; Pardo, M. Fndc5/irisin is not only a myokine but also an adipokine. PLoS ONE 2013, 8, e60563. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Gao, R.; Bei, Y.; Li, J.; Zhang, H.; Zhou, Y.; Yao, W.; Xu, D.; Zhou, F.; Jin, M.; et al. Serum irisin predicts mortality risk in acute heart failure patients. Cell. Physiol. Biochem. 2017, 42, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Crujeiras, A.B.; Zulet, M.A.; Lopez-Legarrea, P.; de la Iglesia, R.; Pardo, M.; Carreira, M.C.; Martinez, J.A.; Casanueva, F.F. Association between circulating irisin levels and the promotion of insulin resistance during the weight maintenance period after a dietary weight-lowering program in obese patients. Metabolism 2014, 63, 520–531. [Google Scholar] [CrossRef]
- Stengel, A.; Hofmann, T.; Goebel-Stengel, M.; Elbelt, U.; Kobelt, P.; Klapp, B.F. Circulating levels of irisin in patients with anorexia nervosa and different stages of obesity correlation with body mass index. Peptides 2013, 39, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Zaichenko, L.; Brinkoetter, M.; Thakkar, B.; Sahin-Efe, A.; Joung, K.E.; Tsoukas, M.A.; Geladari, E.V.; Huh, J.Y.; Dincer, F.; et al. Circulating irisin in relation to insulin resistance and the metabolic syndrome. J. Clin. Endocrinol. Metab. 2013, 98, 4899–4907. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, Y.T.; Zhang, S.; Dubielecka, P.M.; Du, J.; Yano, N.; Chin, Y.E.; Zhuang, S.; Qin, G.; Zhao, T.C. Irisin plays a pivotal role to protect the heart against ischemia and reperfusion injury. J. Cell. Physiol. 2017, 232, 3775–3785. [Google Scholar] [CrossRef]
- Mazloomzadeh, S.; Karami Zarandi, F.; Shoghli, A.; Dinmohammadi, H. Metabolic syndrome, its components and mortality: A population-based study. Med. J. Islam. Repub. Iran 2019, 33, 11. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Hoang, T.; Bu, S.Y.; Kim, J.M.; Choi, J.H.; Park, E.J.; Lee, S.M.; Park, E.M.; Min, J.Y.; Lee, I.S.; et al. Associations of dietary intake with cardiovascular disease, blood pressure, and lipid profile in the korean population: A systematic review and meta-analysis. J. Lipid Atheroscler. 2020, 9, 205–229. [Google Scholar] [CrossRef]
- Khorasani, Z.M.; Bagheri, R.K.; Yaghoubi, M.A.; Chobkar, S.; Aghaee, M.A.; Abbaszadegan, M.R.; Sahebkar, A. The association between serum irisin levels and cardiovascular disease in diabetic patients. Diabetes Metab. Syndr. 2019, 13, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Lecker, S.H.; Zavin, A.; Cao, P.; Arena, R.; Allsup, K.; Daniels, K.M.; Joseph, J.; Schulze, P.C.; Forman, D.E. Expression of the irisin precursor fndc5 in skeletal muscle correlates with aerobic exercise performance in patients with heart failure. Circ. Heart Fail. 2012, 5, 812–818. [Google Scholar] [CrossRef]
- Reisi, J.; Ghaedi, K.; Rajabi, H.; Marandi, S.M. Can resistance exercise alter irisin levels and expression profiles of fndc5 and ucp1 in rats? Asian J. Sports Med. 2016, 7, e35205. [Google Scholar] [CrossRef]
- Fukushima, Y.; Kurose, S.; Shinno, H.; Thi Thu, H.C.; Takao, N.; Tsutsumi, H.; Hasegawa, T.; Nakajima, T.; Kimura, Y. Effects of body weight reduction on serum irisin and metabolic parameters in obese subjects. Diabetes Metab. J. 2016, 40, 386–395. [Google Scholar] [CrossRef]
- Daskalopoulou, S.S.; Cooke, A.B.; Gomez, Y.H.; Mutter, A.F.; Filippaios, A.; Mesfum, E.T.; Mantzoros, C.S. Plasma irisin levels progressively increase in response to increasing exercise workloads in young, healthy, active subjects. Eur. J. Endocrinol. 2014, 171, 343–352. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, H.J.; So, B.; Son, J.S.; Yoon, D.; Song, W. Effect of aerobic training and resistance training on circulating irisin level and their association with change of body composition in overweight/obese adults: A pilot study. Physiol. Res. 2016, 65, 271–279. [Google Scholar] [CrossRef]
- Aydin, S.; Kuloglu, T.; Aydin, S.; Eren, M.N.; Celik, A.; Yilmaz, M.; Kalayci, M.; Sahin, I.; Gungor, O.; Gurel, A.; et al. Cardiac, skeletal muscle and serum irisin responses to with or without water exercise in young and old male rats: Cardiac muscle produces more irisin than skeletal muscle. Peptides 2014, 52, 68–73. [Google Scholar] [CrossRef]
- Ko, J.R.; Seo, D.Y.; Kim, T.N.; Park, S.H.; Kwak, H.B.; Ko, K.S.; Rhee, B.D.; Han, J. Aerobic exercise training decreases hepatic asprosin in diabetic rats. J. Clin. Med. 2019, 8, 666. [Google Scholar] [CrossRef]
- Seo, D.Y.; Yoon, C.S.; Dizon, L.A.; Lee, S.R.; Youm, J.B.; Yang, W.S.; Kwak, H.B.; Ko, T.H.; Kim, H.K.; Han, J.; et al. Circadian modulation of the cardiac proteome underpins differential adaptation to morning and evening exercise training: An lc-ms/ms analysis. Pflug. Arch. 2020, 472, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, M.; Kawamura, T.; Li, L.; Sasaki, Y.; Matsumoto, K.; Kataoka, H.; Ito, O.; Minami, N.; Sato, T.; Ootaka, T.; et al. Combination of exercise and enalapril enhances renoprotective and peripheral effects in rats with renal ablation. Am. J. Hypertens 2006, 19, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Tavassoli, H.; Heidarianpour, A.; Hedayati, M. The effects of resistance exercise training followed by de-training on irisin and some metabolic parameters in type 2 diabetic rat model. Arch. Physiol. Biochem. 2019, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Li, H.; Shen, S.-W.; Shen, Z.-H.; Xu, M.; Yang, C.-J.; Li, F.; Feng, Y.-B.; Yun, J.-T.; Wang, L.; et al. Swimming exercise increases serum irisin level and reduces body fat mass in high-fat-diet fed wistar rats. Lipids Health Dis. 2016, 15, 93. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.S. The potential of irisin as a therapeutic for diabetes. Future Med. Chem. 2017, 9, 529–532. [Google Scholar] [CrossRef]
- Xin, C.; Liu, J.; Zhang, J.; Zhu, D.; Wang, H.; Xiong, L.; Lee, Y.; Ye, J.; Lian, K.; Xu, C.; et al. Irisin improves fatty acid oxidation and glucose utilization in type 2 diabetes by regulating the ampk signaling pathway. Int. J. Obes. (Lond.) 2016, 40, 443–451. [Google Scholar] [CrossRef]
- Miyamoto-Mikami, E.; Sato, K.; Kurihara, T.; Hasegawa, N.; Fujie, S.; Fujita, S.; Sanada, K.; Hamaoka, T.; Tabata, I.; Iemitsu, M. Endurance training-induced increase in circulating irisin levels is associated with reduction of abdominal visceral fat in middle-aged and older adults. PLoS ONE 2015, 10, e0120354. [Google Scholar] [CrossRef]
- Hwang, Y.C.; Jeon, W.S.; Park, C.Y.; Youn, B.S. The ratio of skeletal muscle mass to visceral fat area is a main determinant linking circulating irisin to metabolic phenotype. Cardiovasc. Diabetol. 2016, 15, 9. [Google Scholar] [CrossRef]
- Kusminski, C.M.; Holland, W.L.; Sun, K.; Park, J.; Spurgin, S.B.; Lin, Y.; Askew, G.R.; Simcox, J.A.; McClain, D.A.; Li, C.; et al. Mitoneet-driven alterations in adipocyte mitochondrial activity reveal a crucial adaptive process that preserves insulin sensitivity in obesity. Nat. Med. 2012, 18, 1539–1549. [Google Scholar] [CrossRef]
- Catalano, K.J.; Stefanovski, D.; Bergman, R.N. Critical role of the mesenteric depot versus other intra-abdominal adipose depots in the development of insulin resistance in young rats. Diabetes 2010, 59, 1416–1423. [Google Scholar] [CrossRef]
- Staiger, H.; Bohm, A.; Scheler, M.; Berti, L.; Machann, J.; Schick, F.; Machicao, F.; Fritsche, A.; Stefan, N.; Weigert, C.; et al. Common genetic variation in the human fndc5 locus, encoding the novel muscle-derived ‘browning’ factor irisin, determines insulin sensitivity. PLoS ONE 2013, 8, e61903. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.Y.; Dincer, F.; Mesfum, E.; Mantzoros, C.S. Irisin stimulates muscle growth-related genes and regulates adipocyte differentiation and metabolism in humans. Int. J. Obes. (Lond.) 2014, 38, 1538–1544. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Zhang, R.; Jiang, F.; Wang, J.; Chen, M.; Peng, D.; Yan, J.; Bao, Y.; Hu, C.; Jia, W. An interaction between a fndc5 variant and obesity modulates glucose metabolism in a chinese han population. PLoS ONE 2014, 9, e109957. [Google Scholar] [CrossRef] [PubMed]
- Tanisawa, K.; Taniguchi, H.; Sun, X.; Ito, T.; Cao, Z.B.; Sakamoto, S.; Higuchi, M. Common single nucleotide polymorphisms in the fndc5 gene are associated with glucose metabolism but do not affect serum irisin levels in japanese men with low fitness levels. Metabolism 2014, 63, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Mazur-Bialy, A.I.; Bilski, J.; Pochec, E.; Brzozowski, T. New insight into the direct anti-inflammatory activity of a myokine irisin against proinflammatory activation of adipocytes. Implication for exercise in obesity. J. Physiol. Pharmacol. 2017, 68, 243–251. [Google Scholar] [PubMed]
- Korta, P.; Pocheć, E.; Mazur-Biały, A. Irisin as a multifunctional protein: Implications for health and certain diseases. Medicina (Kaunas) 2019, 55, 485. [Google Scholar] [CrossRef] [PubMed]
- Swaroop, J.J.; Rajarajeswari, D.; Naidu, J.N. Association of tnf-α with insulin resistance in type 2 diabetes mellitus. Indian J. Med. Res. 2012, 135, 127–130. [Google Scholar] [CrossRef]
- Rehman, K.; Akash, M.S.H. Mechanisms of inflammatory responses and development of insulin resistance: How are they interlinked? J. Biomed. Sci. 2016, 23, 87. [Google Scholar] [CrossRef]
- Bers, D.M. Calcium cycling and signaling in cardiac myocytes. Annu. Rev. Physiol. 2008, 70, 23–49. [Google Scholar] [CrossRef]
- Li, R.L.; Wu, S.S.; Wu, Y.; Wang, X.X.; Chen, H.Y.; Xin, J.J.; Li, H.; Lan, J.; Xue, K.Y.; Li, X.; et al. Irisin alleviates pressure overload-induced cardiac hypertrophy by inducing protective autophagy via mTOR-independent activation of the AMPK-ULK1 pathway. J. Mol. Cell. Cardiol. 2018, 121, 242–255. [Google Scholar] [CrossRef]
- Hassaan, P.S.; Nassar, S.Z.; Issa, Y.; Zahran, N. Irisin vs. Treadmill exercise in post myocardial infarction cardiac rehabilitation in rats. Arch. Med. Res. 2019, 50, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Hulsmans, M.; Clauss, S.; Xiao, L.; Aguirre, A.D.; King, K.R.; Hanley, A.; Hucker, W.J.; Wulfers, E.M.; Seemann, G.; Courties, G.; et al. Macrophages facilitate electrical conduction in the heart. Cell 2017, 169, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Xu, M.; Bryant, J.L.; Ma, J.; Xu, X. Exercise-induced myokine fndc5/irisin functions in cardiovascular protection and intracerebral retrieval of synaptic plasticity. Cell Biosci. 2019, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Silvestrini, A.; Bruno, C.; Vergani, E.; Venuti, A.; Favuzzi, A.M.R.; Guidi, F.; Nicolotti, N.; Meucci, E.; Mordente, A.; Mancini, A. Circulating irisin levels in heart failure with preserved or reduced ejection fraction: A pilot study. PLoS ONE 2019, 14, e0210320. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters Mean (SD) | CON (n = 9) | EXE (n = 9) | z | p-Value |
|---|---|---|---|---|
| Body weight (g) | 542.88 (44.61) | 512.67 (49.76) | 1.28 | 0.20 |
| Heart weight (g) | 1.33 (0.13) | 1.19 (0.08) | 2.06 | 0.04 |
| Abdominal visceral fat weight (g) | 7.99 (4.67) | 3.60 (1.50) | 2.06 | 0.04 |
| Epididymal fat weight (g) | 8.98 (3.86) | 3.27 (1.33) | 3.14 | 0.002 |
| Parameters Mean (SD) | CON (n = 9) | EXE (n = 9) | z | p-Value |
|---|---|---|---|---|
| TC (mg/dL) | 55.11 (6.94) | 48.00 (6.83) | 2.07 | 0.04 |
| HDL-C (mg/dL) | 45.11 (5.69) | 38.88 (6.49) | 1.88 | 0.06 |
| LDL-C (mg/dL) | 15.89 (4.81) | 14.44 (3.43) | 0.80 | 0.42 |
| TG (mg/dL) | 37.22 (5.33) | 34.44 (5.94) | 1.33 | 0.18 |
| Glucose (mg/dL) | 129.89 (23.98) | 110.25 (6.14) | 1.69 | 0.09 |
| Insulin (ng/mL) | 0.38 (0.37) | 0.22 (0.05) | 1.11 | 0.27 |
| Parameters Mean (SD) | CON (n = 9) | EXE (n = 9) | z | p-Value |
|---|---|---|---|---|
| Irisin (ng/mL) | 332.64 (94.52) | 486.78 (101.64) | −2.21 | 0.003 |
| TNF-α (pg/mL) | 22.84 (1.59) | 25.67 (1.03) | −1.29 | 0.20 |
| IL-6 (pg/mL) | 16.35 (0.89) | 17.19 (1.03) | −1.88 | 0.06 |
| hs-CRP (μg/mL) | 0.4 (0.6) | 0.19 (0.03) | 1.37 | 0.17 |
| MMP9 (ng/mL) | 1.82 (0.39) | 1.81 (0.45) | 0.27 | 0.79 |
| Parameters Mean (SD) | CON (n = 9) | EXE (n = 9) | z | p-Value |
|---|---|---|---|---|
| M-Mode | ||||
| IVSd (mm) | 1.59 (0.18) | 1.60 (0.12) | −0.40 | 0.69 |
| IVSs (mm) | 2.29 (0.25) | 2.44 (0.28) | −0.58 | 0.56 |
| LVIDd (mm) | 8.80 (0.56) | 9.15 (0.88) | −0.71 | 0.45 |
| LVIDs (mm) | 6.47 (0.69) | 6.40 (1.05) | 0.49 | 0.63 |
| LVPWd (mm) | 1.89 (0.30) | 1.92 (0.20) | −0.50 | 0.62 |
| LVPWs (mm) | 2.36 (0.22) | 2.22 (0.37) | 1.29 | 0.20 |
| Ejection fraction (%) | 57.20 (6.42) | 64.32 (6.82) | −2.12 | 0.03 |
| Fractional shortening (%) | 26.62 (3.81) | 31.37 (4.70) | −2.12 | 0.03 |
| LV Mass (g) | 1.53 (0.14) | 1.65 (0.13) | −1.84 | 0.07 |
| B-Mode | ||||
| Heart Rate (bpm) | 327.39 (36.31) | 349.90 (43.03) | −1.11 | 0.23 |
| Cardiac output (ml/min) | 0.27 (0.03) | 0.35 (0.04) | −3.09 | 0.002 |
| Stroke volume (uL) | 0.83 (0.11) | 1.01 (0.17) | −2.30 | 0.02 |
| End-Diastolic volume (uL) | 1.32 (0.17) | 1.77 (0.44) | −2.38 | 0.02 |
| End-Systolic volume (uL) | 0.53 (0.10) | 0.84 (0.27) | −2.38 | 0.02 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, D.Y.; Bae, J.H.; Kim, T.N.; Kwak, H.-B.; Kha, P.T.; Han, J. Exercise-Induced Circulating Irisin Level Is Correlated with Improved Cardiac Function in Rats. Int. J. Environ. Res. Public Health 2020, 17, 3863. https://doi.org/10.3390/ijerph17113863
Seo DY, Bae JH, Kim TN, Kwak H-B, Kha PT, Han J. Exercise-Induced Circulating Irisin Level Is Correlated with Improved Cardiac Function in Rats. International Journal of Environmental Research and Public Health. 2020; 17(11):3863. https://doi.org/10.3390/ijerph17113863
Chicago/Turabian StyleSeo, Dae Yun, Jun Hyun Bae, Tae Nyun Kim, Hyo-Bum Kwak, Pham Trong Kha, and Jin Han. 2020. "Exercise-Induced Circulating Irisin Level Is Correlated with Improved Cardiac Function in Rats" International Journal of Environmental Research and Public Health 17, no. 11: 3863. https://doi.org/10.3390/ijerph17113863
APA StyleSeo, D. Y., Bae, J. H., Kim, T. N., Kwak, H.-B., Kha, P. T., & Han, J. (2020). Exercise-Induced Circulating Irisin Level Is Correlated with Improved Cardiac Function in Rats. International Journal of Environmental Research and Public Health, 17(11), 3863. https://doi.org/10.3390/ijerph17113863