Protective Effects of Ammannia baccifera Against CCl4-Induced Oxidative Stress in Rats

 , ,
, , 

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of A. baccifera Extract
2.3. Phytochemical Analysis by UHPLC-QTOF-MS
2.4. Animals
2.5. Experimental Procedure
- Group I: control − olive oil (1 mL/kg)
- Group II: toxic control − 30% CCl4
- Group III: EEAB 250 mg/kg + 30% CCl4
- Group IV: EEAB 500 mg/kg + 30% CCl4
2.6. Quantification of Lipid Peroxidation (LPO) and Analysis of Lipids
2.7. Analysis of Protein Damage: Estimation of Protein Carbonyl and Total Sulfhydryl (Thiol) Contents
2.8. Antioxidants
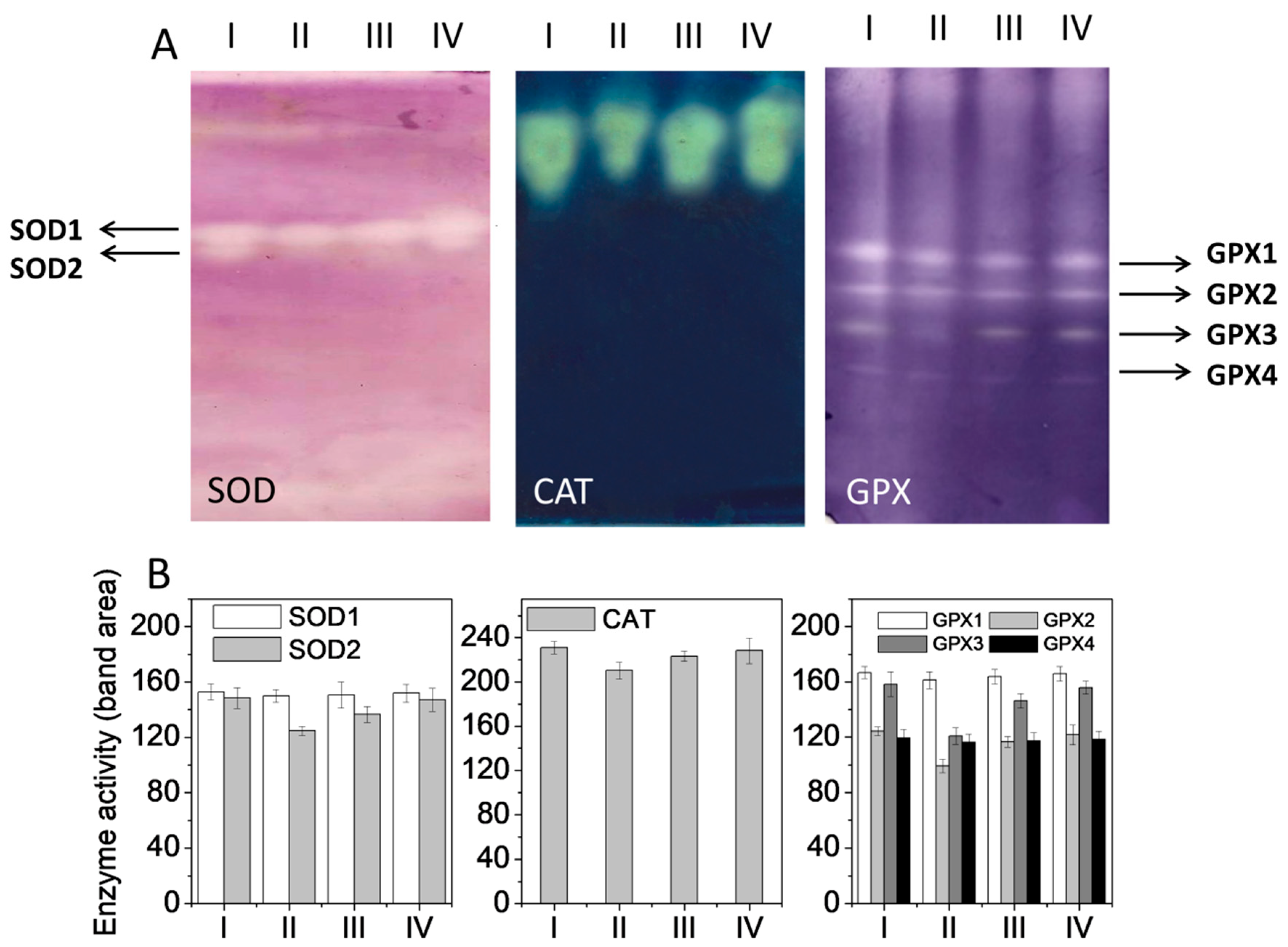
2.9. Separation and Quantification of Isozyme Response by Electrophoresis
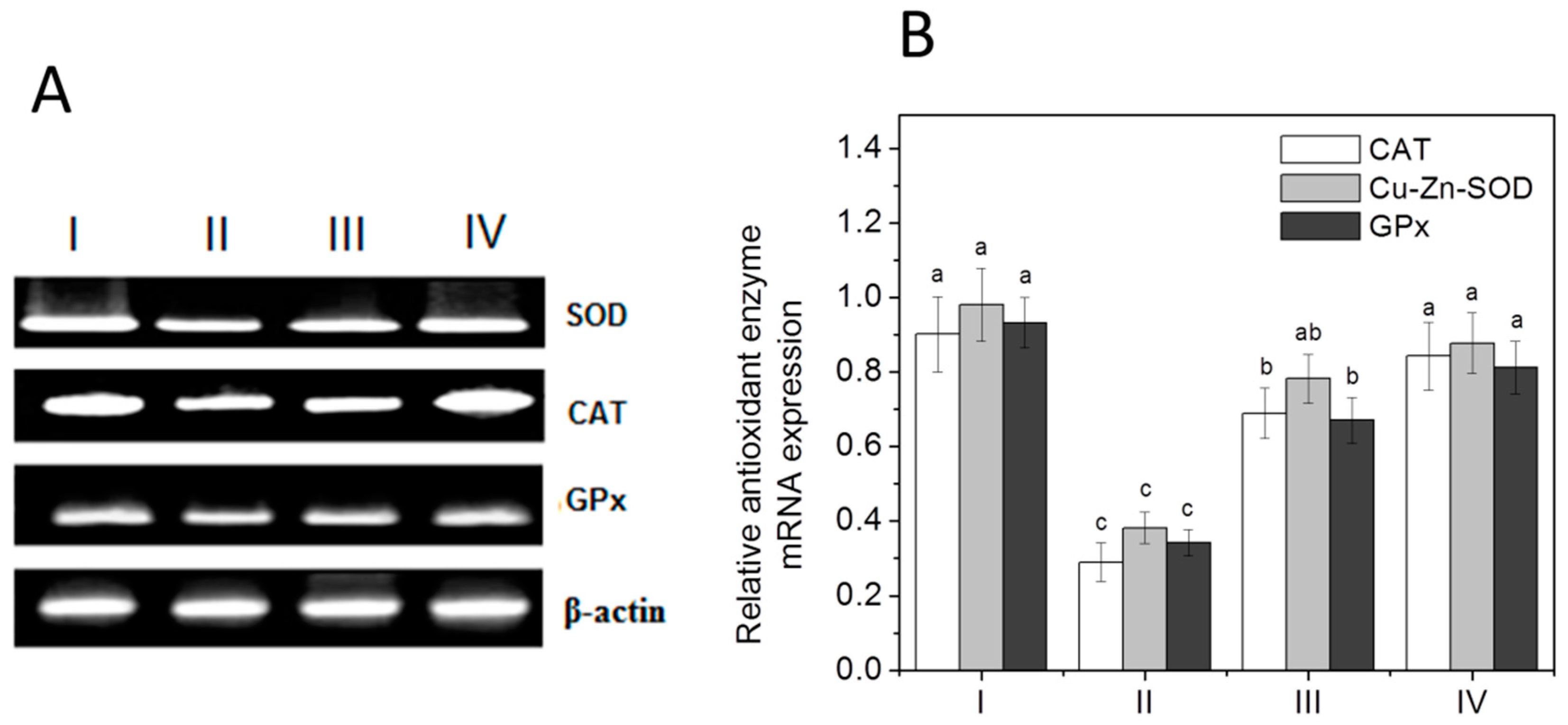
2.10. RNA Extraction and Quantification by a Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
2.11. Statistical Analysis
3. Results
3.1. Identification of Metabolites by UHPLC-QTOF-MS
3.2. Lipid Peroxidation and Lipid Profile
3.3. Total Protein, Protein Carbonyl, and Total Sulfhydryl Content
3.4. Spectrophotometric Analysis of Antioxidants
3.5. Expression of Antioxidant Enzyme Proteins
3.6. Antioxidant Enzymes: mRNA Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brautbar, N.; Williams, J., II. Industrial solvents and liver toxicity: Risk assessment, risk factors and mechanisms. Int. J. Hyg. Environ. Health 2002, 205, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Boll, M.; Weber, L.W.; Becker, E.; Stampfl, A. Pathogenesis of carbon tetrachloride-induced hepatocyte injury bioactivation of CCI4 by cytochrome P450 and effects on lipid homeostasis. Z. Naturforsch. C 2001, 56, 111–121. [Google Scholar] [CrossRef]
- Hensley, K.; Robinson, K.A.; Gabbita, S.P.; Salsman, S.; Floyd, R.A. Reactive oxygen species, cell signaling, and cell injury. Free Radic. Biol. Med. 2000, 28, 1456–1462. [Google Scholar] [CrossRef]
- Kadri, A.; Zarai, Z.; Ben Chobba, I.; Bekir, A.; Gharsallah, N.; Damak, M.; Gdoura, R. Chemical composition and antioxidant activity of Marrubium vulgare L. essential oil from Tunisia. Afr. J. Biotechnol. 2011, 10, 3908–3914. [Google Scholar]
- Lavanya, G.; Voravuthikunchai, S.P.; Towatana, N.H. Acetone Extract from Rhodomyrtus tomentosa: A Potent Natural Antioxidant. Evid. Based Complementary Altern. Med. 2012, 2012, 535479. [Google Scholar] [CrossRef] [PubMed]
- Modak, M.; Dixit, P.; Londhe, J.; Ghaskadbi, S.; Paul, A.; Devasagayam, T. Indian Herbs and Herbal Drugs Used for the Treatment of Diabetes. J. Clin. Biochem. Nutr. 2007, 40, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Loganayaki, N.; Manian, S. Antitumor activity of the methanolic extract of Ammannia baccifera L. against Dalton’s ascites lymphoma induced ascitic and solid tumors in mice. J. Ethnopharmacol. 2012, 142, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Loganayaki, N.; Siddhuraju, P.; Manian, S. Antioxidant, anti-inflammatory and anti-nociceptive effects of Ammannia baccifera L. (Lythracceae), a folklore medicinal plant. J. Ethnopharmacol. 2012, 140, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Das, C.; Sahoo, D.; Sahoo, A.; Nayak, D. Preliminary phytochemical studies and antimicrobial activity of leaf of Ammannia baccifera (Linn.). Pharmacology 2008, 3, 129–132. [Google Scholar]
- Lavanya, G.; Sivajyothi, R.; Parthasarathy, P.R. In vitro Antioxidant Activity of Ethanolic Extract of Ammannia baccifera Linn. Bioscan 2008, 3, 183–188. [Google Scholar]
- Montoro, P.; Maldini, M.; Russo, M.; Postorino, S.; Piacente, S.; Pizza, C. Metabolic profiling of roots of liquorice (Glycyrrhiza glabra) from different geographical areas by ESI/MS/MS and determination of major metabolites by LC-ESI/MS and LC-ESI/MS/MS. J. Pharm. Biomed. Anal. 2011, 54, 535–544. [Google Scholar] [CrossRef]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Innovation: Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, Y.; Ma, L.; Yi, X.; Ruan, J. Metabolomic Analysis Using Ultra-Performance Liquid Chromatography-Quadrupole-Time of Flight Mass Spectrometry (UPLC-Q-TOF MS) Uncovers the Effects of Light Intensity and Temperature under Shading Treatments on the Metabolites in Tea. PLoS ONE 2014, 9, e112572. [Google Scholar] [CrossRef] [PubMed]
- Lavanya, G.; Manjunath, M.; Sivajyothi, R.; Parthasarathy, P.R. Safety evaluation of the ethanol extract of Ammannia baccifera (Lythraceae): Assessment of Acute and Subacute toxicity. J. Pharm. Res. 2010, 3, 2634–2637. [Google Scholar]
- Luo, Y.J.; Yu, J.P.; Shi, Z.H.; Wang, L. Ginkgo biloba extract reverses CCl4-induced liver fibrosis in rats. World J. Gastroenterol. 2004, 10, 1037–1042. [Google Scholar] [CrossRef] [PubMed]
- Manubolu, M.; Goodla, L.; Ravilla, S.; Thanasekaran, J.; Dutta, P.; Malmlof, K.; Obulum, V.R. Protective effect of Actiniopteris radiata (Sw.) Link. against CCl(4) induced oxidative stress in albino rats. J. Ethnopharmacol. 2014, 153, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Parekh, A.C.; Jung, D.H. Cholesterol determination with ferric acetate-uranium acetate and sulfuric acid-ferrous sulfate reagents. Anal. Chem. 1970, 42, 1423–1427. [Google Scholar] [CrossRef]
- Rice, E.W. Triglycerides in serum. In Standards Methods in Clinical Chemistry; Roedrick, P., McDonal, R.P., Eds.; Academic Press: New York, NY, USA, 1970; p. 215. [Google Scholar]
- Van Handel, E. Suggested modifications of the micro determination of triglycerides. Clin. Chem. 1961, 7, 249–251. [Google Scholar]
- Rouser, G.; Fkeischer, S.; Yamamoto, A. Two dimensional then layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 1970, 5, 494–496. [Google Scholar] [CrossRef]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar] [CrossRef]
- Lavanya, G.; Sivajyothi, R.; Manjunath, M.; Parthasarathy, P. Fate of biomolecules during carbon tetrachloride induced oxidative stress and protective nature of Ammannia baccifera Linn.: A natural antioxidant. Int. J. Green Pharm. 2009, 3, 300–305. [Google Scholar]
- Misra, H.P.; Fridovich, I. The Role of Superoxide Anion in the Autoxidation of Epinephrine and a Simple Assay for Superoxide Dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Wendel, A.; Feuerstein, S.; Konz, K.-H. Acute paracetamol intoxication of starved mice leads to lipid peroxidation in vivo. Biochem. Pharmacol. 1979, 28, 2051–2055. [Google Scholar] [CrossRef]
- Moron, M.S.; Depierre, J.W.; Mannervik, B. Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochim. Biophys. Acta (BBA) Gen. Subj. 1979, 582, 67–78. [Google Scholar] [CrossRef]
- Omaye, S.T.; Turnbull, J.D.; Sauberlich, H.E. Selected methods for the determination of ascorbic acid in animal cells, tissues, and fluids. Methods Enzymol. 1979, 62, 3–11. [Google Scholar] [PubMed]
- Naziroglu, M.; Karaoglu, A.; Aksoy, A.O. Selenium and high dose vitamin E administration protects cisplatin-induced oxidative damage to renal, liver and lens tissues in rats. Toxicology 2004, 195, 221–230. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Limaye, P.V.; Raghuram, N.; Sivakami, S. Oxidative stress and gene expression of antioxidant enzymes in the renal cortex of streptozotocin-induced diabetic rats. Mol. Cell. Biochem. 2003, 243, 147–152. [Google Scholar] [CrossRef]
- Weber, L.W.; Boll, M.; Stampfl, A. Hepatotoxicity and mechanism of action of haloalkanes: Carbon tetrachloride as a toxicological model. Crit. Rev. Toxicol. 2003, 33, 105–136. [Google Scholar] [CrossRef]
- Basu, S. Carbon tetrachloride-induced lipid peroxidation: Eicosanoid formation and their regulation by antioxidant nutrients. Toxicology 2003, 189, 113–127. [Google Scholar] [CrossRef]
- Pathakoti, K.; Goodla, L.; Manubolu, M.; Tencomnao, T. Metabolic Alterations and the Protective Effect of Punicalagin Against Glutamate-Induced Oxidative Toxicity in HT22 Cells. Neurotox. Res. 2017, 31, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Suman, T.Y.; Elumalai, D.; Kaleena, P.K.; Rajasree, S.R.R. GC-MS analysis of bioactive components and synthesis of silver nanoparticle using Ammannia baccifera aerial extract and its larvicidal activity against malaria and filariasis vectors. Ind. Crops Prod. 2013, 47, 239–245. [Google Scholar] [CrossRef]
- Michel, T.; Halabalaki, M.; Skaltsounis, A.L. New concepts, experimental approaches, and dereplication strategies for the discovery of novel phytoestrogens from natural sources. Planta Med. 2013, 79, 514–532. [Google Scholar] [CrossRef]
- Koolen, H.H.F.; da Silva, F.M.A.; Gozzo, F.b.C.; de Souza, A.Q.L.; de Souza, A.D.L. Antioxidant, antimicrobial activities and characterization of phenolic compounds from buriti (Mauritia flexuosa L. f.) by UPLC-ESI-MS/MS. Food Res. Int. 2013, 51, 467–473. [Google Scholar] [CrossRef]
- Choi, H.; You, Y.; Hwang, K.; Lee, J.; Chun, J.; Chung, J.W.; Shim, S.; Park, C.-S.; Jun, W. Isolation and identification of compound from dropwort (Oenanthe javanica) with protective potential against oxidative stress in HepG2 cells. Food Sci. Biotechnol. 2011, 20, 1743–1746. [Google Scholar] [CrossRef]
- Choi, J.H.; Kim, D.W.; Yun, N.; Choi, J.S.; Islam, M.N.; Kim, Y.S.; Lee, S.M. Protective effects of hyperoside against carbon tetrachloride-induced liver damage in mice. J. Nat. Prod. 2011, 74, 1055–1060. [Google Scholar] [CrossRef]
- Mikhaeil, B.R.; Badria, F.A.; Maatooq, G.T.; Amer, M.M.A. Antioxidant and Immunomodulatory Constituents of Henna Leaves. Z. Nat. C 2004, 59, 468. [Google Scholar] [CrossRef]
- Kakkar, S.; Bais, S. A review on protocatechuic Acid and its pharmacological potential. ISRN Pharmacol. 2014, 2014, 952943. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-C.; Liao, C.-C.; Peng, C.-C.; Lim, J.-M.; Siao, J.-H.; Wei, C.-M.; Chen, C.-C.; Wu, C.-S.; Chang, T.-M. Dihydromyricetin from Ampelopsis grossedentata inhibits melanogenesis through down-regulation of MAPK, PKA and PKC signaling pathways. Chem. Biol. Interact. 2016, 258, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.K.; Granner, D.K.; Mayes, P.A.; Rodwell, V.W. Harper’s Biochemistry, 23rd ed.; Appleton and Lange/McGraw-Hill: Stamford, CT, USA, 1993. [Google Scholar]
- Lamb, R.G.; Snyder, J.W.; Coleman, J.B. New trends in the prevention of hepatocellular death. Modifiers of calcium movement and of membrane phospholipid metabolism. In Liver Drugs: From Experimental Pharmacology to Therapeutic Application; Testa, B., Perrissaud, D., Eds.; CRC Press: Boca Raton, FL, USA, 1988. [Google Scholar]
- Coleman, J.B.; Condie, L.W.; Lamb, R.G. The influence of CCl4 biotransformation on the activation of rat liver phospholipase C in vitro. Toxicol. Appl. Pharmacol. 1988, 95, 200–207. [Google Scholar] [CrossRef]
- Morrow, J.D.; Awad, J.A.; Boss, H.J.; Blair, I.A.; Roberts, L.J., 2nd. Non-cyclooxygenase-derived prostanoids (F2-isoprostanes) are formed in situ on phospholipids. Proc. Natl. Acad. Sci. USA 1992, 89, 10721–10725. [Google Scholar] [CrossRef] [PubMed]
- Wiggins, D.; Gibbons, G.F. Origin of hepatic very-low-density lipoprotein triacylglycerol: The contribution of cellular phospholipid. Biochem. J. 1996, 320, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Kamalakkannan, N.; Rukkumani, R.; Viswanathan, P.; Rajasekharan, K.; Menon, V.P. Effect of curcumin and its analogue on lipids in carbon tetrachloride-induced hepatotoxicity: A comparative study. Pharm. Biol. 2005, 43, 460–466. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Ritesh, K.R.; Suganya, A.; Dileepkumar, H.V.; Rajashekar, Y.; Shivanandappa, T. A single acute hepatotoxic dose of CCl4 causes oxidative stress in the rat brain. Toxicol. Rep. 2015, 2, 891–895. [Google Scholar] [CrossRef]
- Miesel, R.; Sanocka, D.; Kurpisz, M.; Kroger, H. Antiinflammatory effects of NADPH oxidase inhibitors. Inflammation 1995, 19, 347–362. [Google Scholar] [CrossRef]
- Aquilano, K.; Baldelli, S.; Ciriolo, M.R. Glutathione: New roles in redox signaling for an old antioxidant. Front. Pharmacol. 2014, 5, 196. [Google Scholar] [CrossRef] [PubMed]
- Al-Dbass, A.M.; Al-Daihan, S.K.; Bhat, R.S. Agaricus blazei Murill as an efficient hepatoprotective and antioxidant agent against CCl4-induced liver injury in rats. Saudi J. Biol. Sci. 2012, 19, 303–309. [Google Scholar] [CrossRef]
- Winkler, B.S. Unequivocal evidence in support of the nonenzymatic redox coupling between glutathione/glutathione disulfide and ascorbic acid/dehydroascorbic acid. Biochim. Biophys. Acta 1992, 27, 287–290. [Google Scholar] [CrossRef]
- Zhu, R.; Wang, Y.; Zhang, L.; Guo, Q. Oxidative stress and liver disease. Hepatol. Res. 2012, 42, 741–749. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, MI, USA, 2007. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [PubMed]
- Mates, J.M. Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology. Toxicology 2000, 153, 83–104. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, J.; Waalkes, M.; Kang, Y.J. Changes in the gene expression associated with carbon tetrachloride-induced liver fibrosis persist after cessation of dosing in mice. Toxicol. Sci. 2004, 79, 404–410. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Harries, H.M.; Fletcher, S.T.; Duggan, C.M.; Baker, V.A. The use of genomics technology to investigate gene expression changes in cultured human liver cells. Toxicol. In Vitr. 2001, 15, 399–405. [Google Scholar] [CrossRef]
- El-Sayed, Y.S.; Lebda, M.A.; Hassinin, M.; Neoman, S.A. Chicory (Cichorium intybus L.) root extract regulates the oxidative status and antioxidant gene transcripts in CCl4-induced hepatotoxicity. PLoS ONE 2015, 10, e0121549. [Google Scholar] [CrossRef] [PubMed]
- Szymonik-Lesiuk, S.; Czechowska, G.; Stryjecka-Zimmer, M.; Slomka, M.; Madro, A.; Celinski, K.; Wielosz, M. Catalase, superoxide dismutase, and glutathione peroxidase activities in various rat tissues after carbon tetrachloride intoxication. J. Hepatobiliary Pancreat. Surg. 2003, 10, 309–315. [Google Scholar] [CrossRef]
- Sanchez-Reus, M.I.; Gomez del Rio, M.A.; Iglesias, I.; Elorza, M.; Slowing, K.; Benedi, J. Standardized Hypericum perforatum reduces oxidative stress and increases gene expressionof antioxidant enzymes on rotenone-exposed rats. Neuropharmacology 2007, 52, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B.; Gao, W.; Li, L.; Niu, S.M.; Zhao, L.; Liu, J.; Shi, L.S.; Fu, M.; Liu, F. Rose (Rosa rugosa)-flower extract increases the activities of antioxidant enzymes and their gene expression and reduces lipid peroxidation. Biochem. Cell Biol. 2005, 83, 78–85. [Google Scholar] [CrossRef] [PubMed]
- El-baky, A.; Hanaa, H.; Hussein, M. Influence of salinity on lipid peroxidation, antioxidant enzymes and electrophoretic patterns of protein and isoenzymes in leaves of some onion cultivars. Asian J. Plant Sci. 2003, 2, 633–638. [Google Scholar]



{kind=link}
{kind=link}
| Gene Description | Primer Sequence (5′→3′) | Gene Bank Accession No. | Length (bp) |
|---|---|---|---|
| Cu-Zn-SOD | F: 5′GCAGAAGGCAAGCGGTGAAC | X05634 | 387 |
| R: 5′TAGCAGGACAGCAGATGAGT | |||
| CAT | F: 5′GCGAATGGAGAGGCAGTGTAC | AH004967 | 670 |
| R:5′GAGTGACGTTGTCTTCATTAGCACTG | |||
| GPx | F: 5′CTCTCCGCGGTGGCACAGT | M21210 | 290 |
| R: 5′CCACCACCGGGTCGGACATAC | |||
| B-Actin * | F: 5′CTGCTTGCTGATCCACA | V01217 | 505 |
| R: 5′CTGACCGAGCGTGGCTAC |
| S. No. | RT (min) | Mass | Tentative Identification | Formula | DB Diff (ppm) |
|---|---|---|---|---|---|
| Hydroxycinnamic Acids | |||||
| 1 | 4.37 | 354.0958 | Chlorogenic acid | C16H18O9 | −2.09 |
| Flavonoids | |||||
| 2 | 4.67; 6.65 | 448.0992 | Quercetin | C21H20O11 | 2.96; 4.08 |
| 3 | 4.67; 6.43; 6.65; 9.16 | 304.0578 | Pentahydroxy flavanone | C15H12O7 | 1.73; 2.58; 3.28; 3.29 |
| 4 | 6.05 | 432.1052 | Cosmosiin | C21H20O10 | 1.1 |
| 5 | 6.02; 6.22; | 320.0527 | Dihydromyricetin | C15H12O8 | 1.75; 3.31 |
| 6 | 5.03 | 344.086 | Eupatorin | C18H16O7 | 10.49 |
| 7 | 6.72 | 317.065 | Petunidin | C16H13O7 | 3.71 |
| 8 | 0.82 | 138.0675 | 4-Hydroxyphenyl ethanol | C8H10O2 | 4.38 |
| 9 | 6.78 | 154.026 | Protocatechuic acid | C7H6O4 | 3.65 |
| Terpenoids | |||||
| 10 | 4.56; 4.79 | 420.1618 | Lamioside | C18H28O11 | 3.26; 1.41 |
| 11 | 14.7; 18.06 | 328.1665 | Crocetin | C20H24O4 | 2.83; 2.38 |
| 12 | 4.07; 4.17 | 526.2536 | Khayasin C | C30H38O8 | 5.75; 6.42 |
| 13 | 4.90 | 166.099 | Perillic acid | C10H14O2 | 2.03 |
| 14 | 6.61 | 150.1038 | (-)-Isopiperitenone | C10H14O | 4.16 |
| 15 | 4.66 | 406.1458 | 10-Hydroxyloganin | C17H26O11 | 4.11 |
| 16 | 4.9 | 152.1197 | (-)-trans-Carveol | C10H16O | 2.88 |
| Parameter | Group I | Group II | Group III | Group IV | p-Value |
|---|---|---|---|---|---|
| LPO | 1.83 ± 0.01 a | 3.94 ± 0.01 b | 3.08 ± 0.01 b,c | 2.15 ± 0.02 a | 0.041 |
| Triacylglycerides | 3.21 ± 0.01 a | 6.57 ± 0.05 b | 4.82 ± 0.04 c | 3.79 ± 0.02 a,c | 0.010 |
| Cholesterol | 3.10 ± 0.04 a | 4.22 ± 0.05 b | 4.02 ± 0.02 c | 3.72 ± 0.02 a | 0.010 |
| Phospholipids | 14.11 ± 0.03 a | 10.16 ± 0.05 b | 12.75 ± 0.16 c | 13.85± 0.04 a | 0.045 |
| Parameter | Group I | Group II | Group III | Group IV | p-Value |
|---|---|---|---|---|---|
| Total Protein | 154.23 ± 0.54 a | 128.9 ± 0.32 b | 135.92 ± 0.45 c | 147.95 ± 0.39 a | 0.006 |
| Protein Carbonyl | 3.91 ± 0.01 c | 21.47 ± 0.17 a | 17.94 ± 0.15 d | 7.50 ± 0.22 a | 0.045 |
| Total Sulfhydryl | 3.10 ± 0.021 a | 0.91 ± 0.02 b | 1.53 ± 0.01 c | 2.87 ± 0.02 a | 0.030 |
| Parameter | Group I | Group II | Group III | Group IV | p-Value |
|---|---|---|---|---|---|
| SOD | 0.54 ± 0.01 a | 0.19 ± 0.00 b | 0.34 ± 0.00 c | 0.46 ± 0.00 d | 0.000 |
| CAT | 78.27 ± 0.57 a | 32.73 ± 0.16 b | 48.68 ± 0.16 c | 68.77 ± 0.27 d | 0.030 |
| GPx | 60.77 ± 0.80 a | 27.57 ± 0.64 b | 40.86 ± 0.28 c | 55.89 ± 0.29 c | 0.007 |
| GSH | 5.61 ± 0.15 a | 1.29 ± 0.02 b | 2.92 ± 0.02 c | 4.73 ± 0.15 d | 0.033 |
| Vitamin C | 3.10 ± 0.02 a | 2.14 ± 0.06 b | 2.56 ± 0.03 c | 2.81 ± 0.01 c | 0.014 |
| Vitamin E | 1.81 ± 0.01 a | 0.89 ± 0.02 b | 1.13 ± 0.03 c | 1.55 ± 0.02 d | 0.035 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goodla, L.; Manubolu, M.; Pathakoti, K.; Jayakumar, T.; Sheu, J.-R.; Fraker, M.; Tchounwou, P.B.; Poondamalli, P.R. Protective Effects of Ammannia baccifera Against CCl4-Induced Oxidative Stress in Rats. Int. J. Environ. Res. Public Health 2019, 16, 1440. https://doi.org/10.3390/ijerph16081440
Goodla L, Manubolu M, Pathakoti K, Jayakumar T, Sheu J-R, Fraker M, Tchounwou PB, Poondamalli PR. Protective Effects of Ammannia baccifera Against CCl4-Induced Oxidative Stress in Rats. International Journal of Environmental Research and Public Health. 2019; 16(8):1440. https://doi.org/10.3390/ijerph16081440
Chicago/Turabian StyleGoodla, Lavanya, Manjunath Manubolu, Kavitha Pathakoti, Thanasekaran Jayakumar, Jeon-Rong Sheu, Mike Fraker, Paul B. Tchounwou, and Parthasarathy R. Poondamalli. 2019. "Protective Effects of Ammannia baccifera Against CCl4-Induced Oxidative Stress in Rats" International Journal of Environmental Research and Public Health 16, no. 8: 1440. https://doi.org/10.3390/ijerph16081440
APA StyleGoodla, L., Manubolu, M., Pathakoti, K., Jayakumar, T., Sheu, J.-R., Fraker, M., Tchounwou, P. B., & Poondamalli, P. R. (2019). Protective Effects of Ammannia baccifera Against CCl4-Induced Oxidative Stress in Rats. International Journal of Environmental Research and Public Health, 16(8), 1440. https://doi.org/10.3390/ijerph16081440