Concurrent Drought and Temperature Stress in Rice—A Possible Result of the Predicted Climate Change: Effects on Yield Attributes, Eating Characteristics, and Health Promoting Compounds

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation
Plant Material
2.2. Growth Chamber
2.3. Sowing and Watering
2.4. Drought Treatments
2.5. Data Recording in the Growth Chambers
2.6. Analysis of Nutritional Quality
2.6.1. Sample Preparation
2.6.2. Total Protein Content
2.6.3. Total Phenolic Content and Total Antioxidant Capacity
2.6.4. Amylose Content
2.6.5. Gel Consistency
2.6.6. Gelatinization Temperature
2.7. Statistical Analysis
3. Results
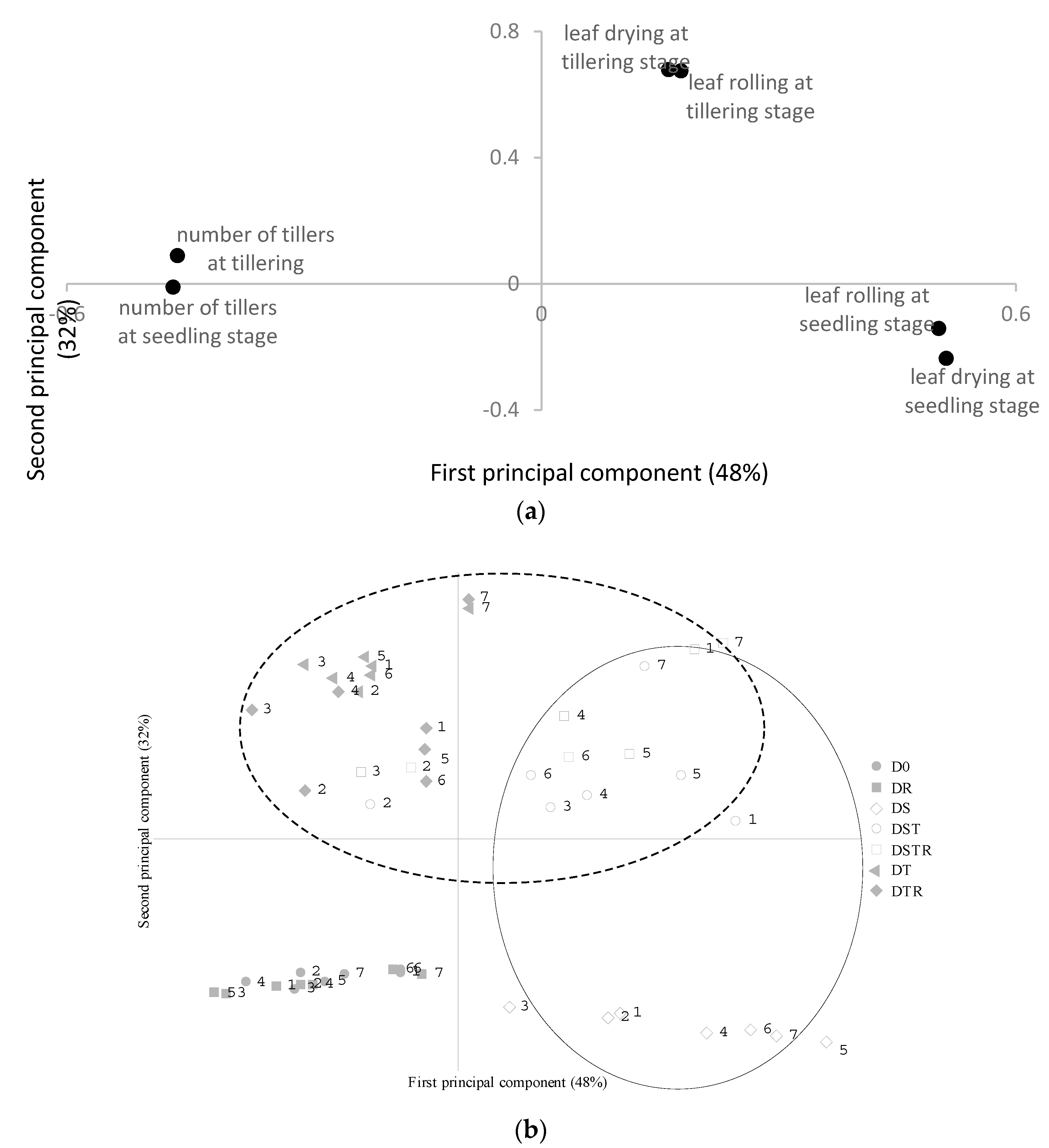
3.1. Early Plant Growth—Drought Sensitivity Score, Number of Tillers Plant−1 and Time to Flowering
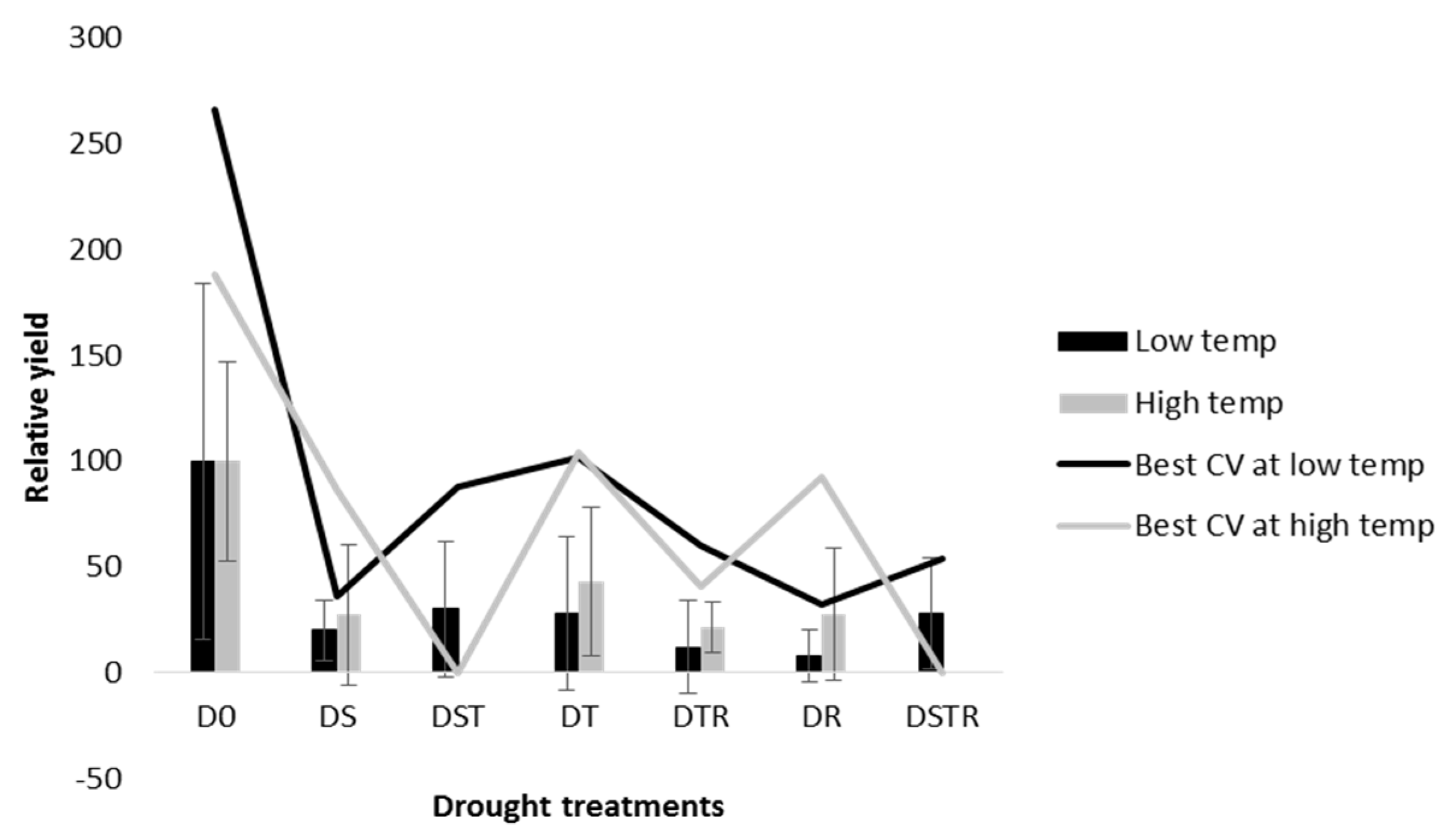
3.2. Rice Production—Grain Yield and Yield Components
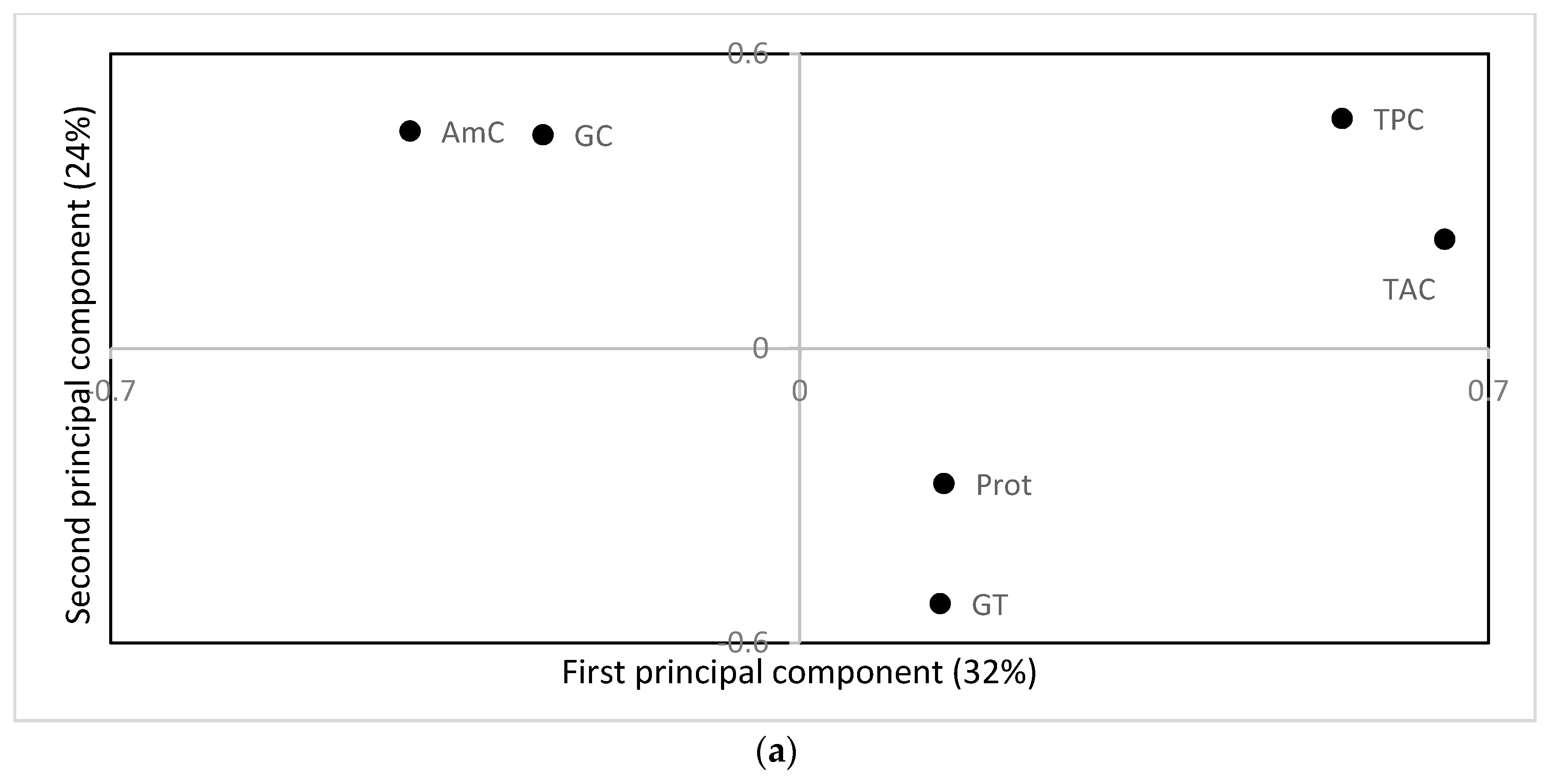
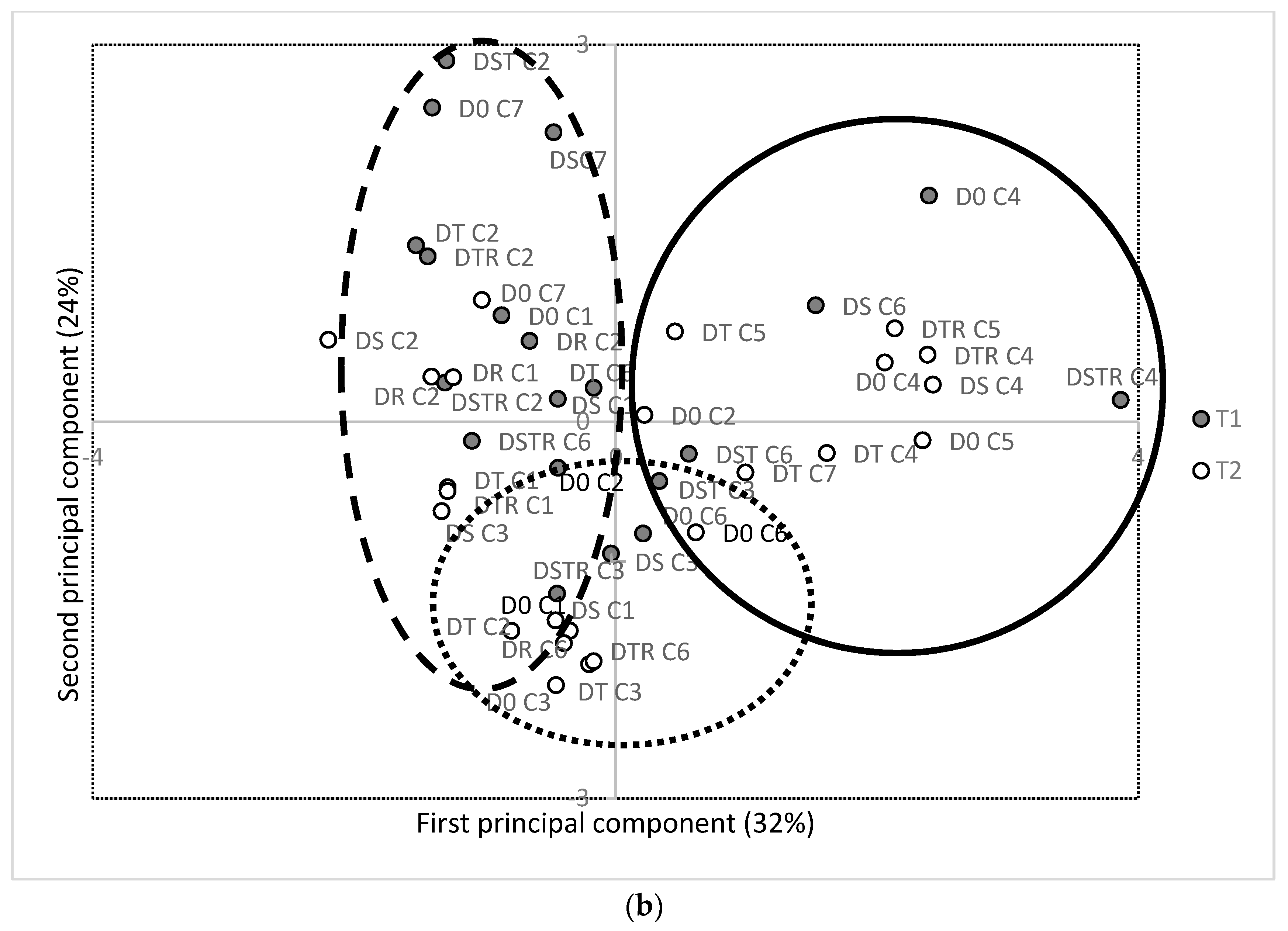
3.3. Eating Quality and Health Promoting Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krishnan, P.; Ramakrishnan, B.; Reddy, K.R.; Reddy, V.R. High-temperature effects on rice growth, yield, and grain quality. In Advances in agronomy; Sparks, D.L., Ed.; Academic Press: Burlington, NJ, USA, 2011; Volume 111, pp. 87–206. [Google Scholar]
- Intergovernmental Panel Climate Change (IPCC). Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change 2014; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; 151p. [Google Scholar]
- Mukamuhirwa, A.; Persson Hovmalm, H.; Ortiz, R.; Nyamangyoku, O.; Prieto-Linde, M.L.; Ekholm, A.; Johansson, E. Effect of intermittent drought on grain yield and quality of rice (Oryza sativa L.) grown in Rwanda. (in press)
- The 2016 Report on Future Climate of Africa. Available online: http://2016report.futureclimateafrica.org/reader/east-africa/Rwanda-factsheet-climate-information-for-an-uncertain-future/ (accessed on 10 January 2019).
- Wassmann, R.; Jagadish, S.V.K.S.; Heuer, S.; Ismail, A.; Redona, E.; Serraj, R.; Singh, R.K.; Howell, G.; Pathak, H.; Sumfleth, K. Climate change affecting rice production: The physiological and agronomic basis for possible adaptation strategies. Adv. Agron. 2009, 101, 59–121. [Google Scholar]
- Wopereis, M.C.S.; Defoer, T.; Idinoba, P.; Diack, S.; Dugué, M.J. Participatory Learning and Action Research (PLAR) for Integrated Rice Management (IRM) in Inland Valleys of Sub-Saharan Africa: Technical Manual; WARDA Training Series; Africa Rice Center: Cotonou, Benin, 2008; 128p. [Google Scholar]
- Oh-e, I.; Saitoh, K.; Kuroda, T. Effects of high temperature on growth, yield and dry-matter production of rice grown in the paddy field. Plant Prod. Sci. 2007, 10, 412–422. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Craufurd, P.Q.; Wheeler, T.R. High temperature stress and spikelet fertility in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Shimono, H.; Hasegawa, T.; Iwama, K. Response of growth and grain yield in paddy rice to cool water at different growth stages. Field Crops Res. 2002, 73, 67–79. [Google Scholar] [CrossRef]
- Emam, M.M.; Khattab, H.E.; Helal, N.M.; Deraz, E.A. Effect of selenium and silicon on yield quality of rice plant grown under drought stress. AJCS 2014, 8, 596–605. [Google Scholar]
- Vidal, V.; Pons, B.; Brunnschweiler, J.; Handschin, S.; Rouau, X.; Mestres, C. Cooking behaviour of rice in relation to kernel physicochemical properties. J. Agric. Food Chem. 2007, 55, 336–346. [Google Scholar] [CrossRef]
- Bouman, B.A.M.; Lampayan, R.M.; Tuong, T.P. Water Management in Irrigated Rice: Coping with Water Scarcity; IRRI: Los Banos, Philippines, 2007. [Google Scholar]
- IRRI. Available online: http://irri.org/our-work/research/better-rice-varieties/climate-change-ready-rice (accessed on 22 August 2018).
- Zandalinas, S.I.; Mittlerb, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant 2017. [Google Scholar] [CrossRef] [PubMed]
- Zwart, S.J.; Bastiaansen, W.G.M. Review of measured crop water productivity values for irrigated wheat, rice, cotton and maize. Agric. Water Manag. 2004, 69, 115–133. [Google Scholar] [CrossRef]
- ISAR. Description of New Rice Varieties Introduced in Rwanda; Rwanda Agricultural Research Institute: Kigali, Rwanda, 2010. [Google Scholar]
- Porter, A.S.; Evans-Fitz Gerald, C.; McElwain, J.C.; Yiotis, C.; Elliott-Kingston, C. How well do you know your growth chambers? Testing for chamber effect using plant traits. Plant Meth. 2015, 11, 44. [Google Scholar] [CrossRef]
- Johansson, E.; Oscarson, P.; Lundborg, T. Effect of planting date on flowering time in wheat. Physiol. Plant 1996, 96, 338–341. [Google Scholar] [CrossRef]
- Kohl, K. Growing rice in controlled environments. Ann. Appl. Biol. 2015, 167, 157–177. [Google Scholar] [CrossRef]
- Cheng, W.; Sakai, H.; Yagi, K.; Hasegawa, T. Interactions of elevated [CO2] and night temperature on rice growth and yield. Agric. For. Meteorol. 2009, 149, 51–58. [Google Scholar] [CrossRef]
- Harrington, S. Greenhouse Procedures Manual, Growing Rice under Controlled Conditions; McCouch Research Program; Cornell University: Ithaca, NY, USA, 2010. [Google Scholar]
- Best, R. Some Aspects of Photoperiodism in Rice (Oryza sativa L.); Elsevier: Amsterdam, The Netherlands, 1961; 87p. [Google Scholar]
- Hirai, G.; Okumura, T.; Takeuchi, S.; Tanaka, O.; Chujo, H.; Tanaka, N. Studies on the effect of the relative humidity of the atmosphere on the growth and physiology of rice plants: Effect of ambient humidity on the translocation of assimilated 13C in leaves. Jpn. J. Crop Sci. 1996, 65, 460–464. [Google Scholar] [CrossRef]
- IRRI. Standards Evaluation System for Rice. 2002. Available online: http://www.knowledgebank.irri.org/images/docs/rice-standard-evaluation-system.pdf (accessed on 20 September 2015).
- McGeehan, S.L.; Naylor, D.V. Automated instrumental analysis of carbon and nitrogen in plant and soil samples. Commun. Soil Sci. Plant Anal. 1988, 19, 493–505. [Google Scholar] [CrossRef]
- International Organization for Standardization, ISO16634-2. Food Products—Determination of the Total Nitrogen Content by Combustion according to the Dumas Principle and Calculation of the Crude Protein Content—Part 2: Cereals, Pulses and Milled Cereal Products. 2016. Available online: https://www.iso.org/obp/ui/#iso:std:iso:16634:-2:ed-1:v1:en (accessed on 18 February 2018).
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar]
- Pérez-Jiménez, J.; Saura-Calixto, F. Literature data may underestimate the actual antioxidant capacity of cereals. J. Agric. Food Chem. 2005, 53, 5036–5040. [Google Scholar] [CrossRef]
- Mukamuhirwa, A.; Hovmalm, H.P.; Ortiz, R.; Nyamangyoku, O.; Johansson, E. Quality and grain yield attributes of Rwandan rice (Oryza sativa L.) cultivars grown in a biotron applying two NPK levels. J. Food Qual. 2018. [Google Scholar] [CrossRef]
- Hofvander, P.; Andersson, M.; Larsson, C.T.; Larsson, H. Field performance and starch characteristics of high amylose potatoes obtained by antisense gene targeting of two branching enzymes. Plant Biotechnol. J. 2004, 2, 311–320. [Google Scholar] [CrossRef]
- Perez, C.M.; Juliano, B.O. Modification of the simplified amylose test for milled rice. Starch 1978, 30, 424–426. [Google Scholar] [CrossRef]
- Cagampang, G.B.; Perez, C.M.; Juliano, B.O. A gel consistency test for eating quality of rice. J. Sci. Food Agric. 1973, 24, 1589–1594. [Google Scholar] [CrossRef]
- Gunartne, A.; Ratnayaka, U.P.; Sirisena, N.; Ratnayaka, J.; Kong, X.; Arachchi, L.V.; Corke, H. Effect of soil moisture stress from flowering to grain maturity on functional properties of Sri Lankan rice flour. Starch 2011, 63, 283–290. [Google Scholar] [CrossRef]
- Malik, A.H.; Kuktaite, R.; Johansson, E. Combined effect of genetic and environmental factors on the accumulation of proteins in the wheat grain and their relationship to bread-making quality. J. Cer. Sci. 2013, 57, 170–174. [Google Scholar] [CrossRef]
- SAS. Statistical Analysis System, User’s Guide, 7th ed.; SAS Inst. Inc.: Cary, NC, USA, 2004. [Google Scholar]
- Mathew, I.; Shimelis, H.; Mwadzingeni, L.; Zengeni, R.; Mutema, M.; Chaplot, V. Variance components and heritability of traits related to root:shoot biomass allocation and drought tolerance in wheat. Euphytica 2018, 214, 225. [Google Scholar] [CrossRef]
- Parent, B.; Suard, B.; Serraj, R.; Tardieu, F. Rice leaf growth and water potential are resilient to evaporative demand and soil water deficit once the effects of root systems are neutralized. Plant Cell Environ. 2010, 33, 1256–1267. [Google Scholar] [CrossRef]
- Katoa, Y.; Kamoshita, A.; Yamagishi, J. Preflowering abortion reduces spikelet number in upland rice (Oryza sativa L.) under water stress. Crop Sci. Soc. Am. 2008, 48, 2389–2395. [Google Scholar] [CrossRef]
- Terra, T.G.R.; Leal, T.C.A.B.; Hideo, P.; Range, N.; Barros, H.B.; Dos Santos, A.C. Tolerance to drought in rice cultivars in Southern Cerrado area from Tocantins state, Brazil. Acta Sci. Agric. 2010, 32, 715–719. [Google Scholar]
- Rang, Z.W.; Jagadish, S.V.K.; Zhou, Q.M.; Craufurd, P.Q.; Heuer, S. Effect of high temperature and water stress on pollen germination and spikelet fertility in rice. Environ. Exp. Bot. 2011, 70, 58–65. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhang, Y.; Xiang, J.; Uphoff, N.T.; Pan, X.; Zhu, D. Effects of low temperature stress on spikelet-related parameters during anthesis in indica-japonica hybrid rice. Front. Plant Sci. 2017, 8, 1350. [Google Scholar] [CrossRef]
- Ahmed, N.; Tetlow, I.J.; Nawaz, S.; Iqbal, A.; Mubin, M.; Rehman, M.S.; Butt, A.; Lightfootc, D.A.; Maekawa, M. Effect of high temperature on grain filling period, yield, amylose content and activity of starch biosynthesis enzymes in endosperm of basmati rice. J. Sci. Food Agric. 2015, 95, 2237–2243. [Google Scholar] [CrossRef]
- Ghosh, T.; Rai, M.; Tyagi, W.; Challam, C. Seedling stage low temperature response in tolerant and susceptible rice genotypes suggests role of relative water content and members of OsSNAC gene family. Plant Sign. Behav. 2016, 11, e1138192. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Salam, M.U.; Jones, J.W.; Jones, J.W.G. Phasic development of rice seedlings. Agron. J. 1997, 89, 653–658. [Google Scholar] [CrossRef]
- Bunnag, S.; Pongthai, P. Selection of rice (Oryza sativa L) cultivars tolerant to drought stress at the vegetative stage under field conditions. Am. J. Plant Sci. 2013, 4, 1701–1708. [Google Scholar] [CrossRef]
- Shao, Y.; Tang, F.; Huang, Y.; Xu, F.; Chen, Y.; Tong, C.; Chen, H.; Bao, J. Analysis of genotype × environment interactions for polyphenols and antioxidant capacity of rice by association mapping. J. Agric. Food Chem. 2014, 62, 5361–5368. [Google Scholar] [CrossRef]
- Pang, Y.; Ahmed, S.; Xu, Y.; Beta, T.; Zhu, Z.; Shao, Y.; Bao, J. Bound phenolic compounds and antioxidant properties of whole grain and bran of white, red and black rice. Food Chem. 2018, 240, 212–221. [Google Scholar] [CrossRef]
- Shao, Y.; Jin, L.; Zhang, G.; Lu, Y.; Shen, Y.; Bao, J. Association mapping of grain color, phenolic content, flavonoid content and antioxidant capacity in dehulled rice. Theor. Appl. Genet. 2011, 22, 1005–1016. [Google Scholar] [CrossRef]
- Mall, A.K.; Swain, P.; Das, S.; Singh, O.N.; Kumar, A. Effect of drought on yield and drought susceptibility index for quality characters of promising rice genotypes. Cer. Res. Commun. 2011, 39, 22–31. [Google Scholar] [CrossRef]
- Fofana, M.; Cherif, M.; Kone, B.; Futakuchi, K.; Audebert, A. Effect of water deficit at grain repining stage on rice grain quality. J. Agric. Biotechnol. Sustain. Dev. Sci. 2010, 9, 287–293. [Google Scholar]
- Sar, S.; Tizzotti, M.J.; Hasjim, J.; Gilbert, R.G. Effects of rice variety and growth location in Cambodia on grain composition and starch structure. Rice Sci. 2014, 21, 47–58. [Google Scholar] [CrossRef]
- Zaman, F.U.; Siddiq, E.A.; Prasad, A.B. Genetical analysis of gel consistency in rice. Indian J. Genet. Plant Breed. 1985, 45, 111–118. [Google Scholar]
- Wang, S.; Callaway, R.M.; Zhou, D.; Weiner, J. Experience of inundation or drought alters the responses of plants to subsequent water conditions. J. Ecol. 2016. [Google Scholar] [CrossRef]
- Walter, J.; Jentsch, A.; Beierkuhnlein, C.; Kreyling, J. Ecological stress memory and cross stress tolerance in plants in the face of climate extremes. Environ. Exp. Bot. 2013, 94, 3–8. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Matthes, M.C.; Napier, J.A.; Pickett, J.A. Stressful ‘‘memories’’ of plants: Evidence and possible mechanisms. Plant Sci. 2007, 173, 603–608. [Google Scholar] [CrossRef]
- Madlung, A.; Comai, L. The effect of stress on genome regulation and structure. Ann. Bot. 2004, 94, 481–495. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Name/code | Popular Name | Characteristics | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Type | Plant Height | Flag Leaf | Panicle Exertion | Tillering Ability | Lifespan (Days) | Potential Yield (t ha−1) | |||
| 1 | N/A | ‘Ingwizabukungu’ | Indica | Intermediate | Intermediate | N/A | N/A | N/A | N/A |
| 2 | WAT 1395-B-24-2 | ‘Intsindagirabigega’ | Indica | Intermediate | Intermediate | Well exerted | Medium | 120–150 | 8.0 |
| 3 | WITA 4 | ‘Jyambere’ | Iindica | Intermediate | Intermediate | Moderate well exerted | Medium | 152 | 10.9 |
| 4 | WAB923-B-6-AL1 | ‘Mpembuke’ | Iindica | Intermediate | Intermediate | Moderate well exerted | Medium | 170 | 8.3 |
| 5 | WAB 569-35-1-1-1-HB | ‘Ndamirabahinzi’ | Indica | Intermediate | Intermediate | Well exerted | Medium | 143 | 7.6 |
| 6 | WAB 880-1-38-20-28-P1-HB | ‘Nemeyubutaka’ | Indica | Intermediate | Erect | Well exerted | Medium | 152 | 9.3 |
| 7 | Zong geng | ‘Kigoli’ | Japonica | Tall | Intermediate | Moderate well exerted | Medium | 180 | 6.0 |
| No of Tillers Plant−1 | Leaf Rolling | Leaf Drying | ||||
|---|---|---|---|---|---|---|
| S | Tl | S | Tl | S | Tl | |
| T | 0.49 | 0.78 | 0.07 | 1.2 | 1.21 | 2.22 |
| D | 7.71 | 6.88 | 65.9 | 43.6 | 70.40 | 59.60 |
| C | 7.71 | 13.6 | 1.0 | 1.4 | 1.64 | 2.16 |
| TD | 2.82 | 0.02 | 10.6 | 12.0 | 5.11 | 2.77 |
| TC | 2.82 | 0.30 | 0.38 | 2.1 | 0.29 | 0.72 |
| DC | 9.24 | 8.85 | 66.9 | 44.8 | 71.70 | 61.00 |
| TDC | 3.17 | 0.08 | 10.9 | 12.3 | 5.31 | 2.95 |
| Cultivar | Recovery Rate (%) | Time to Flowering (DAS) | |
|---|---|---|---|
| S | Tl | ||
| ‘Ingwizabukungu’ | 65.0 a | 38.0 a | 91.6 b |
| ‘Intsindagirabigega’ | 92.5 a | 68.0 a | 113.1 a |
| ‘Jyambere’ | 87.5 a | 52.0 a | 112.3 a |
| ‘Mpembuke’ | 62.5 a | 50.0 a | 113.9 a |
| ‘Ndamirabahinzi’ | 45.0 a | 48.0 a | 105.8 ab |
| ‘Nemeyubutaka’ | 73.3 a | 65.5 a | 107.7 ab |
| ‘Zong geng’ | 60.0 a | 22.2 a | 121.5 a |
| Temperature | |||
| Low | 75.0 a | 54.86 a | 120.5 a |
| High | 64.3 a | 44.57 a | 90.3 b |
| Drought | |||
| D0 | 95.7 a | 87.1 a | 94.8 b |
| DS | 38.6 c | 32.9 c | 111.4 b |
| DST | 64.3 b | 35.7 c | 133.7 a |
| DT | 100.0 a | 41.4 bc | 105.8 b |
| DTR | 98.6 a | 45.7 bc | 111.7 b |
| DR | 95.7 a | 77.1 ab | 103.7 b |
| DSTR | 80.0 ab | 38.6 c | 133.2 a |
| Treatment | Cultivar | Plants per Temp × Drought with No Panicles (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Temp Drought | 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| Low D0 | + | + | - | + | + | + | + | 37.1 ab |
| Low DS | + | + | + | - | - | + | + | 80.0 cde |
| Low DST | + | + | + | + | + | + | + | 57.1 bcd |
| Low DT | + | + | + | + | - | + | - | 74.3 cde |
| Low DTR | + | + | + | - | + | - | - | 74.3 cde |
| Low DR | + | + | - | - | + | - | - | 85.7 de |
| Low DSTR | - | + | + | + | + | + | - | 65.7 bcd |
| High D0 | + | + | + | + | + | + | + | 11.4 a |
| High DS | + | + | + | + | - | - | - | 74.3 cde |
| High DST | - | - | - | - | - | - | - | 100.0 e |
| High DT | + | + | + | + | + | + | - | 54.3 bcd |
| High DTR | + | + | + | + | + | + | - | 65.7 bcd |
| High DR | + | + | + | + | + | + | - | 51.4 bc |
| High DSTR | - | - | - | - | - | - | - | 100.0 e |
| Plants per cultivar with no panicles (%) | 62.9 ab | 50.0 a | 61.4 ab | 68.6 ab | 72.9 bc | 58.6 ab | 91.4 c | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mukamuhirwa, A.; Persson Hovmalm, H.; Bolinsson, H.; Ortiz, R.; Nyamangyoku, O.; Johansson, E. Concurrent Drought and Temperature Stress in Rice—A Possible Result of the Predicted Climate Change: Effects on Yield Attributes, Eating Characteristics, and Health Promoting Compounds. Int. J. Environ. Res. Public Health 2019, 16, 1043. https://doi.org/10.3390/ijerph16061043
Mukamuhirwa A, Persson Hovmalm H, Bolinsson H, Ortiz R, Nyamangyoku O, Johansson E. Concurrent Drought and Temperature Stress in Rice—A Possible Result of the Predicted Climate Change: Effects on Yield Attributes, Eating Characteristics, and Health Promoting Compounds. International Journal of Environmental Research and Public Health. 2019; 16(6):1043. https://doi.org/10.3390/ijerph16061043
Chicago/Turabian StyleMukamuhirwa, Alphonsine, Helena Persson Hovmalm, Hans Bolinsson, Rodomiro Ortiz, Obedi Nyamangyoku, and Eva Johansson. 2019. "Concurrent Drought and Temperature Stress in Rice—A Possible Result of the Predicted Climate Change: Effects on Yield Attributes, Eating Characteristics, and Health Promoting Compounds" International Journal of Environmental Research and Public Health 16, no. 6: 1043. https://doi.org/10.3390/ijerph16061043
APA StyleMukamuhirwa, A., Persson Hovmalm, H., Bolinsson, H., Ortiz, R., Nyamangyoku, O., & Johansson, E. (2019). Concurrent Drought and Temperature Stress in Rice—A Possible Result of the Predicted Climate Change: Effects on Yield Attributes, Eating Characteristics, and Health Promoting Compounds. International Journal of Environmental Research and Public Health, 16(6), 1043. https://doi.org/10.3390/ijerph16061043