

Marine-Derived Polysaccharides and Their Potential Health Benefits in Nutraceutical Applications




Abstract
1. Introduction
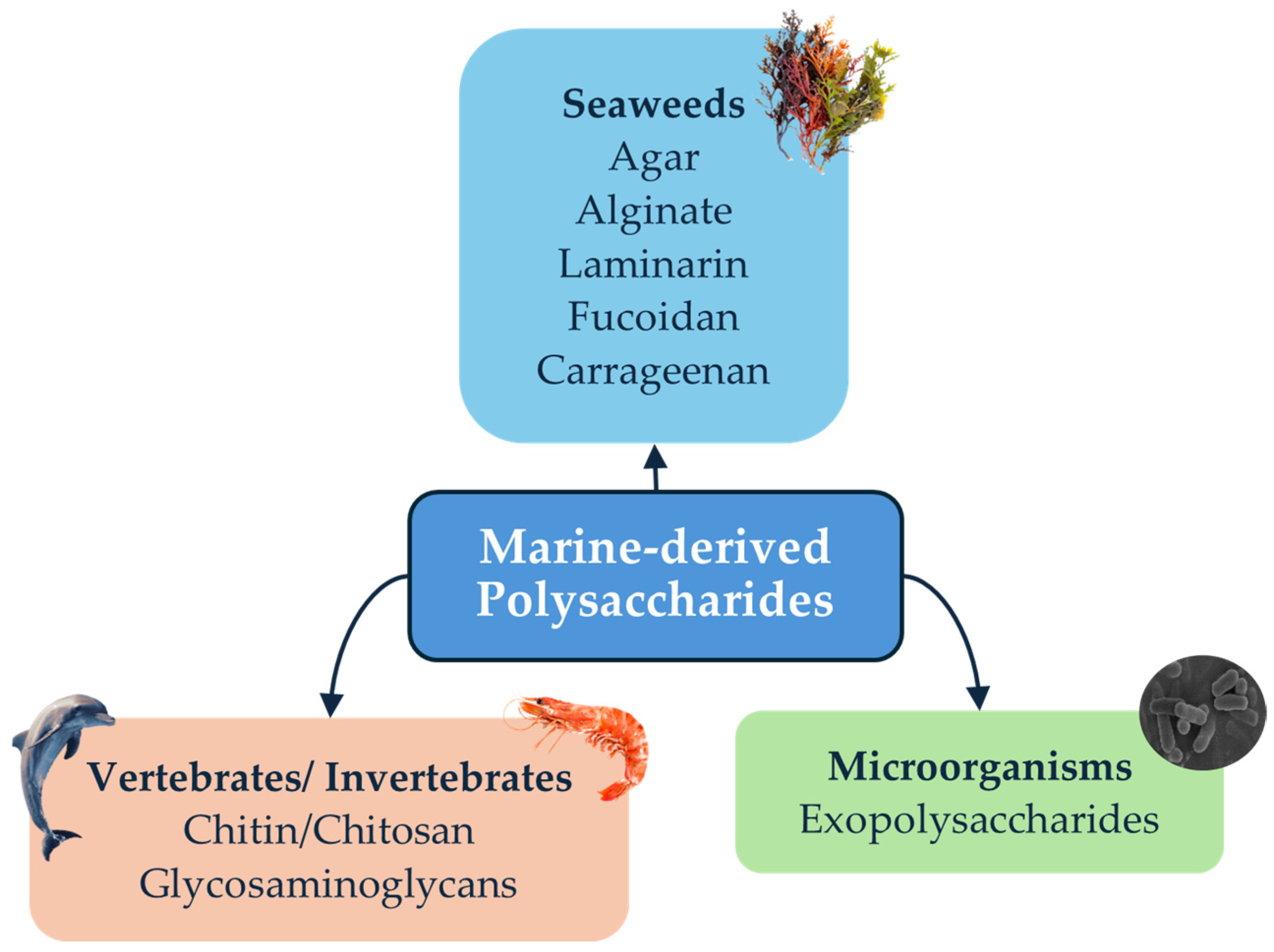
2. Marine-Derived Polysaccharides
2.1. Brown Macroalgae Polysaccharides
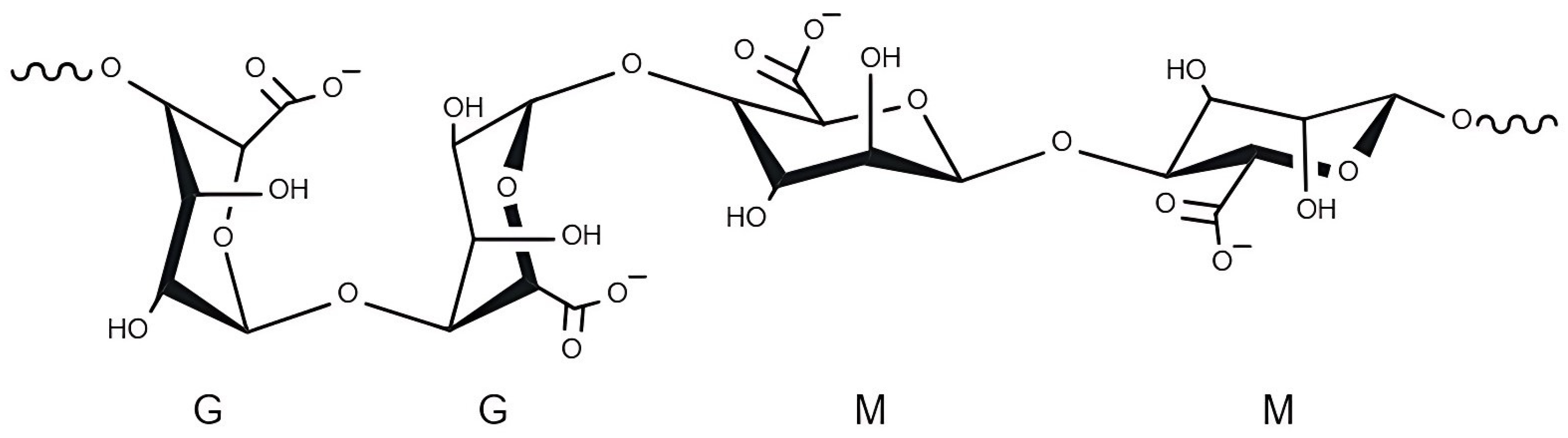
2.1.1. Alginate
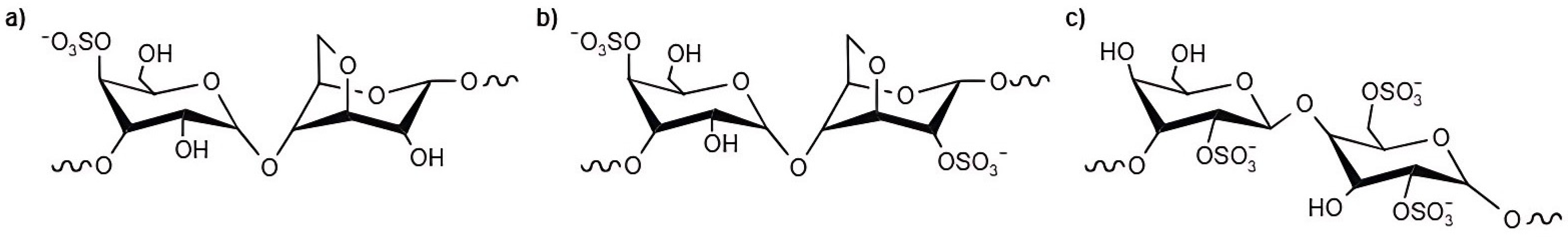
2.1.2. Fucoidan
2.1.3. Laminarin
2.2. Red Macroalgae Polysaccharides
2.2.1. Agar
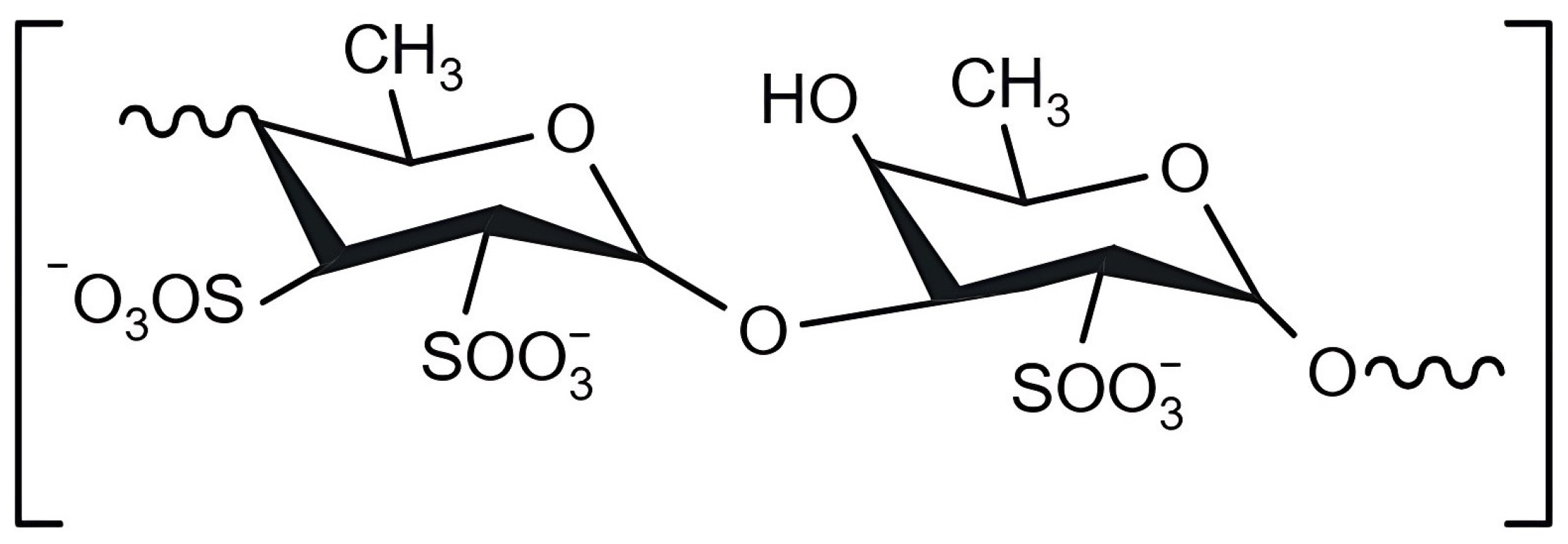
2.2.2. Carrageenan
2.3. Marine Invertebrate and Vertebrate Polysaccharides
2.3.1. Chitin/Chitosan
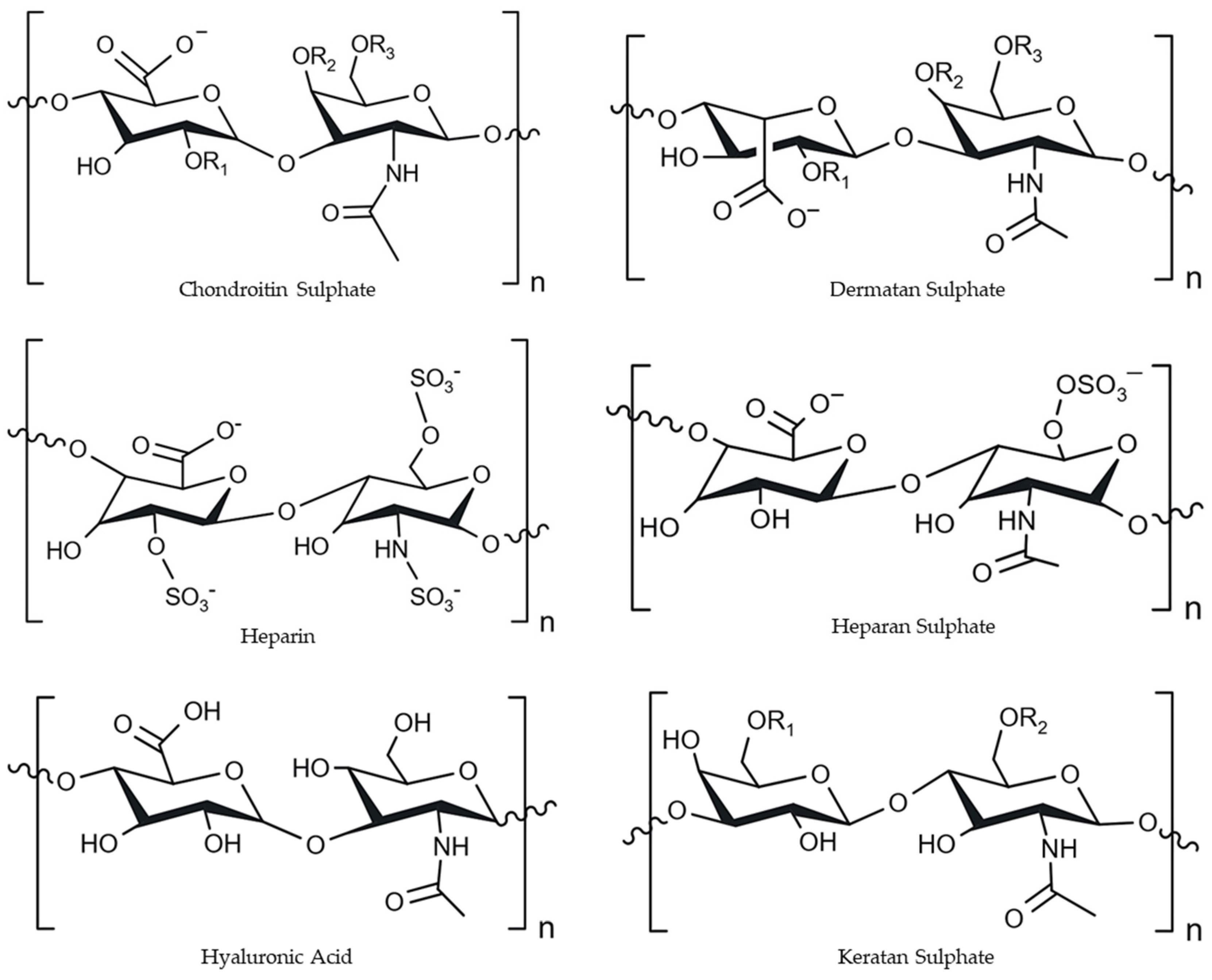
2.3.2. Glycosaminoglycans
2.4. Marine Microorganisms: Polysaccharides
Exopolysaccharides (EPSs)
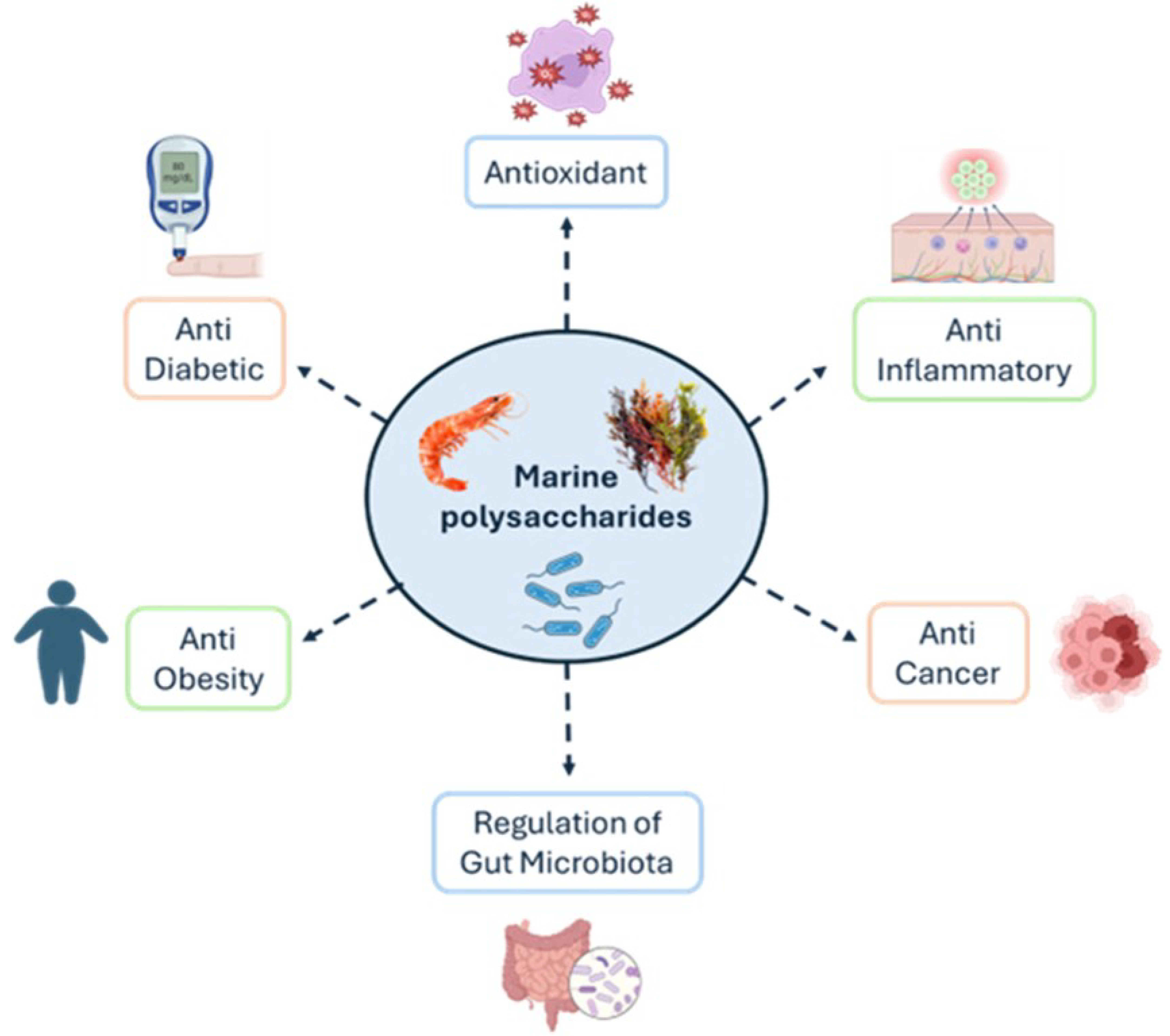
3. Biological Properties of Marine-Derived Polysaccharides
3.1. Antioxidant

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioactive Polysaccharides | Polysaccharide Source | MW | Models | Dose Period | Experimental Method | Results | Ref. |
|---|---|---|---|---|---|---|---|
| Alginate | _ | 50–250 kDa | In vitro | 10−4–1 (w/v) | ABTS and SOD assays | Radical scavenging activity | [92] |
| Fucoidan | Fucus vesiculosus | _ | Hyperoxaluric Wistar rats | 5 mg/kg body weight, 28 days | Estimation of plasma malondialdehyde (MDA) | Decrease in MDA levels | [93] |
| Laminarin | _ | _ | Porcine early-stage embryo | 20 μg/mL | Intracellular ROS levels and GSH assay | ROS scavenging activity, increase in GSH levels | [94] |
| Laminarin | Laminaria hyperborea | 5.7–6.2 kDa | In vitro | 1 mg/mL | DPPH and FRAP assays | Radical scavenging activity | [95] |
| Agar | Gracilaria tenuistipitata | _ | In vitro | 1–10 mg/mL | DPPH assay | Radical scavenging activity | [96] |
| Agaro- oligosaccharides | _ | 500–2500 Da | Human hepatocyte L-02 | An array of concentrations (in vitro study) | Intracellular oxidant stress assay | Radical scavenging activity | [101] |
| Mature Wistar rat | 200–600 mg/kg, 10 days (in vivo study) | Biochemical assays | Inhibiting MDA, AST, and ALT | ||||
| Carrageenan | Eucheuma gelatinae | 2.635 MDa and 2.58 MDa | In vitro | 29.19 mg/mL | Total antioxidant and reducing power activity | Reducing power | [97] |
| Chitosan | _ | 100 kDa | Healthy individuals | 540 mg/day, 2 months | Total plasma antioxidant capacity (TPAC), DPPH and ABTS assays | Increase in TPA; Radical scavenging activity | [98] |
| Chitosan | Thenus unimaculatus | _ | In vitro | 0.5–5 mg/mL | DPPH, ferrous ion chelating, and hydroxyl radical scavenging assay | Radical scavenging activity | [89] |
| GAG | Rapana venosa | _ | In vitro | 1–5 mg/mL | ABTS and FRAP assays | Radical scavenging activity | [67] |
| LSP and LMP | Lapemis curtus | 84–89 kDa | In vitro | An array of concentrations | DPPH and ferrous ion chelating assays | Radical scavenging activity | [102] |
| EPS273 | Pseudomonas stutzeri 273 | 190 kDa | In vitro | An array of concentrations | Hydroxyl radical and SOD | Radical scavenging activity | [99] |
| EPS | Leptolyngbya sp. IkmLPT16 | _ | In vitro | 2–100 mg/mL | DPPH assay | Radical scavenging activity | [100] |
3.2. Anti-Inflammatory
| Bioactive Polysaccharides | Polysaccharide Source | MW | Models | Dose, Period | Experimental Method | Results | Ref. |
|---|---|---|---|---|---|---|---|
| Alginate | Macrocystis pyrifera | _ | Rat peritoneal mast cells | 0.01–1 mg/ mL (in vitro study) | Histidine decarboxylase and interleukins-1β, 6, 8, and tumour necrosis factor-α secretion assays | Inhibited histamine release, IL-1β, and TNF-α | [106] |
| Male Wistar rats | 0.25–1 g/kg (in vivo study) | Compound 48/80-induced systemic anaphylactic shock; Passive cutaneous anaphylaxis | Inhibited compound 48/80, inhibited passive cutaneous anaphylaxis | ||||
| Fucoidan | Saccharina japonica | _ | Streptozotocin-induced diabetic rats | 100 mg/kg/day, 2 months | Determination of inflammatory biomarkers | Reduced IL-1β, IL-6, and TNF-α levels | [107] |
| Laminarin | Laminaria spp. | _ | Pigs challenged with dextran sodium sulphate (DSS) | 200 ppm, 35 days | Colonic gene expression | Alterations in the pattern of co-expressed genes | [108] |
| Agaro- oligosaccharides | _ | _ | Colitis-induced C57BL/6 mice | 4 g/kg/day, 6 days | Measurement of myeloperoxidase activity (MPO); Immunosorbent assay of tumour necrosis factor-α | Induced HO-1 expression; Suppressed TNF-α, IL-12, and IL-17A expressions | [113] |
| Chitosan | _ | 3–200 kDa | LPS-induced Raw 264.7 cells | 0–1600 μg/mL (in vitro study) | Determination of NO, IL-6, and TNF-α in the supernatant of the cells | Decreased the levels of NO, IL-6, and TNF-α | [110] |
| DSS-induced UC mice | 150 mg/kg and 300 mg/kg, 10 days (in vivo study) | Determination of IL-6, TNF-α, IL-1β, IL-10, and IgG in serum of mice | Decreased inflammatory cell infiltration and IL-6, TNF-α, IL-1β, IL-10, and IgG levels | ||||
| PVP-2 | Perna viridis | _ | LPS-induced Raw 264.7 cells | 1–10 μg/mL (in vitro study) | COX and 5-LOX inhibition assays | Decreased the levels of COX 1 and 2, 5-LOX, and NO in the cells | [111] |
| Carrageenan-induced paw oedema mice | 22–110 mg/kg body weight (BW), 10 days (in vivo study) | Determination of NO Effects of PVP-2 in carrageenan-induced paw oedema | Reduced paw oedema in mice | ||||
| D-SBSG | Aristichthys nobilis | 4.96 kDa | Raw 264.7 cells | 0.25–1 mg/mL | NO and cytokine secretion assays ROS generation assay | Decreased the levels of NO, IL-6, IL-1β, IL-10 and ROS | [114] |
| EPSSE | Pseudoalteromonas shioyasakiensis SE | 8 kDa | In vitro | 0.01–100 μg/mL | N, N, N′, N′-tetramethyl-p-phenylenediamine (TMPD) assay method | Inhibited COX-1 and COX-2 | [112] |
3.3. Anti-Cancer
| Bioactive Polysaccharides | Polysaccharide Source | MW | Models | Dose, Period | Experimental Method | Results | Ref. |
|---|---|---|---|---|---|---|---|
| Alginate | _ | _ | Breast cancer MCF-7 cell lines | 6.25–50 μg/mL | 3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide (MTT) assay | Ca-alginate-based AGN-loaded beads decreased MCF-7 cell viability | [119] |
| Fucoidan | Sargassum cinereum | _ | Colon cancer cell line Caco-2 | 10–1000 μg/mL | MTT assay | Inhibited growth of Caco-2 cells | [122] |
| Laminarin | Turbinaria ornata | 689–2634 Da | Retinoblastoma Y79 cancer cells | An array of concentrations | MTT assay | Inhibited growth of Y79 cells | [120] |
| Laminarin | _ | _ | HepG2 cells | 10–40 μM | MTT assay | Inhibited autophagy; Induced apoptosis | [121] |
| Agar | Laminaria digitata | _ | DMBA-induced skin cancer mice | 60–120 mg/mL, 63 days (in vivo study) | In Vivo Anticancer Assay | Normalised food intake, water intake, body weight | [123] |
| NTT 3T3 mice fibroblast cells | An array of concentrations (in vitro study) | MTT Assay | Inhibited tumour development; Inhibited growth of NTT 3T3 cells | ||||
| Agaro-oligosaccharides | _ | _ | HCT-116 cells | 10–100 µg/mL (MTT assay) | MTT Assay | Inhibited the growth of HCT-116 cells | [128] |
| 0–100 µg/mL (DAPI assay) | 40,6-Diamidino-2-Phenylindole (DAPI) staining assay | Induced apoptosis | |||||
| ʎ Carrageenan Oligosaccharides | Kappaphycopsis cottonii | 2 kDa | BGC-823 cells BALB/c mice | 1–20 μg/mL | Cell counting kit-8 (CCK-8) assay Annexin V/PI test; Neutral red uptake assay; In vivo phagocytosis assay | Increased TNF-α and IFN-γ levels; Induced apoptosis; Improved spleen and thymus indexes; Inhibited the growth of BGC-823-derived tumours | [129] |
| Chitosan | _ | 600–800 kDa and 100–300 kDa | MCF-7, HeLa and Saos-2 cells | 0.25–4 mg/mL | MTT assay | Inhibited cell proliferation in all cell lines | [125] |
| 1–2 mg/mL | Annexin V/PI test | Induced apoptosis and necrosis | |||||
| Chondroitin sulphate | Litopenaeus vannamei | _ | Murine melanoma cells (B16F10) | 25–100 μg/mL | Cell death, colony formation, wound healing, Transwell migration, and Matrigel endothelial cell tube formation assays; TNF-α quantification | Reduced tumour colony formation, cell migration, and tubular structure formation; Decreased melanin and TNF-α levels | [126] |
| GAG | Oreochromis niloticus | _ | HTC and SH-SY5Y cells | 1–100 μg/mL | MTT assay | Inhibited growth of HTC and SH-SY5Y cell lines | [130] |
| EPS | Bacillus sp. | 37.6–51.9 kDa | HepG2 cells | 125–1000 μg/mL | Neutral red uptake assay | Cytotoxic activity towards HepG2 cells | [127] |
3.4. Regulation of Gut Microbiota
| Bioactive Polysaccharides | Polysaccharide Source | MW | Models | Dose, Period | Experimental Method | Results | Ref. |
|---|---|---|---|---|---|---|---|
| Alginate oligosaccharide | _ | _ | HFD-fed C57BL/6J mice | 5 g/100 g mice feed, 10 days | Histopathological examination; Gut microbiota analysis; SCFAs, serum biochemical analysis; Insulin and endotoxin quantifications | Reduced fat accumulation and inflammation in the liver; Regulated intestinal flora composition; Reduced TG and LDL-C levels; Increased SCFAs levels | [134] |
| Fucoidan | Laminaria japonica | 250 kDa | CTX treated mice | 20 or 40 mg/kg BW, 19 days | Histopathological examination; Gut microbiota analysis | Reversed the damage in the spleen and thymus; Regulated intestinal flora composition | [135] |
| Fucoidan | Laminaria japonica | 14.6 kDa | Diabetes mellitus (DM) model mice | 100 mg/kg, 13 weeks | Serum biochemical analysis; Histopathological examination; Gut microbiota analysis; Non-targeted metabolomics and bile acid analysis | Reduced TG, total cholesterol (TC), and (LDL-C); Reduced liver damage; Regulated intestinal flora composition; Modulated bile acid content | [136] |
| SHNP (laminarin-type β-(1→3)-glucan) | Sargassum henslowianum | 8.4 kDa | Faecal inocula from healthy individuals | 20 mg/mL (in vitro study) | In vitro fermentation of human faecal microbiota; SCFA analysis | Regulated intestinal flora composition; Increased SCFA levels | [137] |
| Neoagarotetraose, NA4 (from agar) | _ | 630 Da | HFD-fed C57BL/6 mice | 200–800 mg/kg/day, 12 weeks | Biochemical assays; Histochemical staining SCFA analysis; Gut microbiota analysis; Real-time quantitative PCR | Reduced insulin resistance and improved glucose tolerance; Reduced fat deposition and ballooning in hepatocytes; Increased SCFA levels; Regulated intestinal flora composition; Reversed damage to the intestinal barrier | [138] |
| κ Carrageenan | _ | _ | HFD-fed C57BL/6J mice | 0.2–1% (w/w), 8 weeks | Oral glucose tolerance test; Serum biochemical analysis; Histopathological examination; RT-qPCR; SCFA analysis | Reduced weight gain; Reduced TG, TC, and LDL-C; Reversed damage to the intestinal barrier; Regulated intestinal flora composition; Increased ratio of p-AMPK/AMPK; Increased SCFA levels | [139] |
| Chitin derivatives | _ | 20–50 kDa | DSS-induced UC C57BL/6 mice | An array of concentrations 14 days | Disease activity index (DAI) analysis; Enzyme-linked immunosorbent assay (ELISA); RT-qPCR; SCFA analysis | Reduced weight loss; The colonic stratified structure remained mostly intact; Reversed damage to the intestinal barrier; Regulated intestinal flora composition; Increased SCFA levels | [140] |
| Chitosan | _ | _ | Grass carp infected with F. columnare Grass carp liver cells (L8824) | 0.03 g/1000 g feed (in vivo study), 56 days | Serum biochemical analysis; Histopathological examination; 16s rRNA gene; sequencing | Reduction in mortality and increased immunity; Improved intestinal flora diversity | [142] |
| SBSG | Aristichthys nobilis | 54 kDa | In vitro | 0–2 mg/mL (in vitro study) | Simulated in vitro digestion and in vitro evaluation of prebiotic activity | SBSG does not degrade in the upper gastrointestinal tract; Regulated intestinal flora composition | [141] |
| C57BL/6J mice | 50–200 mg/kg (in vivo study) 38 days | Histopathological examination | Reversed the damage in the jejunum | ||||
| EPS | Lactic acid bacteria | _ | HT-29 epithelial cells | 0.001–1% (w/v) | Digestibility assay; Growth and EPS use by probiotic bacteria; Fermentation through a continuous culture system; SCFA analysis | Growth of probiotic bacteria and inhibition of food-borne pathogens; Production of vitamins B1, B2, and folic acid; Increased SCFA levels | [87] |
3.5. Anti-Obesity and Anti-Diabetic Activity
| Bioactive Polysaccharides | Polysaccharide Source | MW | Models | Doses | Experimental Method | Results | Ref. |
|---|---|---|---|---|---|---|---|
| Alginate | Laminaria japonica | 110 kDa | HFD-fed BALB/c mice | 0.3%, 11 weeks | Serum biochemical analysis; Histopathological examination; Faecal transplantation; SCFA analysis | Reduced TG, TC, andLDL-C levels; Reduced weight gain, fat accumulation in the liver, and epididymal adipose tissue; Regulated intestinalflora composition; Increased SCFA levels | [143] |
| Fucoidan | Sargassum fusiforme | 205.8 kDa | HFD- and streptozotocin (STZ)-induced T2DM mice | 100 mg/kg; BW/day, 1 month | Intraperitoneal glucose tolerance test (IPGTT); Serum biochemical analysis; Histopathological examination; 16s rRNA gene sequencing | Decreased polydipsia and polyphagia; Reduced TG, TC, and LDL-C levels; Reduced weight gain, fat accumulation in the liver, heart, and adipose tissue; Regulated intestinal flora composition | [146] |
| Laminarioligosaccharides | Laminaria digitata | _ | In vitro | 10–30 μg | α-Amylase and α-glucosidase inhibition assay | Inhibited α-amylase and α-glucosidase | [147] |
| Laminarin | _ | _ | HFD-fed BALB/c mice | 1%, 6 weeks | 16s rRNA gene sequencing; Microbial community analysis; Metagenome sequences analysis | Decreased weight gain Regulated intestinal flora composition | [148] |
| Agaropectin derived oligosaccharides | Gloiopeltis furcata | 1500 Da | HepG2 cells | An array of concentrations | Glucose consumption and insulin sensitivity assays; TG, TC, and superoxide dismutase (SOD) assays | Enhanced insulin sensitivity and glucose metabolism; Decreased accumulation of lipids and improved lipid metabolism | [149] |
| Carrageenan | Eucheuma spinosum | 1398 Da | HFD-fed Wistar rats | 1–3%, 30 days | Serum biochemical analysis | Reduced TG, TC, and LDL-C levels; Decreased weight gain and food availability; Increase in faecal moisture | [151] |
| ι- Carrageenan | Sarconema filiforme | _ | HFD-fed Wistar rats | 5% (~1.05 g/day), 4 months | Serum biochemical analysis; Glucose consumption and insulin tolerance assays; Microbial community analysis | Decreased weight gain, food intake, systolic blood pressure, and TC; Regulated intestinal flora composition | [53] |
| Thiolated chitosan | _ | _ | IEC-6 cells | 6 mg/mL (in vitro study) | In vitro mucus adhesion test | Exhibited mucoadhesive properties | [152] |
| HFD-fed male C57BL/6 mice | 250 mg/kg/ day (in vivo study), 8 weeks | Oral glucose tolerance test (OGTT) and intraperitoneal glucose tolerance test (IPGTT); In vivo adhesion test; Serum biochemical analysis | Reduced lipid accumulation; Reduced TG, TC, and LDL-C levels; Controlled weight gain | ||||
| Chondroitin sulphate oligosaccharides | Raja pulchra | 0.46–250 kDa | 3T3-L1 preadipocyte cells | An array of concentrations (in vitro study) | In vitro pancreatic lipase assay; Triglyceride E-test kit | Inhibition of lipase activity, absorption of TG, and lipid accumulation | [153] |
| HFD-fed male C57BL/6 mice | 50 mg/ 5 mL/kg/day (in vivo study), 8 weeks | Serum biochemical analysis | Decreased full body, liver, and adipose tissue weight | ||||
| EPS | Streptomyces vinaceusdrappus | 51 kDa | In vitro | 1.95 to 1000 μg/mL | Inhibition of lipase enzyme assay; α-Amylase and α-glucosidase inhibition assay | Inhibited lipase enzyme, α-amylase, and α-glucosidase | [154] |
| PJ1-1 | Penicillium janthinellum N29 | 10.24 kDa | In vitro | 0.08–5.00 mg/mL (in vitro study) | α-glucosidase inhibitory assay | Inhibited α-glucosidase | |
| Type 2 diabetes C57BL/6J male mice | 100–400 mg/kg/day, 35 days (in vivo study) | Serum biochemical analysis of mice; Glucose consumption and insulin sensitivity assays | Decreased lipid accumulation; Reduced TG, TC, and LDL-C levels, and increased HDL; Increased insulin sensitivity and glucose tolerance and reduced blood glucose level | [155] |
4. Functional Properties of Marine-Derived Polysaccharides
5. Applications of Marine Polysaccharides in Nutraceuticals
6. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanjra, M.A.; Qureshi, M.E. Global water crisis and future food security in an era of climate change. Food Policy 2010, 35, 365–377. [Google Scholar] [CrossRef]
- FAO—Dietary Guidelines and Sustainability. Available online: https://www.fao.org/nutrition/education/food-dietary-guidelines/background/sustainable-dietary%20guidelines/en/ (accessed on 8 October 2024).
- Tavares, J.O.; Cotas, J.; Valado, A.; Pereira, L. Algae Food Products as a Healthcare Solution. Mar. Drugs 2023, 21, 578. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.F.; Rezaei, M.; McClements, D.J. Bioactive functional ingredients from aquatic origin: A review of recent progress in marine-derived nutraceuticals. Crit. Rev. Food Sci. Nutr. 2022, 62, 1242–1269. [Google Scholar] [CrossRef]
- Šimat, V.; Elabed, N.; Kulawik, P.; Ceylan, Z.; Jamroz, E.; Yazgan, H.; Čagalj, M.; Regenstein, J.M.; Özogul, F. Recent advances in marine-based nutraceuticals and their health benefits. Mar. Drugs 2020, 18, 627. [Google Scholar] [CrossRef] [PubMed]
- Pop, O.L.; Salanţă, L.C.; Pop, C.R.; Coldea, T.; Socaci, S.A.; Suharoschi, R.; Vodnar, D.C. Prebiotics and Dairy Applications. In Dietary Fiber: Properties, Recovery, and Applications; Elsevier: Amsterdam, The Netherlands, 2019; pp. 247–277. [Google Scholar] [CrossRef]
- Cotas, J.; Leandro, A.; Pacheco, D.; Gonçalves, A.M.M.; Pereira, L. A comprehensive review of the nutraceutical and therapeutic applications of red seaweeds (Rhodophyta). Life 2020, 10, 19. [Google Scholar] [CrossRef]
- Grand View Research. Available online: https://www.grandviewresearch.com/industry-analysis/nutraceuticals-market (accessed on 8 October 2024).
- Senadheera, T.R.L.; Hossain, A.; Shahidi, F. Marine Bioactives and Their Application in the Food Industry: A Review. Appl. Sci. 2023, 13, 12088. [Google Scholar] [CrossRef]
- Barzkar, N.; Babich, O.; Das, R.; Sukhikh, S.; Tamadoni Jahromi, S.; Sohail, M. Marine Bacterial Dextranases: Fundamentals and Applications. Molecules 2022, 27, 5533. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Dai, Z.; Zhang, W.; Fan, S.; Liu, H.; Liu, R.; Zhao, T. Antiobesity, Antidiabetic, Antioxidative, and Antihyperlipidemic Activities of Bioactive Seaweed Substances. In Bioactive Seaweeds for Food Applications: Natural Ingredients for Healthy Diets; Elsevier: Amsterdam, The Netherlands, 2018; pp. 239–253. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S.K. Application of Marine Algae Derived Nutraceuticals in the Food Industry. Mar. Algae Extr. Process. Prod. Appl. 2015, 2, 627–638. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Osborne, S.; Masci, P.; Gobe, G. Marine-based nutraceuticals: An innovative trend in the food and supplement industries. Mar. Drugs 2015, 13, 6336–6351. [Google Scholar] [CrossRef] [PubMed]
- El-Beltagi, H.S.; Mohamed, A.A.; Mohamed, H.I.; Ramadan, K.M.A.; Barqawi, A.A.; Mansour, A.T. Phytochemical and Potential Properties of Seaweeds and Their Recent Applications: A Review. Mar. Drugs 2022, 20, 342. [Google Scholar] [CrossRef] [PubMed]
- Sudha, P.N.; Aisverya, S.; Nithya, R.; Vijayalakshmi, K. Industrial Applications of Marine Carbohydrates. In Advances in Food and Nutrition Research, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 73, pp. 145–181. [Google Scholar] [CrossRef]
- Ruocco, N.; Costantini, S.; Guariniello, S.; Costantini, M. Polysaccharides from the marine environment with pharmacological, cosmeceutical and nutraceutical potential. Molecules 2016, 21, 551. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.B.A.; Adel, M.; Karimi, P.; Peidayesh, M. Pharmaceutical, Cosmeceutical, and Traditional Applications of Marine Carbohydrates. In Advances in Food and Nutrition Research, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 73, pp. 197–220. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Nagahawatta, D.P.; Fernando, I.P.S.; Kim, Y.T.; Kim, J.S.; Kim, W.S.; Lee, J.S.; Jeon, Y.J. A Review on Fucoidan Structure, Extraction Techniques, and Its Role as an Immunomodulatory Agent. Mar. Drugs 2022, 20, 755. [Google Scholar] [CrossRef]
- Zargarzadeh, M.; Amaral, A.J.R.; Custódio, C.A.; Mano, J.F. Biomedical applications of laminarin. Carbohydr. Polym. 2020, 232, 115774. [Google Scholar] [CrossRef] [PubMed]
- Cotas, J.; Pacheco, D.; Araujo, G.S.; Valado, A.; Critchley, A.T.; Pereira, L. On the Health Benefits vs. Risks of Seaweeds and Their Constituents: The Curious Case of the Polymer Paradigm. Mar. Drugs 2021, 19, 164. [Google Scholar] [CrossRef]
- Mortensen, A.; Aguilar, F.; Crebelli, R.; Di Domenico, A.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; Lambrée, C.; Leblanc, J.C.; et al. Re-evaluation of agar (E 406) as a food additive. EFSA J. 2016, 14, e04645. [Google Scholar] [CrossRef]
- Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; Kuhnle, G.G.; et al. Re-evaluation of alginic acid and its sodium, potassium, ammonium and calcium salts (E 400-E 404) as food additives. EFSA J. 2017, 15, e05049. [Google Scholar] [CrossRef]
- Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; Kuhnle, G.G.; et al. Re-evaluation of carrageenan (E 407) and processed Eucheuma seaweed (E 407a) as food additives. EFSA J. 2018, 16, e05238. [Google Scholar] [CrossRef]
- Dobrinčić, A.; Balbino, S.; Zorić, Z.; Pedisić, S.; Kovačević, D.B.; Garofulić, I.E.; Dragović-Uzelac, V. Advanced technologies for the extraction of marine brown algal polysaccharides. Mar. Drugs 2020, 18, 168. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, structure and biofunctional activities of laminarin from brown algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Moussian, B. Chitin: Structure, Chemistry and Biology. In Targeting Chitin-containing Organisms; Advances in Experimental Medicine and Biology; Yang, Q., Fukamizo, T., Eds.; Springer: Singapore, 2019; Volume 1142, pp. 5–18. [Google Scholar] [CrossRef]
- Ganesan, A.R.; Saravana Guru, M.; Balasubramanian, B.; Mohan, K.; Chao Liu, W.; Valan Arasu, M.; Abdullah Al-Dhabi, N.; Duraipandiyan, V.; Ignacimuthu, S.; Sudhakar, M.P.; et al. Biopolymer from edible marine invertebrates: A potential functional food. J. King Saud Univ. Sci. 2020, 32, 1772–1777. [Google Scholar] [CrossRef]
- Guan, X.; Wang, F.; Zhou, B.; Sang, X.; Zhao, Q. The nutritional function of active polysaccharides from marine animals: A review. Food Biosci. 2024, 58, 103693. [Google Scholar] [CrossRef]
- Orlińska, K.; Komosińska-Vassev, K.; Olczyk, K.; Kowalczyk, A.; Olczyk, P. Glycosaminoglycans—Types, structure, functions, and the role in wound healing processes. Ann. Acad. Med. Silesiens. 2023, 77, 204–216. [Google Scholar] [CrossRef]
- Zaghloul, E.H.; Ibrahim, M.I.A. Production and Characterization of Exopolysaccharide from Newly Isolated Marine Probiotic Lactiplantibacillus plantarum EI6 with in vitro Wound Healing Activity. Front. Microbiol. 2022, 13, 903363. [Google Scholar] [CrossRef] [PubMed]
- Jeewon, R.; Aullybux, A.A.; Puchooa, D.; Nazurally, N.; Alrefaei, A.F.; Zhang, Y. Marine Microbial Polysaccharides: An Untapped Resource for Biotechnological Applications. Mar. Drugs 2023, 21, 420. [Google Scholar] [CrossRef] [PubMed]
- Roca, C.; Lehmann, M.; Torres, C.A.; Baptista, S.; Gaudêncio, S.P.; Freitas, F.; Reis, M.A. Exopolysaccharide production by a marine Pseudoalteromonas sp. strain isolated from Madeira Archipelago ocean sediments. N. Biotechnol. 2016, 33, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Lakmal, H.H.C.; Lee, J.H.; Jeon, Y.J. Enzyme-Assisted Extraction of a Marine Algal Polysaccharide, Fucoidan and Bioactivities. In Polysaccharides: Bioactivity and Biotechnology; Ramawat, K., Mérillon, J.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1065–1077. [Google Scholar] [CrossRef]
- Nussinovitch, A. Alginates. In Hydrocolloid Applications, Gum Technology in the Food and Other Industries, 1st ed.; Springer: New York, NY, USA, 1997; pp. 19–39. [Google Scholar] [CrossRef]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr. Polym. 2007, 69, 530–537. [Google Scholar] [CrossRef]
- Abraham, R.E.; Su, P.; Puri, M.; Raston, C.L.; Zhang, W. Optimisation of biorefinery production of alginate, fucoidan and laminarin from brown seaweed Durvillaea potatorum. Algal Res. 2019, 38, 101389. [Google Scholar] [CrossRef]
- Hernandez-Carmona, G.; Freile-Pelegrin, Y.; Garibay, E. Conventional and Alternative Technologies for the Extraction of Algal Polysaccharides. In Functional Ingredients from Algae for Foods and Nutraceuticals; Domínguez, H., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 475–516. [Google Scholar] [CrossRef]
- Sterner, M.; Gröndahl, F. Extraction of laminarin from Saccharina latissima seaweed using cross-flow filtration. J. Appl. Phycol. 2021, 33, 1825–1844. [Google Scholar] [CrossRef]
- Skjåk-Bræk, G.; Draget, K.I. Alginates: Properties and Applications. In Polymer Science: A Comprehensive Reference; Matyjaszewski, K., Möller, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 10, pp. 213–219. [Google Scholar] [CrossRef]
- Hahn, T.; Lang, S.; Ulber, R.; Muffler, K. Novel procedures for the extraction of fucoidan from brown algae. Process Biochem. 2012, 47, 1691–1698. [Google Scholar] [CrossRef]
- Rodriguez-Jasso, R.M.; Mussatto, S.I.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Microwave-assisted extraction of sulfated polysaccharides (fucoidan) from brown seaweed. Carbohydr. Polym. 2011, 86, 1137–1144. [Google Scholar] [CrossRef]
- Zayed, A.; Ulber, R. Fucoidans: Downstream processes and recent applications. Mar. Drugs 2020, 18, 170. [Google Scholar] [CrossRef] [PubMed]
- Sellimi, S.; Maalej, H.; Rekik, D.M.; Benslima, A.; Ksouda, G.; Hamdi, M.; Sahnoun, Z.; Li, S.; Nasri, M.; Hajji, M. Antioxidant, antibacterial and in vivo wound healing properties of laminaran purified from Cystoseira barbata seaweed. Int. J. Biol. Macromol. 2018, 119, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Carpena, M.; Garcia-Perez, P.; Garcia-Oliveira, P.; Chamorro, F.; Otero, P.; Lourenço-Lopes, C.; Cao, H.; Simal-Gandara, J.; Prieto, M.A. Biological Properties and Potential of Compounds Extracted from Red Seaweeds. In Phytochemistry Reviews; Springer: Berlin/Heidelberg, Germany, 2022; Volume 22, pp. 1–32. [Google Scholar] [CrossRef]
- Zhu, B.; Ni, F.; Xiong, Q.; Yao, Z. Marine oligosaccharides originated from seaweeds: Source, preparation, structure, physiological activity and applications. Crit. Rev. Food Sci. Nutr. 2021, 61, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Imeson, A. Agar. In Food Stabilisers, Thickeners and Gelling Agents; Imeson, A., Ed.; Wiley-Blackwell: New Jersey, NJ, USA, 2009; pp. 31–49. [Google Scholar] [CrossRef]
- Torres, M.D.; Flórez-Fernández, N.; Domínguez, H. Integral Utilization of Red Seaweed for Bioactive Production. Mar. Drugs 2019, 17, 314. [Google Scholar] [CrossRef] [PubMed]
- Ścieszka, S.; Klewicka, E. Algae in food: A general review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3538–3547. [Google Scholar] [CrossRef] [PubMed]
- Armisén, R.; Galatas, F. Agar. In Handbook of Hydrocolloids, 2nd ed.; Phillips, G.O., Williams, P.A., Eds.; Woodhead Publishing: Sawston, UK, 2009; pp. 82–107. [Google Scholar] [CrossRef]
- Cian, R.E.; Drago, S.R.; de Medina, F.S.; Martínez-Augustin, O. Proteins and Carbohydrates from Red Seaweeds: Evidence for Beneficial Effects on Gut Function and Microbiota. Mar. Drugs 2015, 13, 5358–5383. [Google Scholar] [CrossRef]
- Prasetyaningrum, A.; Praptyana, I.R.; Nurfiningsih; Ratnawati. Carrageenan: Nutraceutical and functional food as future food. IOP Conf. Ser. Earth Environ. Sci. 2019, 292, 012068. [Google Scholar] [CrossRef]
- Pangestuti, R.; Kim, S.K. Biological activities of carrageenan. Adv. Food Nutr. Res. 2014, 72, 113–124. [Google Scholar] [CrossRef] [PubMed]
- du Preez, R.; Paul, N.; Mouatt, P.; Majzoub, M.E.; Thomas, T.; Panchal, S.K.; Brown, L. Carrageenans from the Red Seaweed Sarconema filiforme Attenuate Symptoms of Diet-Induced Metabolic Syndrome in Rats. Mar. Drugs 2020, 18, 97. [Google Scholar] [CrossRef]
- Dong, Y.; Wei, Z.; Xue, C. Recent advances in carrageenan-based delivery systems for bioactive ingredients: A review. Trends Food Sci. Technol. 2021, 112, 348–361. [Google Scholar] [CrossRef]
- San Jose State Unviversity. Available online: https://mlml.sjsu.edu/geooce/ (accessed on 16 December 2024).
- Aldairi, A.F.; Ogundipe, O.D.; Pye, D.A. Antiproliferative activity of glycosaminoglycan-like polysaccharides derived from marine molluscs. Mar. Drugs 2018, 16, 63. [Google Scholar] [CrossRef]
- Yamada, S.; Sugahara, K.; Özbek, S. Evolution of glycosaminoglycans: Comparative biochemical study. Commun. Integr. Biol. 2011, 4, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed]
- Vieira, H.; Lestre, G.M.; Solstad, R.G.; Cabral, A.E.; Botelho, A.; Helbig, C.; Coppola, D.; de Pascale, D.; Robbens, J.; Raes, K.; et al. Current and Expected Trends for the Marine Chitin/Chitosan and Collagen Value Chains. Mar. Drugs 2023, 21, 605. [Google Scholar] [CrossRef] [PubMed]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Dutta, P.K.; Dutta, J.; Tripathi, V.S. Chitin and chitosan: Chemistry, properties and applications. J. Sci. Ind. Res. 2004, 63, 20–31. [Google Scholar]
- Aranaz, I.; Mengibar, M.; Harris, R.; Panos, I.; Miralles, B.; Acosta, N.; Galed, G.; Heras, A. Functional Characterization of Chitin and Chitosan. Curr. Chem. Biol. 2012, 3, 203–230. [Google Scholar] [CrossRef]
- Iber, B.T.; Kasan, N.A.; Torsabo, D.; Omuwa, J.W. A review of various sources of chitin and chitosan in nature. J. Renew. Mater. 2022, 10, 1097–1123. [Google Scholar] [CrossRef]
- Piekarska, K.; Sikora, M.; Owczarek, M. Chitin and Chitosan as Polymers of the Future—Obtaining, Modification, Life Cycle Assessment and Main Directions of Application. Polymers 2023, 15, 793. [Google Scholar] [CrossRef] [PubMed]
- Khurshid, C.; Pye, D.A. Isolation and Composition Analysis of Bioactive Glycosaminoglycans from Whelk. Mar. Drugs 2018, 16, 171. [Google Scholar] [CrossRef]
- Shen, S.; Chen, X.; Shen, Z.; Chen, H. Marine polysaccharides for wound dressings application: An overview. Pharmaceutics 2021, 13, 1666. [Google Scholar] [CrossRef] [PubMed]
- Gaspar-Pintiliescu, A.; Stefan, L.M.; Mihai, E.; Sanda, C.; Manoiu, V.S.; Berger, D.; Craciunescu, O. Antioxidant and antiproliferative effect of a glycosaminoglycan extract from Rapana venosa marine snail. PLoS ONE 2024, 19, e0297803. [Google Scholar] [CrossRef]
- Choi, B.D.; Choi, Y.J. Nutraceutical functionalities of polysaccharides from marine invertebrates. Adv. Food Nutr. Res. 2012, 65, 11–30. [Google Scholar] [CrossRef]
- Prestwich, G.D.; Atzet, S. Engineered Natural Materials. In Biomaterials Science, An Introduction to Materials in Medicine, 3rd ed.; Ratner, B.D., Schoen, F.J., Hoffman, A.S., Lemons, J.E., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 195–209. [Google Scholar] [CrossRef]
- Pavão, M.S.G. Glycosaminoglycans analogs from marine invertebrates: Structure, biological effects, and potential as new therapeutics. Front. Cell. Infect. Microbiol. 2014, 4, 123. [Google Scholar] [CrossRef] [PubMed]
- Ameen, F.; AlNadhari, S.; Al-Homaidan, A.A. Marine microorganisms as an untapped source of bioactive compounds. Saudi J. Biol. Sci. 2021, 28, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Gupta, C.; Prakash, D. Nutraceuticals from Microbes of Marine Sources. In Nutraceuticals—Past, Present and Future; Chávarri-Hueda, M., Ed.; IntechOpen: London, UK, 2019; pp. 1–22. [Google Scholar] [CrossRef]
- Dewapriya, P.; Kim, S. kwon. Marine microorganisms: An emerging avenue in modern nutraceuticals and functional foods. Food Res. Int. 2014, 56, 115–125. [Google Scholar] [CrossRef]
- Qi, M.; Zheng, C.; Wu, W.; Yu, G.; Wang, P. Exopolysaccharides from Marine Microbes: Source, Structure and Application. Mar. Drugs 2022, 20, 512. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Bacterial exopolysaccharides from extreme marine habitats: Production, characterization and biological activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef] [PubMed]
- Andrew, M.; Jayaraman, G. Structural features of microbial exopolysaccharides in relation to their antioxidant activity. Carbohydr. Res. 2020, 487, 107881. [Google Scholar] [CrossRef]
- López-Ortega, M.A.; Chavarría-Hernández, N.; del Rocio Lopez-Cuellar, M.; Rodríguez-Hernández, A.I. A review of extracellular polysaccharides from extreme niches: An emerging natural source for the biotechnology. From the adverse to diverse! Int. J. Biol. Macromol. 2021, 177, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-B.; Chen, X.-L.; He, H.-L.; Zhang, X.-Y.; Xie, B.-B.; Yu, Y.; Chen, B.; Zhou, B.-C.; Zhang, Y.Z. Structure and Ecological Roles of a Novel Exopolysaccharide from the Arctic Sea Ice Bacterium Pseudoalteromonas sp. Strain SM20310. Appl. Environ. Microbiol. 2013, 79, 224–230. [Google Scholar] [CrossRef]
- Wei, M.; Geng, L.; Wang, Q.; Yue, Y.; Wang, J.; Wu, N.; Wang, X.; Sun, C.; Zhang, Q. Purification, characterization and immunostimulatory activity of a novel exopolysaccharide from Bacillus sp. H5. Int. J. Biol. Macromol. 2021, 189, 649–656. [Google Scholar] [CrossRef]
- Arun, J.; Selvakumar, S.; Sathishkumar, R.; Moovendhan, M.; Ananthan, G.; Maruthiah, T.; Palavesam, A. In vitro antioxidant activities of an exopolysaccharide from a salt pan bacterium Halolactibacillus miurensis. Carbohydr. Polym. 2017, 155, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Mao, W.; Chen, C.; Kong, X.; Gu, Q.; Li, N.; Liu, X.; Wang, B.; Wang, S.; Xiao, B. Structural elucidation of the exopolysaccharide produced by the mangrove fungus Penicillium solitum. Carbohydr. Polym. 2014, 111, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Yang, J.; Luo, L.; Zhang, X.; Deng, S.; Chen, X.; Li, Y.; Bekhit, A.E.A.; Xu, B.; Huang, R. Ferroptosis Related Immunomodulatory Effect of a Novel Extracellular Polysaccharides from Marine Fungus Aureobasidium melanogenum. Mar. Drugs 2022, 20, 332. [Google Scholar] [CrossRef] [PubMed]
- Parra-Riofrío, G.; García-Márquez, J.; Casas-Arrojo, V.; Uribe-Tapia, E.; Abdala-Díaz, R. Antioxidant and Cytotoxic Effects on Tumor Cells of Exopolysaccharides from Tetraselmis suecica (Kylin) Butcher Grown Under Autotrophic and Heterotrophic Conditions. Mar. Drugs 2020, 18, 534. [Google Scholar] [CrossRef]
- Bafana, A. Characterization and optimization of production of exopolysaccharide from Chlamydomonas reinhardtii. Carbohydr. Polym. 2013, 95, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Drira, M.; Elleuch, J.; Ben Hlima, H.; Hentati, F.; Gardarin, C.; Rihouey, C.; Le Cerf, D.; Michaud, P.; Abdelkafi, S.; Fendri, I. Optimization of Exopolysaccharides Production by Porphyridium sordidum and Their Potential to In-duce Defense Responses in Arabidopsis thaliana against Fusarium oxysporum. Biomolecules 2021, 11, 282. [Google Scholar] [CrossRef] [PubMed]
- Casillo, A.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Exopolysaccharides from Marine and Marine Extremophilic Bacteria: Structures, Properties, Ecological Roles and Applications. Marine Drugs. 2018, 16, 69. [Google Scholar] [CrossRef] [PubMed]
- Hongpattarakere, T.; Cherntong, N.; Wichienchot, S.; Kolida, S.; Rastall, R.A. In vitro prebiotic evaluation of exopolysaccharides produced by marine isolated lactic acid bacteria. Carbohydr. Polym. 2012, 87, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Arasukumar, B.; Prabakaran, G.; Gunalan, B.; Moovendhan, M. Chemical composition, structural features, surface morphology and bioactivities of chitosan derivatives from lobster (Thenus unimaculatus) shells. Int. J. Biol. Macromol. 2019, 135, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Wei, B.; Wang, S.; Ke, S.; Chen, J.; Zhang, H.; Wang, H. The Antioxidant Activity of Polysaccharides Derived from Marine Organisms: An Overview. Mar. Drugs 2019, 17, 674. [Google Scholar] [CrossRef] [PubMed]
- Kelishomi, Z.H.; Goliaei, B.; Mahdavi, H.; Nikoofar, A.; Rahimi, M.; Moosavi-Movahedi, A.A.; Mamashli, F.; Bigdeli, B. Antioxidant activity of low molecular weight alginate produced by thermal treatment. Food Chem. 2016, 196, 897–902. [Google Scholar] [CrossRef]
- Veena, C.K.; Josephine, A.; Preetha, S.P.; Varalakshmi, P.; Sundarapandiyan, R. Renal peroxidative changes mediated by oxalate: The protective role of fucoidan. Life Sci. 2006, 79, 1789–1795. [Google Scholar] [CrossRef]
- Jiang, H.; Liang, S.; Yao, X.R.; Jin, Y.X.; Shen, X.H.; Yuan, B.; Zhang, J.B.; Kim, N.H. Laminarin improves developmental competence of porcine early-stage embryos by inhibiting oxidative stress. Theriogenology 2018, 115, 38–44. [Google Scholar] [CrossRef]
- Rajauria, G.; Ravindran, R.; Garcia-Vaquero, M.; Rai, D.K.; Sweeney, T.; O’Doherty, J. Molecular characteristics and antioxidant activity of laminarin extracted from the seaweed species Laminaria hyperborea, using hydrothermal-assisted extraction and a multi-step purification procedure. Food Hydrocoll. 2021, 112, 106332. [Google Scholar] [CrossRef]
- Mohibbullah, M.; Talha, M.A.; Baten, M.A.; Newaz, A.W.; Choi, S. Yield optimization, physicochemical characterizations, and antioxidant properties of food grade agar from Gracilaria tenuistipitata of Cox’s Bazar coast, Bangladesh. Food Sci. Nutr. 2023, 11, 2852–2863. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.T.; Cuong, D.X.; Thuy, L.H.; Thuan, P.T.; Tuyen, D.T.T.; Mo, V.T.; Dong, D.H. Carrageenan of Red Algae Eucheuma gelatinae: Extraction, Antioxidant Activity, Rheology Characteristics, and Physicochemistry Characterization. Molecules 2022, 27, 1268. [Google Scholar] [CrossRef] [PubMed]
- Anraku, M.; Fujii, T.; Kondo, Y.; Kojima, E.; Hata, T.; Tabuchi, N.; Tsuchiya, D.; Goromaru, T.; Tsutsumi, H.; Kadowaki, D.; et al. Antioxidant properties of high molecular weight dietary chitosan in vitro and in vivo. Carbohydr. Polym. 2011, 83, 501–505. [Google Scholar] [CrossRef]
- Wu, S.; Liu, G.; Jin, W.; Xiu, P.; Sun, C. Antibiofilm and anti-infection of a marine bacterial exopolysaccharide against Pseudomonas aeruginosa. Front. Microbiol. 2016, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Gongi, W.; Cordeiro, N.; Pinchetti, J.L.G.; Ouada, H. Ben. Functional, rheological, and antioxidant properties of extracellular polymeric substances produced by a thermophilic cyanobacterium Leptolyngbya sp. J. Appl. Phycol. 2022, 34, 1423–1434. [Google Scholar] [CrossRef]
- Chen, H.; Yan, X.; Zhu, P.; Lin, J. Antioxidant activity and hepatoprotective potential of agaro- oligosaccharides in vitro and in vivo. Nutr. J. 2006, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.; Han, W.; Zhao, X.; Wang, Q.; Gao, Y.; Deng, S. Glycosaminoglycans from a Sea Snake (Lapemis curtus): Extraction, Structural Characterization and Antioxidant Activity. Mar. Drugs 2018, 16, 170. [Google Scholar] [CrossRef] [PubMed]
- Elbandy, M. Anti-Inflammatory Effects of Marine Bioactive Compounds and Their Potential as Functional Food Ingredients in the Prevention and Treatment of Neuroinflammatory Disorders. Molecules 2022, 28, 2. [Google Scholar] [CrossRef]
- Reddy, S.M.; Suresh, V.; Pitchiah, S.; Subramanian, B. Anti-inflammatory Activities of Sulfated Polysaccharides from Ethanol Crude Extract of Spyrida Species Red Seaweed. Cureus 2023, 15, e50284. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Herath, K.H.I.N.M.; Yang, H.-W.; Choi, C.S.; Jeon, Y.-J. Anti-Inflammatory Mechanisms of Fucoidans to Treat Inflammatory Diseases: A Review. Mar. Drugs 2021, 19, 678. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Lee, S.A.; Moon, P.D.; Na, H.J.; Park, R.K.; Um, J.Y.; Kim, H.M.; Hong, S.H. Alginic acid has anti-anaphylactic effects and inhibits inflammatory cytokine expression via suppression of nuclear factor-κB activation. Clin. Exp. Allergy 2006, 36, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Aleissa, M.S.; Alkahtani, S.; Abd Eldaim, M.A.; Ahmed, A.M.; Bungau, S.G.; Almutairi, B.; Bin-Jumah, M.; AlKahtane, A.A.; Alyousif, M.S.; Abdel-Daim, M.M. Fucoidan Ameliorates Oxidative Stress, Inflammation, DNA Damage, and Hepatorenal Injuries in Diabetic Rats Intoxicated with Aflatoxin B1. Oxidative Med. Cell. Longev. 2020, 2020, 9316751. [Google Scholar] [CrossRef] [PubMed]
- Rattigan, R.; O’Doherty, J.V.; Vigors, S.; Ryan, M.T.; Sebastiano, R.S.; Callanan, J.J.; Thornton, K.; Rajauria, G.; Margassery, L.M.; Dobson, A.D.W.; et al. The Effects of the Marine-Derived Polysaccharides Laminarin and Chitosan on Aspects of Colonic Health in Pigs Challenged with Dextran Sodium Sulphate. Mar. Drugs 2020, 18, 262. [Google Scholar] [CrossRef] [PubMed]
- Amdekar, S.; Roy, P.; Singh, V.; Kumar, A.; Singh, R.; Sharma, P. Anti-inflammatory activity of lactobacillus on carrageenan-induced paw edema in male wistar rats. Int. J. Inflamm. 2012, 2012, 752015. [Google Scholar] [CrossRef]
- Niu, W.; Dong, Y.; Fu, Z.; Lv, J.; Wang, L.; Zhang, Z.; Huo, J.; Ju, J. Effects of molecular weight of chitosan on anti-inflammatory activity and modulation of intestinal microflora in an ulcerative colitis model. Int. J. Biol. Macromol. 2021, 193 Pt B, 1927–1936. [Google Scholar] [CrossRef]
- Pai, A.A.; Chakraborty, K.; Dhara, S.; Raj, A.; Kariyil, B.J.; Anoopraj, R. Therapeutic potential of sulfated glycosaminoglycan from seafood Asian green mussel (Perna viridis): Insights from an in vivo study. Food Biosci. 2024, 61, 104837. [Google Scholar] [CrossRef]
- Hafez, S.S.; Abdallah, S.A.; Moustafa, E.E. Characterization of exopolysaccharide EPSSE from marine Pseudoalteromonas shioyasakiensis and its biological activity potential. Egypt. J. Aquat. Res. 2024, 50, 272–281. [Google Scholar] [CrossRef]
- Higashimura, Y.; Naito, Y.; Takagi, T.; Mizushima, K.; Hirai, Y.; Harusato, A.; Ohnogi, H.; Yamaji, R.; Inui, H.; Nakano, Y.; et al. Oligosaccharides from agar inhibit murine intestinal inflammation through the induction of heme oxygenase-1 expression. J. Gastroenterol. 2013, 48, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhou, S.; Wang, Z.; Liu, S.; Li, R.; Jia, X.; Chen, J.; Liu, X.; Song, B.; Zhong, S. Anticoagulant and anti-inflammatory effects of a degraded sulfate glycosaminoglycan from swimming bladder. Food Res. Int. 2022, 157, 111444. [Google Scholar] [CrossRef]
- Cotas, J.; Pacheco, D.; Gonçalves, A.M.M.; Silva, P.; Carvalho, L.G.; Pereira, L. Seaweeds’ nutraceutical and biomedical potential in cancer therapy: A concise review. J. Cancer Metastasis Treat. 2021, 7, 13. [Google Scholar] [CrossRef]
- Sheikh, I.; Sharma, V.; Tuli, H.S.; Aggarwal, D.; Sankhyan, A.; Vyas, P.; Sharma, A.K.; Bishayee, A. Cancer chemoprevention by flavonoids, dietary polyphenols and terpenoids. Biointerface Res. Appl. Chem. 2021, 11, 8502–8537. [Google Scholar] [CrossRef]
- Ruan, B.F.; Ge, W.W.; Lin, M.X.; Li, Q.S. A Review of the Components of Seaweeds as Potential Candidates in Cancer Therapy. Anti Cancer Agents Med. Chem. 2018, 18, 354–366. [Google Scholar] [CrossRef]
- Ju, H.; Yu, C.; Liu, W.; Li, H.-H.; Fu, Z.; Wu, Y.-C.; Gong, P.-X.; Li, H.-J. Polysaccharides from marine resources exhibit great potential in the treatment of tumor: A review. Carbohydr. Polym. Technol. Appl. 2023, 5, 100308. [Google Scholar] [CrossRef]
- Aldawsari, M.F.; Ahmed, M.M.; Fatima, F.; Anwer, M.K.; Katakam, P.; Khan, A. Development and characterization of calcium-alginate beads of apigenin: In vitro antitumor, antibacterial, and antioxidant activities. Mar. Drugs 2021, 19, 467. [Google Scholar] [CrossRef] [PubMed]
- Remya, R.R.; Rajasree, S.R.R.; Suman, T.Y.; Aranganathan, L.; Gayathri, S.; Gobalakrishnan, M.; Karthih, M.G. Laminarin based AgNPs using brown seaweed Turbinaria ornata and its induction of apoptosis in human retinoblastoma Y79 cancer cell lines. Mater. Res. Express 2018, 5, 035403. [Google Scholar] [CrossRef]
- Cui, D.; Ma, J.; Liang, T.; Sun, L.; Meng, L.; Liang, T.; Li, Q. Selenium nanoparticles fabricated in laminarin polysaccharides solutions exert their cytotoxicities in HepG2 cells by inhibiting autophagy and promoting apoptosis. Int. J. Biol. Macromol. 2019, 137, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Narayani, S.S.; Saravanan, S.; Ravindran, J.; Ramasamy, M.S.; Chitra, J. In vitro anticancer activity of fucoidan extracted from Sargassum cinereum against Caco-2 cells. Int. J. Biol. Macromol. 2019, 138, 618–628. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Thabitha, A.; Vignesh, N.; Manigandan, V.; Saravanan, R.; Daradkeh, G.; Qoronfleh, M.W. Antiskin Cancer and Antioxidant Activities of Formulated Agar from Brown Seaweed Laminaria digitata (Hudson) in Dimethyl Benzanthracene-Induced Swiss Albino Mice. Int. J. Polym. Sci. 2021, 2021, 9930777. [Google Scholar] [CrossRef]
- Souza, R.B.; Frota, A.F.; Silva, J.; Alves, C.; Neugebauer, A.Z.; Pinteus, S.; Rodrigues, J.A.G.; Cordeiro, E.M.S.; de Almeida, R.R.; Pedrosa, R.; et al. In vitro activities of kappa-carrageenan isolated from red marine alga Hypnea musciformis: Antimicrobial, anticancer and neuroprotective potential. Int. J. Biol. Macromol. 2018, 112, 1248–1256. [Google Scholar] [CrossRef] [PubMed]
- Abedia, Z.; Moghadamnia, A.A.; Zabihi, E.; Pourbagher, R.; Ghasemi, M.; Nouri, H.R.; Tashakorian, H.; Jenabian, N. Anticancer properties of chitosan against osteosarcoma, breast cancer and cervical cancer cell lines. Casp. J. Intern. Med. 2019, 10, 439–446. [Google Scholar] [CrossRef]
- Palhares, L.C.G.F.; Barbosa, J.S.; Scortecci, K.C.; Rocha, H.A.O.; Brito, A.S.; Chavante, S.F. In vitro antitumor and anti-angiogenic activities of a shrimp chondroitin sulfate. Int. J. Biol. Macromol. 2020, 162, 1153–1165. [Google Scholar] [CrossRef]
- Abdelnasser, S.M.; Yahya, S.M.M.; Mohamed, W.F.; Asker, M.M.S.; Abu Shady, H.M.; Mahmoud, M.G.; Gadallah, M.A. Antitumor exopolysaccharides derived from novel marine Bacillus: Isolation, characterization aspect and biological activity. Asian Pac. J. Cancer Prev. 2017, 18, 1847–1854. [Google Scholar] [CrossRef]
- Yun, E.J.; Yu, S.; Kim, Y.A.; Liu, J.J.; Kang, N.J.; Jin, Y.S.; Kim, K.H. In vitro prebiotic and anti-colon cancer activities of agar-derived sugars from red seaweeds. Mar. Drugs 2021, 19, 213. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Zhai, L.; Chen, J.; Wang, F.; Chen, H.; Wu, W. The Antitumor Potential of λ-Carrageenan Oligosaccharides on Gastric Carcinoma by Immunomodulation. Nutrients 2023, 15, 2044. [Google Scholar] [CrossRef]
- de Moura, H.C.; Novello, C.R.; Balbinot-Alfaro, E.; Düsman, E.; Barddal, H.P.O.; Almeida, I.V.; Vicentini, V.E.P.; Prentice-Hernández, C.; Alfaro, A.T. Obtaining glycosaminoglycans from tilapia (oreochromis niloticus) scales and evaluation of its anticoagulant and cytotoxic activities: Glycosaminoglycans from tilapia scales: Anticoagulant and cytotoxic activities. Food Res. Int. 2021, 140, 110012. [Google Scholar] [CrossRef] [PubMed]
- Davey, L.; Halperin, S.A.; Lee, S.F. Thiol-Disulfide Exchange in Gram-Positive Firmicutes. Trends Microbiol. 2016, 24, 902–915. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.-Y.; Zhao, C.-N.; Xu, X.-Y.; Tang, G.-Y.; Corke, H.; Gan, R.-Y.; Li, H.-B. Dietary plants, gut microbiota, and obesity: Effects and mechanisms. Trends Food Sci. Technol. 2019, 92, 194–204. [Google Scholar] [CrossRef]
- Shagdarova, B.; Konovalova, M.; Varlamov, V.; Svirshchevskaya, E. Anti-Obesity Effects of Chitosan and Its Derivatives. Polymers 2023, 15, 3967. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Ye, C.; Yuan, J.; Qin, S. Alginate oligosaccharide improves lipid metabolism and inflammation by modulating gut microbiota in high-fat diet fed mice. Appl. Microbiol. Biotechnol. 2020, 104, 3541–3554. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Pu, Q.; Zhao, Q.; Zhou, Y.; Jiang, X.; Han, T. Effects of Fucoidan Isolated from Laminaria japonica on Immune Response and Gut Microbiota in Cyclophosphamide-Treated Mice. Front. Immunol. 2022, 13, 916618. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, J.; Chen, X.; Xue, T.; Xin, J.; Liu, Y.; Wang, X.; Li, X. Laminaria japonica Polysaccharide Regulates Fatty Hepatosis Through Bile Acids and Gut Microbiota in Diabetes Rat. Mar. Biotechnol. 2024, 26, 1165–1178. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhu, L.; Li, Y.; Jiang, S.; Sun, Q.; Xie, E.; Chen, H.; Zhao, Z.; Qiao, W.; Xu, J.; et al. Structure of a laminarin-type β-(1→3)-glucan from brown algae Sargassum henslowianum and its potential on regulating gut microbiota. Carbohydr. Polym. 2021, 255, 117389. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wang, Q.; Lin, F.; Zheng, B.; Huang, Y.; Yang, Y.; Xue, C.; Xiao, M.; Ye, J. Neoagarotetraose alleviates high fat diet induced obesity via white adipocytes browning and regulation of gut microbiota. Carbohydr. Polym. 2022, 296, 119903. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, L.; He, Y.; Zeng, L.; He, J.; Yang, Y.; Zhang, T. Effect of κ-carrageenan on glucolipid metabolism and gut microbiota in high-fat diet-fed mice. J. Funct. Foods 2021, 86, 104707. [Google Scholar] [CrossRef]
- Mei, Z.; Huang, X.; Zhang, H.; Cheng, D.; Xu, X.; Fang, M.; Hu, J.; Liu, Y.; Liang, Y.; Mei, Y. Chitin derivatives ameliorate DSS-induced ulcerative colitis by changing gut microbiota and restoring intestinal barrier function. Int. J. Biol. Macromol. 2022, 202, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Liu, X.; Huang, H.; Zhong, A.; Guo, X.; Chen, J.; Wang, Z.; Cheong, K.L.; Zhong, S. Potential prebiotic activity and improvement effect on intestinal injury of sulfated glycosaminoglycan from swim bladder. Food Biosci. 2024, 60, 104548. [Google Scholar] [CrossRef]
- Zhou, J.; Feng, M.; Zhang, W.; Kuang, R.; Zou, Q.; Su, J.; Yuan, G. Oral administration of hepcidin and chitosan benefits growth, immunity, and gut microbiota in grass carp (Ctenopharyngodon idella). Front. Immunol. 2022, 13, 1075128. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Duan, M.; Jia, J.; Song, S.; Ai, C. Low-molecular alginate improved diet-induced obesity and metabolic syndrome through modulating the gut microbiota in BALB/c mice. Int. J. Biol. Macromol. 2021, 187, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.; Gastaldelli, A.; Yki-Järvinen, H.; Scherer, P.E. Why does obesity cause diabetes? Cell Metab. 2022, 34, 11–20. [Google Scholar] [CrossRef]
- Dirir, A.M.; Daou, M.; Yousef, A.F.; Yousef, L.F. A review of alpha-glucosidase inhibitors from plants as potential candidates for the treatment of type-2 diabetes. Phytochem. Rev. 2022, 21, 1049–1079. [Google Scholar] [CrossRef]
- Wu, Q.; Wu, S.; Cheng, Y.; Zhang, Z.; Mao, G.; Li, S.; Yang, Y.; Zhang, X.; Wu, M.; Tong, H. Sargassum fusiforme fucoidan modifies gut microbiota and intestinal metabolites during alleviation of hyperglycemia in type 2 diabetic mice. Food Funct. 2021, 12, 3572–3585. [Google Scholar] [CrossRef]
- Jayapala, N.; Toragall, V.; Ganesh Kumar, B.S.; Chaudhari, S.R.; Baskaran, V. Preparation, characterization, radical scavenging property and antidiabetic potential of laminarioligosaccharides derived from laminarin. Algal Res. 2022, 63, 102642. [Google Scholar] [CrossRef]
- Nguyen, S.G.; Kim, J.; Guevarra, R.B.; Lee, J.H.; Kim, E.; Kim, S.I.; Unno, T. Laminarin favorably modulates gut microbiota in mice fed a high-fat diet. Food Funct. 2016, 7, 4193–4201. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, H.; Zhang, N.; Cai, C.; Li, G.; Hao, J. Anti-diabetic activities of agaropectin-derived oligosaccharides from Gloiopeltis furcata via regulation of mitochondrial function. Carbohydr. Polym. 2020, 229, 115482. [Google Scholar] [CrossRef] [PubMed]
- Ernst, N.D.; Obarzanek, E. Child Health and Nutrition: Obesity and High Blood Cholesterol. Prev. Med. 1994, 23, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Zhong, W. Antihyperglycemic and antihyperlipidemic effects of low-molecular-weight carrageenan in rats. Open Life Sci. 2018, 13, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.C.; Ho, Y.Y.; Tang, R.C.; Ke, Y.C.; Lin, J.N.; Yang, I.H.; Lin, F.H. Thiolated chitosan as an intestinal absorption carrier with hesperidin encapsulation for obesity treatment. Nutrients 2021, 13, 4405. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Kobayashi, T.; Moroi, S.; Kotake, H.; Ikoma, T.; Saeki, H.; Ura, K.; Takagi, Y. Anti-obesity effects of chondroitin sulfate oligosaccharides from the skate Raja pulchra. Carbohydr. Polym. 2019, 214, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Ghareeb, A.; Fouda, A.; Kishk, R.M.; El Kazzaz, W.M. Unlocking the therapeutic potential of bioactive exopolysaccharide produced by marine actinobacterium Streptomyces vinaceusdrappus AMG31: A novel approach to drug development. Int. J. Biol. Macromol. 2024, 276, 133861. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Tian, Y.; Liu, S.; Chu, X.; Mao, W. Anti-Diabetic Activity of a Novel Exopolysaccharide Produced by the Mangrove Endophytic Fungus Penicillium janthinellum N29. Mar. Drugs 2023, 21, 270. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.W.; Wu, Y.; Ding, H. The range of dietary fibre ingredients and a comparison of their technical functionality. Fibre Rich Wholegrain Foods Improv. Qual. 2013, 237, 96–119. [Google Scholar] [CrossRef]
- Georg Jensen, M.; Knudsen, J.C.; Viereck, N.; Kristensen, M.; Astrup, A. Functionality of alginate based supplements for application in human appetite regulation. Food Chem. 2012, 132, 823–829. [Google Scholar] [CrossRef]
- Moraru, C.; Mincea, M.M.; Frandes, M.; Timar, B.; Ostafe, V. A Meta-Analysis on Randomised Controlled Clinical Trials Evaluating the Effect of the Dietary Supplement Chitosan on Weight Loss, Lipid Parameters and Blood Pressure. Medicina 2018, 54, 109. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wei, W.; Liu, F.; Wang, M.; Zhang, Y.; Du, S. Effects of fucoidan and synbiotics supplementation during bismuth quadruple therapy of Helicobacter pylori infection on gut microbial homeostasis: An open-label, randomized clinical trial. Front. Nutr. 2024, 11, 1407736. [Google Scholar] [CrossRef] [PubMed]
- Himaya, S.; Kim, S.K. Marine Nutraceuticals. In Springer Handbook of Marine Biotechnology; Kim, S.K., Ed.; Springer: Berlin, Germany, 2015; pp. 995–1014. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrasqueira, J.; Bernardino, S.; Bernardino, R.; Afonso, C. Marine-Derived Polysaccharides and Their Potential Health Benefits in Nutraceutical Applications. Mar. Drugs 2025, 23, 60. https://doi.org/10.3390/md23020060
Carrasqueira J, Bernardino S, Bernardino R, Afonso C. Marine-Derived Polysaccharides and Their Potential Health Benefits in Nutraceutical Applications. Marine Drugs. 2025; 23(2):60. https://doi.org/10.3390/md23020060
Chicago/Turabian StyleCarrasqueira, Joana, Susana Bernardino, Raul Bernardino, and Clélia Afonso. 2025. "Marine-Derived Polysaccharides and Their Potential Health Benefits in Nutraceutical Applications" Marine Drugs 23, no. 2: 60. https://doi.org/10.3390/md23020060
APA StyleCarrasqueira, J., Bernardino, S., Bernardino, R., & Afonso, C. (2025). Marine-Derived Polysaccharides and Their Potential Health Benefits in Nutraceutical Applications. Marine Drugs, 23(2), 60. https://doi.org/10.3390/md23020060