
Deoxyvasicinone with Anti-Melanogenic Activity from Marine-Derived Streptomyces sp. CNQ-617

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
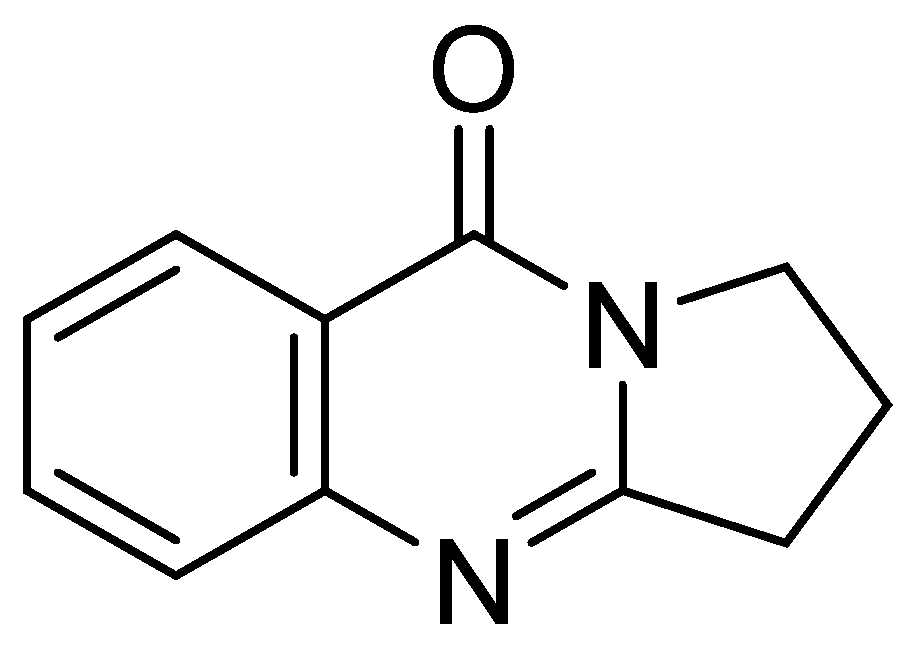
2.1. Identification of Deoxyvasicinone
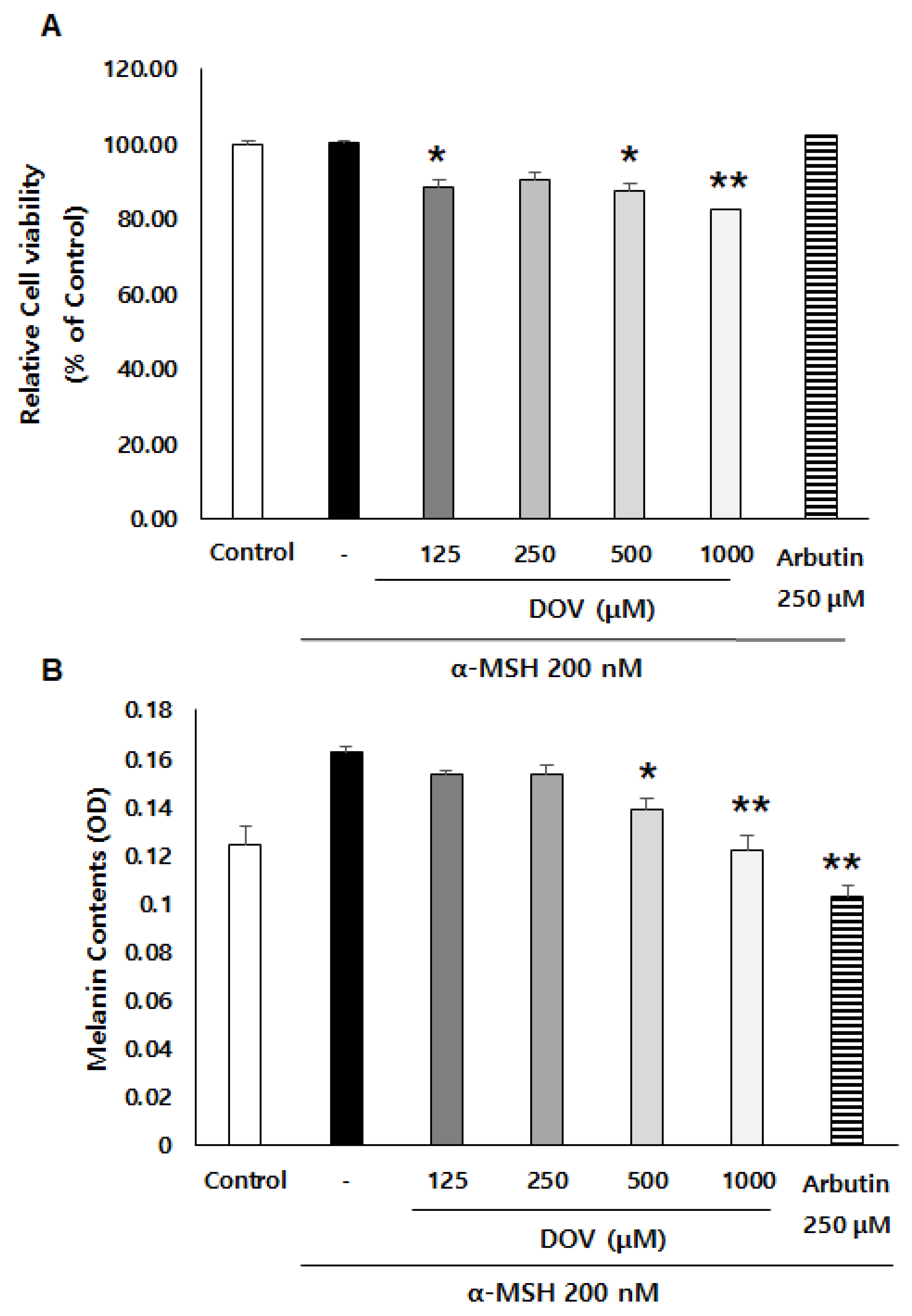
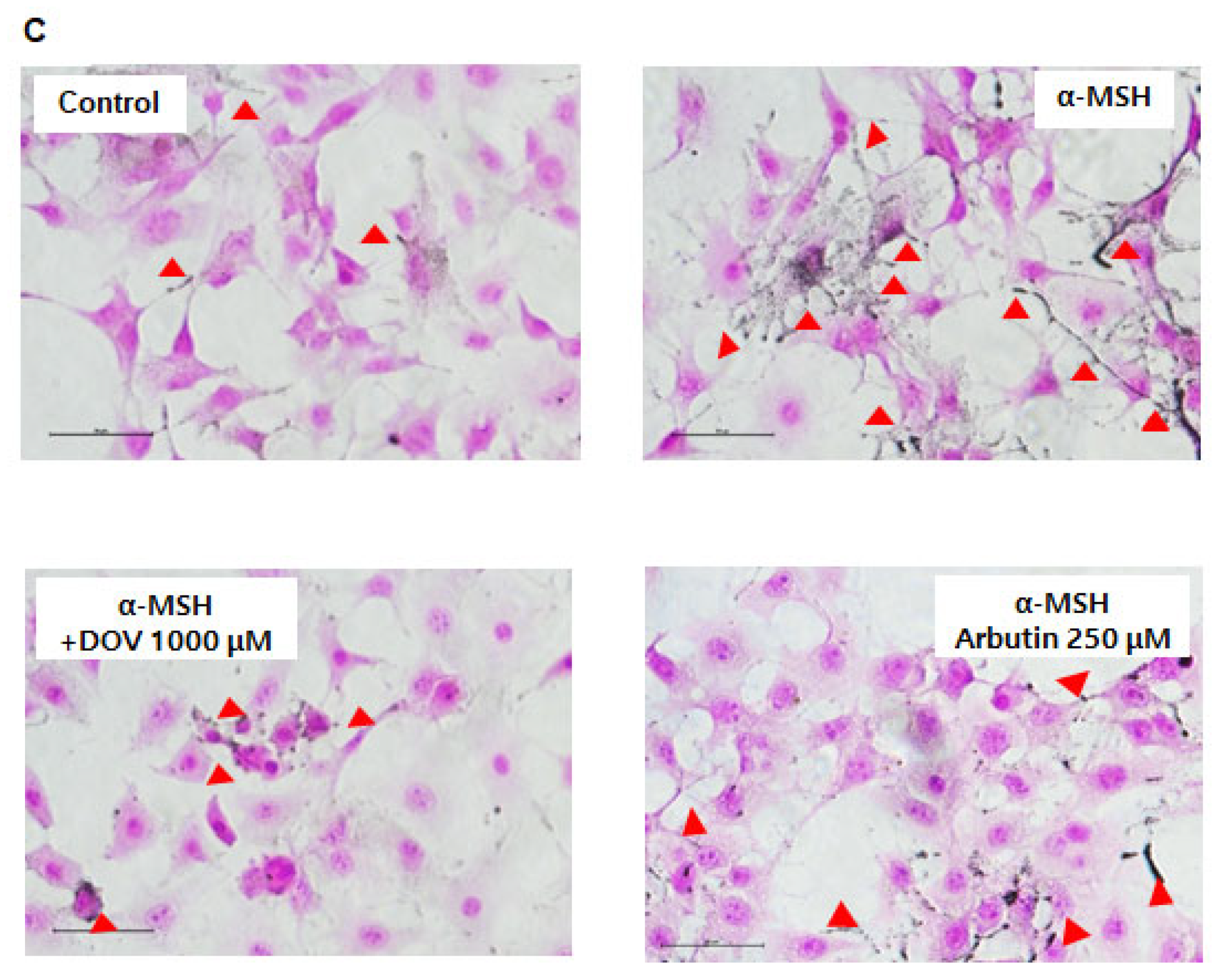
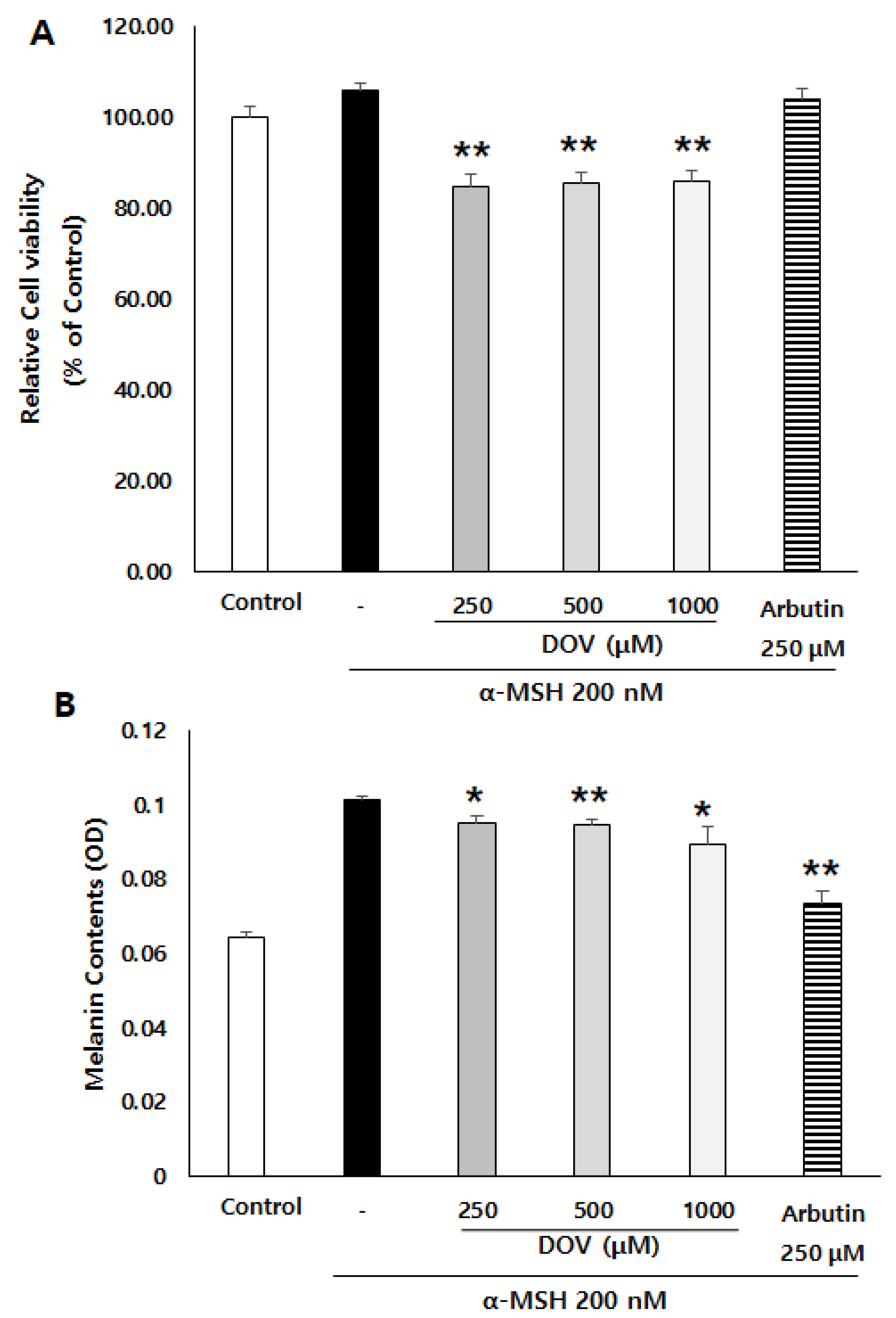
2.2. The Effect of Deoxyvasicinone on Melanogenesis and Viability of Rodent of B16F10 Cells
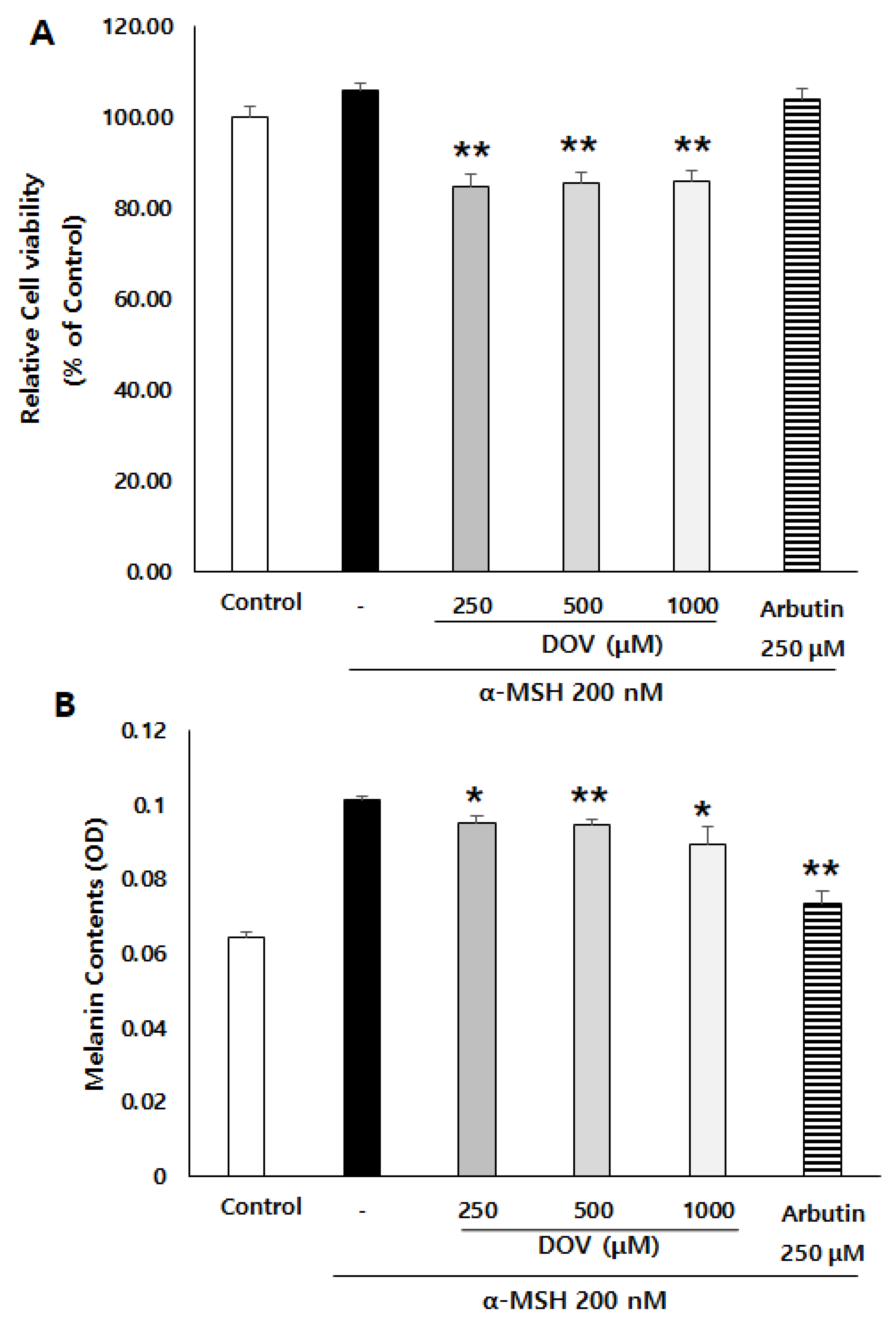
2.3. The Anti-Melanogenic Effects of Deoxyvasicinone upon MNT-1 Cells
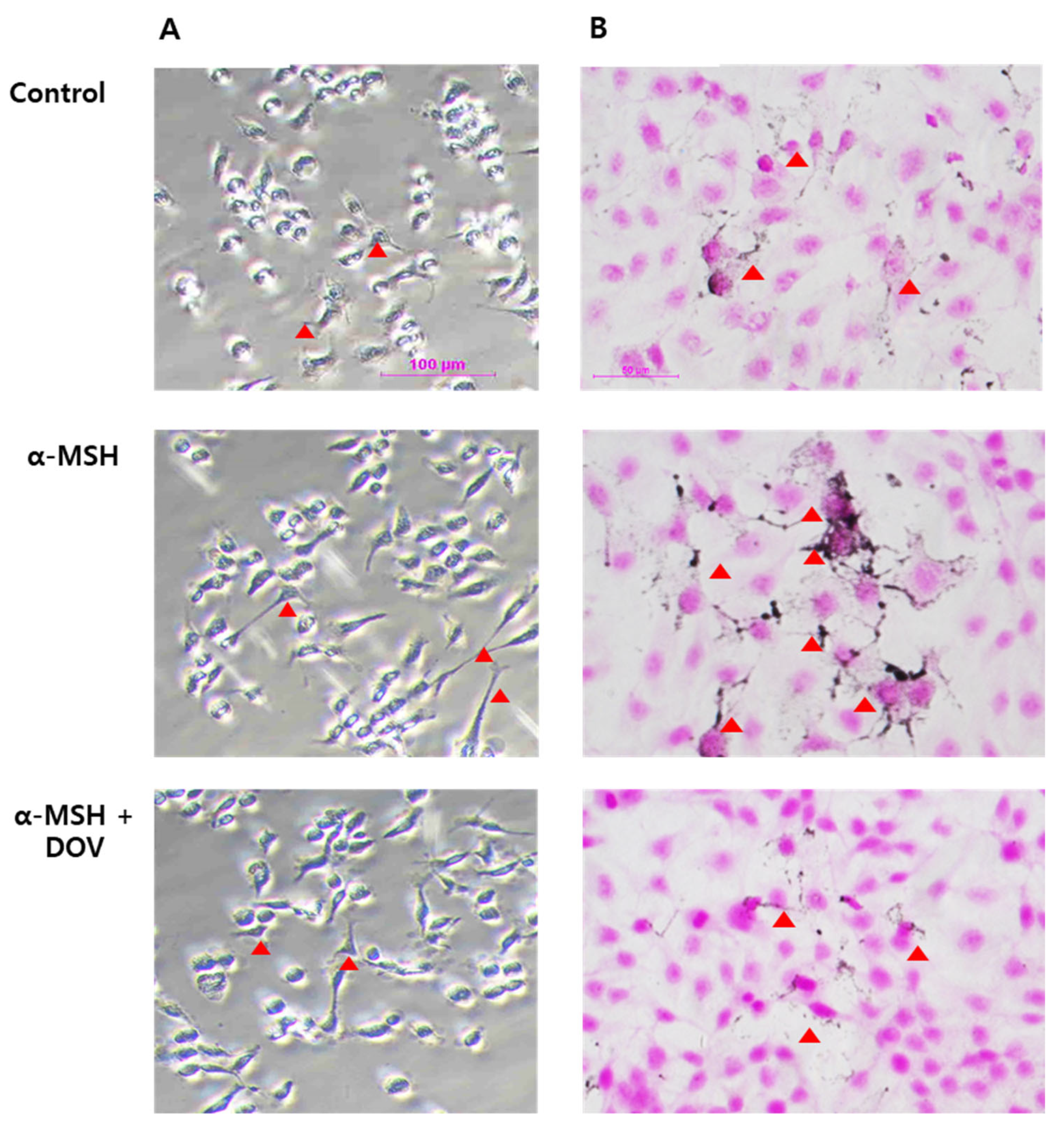
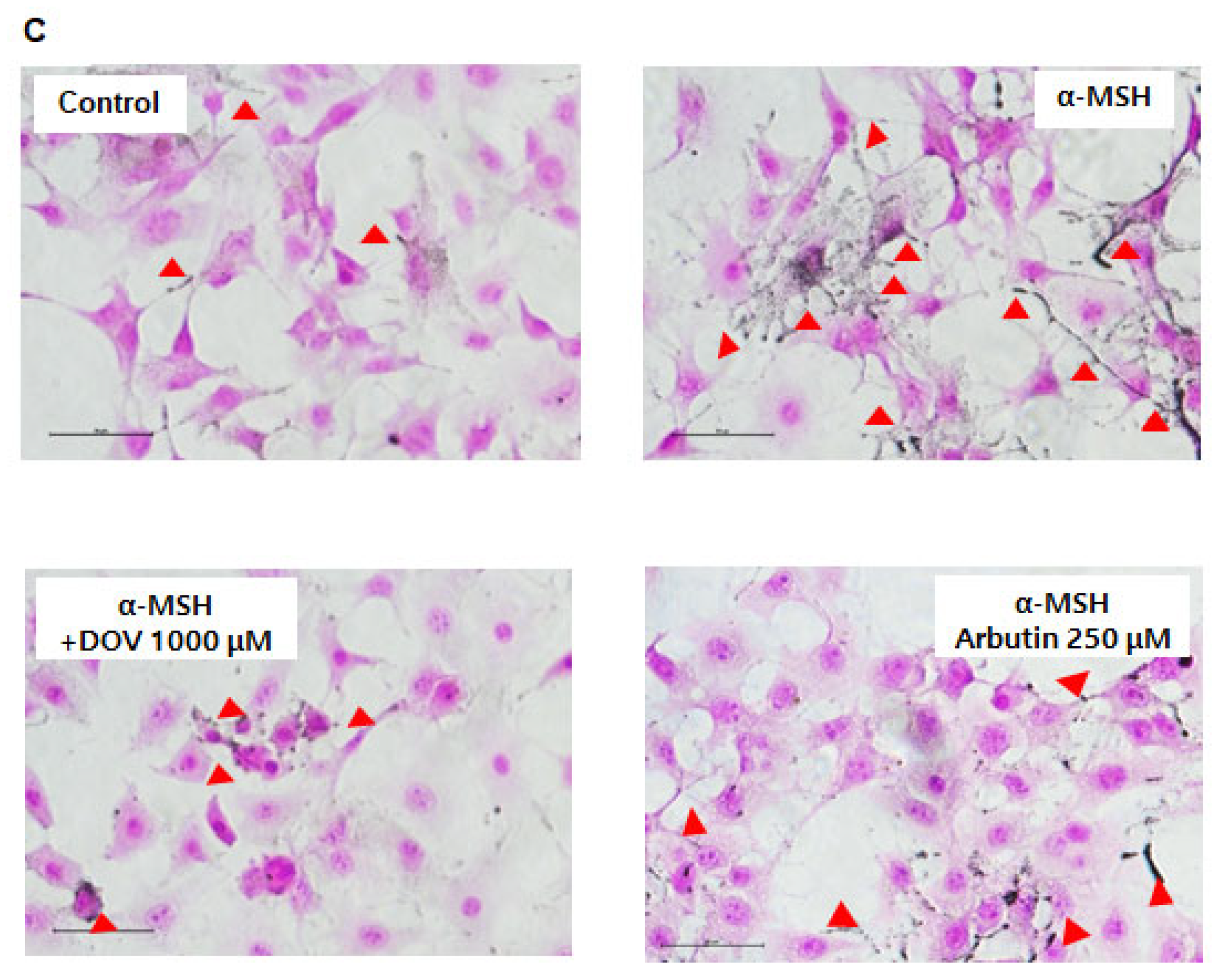
2.4. Effects of Deoxyvasicinone Treatment upon MNT-1 and HaCaT Co-Cultured Cells
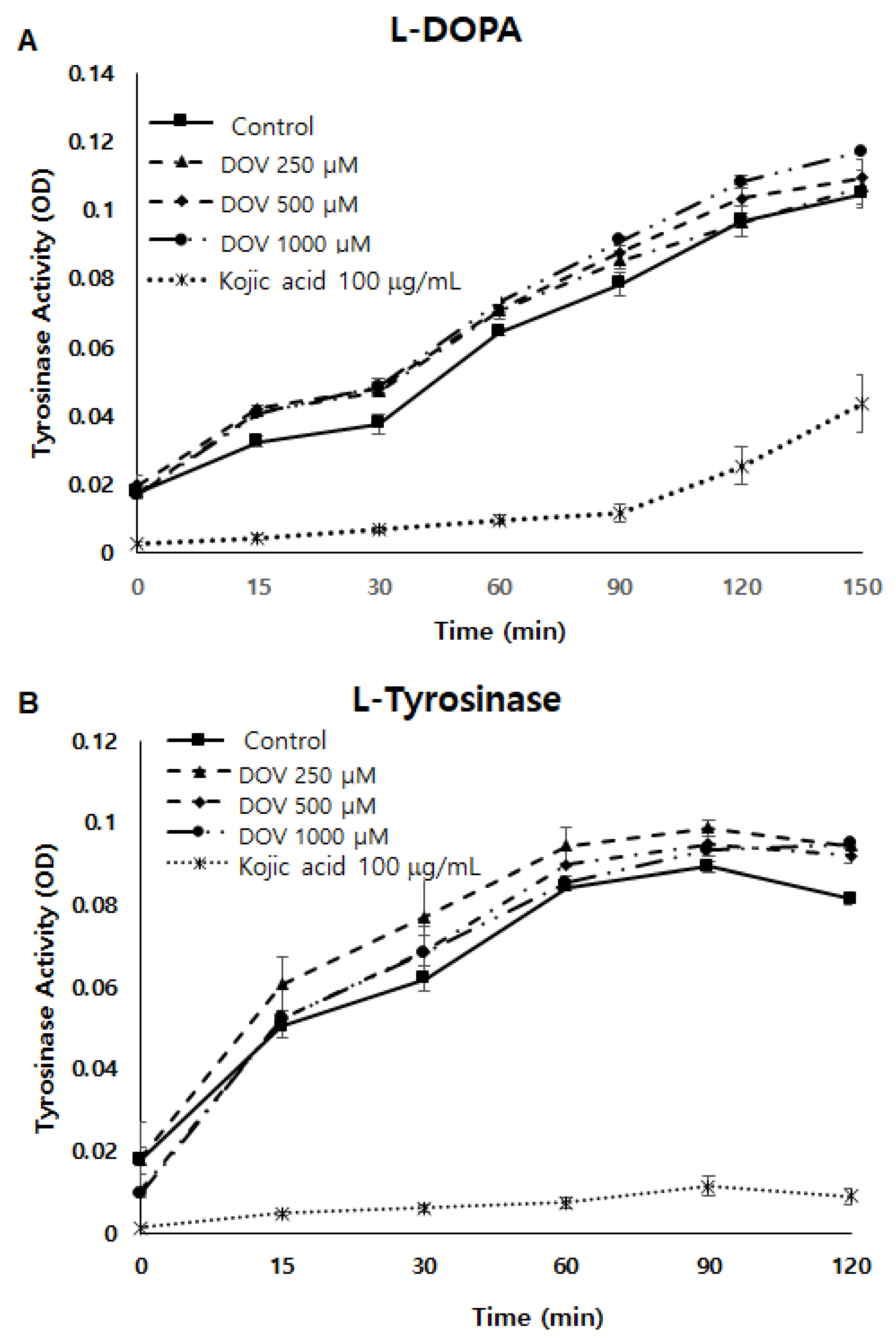
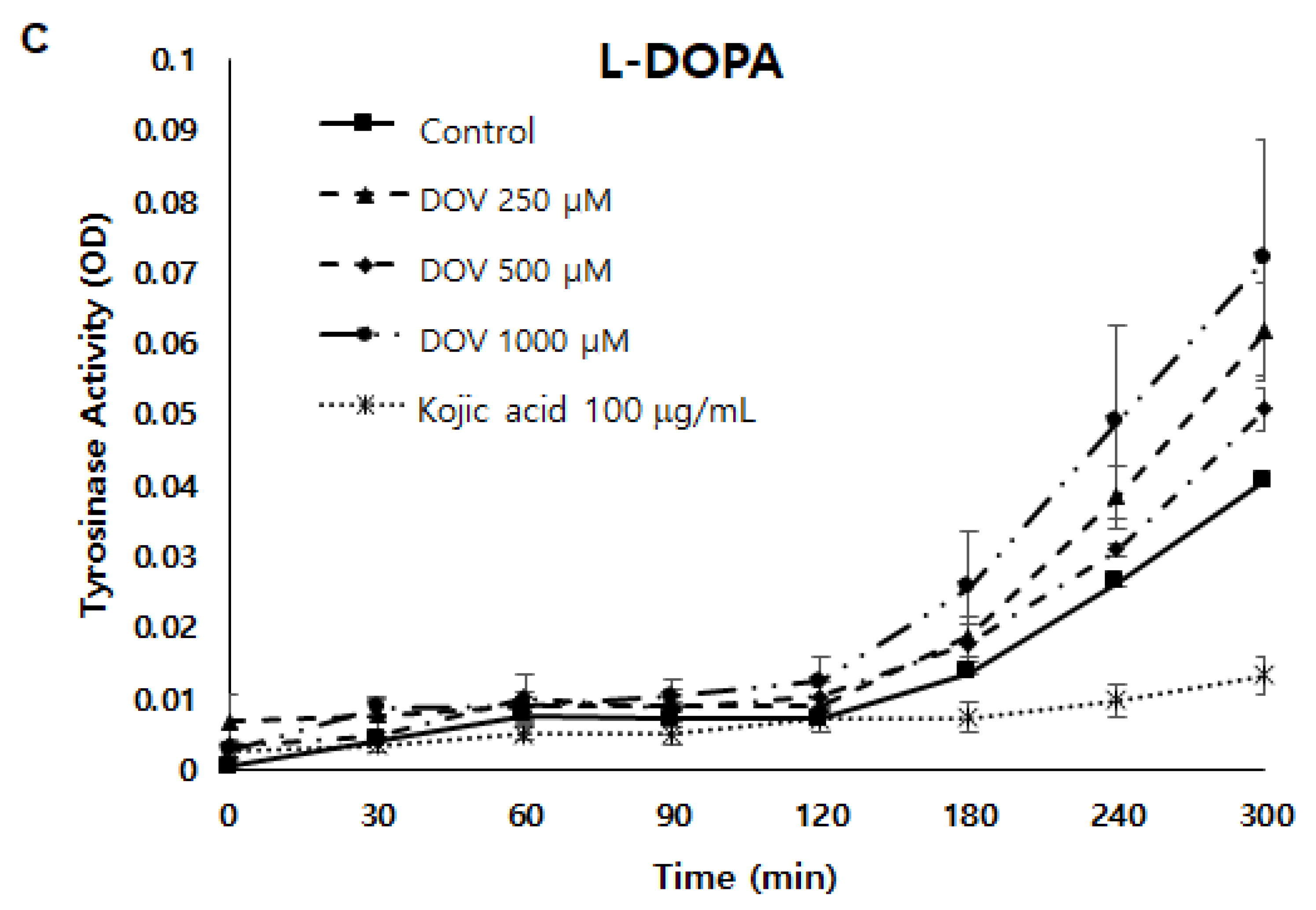
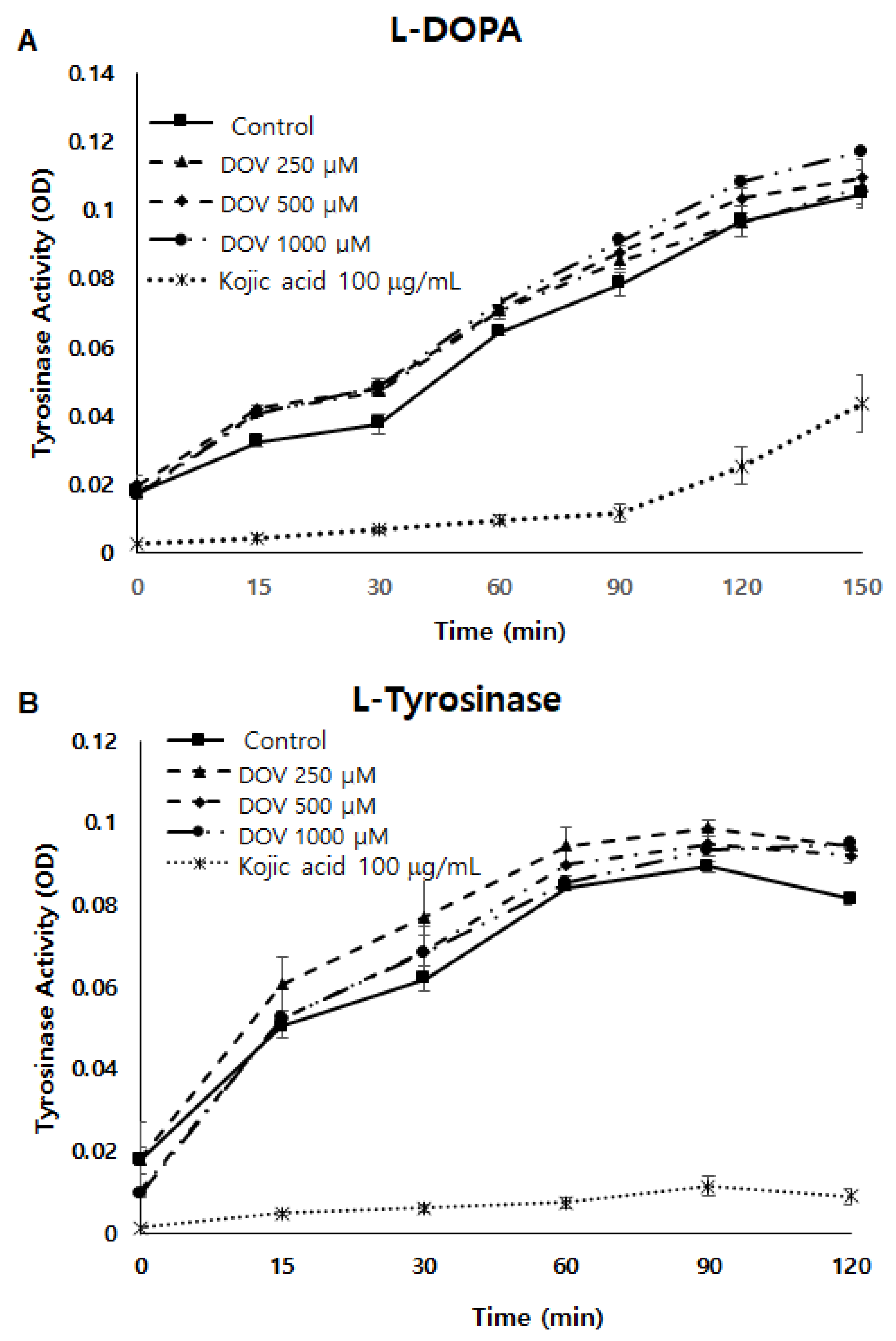
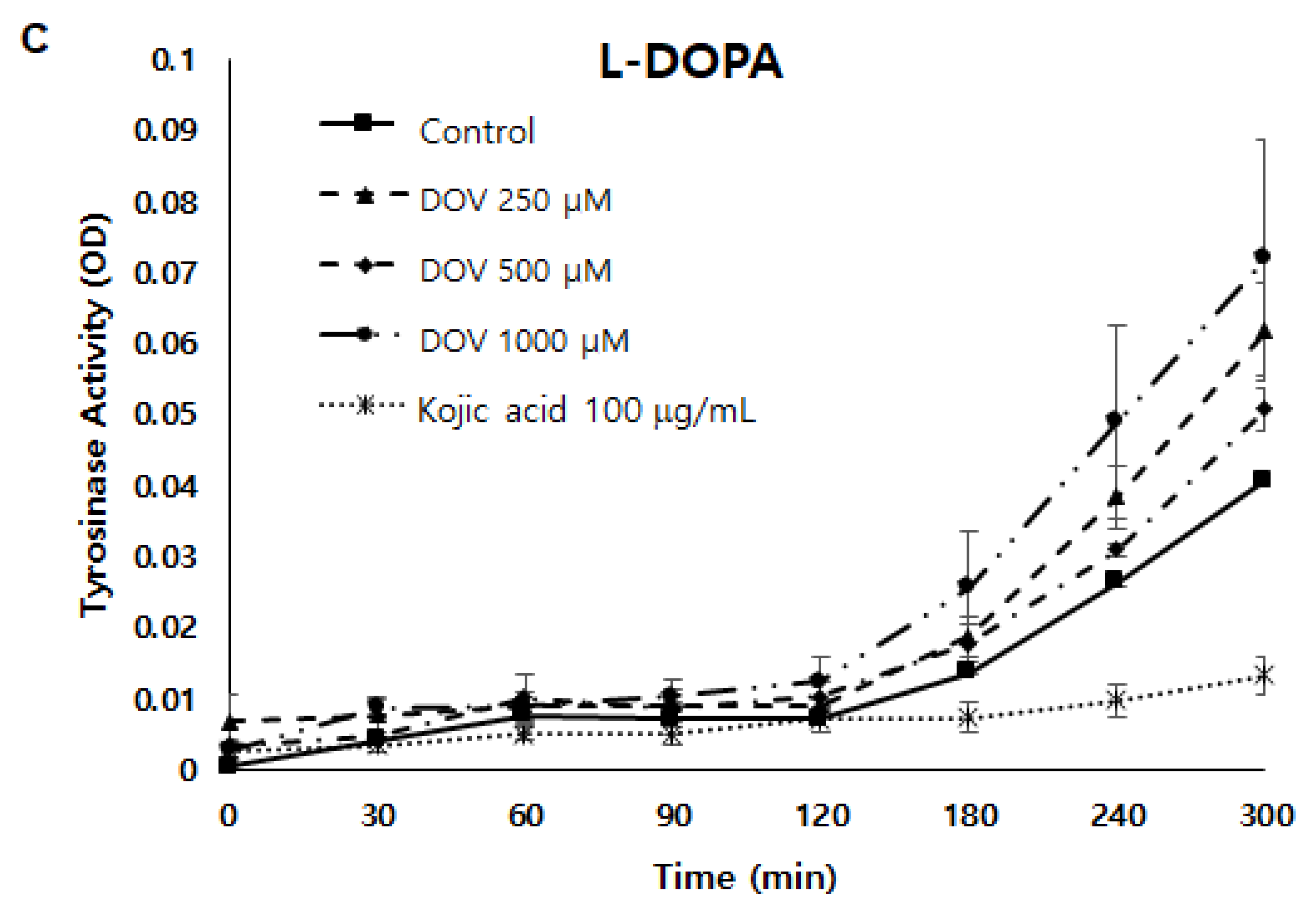
2.5. The Effects of Deoxyvasicinone upon Tyrosinase Enzymatic Activity
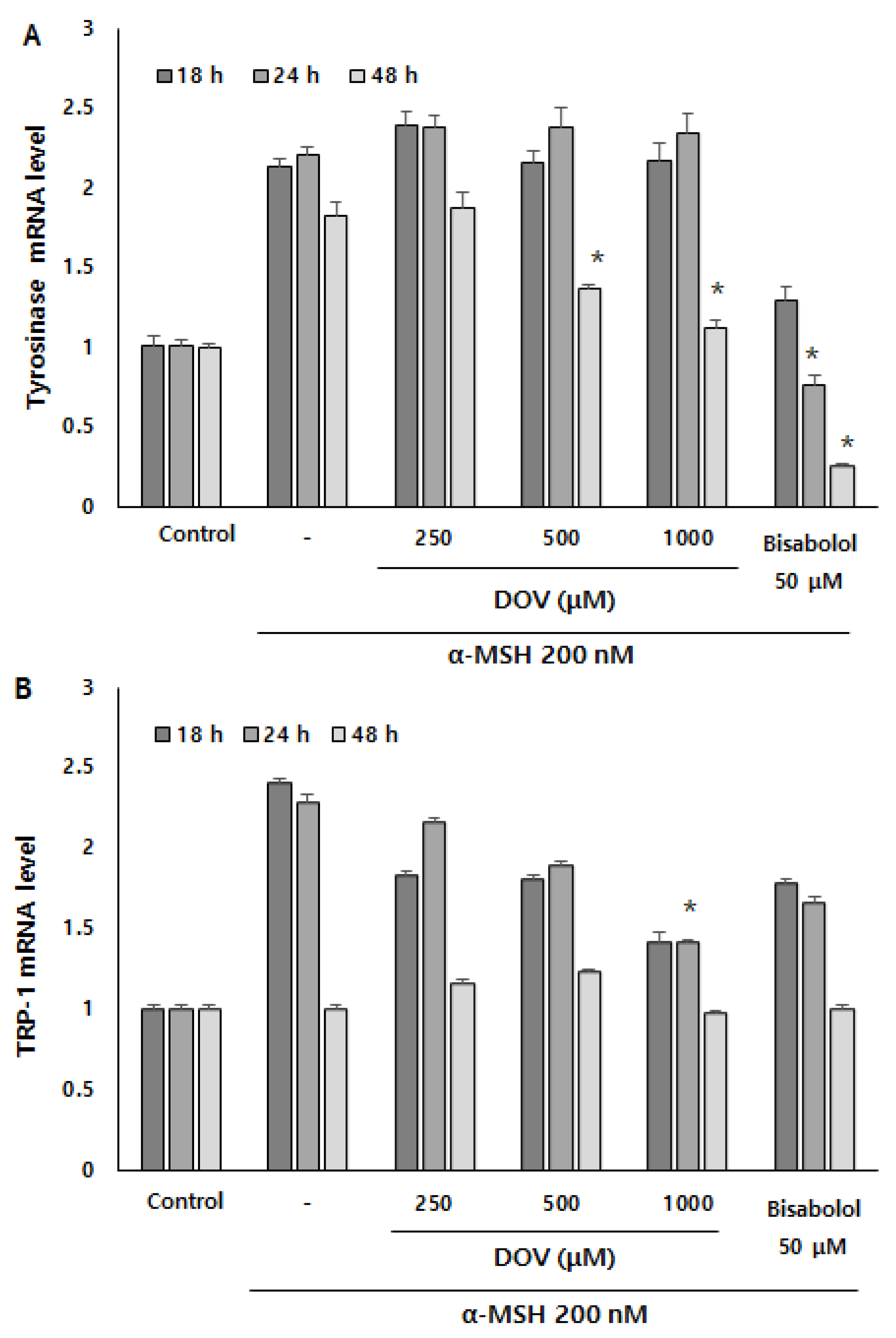
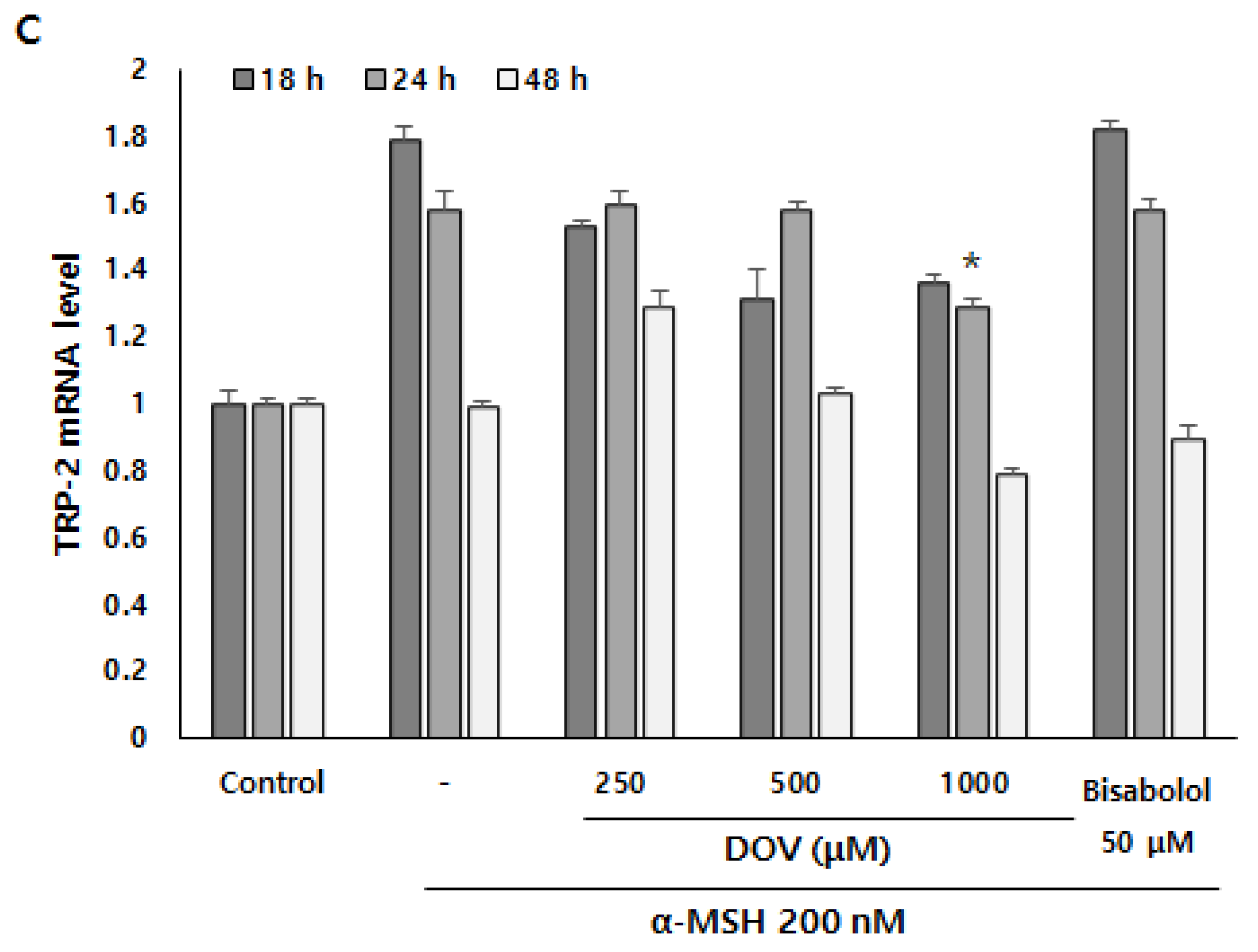
2.6. The Effects of Deoxyvasicinone on mRNA Expression of Melanogenic Enzymes
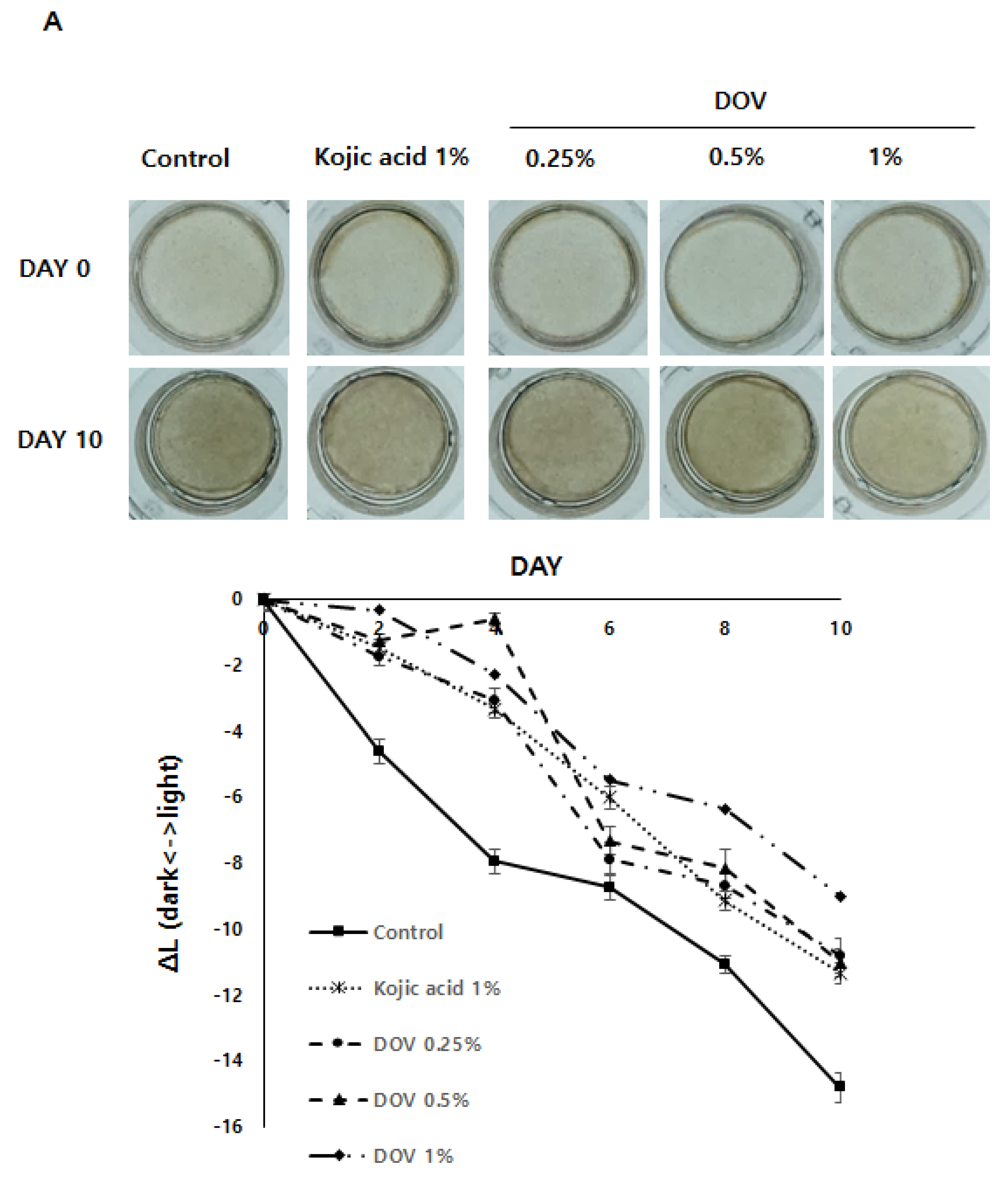
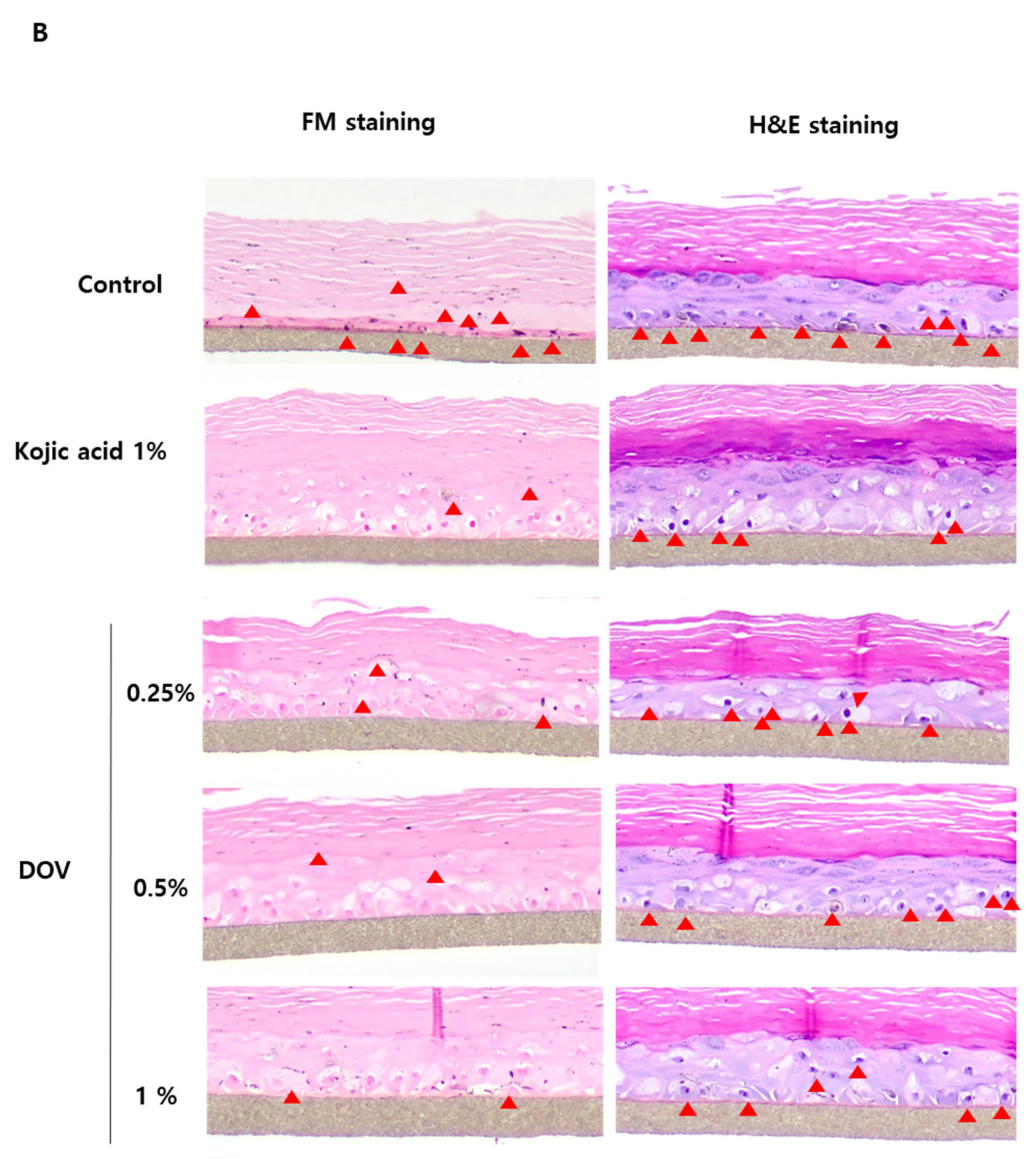
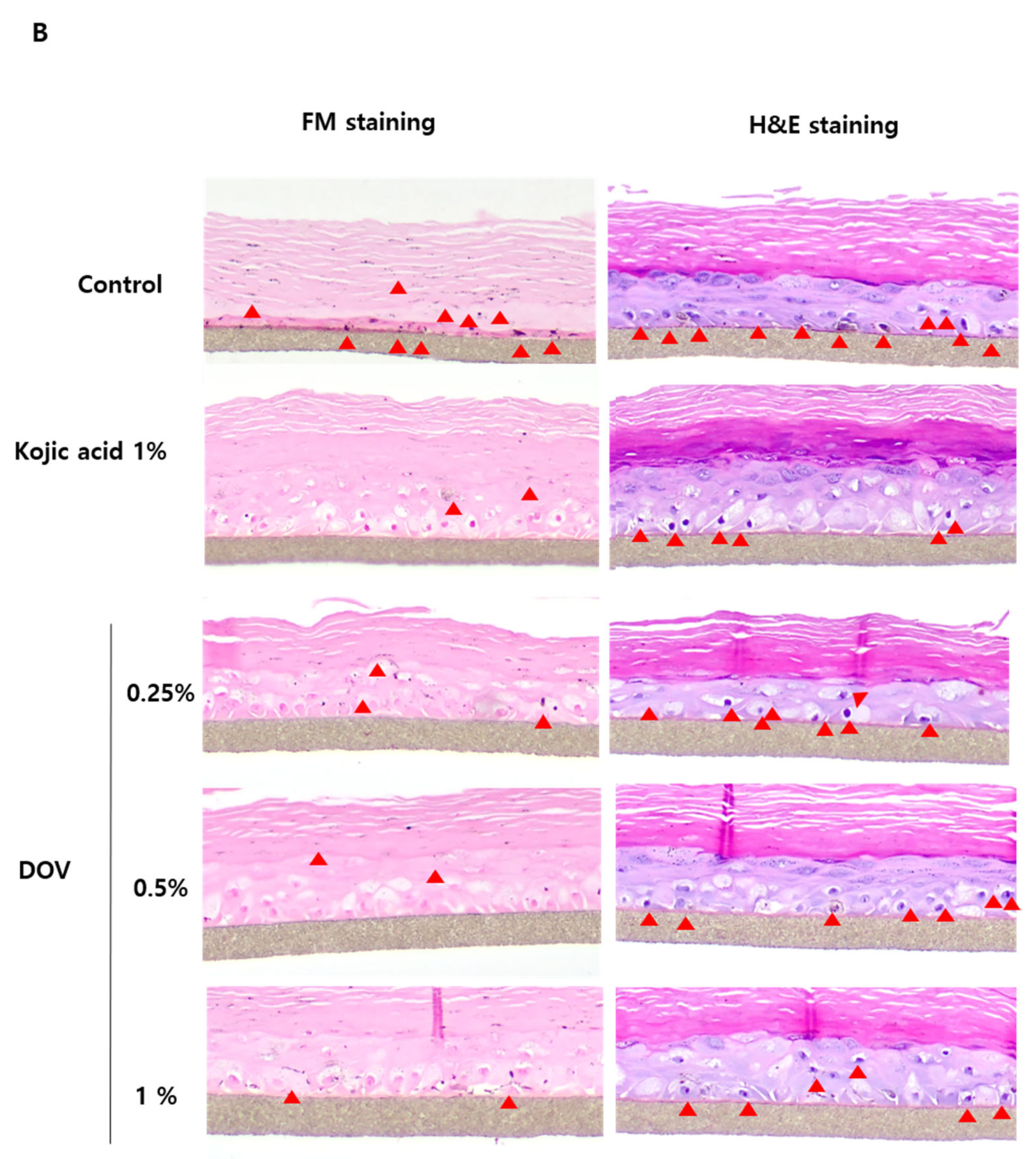
2.7. The Effects of Deoxyvasicinone on the Artificial Human Pigmented Epidermis Model MelanodermTM
3. Discussion
4. Materials and Methods
4.1. General Experimental
4.2. Bacterial Strain
4.3. Cultivation and Culture Extraction
4.4. Isolation
4.5. Cell Culture
4.6. Melanin Assay and Cell Viability Assay (MTT)
4.7. Mushroom Tyrosinase Inhibition Assay
4.8. RNA Isolation and Real-Time PCR
4.9. Skin Whitening Assay Using the Artificial Human Epidermal 3D Skin Model, MelanodermTM
4.10. The Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ito, S.; Wakamatsu, K. Quantitative analysis of eumelanin and pheomelanin in humans, mice, and other animals: A comparative review. Pigment. Cell Res. 2003, 16, 523–531. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaventure, J.; Domingues, M.J.; Larue, L. Cellular and molecular mechanisms controlling the migration of melanocytes and melanoma cells. Pigment. Cell Melanoma Res. 2013, 26, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Borovansky, J.; Riley, P.A. Melanins and Melanosomes: Biosynthesis, Structure, Physiological and Pathological Functions, 1st ed.; John Wiley & Sons: Weinheim, Germany, 2011; pp. 21–47. [Google Scholar]
- Gunia-Krzyżak, A.; Popiol, J.; Marona, H. Melanogenesis inhibitors: Strategies for searching for and evaluation of active compounds. Curr. Med. Chem. 2016, 23, 3548–3574. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.-Y.; Yin, W.-H.; Wang, M.-R.; Dang, Y.-Y.; Ye, X.-Y. Andrographolide suppresses melanin synthesis through Akt/GSK3β/β-catenin signal pathway. J. Dermatol. Sci. 2015, 79, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Del Marmol, V.; Beermann, F. Tyrosinase and related proteins in mammalian pigmentation. FEBS Lett. 1996, 381, 165–168. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Jung, S.-H. Recent development of signaling pathways inhibitors of melanogenesis. Cell. Signal. 2017, 40, 99–115. [Google Scholar] [CrossRef]
- Abbas, K.; Qadir, M.I.; Anwar, S. The role of melanin in skin cancer. Crit. Rev.™ Eukaryot. Gene Expr. 2019, 29, 17–24. [Google Scholar] [CrossRef]
- Fenical, W.; Jenson, P.R. Developing a new resource for drug discovery: Marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2, 666–673. [Google Scholar] [CrossRef]
- Kala, P.R.; Chandrika, V. Effect of different media for isolation, growth and maintenance of actinomycetes from mangrove sediments. Ind. J. Mar. Sci. 1993, 22, 297–299. [Google Scholar]
- Lam, K.S. Discovery of novel metabolites from marine actinomycetes. Curr. Opin. Microbiol. 2006, 9, 245–251. [Google Scholar] [CrossRef] [PubMed]
- De Lima, P.R.E.; da Silva, I.R.; Martins, M.K.; de Azevedo, J.L.; de Araújo, J.M. Antibiotics produced by Streptomyces. Braz. J. Infect. Dis. 2012, 16, 466–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegmann, E.; Mast, Y. Actinomycetes: The Antibiotics Producers. Antibiotics 2019, 8, 105–108. [Google Scholar]
- Potewar, T.M.; Ingale, S.A.; Srinivasan, K.V. Synthesis of tryptanthrin and deoxyvasicinone by a regioselective lithiation-intramolecular electrophilic reaction approach. Arkivoc 2008, 14, 100–108. [Google Scholar] [CrossRef]
- Kim, K.; Leutou, A.S.; Jeong, H.; Kim, D.; Seong, C.N.; Nam, S.-J.; Lim, K.-M. Anti-pigmentary effect of (-)-4-hydroxysattabacin from the marine-derived bacterium Bacillus sp. Mar. Drugs 2017, 15, 138. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Jeong, H.I.; Yang, I.; Nam, S.J.; Lim, K.M. Acremonidin E produced by Penicillium sp. SNF123, a fungal endophyte of Panax ginseng, has antimelanogenic activities. J. Ginseng Res. 2021, 45, 98–107. [Google Scholar] [CrossRef]
- Domingues, L.; Hurbain, I.; Gilles-Marsens, F.; Sirés-Campos, J.; André, N.; Dewulf, M.; Romao, M.; Lesegno, C.V.; Macé, A.-S.; Blouin, C.; et al. Coupling of melanocyte signaling and mechanics by caveolae is required for human skin pigmentation. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Breathnach, A. Melanin hyperpigmentation of skin: Melasma, topical treatment with azelaic acid, and other therapies. Cutis 1996, 57, 36–45. [Google Scholar]
- Statista Home Page. Available online: https://www.statista.com/statistics/863876/global-forecasted-market-value-of-skin-lightening-products/ (accessed on 12 February 2022).
- Jin, H.; Dan, H.-G.; Rao, G.-W. Research progress in quinazoline derivatives as multi-target tyrosine kinase inhibitors. Heterocycl. Commun. 2018, 24, 1–10. [Google Scholar] [CrossRef]
- Khodarahmi, G.; Jafari, E.; Hakimelahi, G.; Abedi, D.; Khajouei, M.R.; Hassanzadeh, F. Synthesis of some new quinazolinone derivatives and evaluation of their antimicrobial activities. IJPR 2012, 11, 789. [Google Scholar]
- Saravanan, G.; Alagarsamy, V.; Prakash, C.R. Synthesis and evaluation of antioxidant activities of novel quinazoline derivatives. Int. J. Pharm. Pharm. Sci. 2010, 2, 83–86. [Google Scholar]
- Sepehri, N.; Iraji, A.; Yavari, A.; Asgari, M.S.; Zamani, S.; Hosseini, S.; Bahadorikhalili, S.; Pirhadi, S.; Larijani, B.; Khoshneviszadeh, M.; et al. The natural-based optimization of kojic acid conjugated to different thio-quinazolinones as potential anti-melanogenesis agents with tyrosinase inhibitory activity. Bioorg. Med. Chem. 2021, 36, 116044. [Google Scholar] [CrossRef] [PubMed]
- Busca, R.; Balloti, R. Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigment Cell Res. 2000, 13, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Pavan, W.J. Transcriptional and signaling regulation in neural crest stem cell-derived melanocyte development: Do all roads lead to Mitf? Cell Res. 2008, 18, 1163–1176. [Google Scholar] [CrossRef]
- Myung, C.H.; Kim, K.; Park, J.I.; Lee, J.E.; Lee, J.A.; Hong, S.C.; Lim, K.M.; Hwang, J.S. 16-Kauren-2-beta-18,19-triol inhibits melanosome transport in melanocytes by down-regulation of melanophilin expression. J. Dermatol. Sci. 2020, 97, 101–108. [Google Scholar] [CrossRef]
- Lee, C.-S.; Nam, G.; Bae, I.-H.; Park, J. Whitening efficacy of ginsenoside F1 through inhibition of melanin transfer in cocultured human melanocytes–keratinocytes and three-dimensional human skin equivalent. J. Ginseng Res. 2019, 43, 300. [Google Scholar] [CrossRef]
- Kim, H.R.; Kim, H.; Jung, B.J.; You, G.E.; Jang, S.; Chung, D.K. Lipoteichoic acid isolated from Lactobacillus plantarum inhibits melanogenesis in B16F10 mouse melanoma cells. Mol. Cells. 2015, 38, 163–170. [Google Scholar]
- Liu, W.S.; Kuan, Y.D.; Chiu, K.H.; Wang, W.K.; Chang, F.H.; Liu, C.H.; Lee, C.H. The extract of Rhodobacter sphaeroides inhibits melanogenesis through the MEK/ERK signaling pathway. Mar. Drugs. 2013, 11, 1899–1908. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.O.; Jeong, H.; Cho, Y.; Lee, E.O.; Jang, H.W.; Kim, J.; Nam, K.; Lim, K.M. Lysates of a probiotic, Lactobacillus rhamnosus, can improve skin barrier function in a reconstructed human epidermis model. Int. J. Mol. Sci. 2019, 20, 4289. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Moon, A. A curcumin analog CA-5f inhibits urokinase-type plasminogen activator and invasive phenotype of triple-negative breast cancer cells. Toxicol. Res. 2021, 38, 19–26. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-e.; Kim, M.-j.; Hillman, P.F.; Oh, D.-C.; Fenical, W.; Nam, S.-J.; Lim, K.-M. Deoxyvasicinone with Anti-Melanogenic Activity from Marine-Derived Streptomyces sp. CNQ-617. Mar. Drugs 2022, 20, 155. https://doi.org/10.3390/md20020155
Lee S-e, Kim M-j, Hillman PF, Oh D-C, Fenical W, Nam S-J, Lim K-M. Deoxyvasicinone with Anti-Melanogenic Activity from Marine-Derived Streptomyces sp. CNQ-617. Marine Drugs. 2022; 20(2):155. https://doi.org/10.3390/md20020155
Chicago/Turabian StyleLee, Se-eun, Min-ju Kim, Prima F. Hillman, Dong-Chan Oh, William Fenical, Sang-Jip Nam, and Kyung-Min Lim. 2022. "Deoxyvasicinone with Anti-Melanogenic Activity from Marine-Derived Streptomyces sp. CNQ-617" Marine Drugs 20, no. 2: 155. https://doi.org/10.3390/md20020155
APA StyleLee, S.-e., Kim, M.-j., Hillman, P. F., Oh, D.-C., Fenical, W., Nam, S.-J., & Lim, K.-M. (2022). Deoxyvasicinone with Anti-Melanogenic Activity from Marine-Derived Streptomyces sp. CNQ-617. Marine Drugs, 20(2), 155. https://doi.org/10.3390/md20020155