
Inhibition of SARS-CoV-2 Virus Entry by the Crude Polysaccharides of Seaweeds and Abalone Viscera In Vitro

 ,
,
Abstract
1. Introduction
2. Results
2.1. Composition Analysis of Crude Polysaccharides
2.2. Monosaccharide Composition of Crude Polysaccharides
2.3. Trace Elements in the Crude Polysaccharides
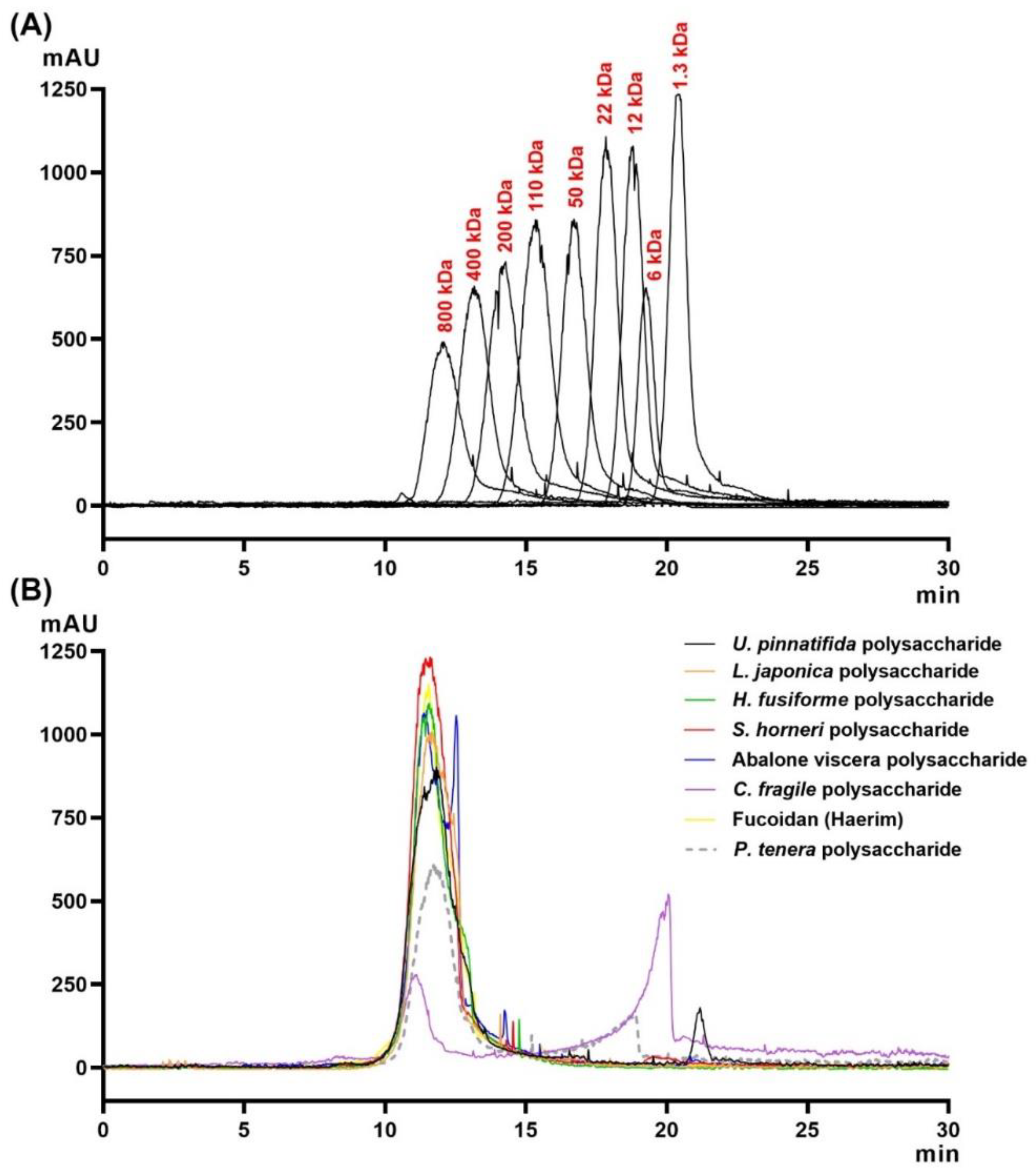
2.4. Molecular Weight Distribution of Crude Polysaccharides
2.5. Cytotoxicity Assay
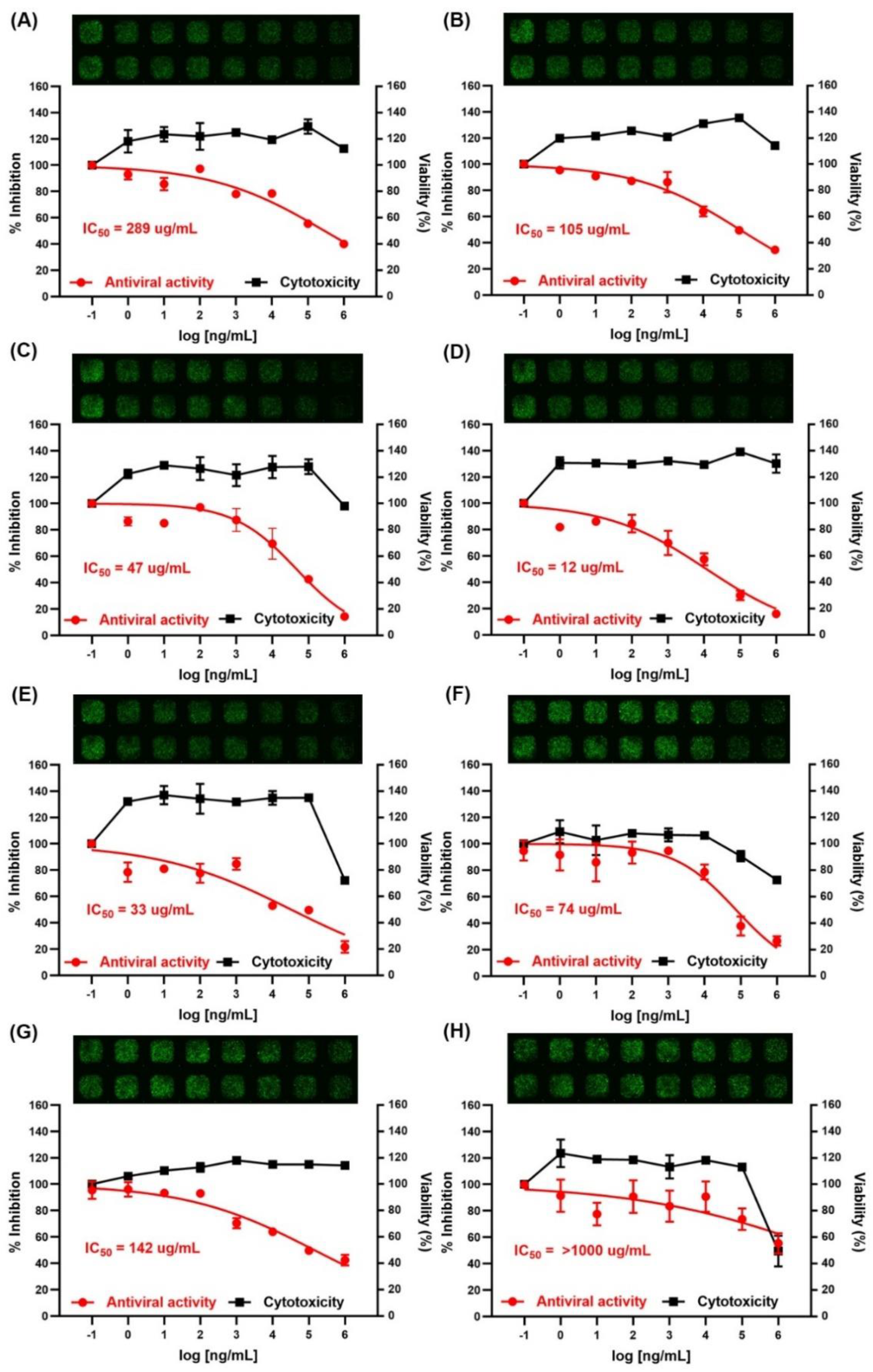
2.6. Inhibition of Viral Infection
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Seaweed Collection
4.3. Preparation of the Crude Polysaccharides
4.4. Total Carbohydrate Content
4.5. Determination of Sulfate Contents
4.6. Total Phenolic Content Analysis
4.7. BCA Assay
4.8. Monosaccharide Analysis of Crude Polysaccharides
4.9. Analysis of Minerals and Trace Elements
4.10. Estimation of Average Molecular Weights
4.11. SARS-CoV-2 Pseudovirus and Cell
4.12. Cytotoxicity Assay
4.13. Inhibition of Viral Infection
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- COVID-19 Case Tracker. Available online: https://coronavirus.jhu.edu/#covid-19-basics2020 (accessed on 15 March 2021).
- U.S. Food and Drug Administration. COVID-19 Vaccines. Available online: https://www.fda.gov/emergency-preparedness-and-response/coronavirus-disease-2019-covid-19/covid-19-vaccines#news (accessed on 15 March 2021).
- COVID-19 Vaccine Tracker. Available online: https://www.bloomberg.com/graphics/covid-vaccine-tracker-global-distribution/ (accessed on 15 March 2021).
- Coronavirus (COVID-19) Vaccinations Tracker. Available online: https://ourworldindata.org/covid-vaccinations (accessed on 15 March 2021).
- Brown, E.S.; Allsopp, P.J.; Magee, P.J.; Gill, C.I.R.; Nitecki, S.; Strain, C.R.; McSorley, E.M. Seaweed and human health. Nutr. Rev. 2014, 72, 205–216. [Google Scholar] [CrossRef] [PubMed]
- MacArtain, P.; Gill, C.I.R.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65 Pt 1, 535–543. [Google Scholar] [CrossRef]
- Hans, N.; Malik, A.; Naik, S. Antiviral activity of sulfated polysaccharides from marine algae and its application in combating COVID-19: Mini review. Bioresour. Technol. Rep. 2021, 13, 100623. [Google Scholar] [CrossRef] [PubMed]
- Kwon, P.S.; Oh, H.; Kwon, S.-J.; Jin, W.; Zhang, F.; Fraser, K.; Hong, J.J.; Linhardt, R.J.; Dordick, J.S. Sulfated polysaccharides effectively inhibit SARS-CoV-2 in vitro. Cell Discov. 2020, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Luthull, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic effects of fucoidan: A review on recent studies. Mar. Drugs 2019, 17, 487. [Google Scholar] [CrossRef] [PubMed]
- Fitton, J.H.; Park, A.Y.; Karpiniec, S.S.; Stringer, D.N. Fucoidan and lung function: Value in viral infection. Mar. Drugs 2021, 19, 4. [Google Scholar] [CrossRef]
- Tamama, K. Potential benefits of dietary seaweeds as protection against COVID-19. Nutr. Rev. 2020, nuaa126. [Google Scholar] [CrossRef]
- Kandeel, M.; Al-Nazawi, M. Virtual screening and repurposing of FDA approved drugs against COVID-19 main protease. Life Sci. 2020, 251, 117627. [Google Scholar] [CrossRef] [PubMed]
- Qamar, M.T.U.; Alqahtani, S.M.; Alamri, M.A.; Chen, L.L. Structural basis of SARS-CoV-2 3CLpro and anti-COVID-19 drug discovery from medicinal plants. J. Pharm. Anal. 2020, 10, 313–319. [Google Scholar] [CrossRef]
- Guo, S.; Wang, J.; He, C.; Wei, H.; Ma, Y.; Xiong, H. Preparation and antioxidant activities of polysaccharides obtained from abalone viscera by combination of enzymolysis and multiple separation methods. J. Food Sci. 2020, 85, 4260–4270. [Google Scholar] [CrossRef]
- Alcantara, L.; Noro, T. Growth of the abalone Haliotis diversicolor (Reeve) fed with macroalgae in floating net cage and plastic tank. Aquac. Res. 2006, 37, 708–717. [Google Scholar] [CrossRef]
- Zheng, P.; Hao, G.; Weng, W.; Ren, H. Antioxidant activities of hydrolysates from abalone viscera using subcritical water-assisted enzymatic hydrolysis. Food Bioproc. Tech. 2019, 12, 910–918. [Google Scholar] [CrossRef]
- Mai, K.; Mercer, J.P.; Donlon, J. Comparative studies on the nutrition of two species of abalone, Haliotis tuberculata L. and Haliotis discus hannai Ino. V. The role of polyunsaturated fatty acids of macroalgae in abalone nutrition. Aquaculture 1996, 139, 77–89. [Google Scholar] [CrossRef]
- Naidoo, K.; Maneveldt, G.; Ruck, K.; Bolton, J.J. A comparison of various seaweed based diets and formulated feed on growth rate of abalone in a land-based aquaculture system. J. Appl. Phycol. 2006, 18, 437–443. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Masci, P.P.; Gobe, G.C.; Osborne, S.A. Therapeutic potential of abalone and status of bioactive molecules: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1742–1748. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Dodgson, K.S.; Price, R.G. A note on the determination of the ester sulfate content of sulfated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Daly, J.L.; Simonetti, B.; Klein, K.; Chen, K.-E.; Williamson, M.K.; Antón-Plágaro, C.; Shoemark, D.K.; Simón-Gracia, L.; Bauer, M.; Hollandi, R.; et al. Neuropilin-1 is a host factor for SARS-CoV-2 infection. Science 2020, 370, 861–865. [Google Scholar] [CrossRef]
- Jin, W.; Zhang, W.; Mitra, D.; McCandless, M.G.; Sharma, P.; Tandon, R.; Zhang, E.; Linhardt, R.J. The structure-activity relationship of the interactions of SARS-CoV-2 spike glycoproteins with glucuronomannan and sulfated galactofucan from Saccharina japonica. Int. J. Biol. Macromol. 2020, 163, 1649–1658. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Peng, H.; Wang, Q.; Liu, Z.; Dong, X.; Wen, C.; Ai, C.; Zhang, Y.; Wang, Z.; Zhu, B. Inhibitory activities of marine sulfated polysaccharides against SARS-CoV-2. Food Funct. 2020, 11, 7415–7420. [Google Scholar] [CrossRef]
- Hao, G.; Cao, W.; Li, T.; Chen, J.; Zhang, J.; Weng, W.; Osako, K.; Ren, H. Effect of temperature on chemical properties and antioxidant activities of abalone viscera subcritical water extract. J. Supercrit. Fluids 2019, 147, 17–23. [Google Scholar] [CrossRef]
- Sun, L.; Zhu, B.; Li, D.; Wang, L.; Dong, X.; Murata, Y.; Xing, R.; Dong, Y. Purification and bioactivity of a sulphated polysaccharide conjugate ACP I from viscera of abalone Haliotis Discus Hannai Ino. Food Agric. Immunol. 2010, 21, 15–26. [Google Scholar] [CrossRef]
- Zhang, R.; Shi, Y.; Zheng, J.; Mao, X.; Liu, Z.; Chen, Q.; Wang, Q. Effects of polysaccharides from abalone viscera (Haliotis discus hannai Ino) on MGC 803 cells proliferation. Int. J. Biol. Macromol. 2018, 106, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Vishchuk, O.S.; Ermakova, S.P.; Zvyagintseva, T.N. Sulfated polysaccharides from brown seaweeds Saccharina japonica and Undaria pinnatifida: Isolation, structural characteristics, and antitumor activity. Carbohydr. Res. 2011, 346, 2769–2776. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Ren, L.; Liu, B.; Zhang, Q.; Zhong, W. Structural features of sulfated glucuronomannan oligosaccharides and their antioxidant activity. Mar. Drugs 2018, 16, 291. [Google Scholar] [CrossRef]
- Ciancia, M.; Quintana, I.; Vizcargüénaga, M.I.; Kasulin, L.; de Dios, A.; Estevez, J.M.; Cerezo, A.S. Polysaccharides from the green seaweeds Codium fragile and C. vermilara with controversial effects on hemostasis. Int. J. Biol. Macromol. 2007, 41, 641–649. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.; Hamilton, J.; Rebers, P.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Fu, D.T.; Oneill, R.A. Monosaccharide composition analysis of oligosaccharides and glycoproteins by high-performance liquid chromatography. Anal. Biochem. 1995, 227, 377–384. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Samples | Total (% of Seaweed Water Extract) | |||
|---|---|---|---|---|
| Carbohydrate | Protein | Sulfate Ion | Phenolic Content | |
| Undaria pinnatifida (sporophyll) | 60.3 ± 1.97 | 2.6 ± 0.50 | 36.3 ± 1.4 | 0.15 ± 0.021 |
| Laminaria japonica | 54.6 ± 1.46 | 8.7 ± 1.12 | * n.d. | 0.14 ± 0.070 |
| Hizikia fusiforme | 94.4 ± 1.82 | 10.9 ± 0.43 | 20.4 ± 0.4 | 1.09 ± 0.030 |
| Sargassum horneri | 99.1 ± 3.35 | 4.0 ± 1.14 | 9.8 ± 0.5 | 1.03 ± 0.012 |
| Abalone viscera | 62.7 ± 4.58 | 22.3 ± 1.17 | 0.5 ± 0.1 | 0.43 ± 0.002 |
| Codium fragile | 29.0 ± 1.80 | 1.4 ± 0.44 | 16.8 ± 1.8 | n.d. |
| Fucoidan | 58.6 ± 2.78 | 0.5 ± 0.31 | 30.7 ± 0.8 | 0.04 ± 0.007 |
| Porphyra tenera | 46.0 ± 1.90 | n.d. | n.d. | 0.12 ± 0.026 |
| Monosaccharides (%) | Undaria pinnatifida (Sporophyll) | Laminaria japonica | Hizikia fusiforme | Sargassum horneri | Abalone viscera | Codium fragile | Fucoidan | Porphyra tenera |
|---|---|---|---|---|---|---|---|---|
| Mannose | 5.7 | 15.3 | 16.6 | 7.6 | 8.7 | 4.5 | 4.3 | 4.6 |
| Rhamnose | 5.9 | 8.5 | * n.d. | n.d. | 8.2 | 5.9 | 6.5 | 6.0 |
| Glucose | 2.9 | 3.4 | 3.1 | 7.3 | 3.8 | 3.6 | n.d. | 2.8 |
| Galactose | 30.7 | 10.2 | 9.6 | 6.9 | 12.9 | 6.6 | 7.9 | 32.4 |
| Xylose | 3.5 | 4.1 | 3.7 | 4.5 | 3.7 | n.d. | * n.d. | 3.6 |
| Arabinose | n.d. | n.d. | n.d. | n.d. | n.d. | 5.9 | 6.5 | n.d. |
| Fucose | 50.3 | 22.1 | 41.8 | 66.2 | 37.3 | 2.6 | 12.8 | 4.5 |
| Mineral (μg g−1 of Sample) | Undaria pinnatifida (Sporophyll) | Laminaria japonica | Hizikia fusiforme | Sargassum horneri | Abalone viscera | Codium fragile | Fucoidan | Porphyra tenera |
|---|---|---|---|---|---|---|---|---|
| Ag | 282.1 | 926.3 | 137.2 | 17.3 | 15.2 | 4.8 | * n.d. | n.d. |
| Al | 3.5 | 35.1 | 28.7 | 22.6 | 6.2 | 74.9 | 362.5 | 500.5 |
| As | 33.1 | 18.7 | 15.4 | 1.9 | 29.3 | n.d. | n.d. | n.d. |
| B | 1647.1 | 1098.6 | 215.9 | 122.9 | 65.8 | 59.8 | 89.9 | 23,945.8 |
| Ba | 1.8 | 2.1 | 1.6 | 1.8 | 0.8 | 4.1 | 97.4 | 115.1 |
| Bi | 405.7 | 740.1 | 577.9 | 353.2 | 289.0 | 470.9 | n.d. | n.d. |
| Ca | 95,735.5 | 136,540.2 | 80,633.2 | 107,498.5 | 68,895.3 | 282,637.1 | 82,928.7 | 62,308.7 |
| Cd | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Co | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 274.3 | 118.7 |
| Cr | n.d. | n.d. | 1.0 | n.d. | 0.1 | 1.4 | 23.1 | 273.4 |
| Cu | 15.6 | 44.0 | 25.5 | 12.8 | 14.4 | 13.9 | 244.1 | n.d. |
| Fe | 25.1 | 69.8 | 59.6 | 103.5 | 583.1 | 132.2 | 216.2 | 1362.2 |
| Ga | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| In | 46.0 | n.d. | n.d. | n.d. | n.d. | n.d. | 1044.3 | 2183.4 |
| K | 45,896.1 | 876.0 | 17,531.7 | 9907.2 | 6368.1 | 5456.3 | 9694.1 | 91,790.1 |
| Li | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Mg | 9143.9 | 17,807.9 | 14,562.4 | 15,518.6 | 5590.2 | 6123.8 | 4221.2 | 15,042.7 |
| Mn | 1.1 | 2.2 | 15.0 | 9.7 | 1.8 | 343.2 | 57.0 | 157.4 |
| Na | 8134.2 | 3888.1 | 1068.2 | 424.7 | 3597.4 | 94,248.6 | 7232.6 | 27,630.7 |
| Ni | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 39.4 |
| Pb | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Sr | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Ti | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 19386.2 |
| Zn | 6.1 | 59.3 | 51.0 | 23.7 | 26.9 | 16.3 | 149.6 | 258.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yim, S.-K.; Kim, K.; Kim, I.; Chun, S.; Oh, T.; Kim, J.-U.; Kim, J.; Jung, W.; Moon, H.; Ku, B.; et al. Inhibition of SARS-CoV-2 Virus Entry by the Crude Polysaccharides of Seaweeds and Abalone Viscera In Vitro. Mar. Drugs 2021, 19, 219. https://doi.org/10.3390/md19040219
Yim S-K, Kim K, Kim I, Chun S, Oh T, Kim J-U, Kim J, Jung W, Moon H, Ku B, et al. Inhibition of SARS-CoV-2 Virus Entry by the Crude Polysaccharides of Seaweeds and Abalone Viscera In Vitro. Marine Drugs. 2021; 19(4):219. https://doi.org/10.3390/md19040219
Chicago/Turabian StyleYim, Sung-Kun, Kian Kim, Inhee Kim, SangHo Chun, TaeHwan Oh, Jin-Ung Kim, Jungwon Kim, WooHuk Jung, Hosang Moon, Bosung Ku, and et al. 2021. "Inhibition of SARS-CoV-2 Virus Entry by the Crude Polysaccharides of Seaweeds and Abalone Viscera In Vitro" Marine Drugs 19, no. 4: 219. https://doi.org/10.3390/md19040219
APA StyleYim, S.-K., Kim, K., Kim, I., Chun, S., Oh, T., Kim, J.-U., Kim, J., Jung, W., Moon, H., Ku, B., & Jung, K. (2021). Inhibition of SARS-CoV-2 Virus Entry by the Crude Polysaccharides of Seaweeds and Abalone Viscera In Vitro. Marine Drugs, 19(4), 219. https://doi.org/10.3390/md19040219