
Exploring the Potential of Icelandic Seaweeds Extracts Produced by Aqueous Pulsed Electric Fields-Assisted Extraction for Cosmetic Applications

 ,
,  ,
,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. PEF-Assisted Extraction for the Processing of Icelandic Seaweed Biomass
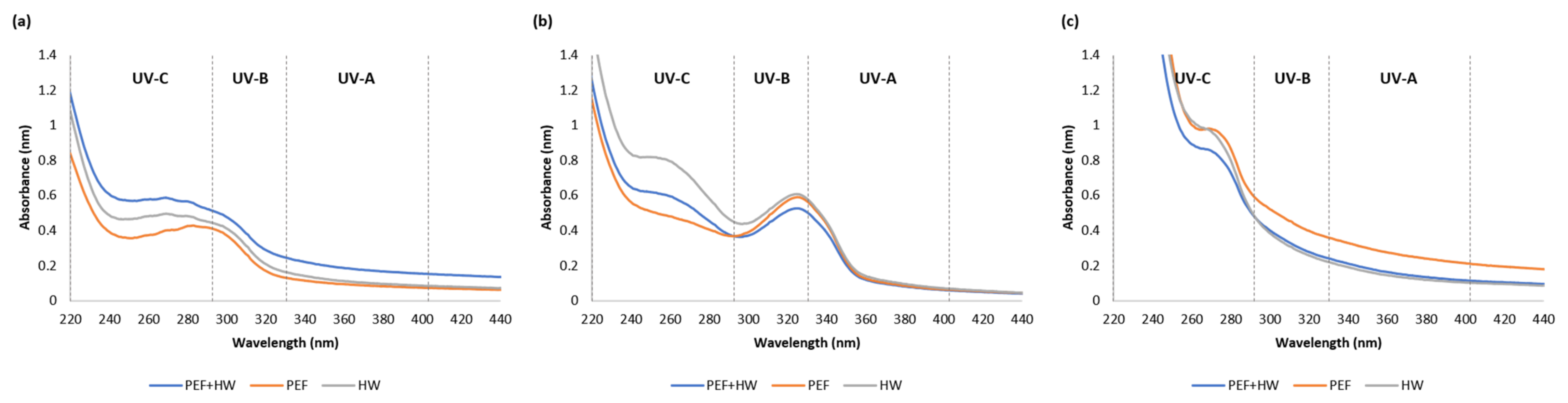
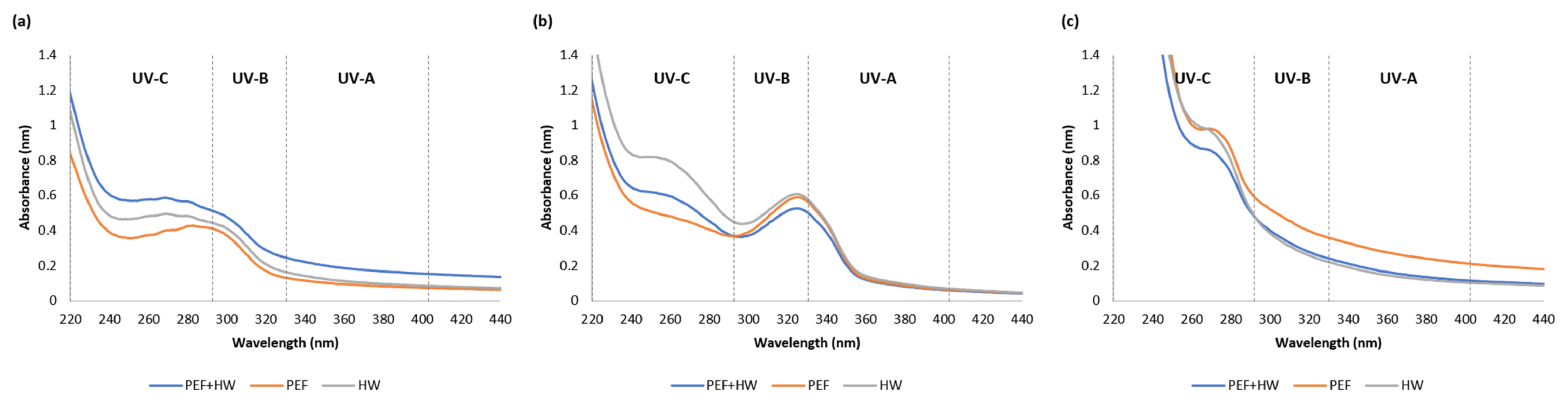
2.2. UV-VIS Absorption Spectra of Icelandic Seaweed Extracts
2.3. Total Phenolic, Flavonoid and Carbohydrate Content of Icelandic Seaweed Extracts
2.4. Antioxidant Capacities of Icelandic Seaweeds Extracts
2.5. Enzymatic Inhibitory Activities of Icelandic Seaweed Extracts
2.5.1. Collagenase Inhibition Activity
2.5.2. Elastase Inhibition Activity
2.5.3. Tyrosinase Inhibition Activity
2.5.4. Hyaluronidase Inhibition Activity
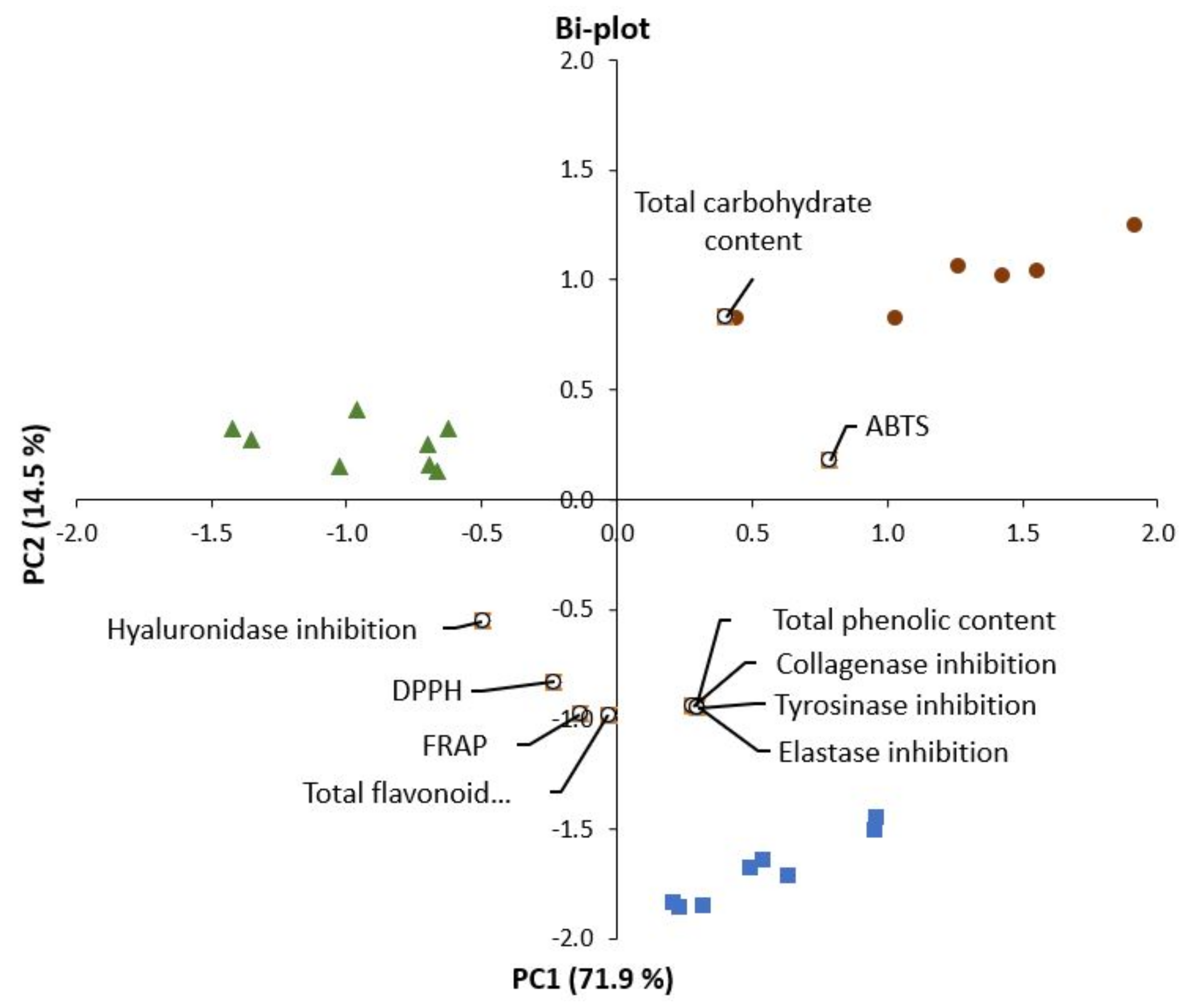
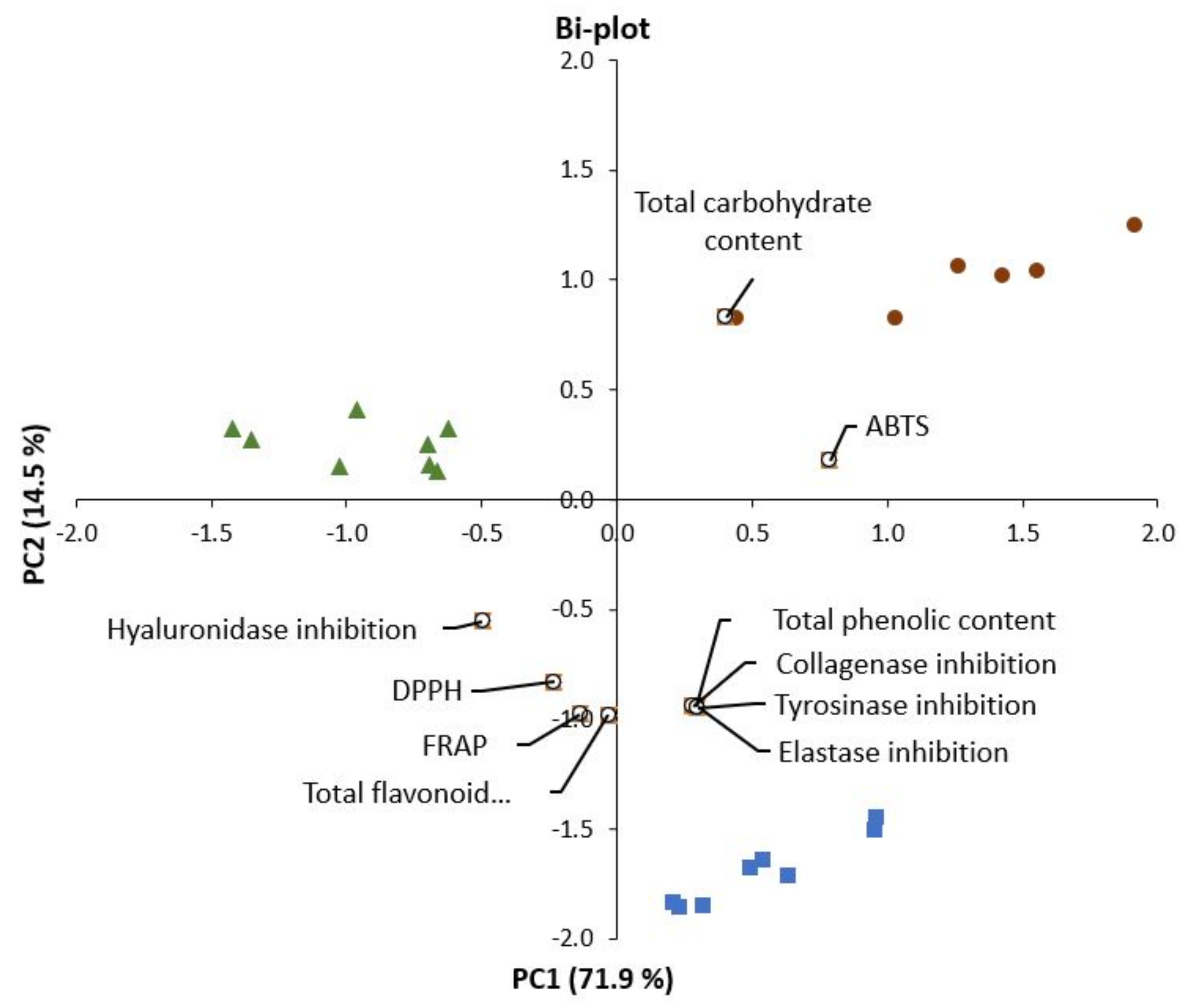
2.6. Correlations between Chemical Compounds and Bioactive Properties
3. Materials and Methods
3.1. Materials
3.2. Experimental Design
3.3. The Extraction of Bioactives from the Icelandic Seaweeds
3.3.1. Extraction Procedures
3.3.2. Conductivity, pH and Temperature Measurements
3.4. Spectral Profiles of the Seaweed Extracts
3.5. Determination of Total Polyphenolic Content
3.6. Determination of Total Flavonoid Content
3.7. Determination of Carbohydrate Content
3.8. Antioxidant Properties of Seaweeds Extracts
3.8.1. 2,2 Diphenyl-1-picrylhydrazyl (DPPH) Free Radical Scavenging Assay
3.8.2. Ferric Ion Reducing Antioxidant Power (FRAP) Assay
3.8.3. 2,2 Azino-bis(3-ethylbenzothiazoline-6-sulfonic Acid) (ABTS) Assay
3.9. Anti-Enzymatic Activities of Seaweeds Extracts
3.9.1. Collagenase Inhibition Assay
3.9.2. Elastase Inhibition Assay
3.9.3. Tyrosinase Inhibition Assay
3.9.4. Hyaluronidase Inhibition Assay
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Ariede, M.B.; Candido, T.M.; Jacome, A.L.M.; Velasco, M.V.R.; de Carvalho, J.C.M.; Baby, A.R. Cosmetic attributes of algae—A review. Algal Res. 2017, 25, 483–487. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Giger-Reverdin, S.; Lessire, M.; Lebas, F.; Ankers, P. Seaweeds for livestock diets: A review. Anim. Feed Sci. Technol. 2016, 212, 1–17. [Google Scholar] [CrossRef]
- O’Connor, J.; Meaney, S.; Williams, G.A.; Hayes, M. Extraction of Protein from Four Different Seaweeds Using Three Different Physical Pre-Treatment Strategies. Molecules 2020, 25, 2005. [Google Scholar] [CrossRef]
- Máximo, P.; Ferreira, L.M.; Branco, P.; Lima, P.; Lourenço, A. Secondary Metabolites and Biological Activity of Invasive Macroalgae of Southern Europe. Mar. Drugs 2018, 16, 265. [Google Scholar] [CrossRef] [Green Version]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for High-Value Products towards Human Health and Nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Zavaglia, A.; Prieto Lage, M.A.; Jimenez-Lopez, C.; Mejuto, J.C.; Simal-Gandara, J. The Potential of Seaweeds as a Source of Functional Ingredients of Prebiotic and Antioxidant Value. Antioxidants 2019, 8, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Sharifi-Rad, J.; Seca, A.M.L.; Pinto, D.C.G.A.; Michalak, I.; Trincone, A.; Mishra, A.P.; Nigam, M.; Zam, W.; Martins, N. Current Trends on Seaweeds: Looking at Chemical Composition, Phytopharmacology, and Cosmetic Applications. Molecules 2019, 24, 4182. [Google Scholar] [CrossRef] [Green Version]
- Ghazali, E.; Soon, P.C.; Mutum, D.S.; Nguyen, B. Health and cosmetics: Investigating consumers’ values for buying organic personal care products. J. Retail. Consum. Serv. 2017, 39, 154–163. [Google Scholar] [CrossRef]
- Amberg, N.; Fogarassy, C. Green Consumer Behavior in the Cosmetics Market. Resources 2019, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L. Seaweeds as Source of Bioactive Substances and Skin Care Therapy—Cosmeceuticals, Algotheraphy, and Thalassotherapy. Cosmetics 2018, 5, 68. [Google Scholar] [CrossRef] [Green Version]
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed marine natural products in the pharmaceutical and cosmeceutical industries: Tips for success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agatonovic-Kustrin, S.; Morton, D. Cosmeceuticals derived from bioactive substances found in marine algae. Oceanography 2013, 1, 106. [Google Scholar]
- Wang, H.-M.D.; Chen, C.-C.; Huynh, P.; Chang, J.-S. Exploring the potential of using algae in cosmetics. Bioresour. Technol. 2015, 184, 355–362. [Google Scholar] [CrossRef]
- Jahan, A.; Ahmad, I.Z.; Fatima, N.; Ansari, V.A.; Akhtar, J. Algal bioactive compounds in the cosmeceutical industry: A review. Phycologia 2017, 56, 410–422. [Google Scholar] [CrossRef]
- Morone, J.; Alfeus, A.; Vasconcelos, V.; Martins, R. Revealing the potential of cyanobacteria in cosmetics and cosmeceuticals—A new bioactive approach. Algal Res. 2019, 41, 101541. [Google Scholar] [CrossRef]
- Cikoš, A.-M.; Jerković, I.; Molnar, M.; Šubarić, D.; Jokić, S. New trends for macroalgal natural products applications. Nat. Prod. Res. 2019, 37, 1–12. [Google Scholar] [CrossRef]
- Thring, T.S.; Hili, P.; Naughton, D.P. Anti-collagenase, anti-elastase and anti-oxidant activities of extracts from 21 plants. BMC Complement. Altern. Med. 2009, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, C.; Sørensen, A.M.; Holdt, S.L.; Akoh, C.C.; Hermund, D.B. Source, Extraction, Characterization, and Applications of Novel Antioxidants from Seaweed. Annu. Rev. Food Sci. Technol. 2019, 10, 541–568. [Google Scholar] [CrossRef]
- Castejón, N.; Señoráns, F.J. Simultaneous extraction and fractionation of omega-3 acylglycerols and glycolipids from wet microalgal biomass of Nannochloropsis gaditana using pressurized liquids. Algal Res. 2019, 37, 74–82. [Google Scholar] [CrossRef]
- Mohamed, M.E.A.; Eissa, A.H.A. Pulsed electric fields for food processing technology. Struct. Funct. Food Eng. 2012, 11, 275–306. [Google Scholar]
- Geada, P.; Rodrigues, R.; Loureiro, L.; Pereira, R.; Fernandes, B.; Teixeira, J.A.; Vasconcelos, V.; Vicente, A.A. Electrotechnologies applied to microalgal biotechnology—Applications, techniques and future trends. Renew. Sustain. Energy Rev. 2018, 94, 656–668. [Google Scholar] [CrossRef] [Green Version]
- Poojary, M.M.; Barba, F.J.; Aliakbarian, B.; Donsì, F.; Pataro, G.; Dias, D.A.; Juliano, P. Innovative alternative technologies to extract carotenoids from microalgae and seaweeds. Mar. Drugs 2016, 14, 214. [Google Scholar] [CrossRef] [PubMed]
- Vorobiev, E.; Lebovka, N. 2—Extraction from Foods and Biomaterials Enhanced by Pulsed Electric Energy. In Innovative Food Processing Technologies; Knoerzer, K., Juliano, P., Smithers, G., Eds.; Woodhead Publishing: Sawston, UK, 2016; pp. 31–56. [Google Scholar]
- Käferböck, A.; Smetana, S.; de Vos, R.; Schwarz, C.; Toepfl, S.; Parniakov, O. Sustainable extraction of valuable components from Spirulina assisted by pulsed electric fields technology. Algal Res. 2020, 48, 101914. [Google Scholar] [CrossRef]
- Parniakov, O.; Barba, F.J.; Grimi, N.; Marchal, L.; Jubeau, S.; Lebovka, N.; Vorobiev, E. Pulsed electric field assisted extraction of nutritionally valuable compounds from microalgae Nannochloropsis spp. using the binary mixture of organic solvents and water. Innov. Food Sci. Emerg. Technol. 2015, 27, 79–85. [Google Scholar] [CrossRef]
- Scherer, D.; Krust, D.; Frey, W.; Mueller, G.; Nick, P.; Gusbeth, C. Pulsed electric field (PEF)-assisted protein recovery from Chlorella vulgaris is mediated by an enzymatic process after cell death. Algal Res. 2019, 41, 101536. [Google Scholar] [CrossRef]
- Naseri, A.; Marinho, G.S.; Holdt, S.L.; Bartela, J.M.; Jacobsen, C. Enzyme-assisted extraction and characterization of protein from red seaweed Palmaria palmata. Algal Res. 2020, 47, 101849. [Google Scholar] [CrossRef]
- Robin, A.; Kazir, M.; Sack, M.; Israel, A.; Frey, W.; Mueller, G.; Livney, Y.D.; Golberg, A. Functional Protein Concentrates Extracted from the Green Marine Macroalga Ulva sp., by High Voltage Pulsed Electric Fields and Mechanical Press. ACS Sustain. Chem. Eng. 2018, 6, 13696–13705. [Google Scholar] [CrossRef]
- Einarsdóttir, R.; Þórarinsdóttir, K.A.; Aðalbjörnsson, B.V.; Guðmundsson, M.; Marteinsdóttir, G.; Kristbergsson, K. The effect of pulsed electric field-assisted treatment parameters on crude aqueous extraction of Laminaria digitata. J. Appl. Phycol. 2021, 33, 3287–3296. [Google Scholar] [CrossRef]
- Postma, P.R.; Cerezo-Chinarro, O.; Akkerman, R.J.; Olivieri, G.; Wijffels, R.H.; Brandenburg, W.A.; Eppink, M.H.M. Biorefinery of the macroalgae Ulva lactuca: Extraction of proteins and carbohydrates by mild disintegration. J. Appl. Phycol. 2018, 30, 1281–1293. [Google Scholar] [CrossRef] [Green Version]
- Zbinden, M.D.A.; Sturm, B.S.M.; Nord, R.D.; Carey, W.J.; Moore, D.; Shinogle, H.; Stagg-Williams, S.M. Pulsed electric field (PEF) as an intensification pretreatment for greener solvent lipid extraction from microalgae. Biotechnol. Bioeng. 2013, 110, 1605–1615. [Google Scholar] [CrossRef]
- Silve, A.; Papachristou, I.; Wüstner, R.; Sträßner, R.; Schirmer, M.; Leber, K.; Guo, B.; Interrante, L.; Posten, C.; Frey, W. Extraction of lipids from wet microalga Auxenochlorella protothecoides using pulsed electric field treatment and ethanol-hexane blends. Algal Res. 2018, 29, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Chittapun, S.; Jonjaroen, V.; Khumrangsee, K.; Charoenrat, T. C-phycocyanin extraction from two freshwater cyanobacteria by freeze thaw and pulsed electric field techniques to improve extraction efficiency and purity. Algal Res. 2020, 46, 101789. [Google Scholar] [CrossRef]
- Aryee, A.N.A.; Agyei, D.; Akanbi, T.O. Recovery and utilization of seaweed pigments in food processing. Curr. Opin. Food Sci. 2018, 19, 113–119. [Google Scholar] [CrossRef]
- Nowacka, M.; Tappi, S.; Wiktor, A.; Rybak, K.; Miszczykowska, A.; Czyzewski, J.; Drozdzal, K.; Witrowa-Rajchert, D.; Tylewicz, U. The Impact of Pulsed Electric Field on the Extraction of Bioactive Compounds from Beetroot. Foods 2019, 8, 244. [Google Scholar] [CrossRef] [Green Version]
- Martínez, J.M.; Delso, C.; Álvarez, I.; Raso, J. Pulsed electric field-assisted extraction of valuable compounds from microorganisms. Comprehen. Rev. Food Sci. Food Saf. 2020, 19, 530–552. [Google Scholar] [CrossRef]
- Pataro, G.; Goettel, M.; Straessner, R.; Gusbeth, C.; Ferrari, G.; Frey, W. Effect of PEF treatment on extraction of valuable compounds from microalgae C. vulgaris. Chem. Eng. Trans. 2017, 57, 67–72. [Google Scholar]
- Brunton, N.P.; Luengo, E. Pulsed Electric Fields for Extraction of Secondary Metabolites from Plants. In Pulsed Electric Fields for Extraction of Secondary Metabolites from Plants; Miklavcic, D., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–15. [Google Scholar]
- Schiener, P.; Black, K.D.; Stanley, M.S.; Green, D.H. The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J. Appl. Phycol. 2015, 27, 363–373. [Google Scholar] [CrossRef]
- Salgado, L.T.; Tomazetto, R.; Cinelli, L.P.; Farina, M.; Filho, G.M.A. The influence of brown algae alginates on phenolic compounds capability of ultraviolet radiation absorption in vitro. Braz. J. Oceanogr. 2007, 55, 145–154. [Google Scholar] [CrossRef]
- Orfanoudaki, M.; Hartmann, A.; Karsten, U.; Ganzera, M. Chemical profiling of mycosporine-like amino acids in twenty-three red algal species. J. Phycol. 2019, 55, 393–403. [Google Scholar] [CrossRef]
- Pangestuti, R.; Siahaan, E.A.; Kim, S.-K. Photoprotective Substances Derived from Marine Algae. Mar. Drugs 2018, 16, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, G.; Figueroa, F.L.; Vega, J.; Chaves, P.; Álvarez-Gómez, F.; Korbee, N.; Bonomi-Barufi, J. Photoprotection properties of marine photosynthetic organisms grown in high ultraviolet exposure areas: Cosmeceutical applications. Algal Res. 2020, 49, 101956. [Google Scholar] [CrossRef]
- Nishida, Y.; Kumagai, Y.; Michiba, S.; Yasui, H.; Kishimura, H. Efficient Extraction and Antioxidant Capacity of Mycosporine-Like Amino Acids from Red Alga Dulse Palmaria palmata in Japan. Mar. Drugs 2020, 18, 502. [Google Scholar] [CrossRef] [PubMed]
- Rehm, E.; Dalgleish, F.; Huot, M.; Matteoli, S.; Archambault, P.; Lambert Girard, S.; Piché, M.; Lagunas-Morales, J. Comparing fluorescent and differential absorption LiDAR techniques for detecting algal biomass with applications to Arctic substrates. In Ocean Sensing and Monitoring X; International Society for Optics and Photonics: Bellingham, WA, USA, 2018; Volume 10631, p. 106310Z. [Google Scholar]
- Wang, T.; Jonsdottir, R.; Ólafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Bedoux, G.; Hardouin, K.; Burlot, A.S.; Bourgougnon, N. Chapter Twelve—Bioactive Components from Seaweeds: Cosmetic Applications and Future Development. In Advances in Botanical Research; Bourgougnon, N., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 71, pp. 345–378. [Google Scholar]
- Roleda, M.Y.; Marfaing, H.; Desnica, N.; Jónsdóttir, R.; Skjermo, J.; Rebours, C.; Nitschke, U. Variations in polyphenol and heavy metal contents of wild-harvested and cultivated seaweed bulk biomass: Health risk assessment and implication for food applications. Food Control 2019, 95, 121–134. [Google Scholar] [CrossRef]
- Ummat, V.; Tiwari, B.K.; Jaiswal, A.K.; Condon, K.; Garcia-Vaquero, M.; O’Doherty, J.; O’Donnell, C.; Rajauria, G. Optimisation of Ultrasound Frequency, Extraction Time and Solvent for the Recovery of Polyphenols, Phlorotannins and Associated Antioxidant Activity from Brown Seaweeds. Mar. Drugs 2020, 18, 250. [Google Scholar] [CrossRef]
- Afonso, C.; Matos, J.; Guarda, I.; Gomes-Bispo, A.; Gomes, R.; Cardoso, C.; Gueifão, S.; Delgado, I.; Coelho, I.; Castanheira, I.; et al. Bioactive and nutritional potential of Alaria esculenta and Saccharina latissima. J. Appl. Phycol. 2021, 33, 501–513. [Google Scholar] [CrossRef]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Gonçalves, A.M.M.; da Silva, G.J.; Pereira, L. Seaweed Phenolics: From Extraction to Applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef]
- Farasat, M.; Khavari-Nejad, R.A.; Nabavi, S.M.; Namjooyan, F. Antioxidant Activity, Total Phenolics and Flavonoid Contents of some Edible Green Seaweeds from Northern Coasts of the Persian Gulf. IJPR 2014, 13, 163–170. [Google Scholar] [PubMed]
- Manivannan, K.; Thirumaran, G.; Devi, G.K.; Anantharaman, P.; Balasubramanian, T. Proximate Composition of Different Group of Seaweeds from Vedalai Coastal Waters (Gulf of Mannar): Southeast Coast of India. Middle East J. Sci. Res. 2009, 4, 72–77. [Google Scholar]
- Mišurcová, L.; Škrovánková, S.; Samek, D.; Ambrožová, J.; Machů, L. Chapter 3—Health Benefits of Algal Polysaccharides in Human Nutrition. In Advances in Food and Nutrition Research; Henry, J., Ed.; Academic Press: Cambridge, MA, USA, 2012; Volume 66, pp. 75–145. [Google Scholar]
- Lafarga, T.; Acién-Fernández, F.G.; Garcia-Vaquero, M. Bioactive peptides and carbohydrates from seaweed for food applications: Natural occurrence, isolation, purification, and identification. Algal Res. 2020, 48, 101909. [Google Scholar] [CrossRef]
- Mutripah, S.; Meinita, M.D.N.; Kang, J.-Y.; Jeong, G.-T.; Susanto, A.B.; Prabowo, R.E.; Hong, Y.-K. Bioethanol production from the hydrolysate of Palmaria palmata using sulfuric acid and fermentation with brewer’s yeast. J. Appl. Phycol. 2014, 26, 687–693. [Google Scholar] [CrossRef]
- Dominguez, H.; Loret, E.P. Ulva lactuca, A Source of Troubles and Potential Riches. Mar. Drugs 2019, 17, 357. [Google Scholar] [CrossRef] [Green Version]
- Kidgell, J.T.; Magnusson, M.; de Nys, R.; Glasson, C.R.K. Ulvan: A systematic review of extraction, composition and function. Algal Res. 2019, 39, 101422. [Google Scholar] [CrossRef]
- Habeebullah, S.F.K.; Alagarsamy, S.; Arnous, A.; Jacobsen, C. Enzymatic extraction of antioxidant ingredients from Danish seaweeds and characterization of active principles. Algal Res. 2021, 56, 102292. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave assisted extraction of phenolic compounds from four economic brown macroalgae species and evaluation of their antioxidant activities and inhibitory effects on α-amylase, α-glucosidase, pancreatic lipase and tyrosinase. Int. Food Res. J. 2018, 113, 288–297. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef]
- Kainama, H.; Fatmawati, S.; Santoso, M.; Papilaya, P.M.; Ersam, T. The Relationship of Free Radical Scavenging and Total Phenolic and Flavonoid Contents of Garcinia lasoar PAM. Pharm. Chem. J. 2020, 53, 1151–1157. [Google Scholar] [CrossRef]
- Dang, T.T.; Van Vuong, Q.; Schreider, M.J.; Bowyer, M.C.; Van Altena, I.A.; Scarlett, C.J. Optimisation of ultrasound-assisted extraction conditions for phenolic content and antioxidant activities of the alga Hormosira banksii using response surface methodology. J. Appl. Phycol. 2017, 29, 3161–3173. [Google Scholar] [CrossRef]
- Couteau, C.; Coiffard, L. Chapter 14—Seaweed Application in Cosmetics. In Seaweed in Health and Disease Prevention; Fleurence, J., Levine, I., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 423–441. [Google Scholar]
- Tsukahara, K.; Takema, Y.; Moriwaki, S.; Tsuji, N.; Suzuki, Y.; Fujimura, T.; Imokawa, G. Selective Inhibition of Skin Fibroblast Elastase Elicits a Concentration-Dependent Prevention of Ultraviolet B-Induced Wrinkle Formation. J. Investig. Dermatol. 2001, 117, 671–677. [Google Scholar] [CrossRef]
- Liyanaarachchi, G.D.; Samarasekera, J.K.R.R.; Mahanama, K.R.R.; Hemalal, K.D.P. Tyrosinase, elastase, hyaluronidase, inhibitory and antioxidant activity of Sri Lankan medicinal plants for novel cosmeceuticals. Ind. Crops Prod. 2018, 111, 597–605. [Google Scholar] [CrossRef]
- Gupta, P.L.; Rajput, M.; Oza, T.; Trivedi, U.; Sanghvi, G. Eminence of Microbial Products in Cosmetic Industry. Nat. Prod. Bioprospect. 2019, 9, 267–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzyme Inhib. Med. 2019, 34, 279–309. [Google Scholar] [CrossRef] [Green Version]
- Couteau, C.; Coiffard, L. Phycocosmetics and Other Marine Cosmetics, Specific Cosmetics Formulated Using Marine Resources. Mar. Drugs 2020, 18, 322. [Google Scholar] [CrossRef]
- Sari, D.M.; Anwar, E.; Arifianti, A.E. Antioxidant and tyrosinase inhibitor activities of ethanol extracts of brown seaweed (Turbinaria conoides) as lightening ingredient. Pharm. J. 2019, 11, 379–382. [Google Scholar] [CrossRef] [Green Version]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.Y.; Baek, N.; Nam, T.-G. Natural, semisynthetic and synthetic tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2016, 31, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Lopes, G.; Gil-Izquierdo, A.; Andrade, P.B.; Sousa, C.; Mouga, T.; Valentão, P. Phlorotannin Extracts from Fucales Characterized by HPLC-DAD-ESI-MSn: Approaches to Hyaluronidase Inhibitory Capacity and Antioxidant Properties. Mar. Drugs 2012, 10, 2766–2781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayad, S.; Nehmé, R.; Tannoury, M.; Lesellier, E.; Pichon, C.; Morin, P. Macroalga Padina pavonica water extracts obtained by pressurized liquid extraction and microwave-assisted extraction inhibit hyaluronidase activity as shown by capillary electrophoresis. J. Chromatogr. A 2017, 1497, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Athukorala, Y.; Kim, K.-N.; Jeon, Y.-J. Antiproliferative and antioxidant properties of an enzymatic hydrolysate from brown alga, Ecklonia cava. Food Chem. Toxicol. 2006, 44, 1065–1074. [Google Scholar] [CrossRef]
- Jiménez-Escrig, A.; Gómez-Ordóñez, E.; Rupérez, P. Brown and red seaweeds as potential sources of antioxidant nutraceuticals. J. Appl. Phycol. 2012, 24, 1123–1132. [Google Scholar] [CrossRef]
- Karawita, R.; Siriwardhana, N.; Lee, K.-W.; Heo, M.-S.; Yeo, I.-K.; Lee, Y.-D.; Jeon, Y.-J. Reactive oxygen species scavenging, metal chelation, reducing power and lipid peroxidation inhibition properties of different solvent fractions from Hizikia fusiformis. Eur. Food Res. Technol. 2005, 220, 363–371. [Google Scholar] [CrossRef]
- Jormalainen, V.; Honkanen, T. Variation in natural selection for growth and phlorotannins in the brown alga Fucus vesiculosus. J. Evolut. Biol. 2004, 17, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Koivikko, R.; Loponen, J.; Pihlaja, K.; Jormalainen, V. High-performance liquid chromatographic analysis of phlorotannins from the brown alga Fucus vesiculosus. Phytochem. Anal. 2007, 18, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Mareček, V.; Mikyška, A.; Hampel, D.; Čejka, P.; Neuwirthová, J.; Malachová, A.; Cerkal, R. ABTS and DPPH methods as a tool for studying antioxidant capacity of spring barley and malt. J. Cereal Sci. 2017, 73, 40–45. [Google Scholar] [CrossRef]
- Asada, M.; Sugie, M.; Inoue, M.; Nakagomi, K.; Hongo, S.; Murata, K.; Irie, S.; Takeuchi, T.; Tomizuka, N.; Oka, S. Inhibitory Effect of Alginic Acids on Hyaluronidase and on Histamine Release from Mast Cells. Biosci. Biotechnol. Biochem. 1997, 61, 1030–1032. [Google Scholar] [CrossRef] [PubMed]
- Mase, T.; Yamauchi, M.; Kato, Y.; Esaki, H.; Isshiki, S. Hyaluronidase-Inhibiting Acidic Polysaccharide Isolated from Porphyridium Purpureum. The Collection of Research Essays in the Suishan Women’s Gakuen University. Nat. Sci. 2013, 44, 105–113. [Google Scholar]
- Tolpeznikaite, E.; Bartkevics, V.; Ruzauskas, M.; Pilkaityte, R.; Viskelis, P.; Urbonaviciene, D.; Zavistanaviciute, P.; Zokaityte, E.; Ruibys, R.; Bartkiene, E. Characterization of Macro- and Microalgae Extracts Bioactive Compounds and Micro- and Macroelements Transition from Algae to Extract. Foods 2021, 10, 2226. [Google Scholar] [CrossRef]
- Gómez, I.; Huovinen, P. Morpho-functional patterns and zonation of South Chilean seaweeds: The importance of photosynthetic and bio-optical traits. Mar. Ecol. Prog. Ser. 2011, 422, 77–91. [Google Scholar] [CrossRef] [Green Version]
- Karsten, U.; Wiencke, C. Factors Controlling the Formation of UV-absorbing Mycosporine-like Amino Acids in the Marine Red Alga Palmaria palmata from Spitsbergen (Norway). J. Plant. Physiol. 1999, 155, 407–415. [Google Scholar] [CrossRef]
- Ummat, V.; Sivagnanam, S.P.; Rajauria, G.; O’Donnell, C.; Tiwari, B.K. Advances in pre-treatment techniques and green extraction technologies for bioactives from seaweeds. Trends Food Sci. Technol 2021, 110, 90–106. [Google Scholar] [CrossRef]
- Boussetta, N.; Lanoisellé, J.-L.; Bedel-Cloutour, C.; Vorobiev, E. Extraction of soluble matter from grape pomace by high voltage electrical discharges for polyphenol recovery: Effect of sulphur dioxide and thermal treatments. J. Food Eng. 2009, 95, 192–198. [Google Scholar] [CrossRef]
- Goettel, M.; Eing, C.; Gusbeth, C.; Straessner, R.; Frey, W. Pulsed electric field assisted extraction of intracellular valuables from microalgae. Algal Res. 2013, 2, 401–408. [Google Scholar] [CrossRef]
- Hwang, P.-A.; Wu, C.-H.; Gau, S.-Y.; Chien, S.-Y.; Hwang, D.-F. Antioxidant and immune-stimulating activities of hot-water extract from seaweed Sargassum hemiphyllum. J. Mar. Sci. Technol. 2010, 18, 41–46. [Google Scholar] [CrossRef]
- Sabeena Farvin, K.H.; Jacobsen, C. Phenolic compounds and antioxidant activities of selected species of seaweeds from Danish coast. Food Chem. 2013, 138, 1670–1681. [Google Scholar] [CrossRef] [PubMed]
- Godlewska, K.; Michalak, I.; Tuhy, L.; Chojnacka, K. Plant Growth Biostimulants Based on Different Methods of Seaweed Extraction with Water. BioMed Res. Int. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Zhang, J.; Shen, J.; Silva, A.; Dennis, D.A.; Barrow, C.J. A Simple 96-Well Microplate Method for Estimation of Total Polyphenol Content in Seaweeds. J. Appl. Phycol. 2006, 18, 445–450. [Google Scholar] [CrossRef] [Green Version]
- Kamtekar, S.; Keer, V.; Patil, V. Estimation of phenolic content, flavonoid content, antioxidant and alpha amylase inhibitory activity of marketed polyherbal formulation. J. Appl. Pharm. Sci. 2014, 4, 61. [Google Scholar]
- Neto, R.; Marçal, C.; Queiros, A.; Abreu, M.; Silva, A.; Cardoso, S. Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as Functional Ingredients. Int. J. Mol. Sci. 2018, 19, 2987. [Google Scholar] [CrossRef] [Green Version]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Eun Lee, K.; Bharadwaj, S.; Yadava, U.; Gu Kang, S. Evaluation of caffeine as inhibitor against collagenase, elastase and tyrosinase using in silico and in vitro approach. J. Enzyme Inhib. Med. Chem. 2019, 34, 927–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Seaweed Specie and Extraction Method | T (°C) | Conductivity before (mS/cm) | Conductivity after (mS/cm) | pH before | pH after |
|---|---|---|---|---|---|
| A. esculenta | |||||
| HW | 95.0 | 16.433 ± 0.260 a | 18.413 ± 0.228 a | 6.49 ± 0.02 a | 6.28 ± 0.05 a |
| PEF | 36.0 | 16.790 ± 0.131 a | 17.713 ± 0.091 a | 6.37 ± 0.01 b | 6.32 ± 0.01 a |
| PEF+HW | 34.3/95 | 16.560 ± 0.259 a | 16.957 ± 1.799 a | 6.42 ± 0.01 c | 6.16 ± 0.03 b |
| P. palmata | |||||
| HW | 95.0 | 8.736 ± 0.09 a | 9.724 ± 0.362 a | 6.46 ± 0.02 a | 6.44 ± 0.04 a |
| PEF | 46.3 | 8.571 ± 0.19 a | 9.214 ± 0.129 a | 6.39 ± 0.02 b | 6.52 ± 0.02 a |
| PEF+HW | 44.3/95 | 8.460 ± 0.17 a | 9.271 ± 0.037 a | 6.42 ± 0.03 a,b | 6.41 ± 0.07 a |
| U. lactuca | |||||
| HW | 95.0 | 6.213 ± 0.02 a | 6.740 ± 0.081 a | 6.07 ± 0.04 a | 6.25 ± 0.05 a |
| PEF | 51.0 | 6.006 ± 0.11 b | 6.261 ± 0.200 b | 5.94 ± 0.04 b | 5.33 ± 0.06 b |
| PEF+HW | 49.3/95 | 6.128 ± 0.01 a,b | 6.437 ± 0.094 a,b | 5.96 ± 0.03 b | 6.12 ± 0.05 c |
| Seaweed Specie and Extraction Method | Total Phenolic Content (µg GAE/g dw) | Total Flavonoid Content (µg QE/g dw) | Total Carbohydrate Content (mg GluE/g dw) |
|---|---|---|---|
| A. esculenta | |||
| HW | 8937.1 ± 785.7 a | 12232.8 ± 1248.7 a | 44.8 ± 1.5 a |
| PEF | 9368.2 ± 407.1 a | 12426.4 ± 848.3 a | 59.6 ± 1.1 b |
| PEF+HW | 8303.8 ± 594.1 a | 11636.8 ± 1393.6 a | 65.2 ± 2.7 c |
| P. palmata | |||
| HW | 1850.5 ± 121.5 a | 805.0 ± 1.9 a | 510.5 ± 61.2 a |
| PEF | 1806.3 ± 104.2 a | 939.0 ± 95.9 a | 401.5 ± 43.8 a |
| PEF+HW | 1761.8 ± 97.8 a | 973.3 ± 45.8 a | 413.8 ± 26.5 a |
| U. lactuca | |||
| HW | 1950.6 ± 109.5 a | 4533.1 ± 89.3 a | 249.5 ± 21.1 a |
| PEF | 1592.0 ± 95.8 b | 3427.3 ± 199.0 b | 106.3 ± 21.2 b |
| PEF+HW | 1709.4 ± 49.4 b | 4496.7 ± 589.4 a | 224.7 ± 19.1 a |
| Seaweed Specie and Extraction Method | DPPH Scavenging Effect (%) | FRAP (µmol TE/100 g dw) | ABTS (µmol TE/100 g dw) |
|---|---|---|---|
| A. esculenta | |||
| HW | 93.8 ± 1.6 a | 984.4 ± 31.3 a | 86.5 ± 15.3 a |
| PEF | 91.8 ± 1.6 a | 960.7± 13.1 a | 89.2 ± 9.8 a |
| PEF + HW | 90.9 ± 1.0 a | 895.7 ± 46.8 a | 106.8 ± 8.3 a |
| P. palmata | |||
| HW | 69.4 ± 7.3 a | 426.3 ± 65.5 a | 113.0 ± 5.6 a |
| PEF | 65.0 ± 7.2 a | 301.0 ± 7.9 a | 101.8 ± 1.5 a |
| PEF + HW | 56.4 ± 3.3 a | 302.7 ± 78.9 a | 97.4 ± 41.1 a |
| U. lactuca | |||
| HW | 71.0 ± 5.7 a | 534.6 ± 42.4 a | 75.7 ± 12.5 a |
| PEF | 86.3 ± 0.5 b | 570.2 ± 26.5 a | 99.5 ± 5.9 a |
| PEF + HW | 71.9 ± 10.0 a | 547.8 ± 38.2 a | 81.6 ± 10.0 a |
| Samples | Collagenase Inhibition (%) | Elastase Inhibition (%) | Tyrosinase Inhibition (%)) | Hyaluronidase Inhibition (%) |
|---|---|---|---|---|
| A. esculenta | ||||
| HW | 71.7 ± 8.6 a | 73.4 ± 2.5 a | 95.5 ± 2.5 a | 100.0 ± 0.0 a |
| PEF | 90.8 ± 3.0 b | 73.8 ± 7.5 a | 92.9 ± 2.6 a | 100.0 ± 0.0 a |
| PEF+HW | 68.3 ± 5.6 a | 71.1 ± 3.0 a | 90.5 ± 4.6 a | 100.0 ± 0.0 a |
| P. palmata | ||||
| HW | 4.3 ± 7.4 a | NI | NI | 91.8 ± 0.5 a |
| PEF | 1.6 ± 1.8 a | NI | NI | 91.9 ± 0.0 a |
| PEF + HW | 2.3 ± 4.0 a | NI | NI | 89.5 ± 2.8 a |
| U. lactuca | ||||
| HW | 2.3 ± 2.1 a | NI | NI | 93.4 ± 0.6 a |
| PEF | 1.7 ± 2.9 a | NI | NI | 96.8 ± 0.4 b |
| PEF + HW | 1.5 ± 2.6 a | NI | NI | 97.3 ± 1.0 b |
| Inhibitor from kit * | 74.9 ± 1.2 | NT | NT | NT |
| EGCG (5 mM) * | 63.15 | NT | NT | NT |
| EGCG (0.5 mM) * | 13.70 | NT | NT | NT |
| Quercetin (1mM) * | NT | 100.0 ± 0.0 | 88.2 ± 1.4 | NT |
| Quercetin (0.5 mM) * | NT | 58.7 ± 11.7 | 74.8 ± 0.5 | NT |
| Tannic acid (1 mM) * | NT | NT | NT | 94.3 ± 0.2 |
| Tannic acid (0.5 mM) * | NT | NT | NT | 88.5 ± 3.2 |
| Chemical Composition | Antioxidant Capacity | Enzyme Inhibition | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variables | TPC | TFC | TCC | DPPH | FRAP | ABTS | Collag | Elast | Tyros | Hyalur |
| TPC | 1 | 0.95 *** | −0.70 *** | 0.74 *** | 0.91 *** | −0.04 | 0.98 *** | 0.99 *** | 0.99 *** | 0.42 * |
| TFC | 1 | −0.84 *** | 0.82 *** | 0.97 *** | −0.24 | 0.93 *** | 0.94 *** | 0.95 *** | 0.54 ** | |
| TCC | 1 | −0.91 *** | −0.86 *** | 0.31 | −0.68 *** | −0.71 *** | −0.71 *** | −0.60 ** | ||
| DPPH | 1 | 0.88 *** | −0.19 | 0.74 *** | 0.75 *** | 0.75 *** | 0.52 ** | |||
| FRAP | 1 | −0.25 | 0.90 *** | 0.91 *** | 0.91 *** | 0.61 ** | ||||
| ABTS | 1 | −0.03 | 0.00 | 0.00 | −0.18 | |||||
| Collag | 1 | 0.98 *** | 0.98 *** | 0.42 * | ||||||
| Elast | 1 | 0.998 *** | 0.43 * | |||||||
| Tyros | 1 | 0.43 * | ||||||||
| Seaweed Species | Extraction Procedure | ||
|---|---|---|---|
| A. esculenta | HW (95 °C, 45 min) n = 3 | PEF (10 min) n = 3 | PEF + HW (10 min + 95 °C, 45 min) n = 3 |
| P. palmata | HW (95 °C, 45 min) n = 3 | PEF (10 min) n = 3 | PEF + HW (10 min + 95 °C, 45 min) n = 3 |
| U. lactuca | HW (95 °C, 45 min) n = 3 | PEF (10 min) n = 3 | PEF + HW (10 min + 95 °C, 45 min) n = 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castejón, N.; Thorarinsdottir, K.A.; Einarsdóttir, R.; Kristbergsson, K.; Marteinsdóttir, G. Exploring the Potential of Icelandic Seaweeds Extracts Produced by Aqueous Pulsed Electric Fields-Assisted Extraction for Cosmetic Applications. Mar. Drugs 2021, 19, 662. https://doi.org/10.3390/md19120662
Castejón N, Thorarinsdottir KA, Einarsdóttir R, Kristbergsson K, Marteinsdóttir G. Exploring the Potential of Icelandic Seaweeds Extracts Produced by Aqueous Pulsed Electric Fields-Assisted Extraction for Cosmetic Applications. Marine Drugs. 2021; 19(12):662. https://doi.org/10.3390/md19120662
Chicago/Turabian StyleCastejón, Natalia, Kristin Anna Thorarinsdottir, Ragnhildur Einarsdóttir, Kristberg Kristbergsson, and Gudrún Marteinsdóttir. 2021. "Exploring the Potential of Icelandic Seaweeds Extracts Produced by Aqueous Pulsed Electric Fields-Assisted Extraction for Cosmetic Applications" Marine Drugs 19, no. 12: 662. https://doi.org/10.3390/md19120662
APA StyleCastejón, N., Thorarinsdottir, K. A., Einarsdóttir, R., Kristbergsson, K., & Marteinsdóttir, G. (2021). Exploring the Potential of Icelandic Seaweeds Extracts Produced by Aqueous Pulsed Electric Fields-Assisted Extraction for Cosmetic Applications. Marine Drugs, 19(12), 662. https://doi.org/10.3390/md19120662