
Wound Healing Metabolites from Peters’ Elephant-Nose Fish Oil: An In Vivo Investigation Supported by In Vitro and In Silico Studies

 , ,
, ,  ,
,  ,
,  ,
,  and
and 
Abstract
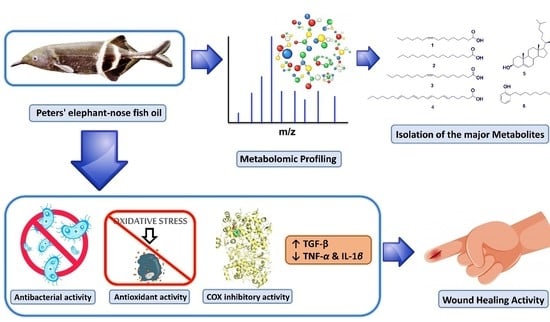
1. Introduction
2. Results
2.1. GC-MS Analysis
2.2. Physicochemical Investigation of Oil
2.3. In Vitro COX-1 and COX-2 Inhibitory Activity
2.4. Molecular Modeling
2.5. In Vitro Antioxidant Activity
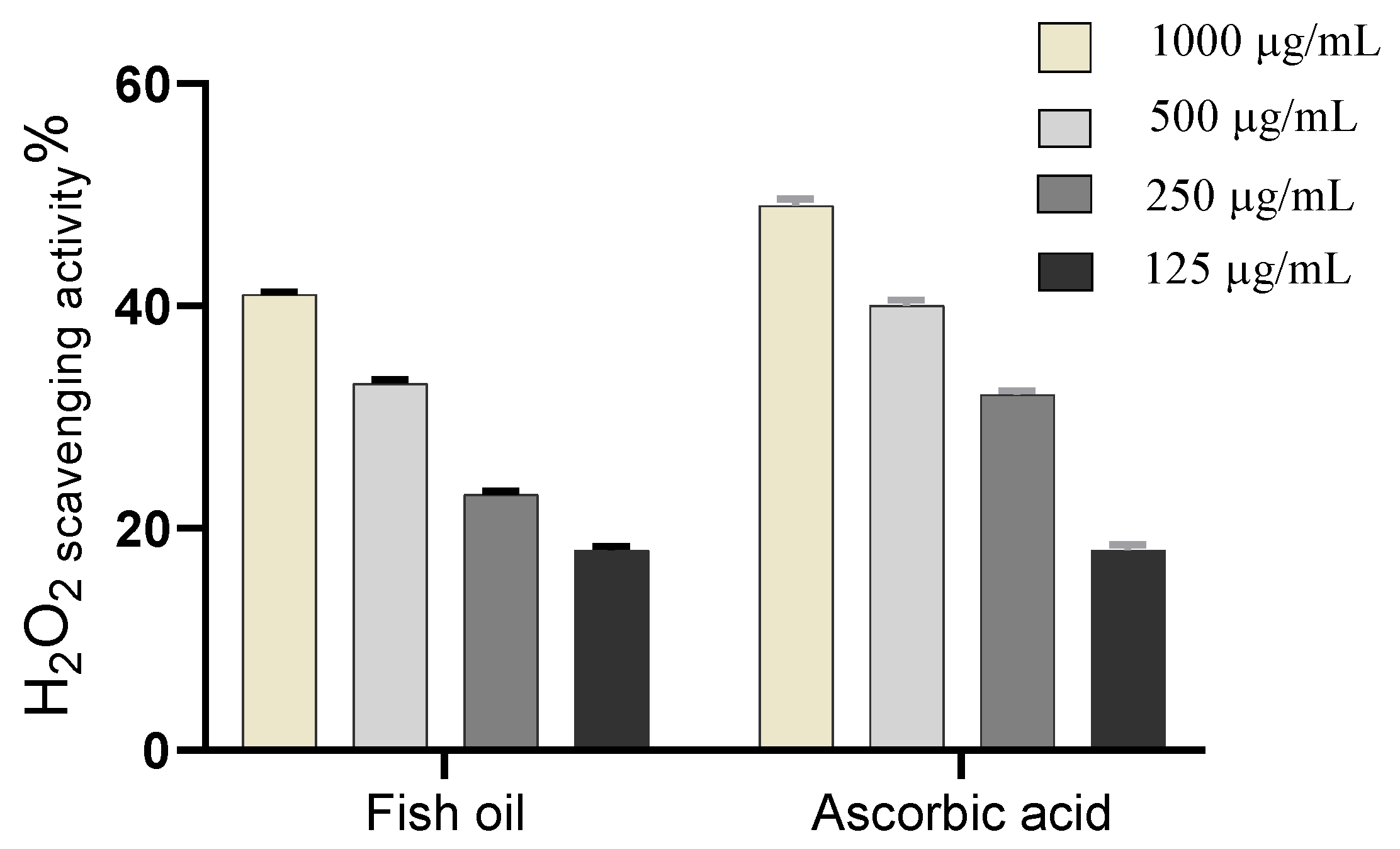
2.5.1. Hydrogen Peroxide Scavenging
2.5.2. Superoxide Radical Scavenging
2.6. Wound Healing Activity
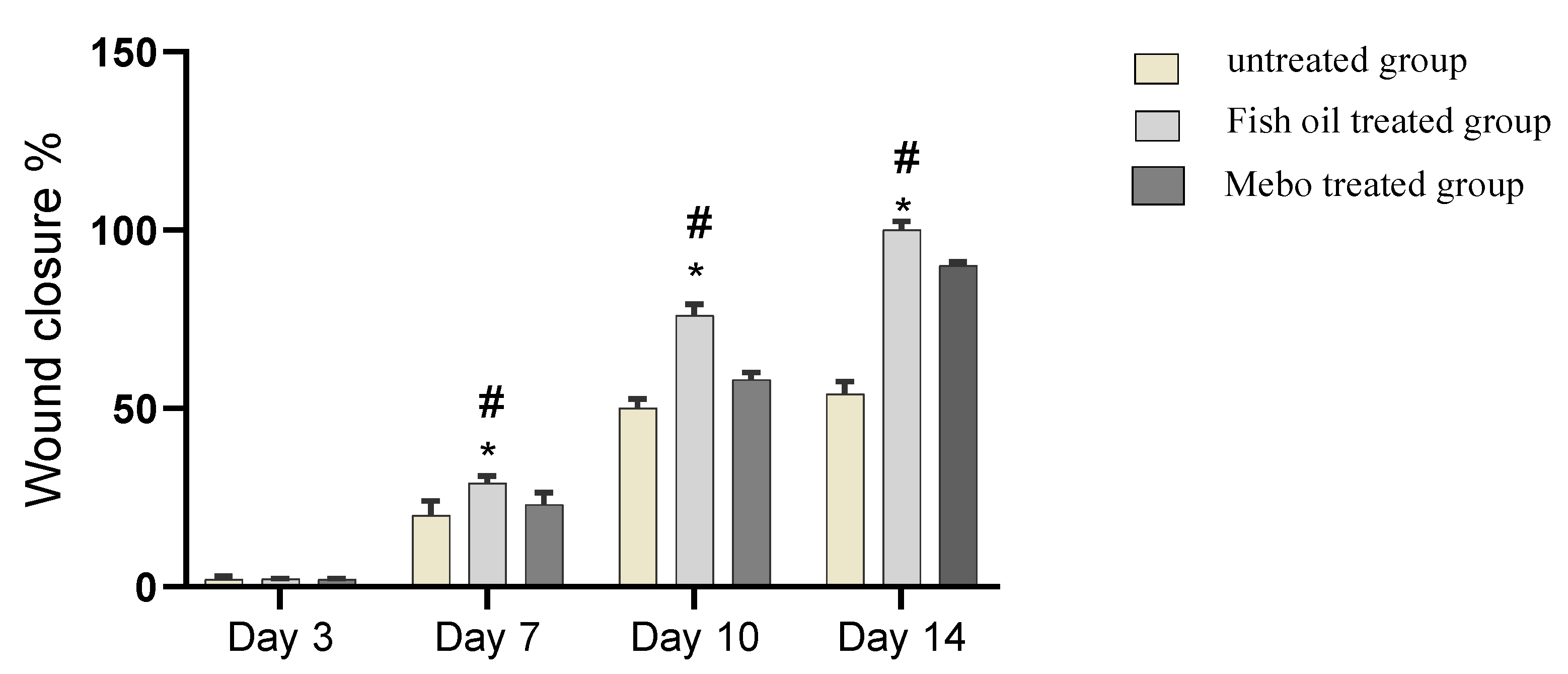
2.6.1. Wound Closure Rate Estimation
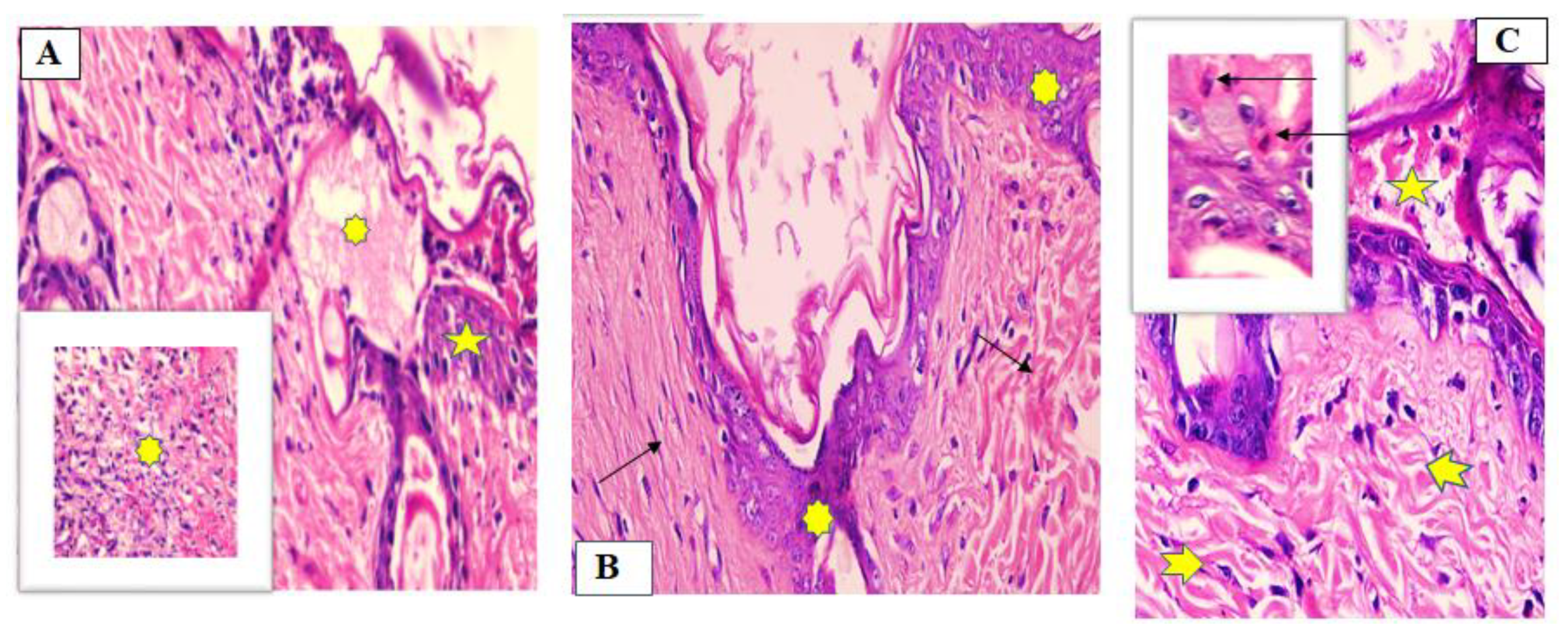
2.6.2. Histopathological Study
7 Days after Treatment
- Group 1 (untreated group)
- Group II (fish oil-treated group)
- Group III (Mebo®-treated group)
14 Days after Treatment
- Group I (untreated group)
- Group II (fish oil-treated group)
- Group III (Mebo®-treated group)
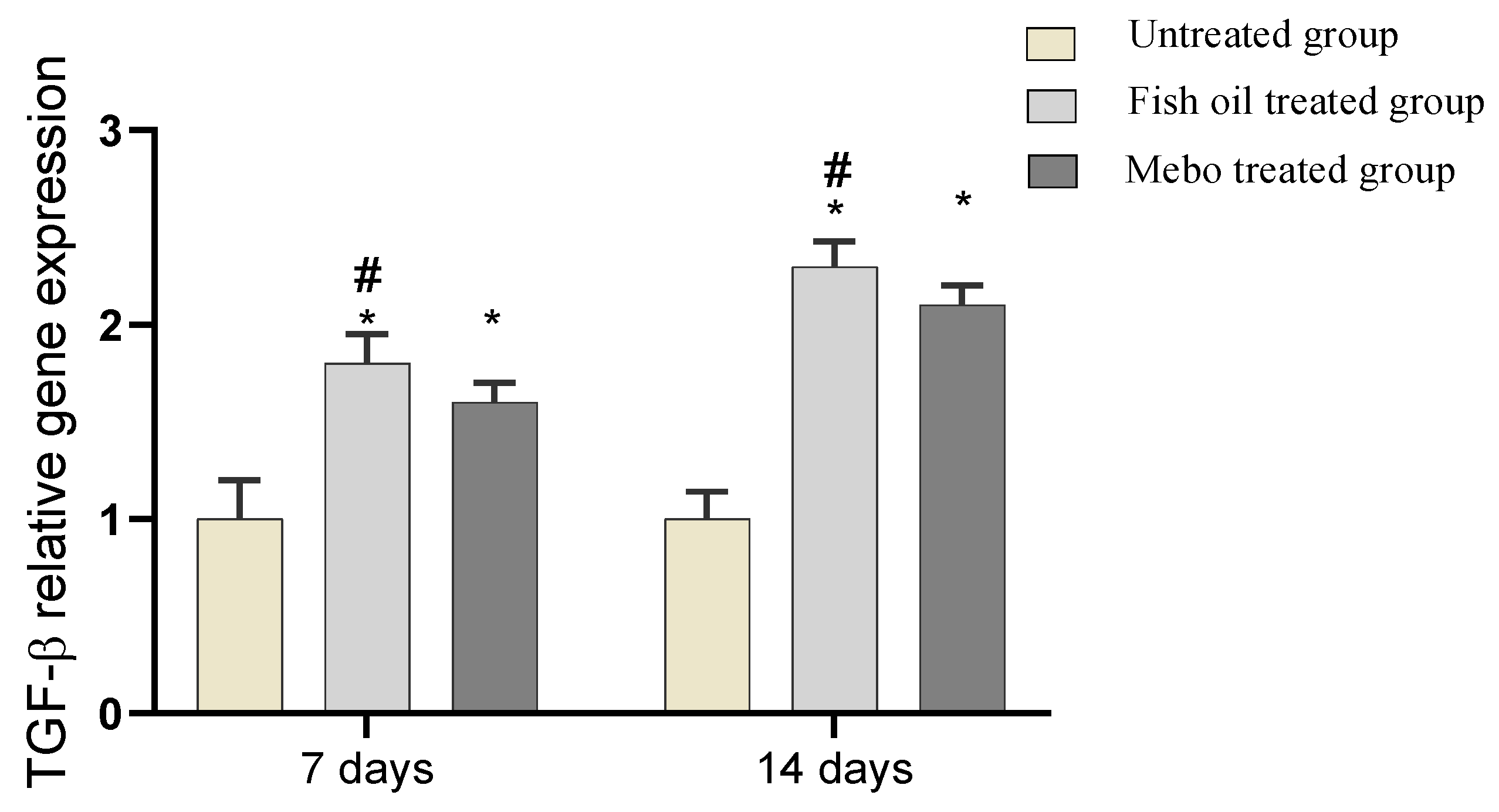
2.6.3. Effect of Fish Oil on mRNA Expression of TGF-β in Experimental Animals
2.6.4. In Vivo Effect of Fish Oil on mRNA Expression of Inflammatory Markers, i.e., TNF-α and IL-1β
3. Discussion
4. Materials and Methods
4.1. Fish Collection
4.2. Chemicals and Reagents
4.3. NMR Spectral Analyses
4.4. Sample Preparation and Lipid Extraction
4.5. Preparation of Fatty Acids Methyl Esters
4.6. GC-MS Analysis of Fatty Acids Methyl Esters
4.7. Isolation and Purification of Compounds
4.8. In Vitro Cyclooxygenases Inhibitory Activity
4.9. Molecular Modeling
4.9.1. Docking Analysis
4.9.2. Molecular Dynamic Simulation
4.10. In Vitro Antioxidant Activity
4.10.1. Hydrogen Peroxide Scavenging Activity
4.10.2. Superoxide Radical Scavenging Activity
4.11. Wound Healing Activity
4.11.1. Animal Treatment
4.11.2. Preparation of the Test Samples for the Bioassay
4.11.3. Circular Excision Wound Model and Experimental Design
4.11.4. Collection of Tissue Samples
4.11.5. Percentage Wound Closure Rate
4.11.6. Histological Study
4.11.7. Gene Expression Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Erkan, N.; Özden, Ö. Proximate composition and mineral contents in aqua cultured sea bass (Dicentrarchus labrax), sea bream (Sparus aurata) analyzed by ICP-MS. Food Chem. 2007, 102, 721–725. [Google Scholar] [CrossRef]
- Petricorena, Z.C. Chemical composition of fish and fishery products. In Handbook of Food Chemistry; Cheung, P.C.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1–28. [Google Scholar]
- Fournier, V.; Destaillats, F.; Juanéda, P.; Dionisi, F.; Lambelet, P.; Sébédio, J.-L.; Berdeaux, O. Thermal degradation of long-chain polyunsaturated fatty acids during deodorization of fish oil. Eur. J. Lipid Sci. Technol. 2006, 108, 33–42. [Google Scholar] [CrossRef]
- Teitelbaum, J.E.; Walker, W.A. Review: The role of omega 3 fatty acids in intestinal inflammation. J. Nutr. Biochem. 2001, 12, 21–32. [Google Scholar] [CrossRef]
- Acar, N.; Chardigny, J.-M.; Bonhomme, B.; Almanza, S.; Doly, M.; Sébédio, J.-L. Long-term intake of trans (n-3) polyunsatu-rated fatty acids reduces the b-wave amplitude of electroretinograms in rats. J. Nutr. 2002, 132, 3151–3154. [Google Scholar] [CrossRef]
- Vancassel, S. Oméga 3 et neurotransmission cérébrale. Oléagineux Corps Gras Lipides 2004, 11, 58–65. [Google Scholar] [CrossRef][Green Version]
- Al-Khudairy, L.; Hartley, L.; Clar, C.; Flowers, N.; Hooper, L.; Rees, K. Omega 6 fatty acids for the primary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2015, CD011094. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Christensen, V.; Walters, C.J.; Ahrens, R.; Alder, J.; Buszowski, J.; Christensen, L.B.; Cheung, W.W.L.; Dunne, J.; Froese, R.; Karpouzi, V.; et al. Database-driven models of the world’s Large Marine Ecosystems. Ecol. Model. 2009, 220, 1984–1996. [Google Scholar] [CrossRef]
- Engelmann, J.; Nöbel, S.; Röver, T.; Von Der Emde, G. The Schnauzenorgan-response of Gnathonemus petersii. Front. Zool. 2009, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Childs, D.R.; Murthy, A.S. Overview of Wound Healing and Management. Surg. Clin. N. Am. 2017, 97, 189–207. [Google Scholar] [CrossRef] [PubMed]
- Pizzimenti, S.; Toaldo, C.; Pettazzoni, P.; Dianzani, M.U.; Barrera, G. The “Two-Faced” Effects of Reactive Oxygen Species and the Lipid Peroxidation Product 4-Hydroxynonenal in the Hallmarks of Cancer. Cancers 2010, 2, 338–363. [Google Scholar] [CrossRef]
- Sun, M.-L.; Zhao, F.; Chen, X.-L.; Zhang, X.-Y.; Zhang, Y.-Z.; Song, X.-Y.; Sun, C.-Y.; Yang, J. Promotion of Wound Healing and Prevention of Frostbite Injury in Rat Skin by Exopolysaccharide from the Arctic Marine Bacterium Polaribacter sp. SM1127. Mar. Drugs 2020, 18, 48. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.R.; Burger, B.; Kühl, C.M.C.; Candreva, T.; Dos Anjos, M.B.P.; Rodrigues, H.G. Wound Healing and Omega-6 Fatty Acids: From Inflammation to Repair. Mediat. Inflamm. 2018, 2018, 2503950. [Google Scholar] [CrossRef]
- Chun, O.K.; Kim, D.-O.; Lee, C.Y. Superoxide Radical Scavenging Activity of the Major Polyphenols in Fresh Plums. J. Agric. Food Chem. 2003, 51, 8067–8072. [Google Scholar] [CrossRef]
- Alexandri, E.; Ahmed, R.; Siddiqui, H.; Choudhary, M.I.; Tsiafoulis, C.G.; Gerothanassis, I.P. High resolution NMR spec-troscopy as a structural and analytical tool for unsaturated lipids in solution. Molecules 2017, 22, 1663. [Google Scholar] [CrossRef] [PubMed]
- Malarvizhi, D.; Anusooriya, P.; Meenakshi, P.; Sundaram, S.; Oirere, E.; Gopalakrishnan, V.K. Isolation, Structural charac-terization of oleic acid from Zaleya decandra root extract. Anal. Chem. Lett. 2016, 6, 669–677. [Google Scholar] [CrossRef]
- Li, D.; Schröder, M.; Vetter, W. Isolation of 6,9,12,15-Hexadecatetraenoic Fatty Acid (16:4n-1) Methyl Ester from Transesterified Fish Oil by HSCCC. Chromatographia 2012, 75, 1–6. [Google Scholar] [CrossRef]
- Pierre, L.L.; Moses, M.N. Isolation and characterisation of stigmasterol and Β-sitosterol from Odontonema strictum (Acan-thaceae). J. Innov. Pharm. Biol. Sci. 2015, 2, 88–95. [Google Scholar]
- Roufogalis, B.D.; Li, Q.; Tran, V.H.; Kable, E.P.W.; Duke, C.C. Investigation of plant-derived phenolic compounds as plasma membrane Ca2+-ATPase inhibitors with potential cardiovascular activity. Drug Dev. Res. 1999, 46, 239–249. [Google Scholar] [CrossRef]
- Pereira, L.M.; Hatanaka, E.; Martins, E.F.; Oliveira, F.; Liberti, E.A.; Farsky, S.H.; Curi, R.; Pithon-Curi, T.C. Effect of oleic and linoleic acids on the inflammatory phase of wound healing in rats. Cell Biochem. Funct. 2008, 26, 197–204. [Google Scholar] [CrossRef]
- Alexander, J.W.; Supp, D. Role of Arginine and Omega-3 Fatty Acids in Wound Healing and Infection. Adv. Wound Care 2014, 3, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Jara, C.P.; Mendes, N.F.; Prado, T.P.D.; de Araújo, E.P. Bioactive Fatty Acids in the Resolution of Chronic Inflammation in Skin Wounds. Adv. Wound Care 2020, 9, 472–490. [Google Scholar] [CrossRef]
- Fuller, B. Role of PGE-2 and other inflammatory mediators in skin aging and their inhibition by topical natural anti-inflammatories. Cosmetics 2019, 6, 6. [Google Scholar] [CrossRef]
- Romana-Souza, B.; dos Santos, J.S.; Bandeira, L.G.; Monte-Alto-Costa, A. Selective inhibition of COX-2 improves cutaneous wound healing of pressure ulcers in mice through reduction of iNOS expression. Life Sci. 2016, 153, 82–92. [Google Scholar] [CrossRef]
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell. Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef]
- Krzyszczyk, P.; Schloss, R.; Palmer, A.; Berthiaume, F. The Role of Macrophages in Acute and Chronic Wound Healing and Interventions to Promote Pro-wound Healing Phenotypes. Front. Physiol. 2018, 9, 419. [Google Scholar] [CrossRef]
- Pachuau, L. Recent developments in novel drug delivery systems for wound healing. Expert Opin. Drug Deliv. 2015, 12, 1895–1909. [Google Scholar] [CrossRef] [PubMed]
- Suguna, L.; Singh, S.; Sivakumar, P.; Sampath, P.; Chandrakasan, G. Influence of Terminalia chebula on dermal wound healing in rats. Phytother. Res. 2002, 16, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Yin, L.; Yang, J.; Shan, G. Emodin, an anthraquinone derivative from Rheum officinale Baill, enhances cutaneous wound healing in rats. Eur. J. Pharmacol. 2007, 567, 177–185. [Google Scholar] [CrossRef]
- Huang, T.-H.; Wang, P.-W.; Yang, S.-C.; Chou, W.-L.; Fang, J.-Y. Cosmetic and Therapeutic Applications of Fish Oil’s Fatty Acids on the Skin. Mar. Drugs 2018, 16, 256. [Google Scholar] [CrossRef]
- Wankell, M.; Munz, B.; Hübner, G.; Hans, W.; Wolf, E.; Goppelt, A.; Werner, S. Impaired wound healing in transgenic mice overexpressing the activin antagonist follistatin in the epidermis. EMBO J. 2001, 20, 5361–5372. [Google Scholar] [CrossRef]
- Beer, H.-D.; Gassmann, M.G.; Munz, B.; Steiling, H.; Engelhardt, F.; Bleuel, K.; Werner, S. Expression and Function of Keratinocyte Growth Factor and Activin in Skin Morphogenesis and Cutaneous Wound Repair. J. Investig. Dermatol. Symp. Proc. 2000, 5, 34–39. [Google Scholar] [CrossRef]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in Wound Healing: A Comprehensive Review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef]
- Haroon, Z.A.; Amin, K.; Saito, W.; Wilson, W.; Greenberg, C.S.; Dewhirst, M.W. SU5416 Delays Wound Healing through Inhibition of TGF-β Activation. Cancer Biol. Ther. 2002, 1, 121–126. [Google Scholar] [CrossRef]
- Feinberg, R.A.; Hokama, L.; De Ruyter, K.; Keen, C.; Kim, I.-S. Operational determinants of caller satisfaction in the call center. Int. J. Serv. Ind. Manag. 2000, 11, 131–141. [Google Scholar] [CrossRef]
- Schultz, G.; Ladwig, G.; Wysocki, A. Extracellular Matrix: Review of Its Roles in Acute and Chronic Wounds. World Wide Wounds. 2005. Available online: http://www.worldwidewounds.com/2005/august/Schultz/Extrace-Matric-Acute-Chronic-Wounds.html (accessed on 10 October 2021).
- Houghton, P.J.; Hylands, P.J.; Mensah, A.Y.; Hensel, A.; Deters, A.M. In vitro tests and ethnopharmacological investiga-tions: Wound healing as an example. J. Ethnopharmacol. 2005, 100, 100–107. [Google Scholar] [CrossRef]
- Siwik, D.A.; Pagano, P.J.; Colucci, W. Oxidative stress regulates collagen synthesis and matrix metalloproteinase activity in cardiac fibroblasts. Am. J. Physiol. Physiol. 2001, 280, C53–C60. [Google Scholar] [CrossRef]
- Rodriguez-Menocal, L.; Shareef, S.; Salgado, M.; Shabbir, A.; Van Badiavas, E. Role of whole bone marrow, whole bone marrow cultured cells, and mesenchymal stem cells in chronic wound healing. Stem Cell Res. Ther. 2015, 6, 24. [Google Scholar] [CrossRef]
- Singh, K.; Agrawal, N.K.; Gupta, S.K.; Sinha, P.; Singh, K. Increased expression of TLR9 associated with pro-inflammatory S100A8 and IL-8 in diabetic wounds could lead to unresolved inflammation in type 2 diabetes mellitus (T2DM) cases with impaired wound healing. J. Diabetes Its Complicat. 2016, 30, 99–108. [Google Scholar] [CrossRef]
- Olaosebikan, B.; Raji, A. Field Guide to Nigerian Freshwater Fishes; New Bussa (Nigeria) Federal College of Freshwater Fisheries Technology: New Bussa, Nigeria, 1998. [Google Scholar]
- Ashworth, M.R.F.; Stahl, E. Thin-Layer Chromatography: A Laboratory Handbook; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Robert, A.; Mfilinge, P.; Limbu, S.M.; Mwita, C.J. Fatty Acid Composition and Levels of Selected Polyunsaturated Fatty Acids in Four Commercial Important Freshwater Fish Species from Lake Victoria, Tanzania. J. Lipids 2014, 2014, 712134. [Google Scholar] [CrossRef]
- Elmaidomy, A.H.; Hassan, H.M.; Amin, E.; Mohamed, W.; Hetta, M.H. Premna odorata Volatile Oil as a New Mycobacterium tuberculosis Growth Inhibitor for the Control of Tuberculosis Disease. Eur. J. Med. Plants 2017, 21, 1–11. [Google Scholar] [CrossRef]
- Demasi, M.; Caughey, G.; James, M.; Cleland, L. Assay of cyclooxygenase-1 and 2 in human monocytes. Inflamm. Res. 2000, 49, 737–743. [Google Scholar] [CrossRef]
- Walker, M.C.; Gierse, J.K. In vitro assays for cyclooxygenase activity and inhibitor characterization. In Cyclooxygenases; Springer: Berlin/Heidelberg, Germany, 2010; pp. 131–144. [Google Scholar]
- Wang, Q.; Lin, B.; Li, Z.; Su, J.; Feng, Y. Cichoric acid ameliorates monosodium urate-induced inflammatory response by reducing NLRP3 inflammasome activation via inhibition of NF-kB signaling pathway. Evid.-Based Complement. Altern. Med. 2021, 2021, 8868527. [Google Scholar] [CrossRef]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput.-Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef]
- Rimon, G.; Sidhu, R.S.; Lauver, D.A.; Lee, J.Y.; Sharma, N.P.; Yuan, C.; Frieler, R.A.; Trievel, R.C.; Lucchesi, B.R.; Smith, W.L. Coxibs interfere with the action of aspirin by binding tightly to one monomer of cyclooxygenase-1. Proc. Natl. Acad. Sci. USA 2010, 107, 28–33. [Google Scholar] [CrossRef]
- Vecchio, A.J.; Simmons, D.M.; Malkowski, M.G. Structural Basis of Fatty Acid Substrate Binding to Cyclooxygenase-2. J. Biol. Chem. 2010, 285, 22152–22163. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef]
- MacKerell, A.D., Jr.; Bashford, D.; Bellott, M.; Dunbrack, R.L., Jr.; Evanseck, J.D.; Field, M.J.; Fischer, S.; Gao, J.; Guo, H.; Ha, S. All-atom empirical potential for molecular modeling and dynamics studies of proteins. J. Phys. Chem. B 1998, 102, 3586–3616. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Jo, S.; Jiang, W.; Lee, H.S.; Roux, B.; Im, W. CHARMM-GUI Ligand Binder for Absolute Binding Free Energy Calculations and Its Application. J. Chem. Inf. Model. 2013, 53, 267–277. [Google Scholar] [CrossRef]
- Hassan, H.A.; Abdel-Aziz, A.F. Evaluation of free radical-scavenging and anti-oxidant properties of black berry against flu-oride toxicity in rats. Food Chem. Toxicol. 2010, 48, 1999–2004. [Google Scholar] [CrossRef] [PubMed]
- Sonboli, A.; Mojarrad, M.; Ebrahimi, S.N.; Enayat, S. Free Radical Scavenging Activity and Total Phenolic Content of Methanolic Extracts from Male Inflorescence of Salix aegyptiaca Grown in Iran. Iran. J. Pharm. Res. IJPR 2010, 9, 293–296. [Google Scholar]
- Tramontina, V.; Machado, M.A.N.; Filho, G.D.R.N.; Kim, S.H.; Vizzioli, M.R.; De Toledo, S. Effect of bismuth subgallate (local hemostatic agent) on wound healing in rats. Histological and histometric findings. Braz. Dent. J. 2002, 13, 11–16. [Google Scholar]
- Hummon, A.B.; Lim, S.R.; Difilippantonio, M.J.; Ried, T. Isolation and solubilization of proteins after TRIzol® extraction of RNA and DNA from patient material following prolonged storage. Biotechniques 2007, 42, 467–472. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Identified Compound | C:D | Type | Area % | RT | RI |
|---|---|---|---|---|---|---|
| 1 | Caprylic acid | C8:0 | SFA | 1.18 | 9.69 | 892 |
| 2 | Pelargonic acid | C9:0 | SFA | 0.44 | 9.39 | 875 |
| 3 | Caproic acid | C10:0 | SFA | 0.22 | 10.05 | 845 |
| 4 | Lauric acid | C12:0 | SFA | 1.53 | 11.18 | 882 |
| 5 | Myristic acid | C14:0 | SFA | 2.80 | 12.92 | 907 |
| 6 | 13-Methyl, Myristic acid | C15:0 | SFA | 3.20 | 14.03 | 909 |
| 7 | Pentadecanoic acid | C15:0 | SFA | 3.26 | 15.11 | 891 |
| 8 | Palmitic acid | C16:0 | SFA | 4.11 * | 15.70 | 810 |
| 9 | 6,9,12-Octadecatrienoic acid | C18:3 (6,9,12) | PUFA | 1.67 | 18.69 | 841 |
| 10 | 7,10,13-Eicosatrienoic acid | C20:3 (7,10,13) | PUFA | 1.57 | 19.57 | 843 |
| 11 | Arachidonic acid | C20:4 (5,8,11,14) | PUFA | 2.45 | 20.60 | 931 |
| 12 | 6,9,12,15-Docosatetraenoic acid | C22:4 (6,9,12,15) | PUFA | 4.63 * | 21.59 | 816 |
| 13 | Eicosa-5,8,11,14,17-pentaenoic acid | C20:5 (5,8,11,14,15) | PUFA | 2.65 | 22.07 | 878 |
| 14 | 7,10-Octadecadienoic acid | C18:2 (7,10) | PUFA | 0.75 | 24.35 | 882 |
| 15 | 10-Heptadecen-8-ynoic acid | C17:2 (8,10) | PUFA | 0.45 | 25.75 | 678 |
| 16 | 9,12-Octadecadienoic acid | C18:2 (9,12) | PUFA | 1.91 | 26.5 | 871 |
| 17 | 4,7,10,13,16,19-Docosahexaenoic acid | C22:6 (4,7,10,13,16,19) | PUFA | 5.55 * | 30.10 | 913 |
| 18 | 7,10,13,16,19-Docosapentaenoic acid | C22:5 (7,10,13,16,19) | PUFA | 0.45 | 31.31 | 855 |
| 19 | Palmitoleic acid | C16:1 (7) | MUFA | 3.40 | 28.68 | 904 |
| 20 | Palmitoleic acid, 15-methyl- | C17:1 (7) | MUFA | 1.02 | 30.54 | 907 |
| 21 | Margaric acid | C17:0 | SFA | 3.47 | 31.23 | 908 |
| 22 | 10-Heptadecenoic acid | C17:1 (10) | MUFA | 1.04 | 32.41 | 830 |
| 23 | Stearic acid | C18:0 | SFA | 2.06 | 33.20 | 914 |
| 24 | Vaccenic acid | C18:1 (7) | MUFA | 1.71 | 32.01 | 910 |
| 25 | Oleic acid | C18:1 (9) | MUFA | 4.99* | 32.51 | 925 |
| 26 | Nonadecanoic acid | C19:0 | SFA | 0.67 | 33.02 | 815 |
| 27 | Arachidic acid | C20:0 | SFA | 1.47 | 36.46 | 904 |
| 28 | Behenic acid | C22:0 | SFA | 0.92 | 39.86 | 899 |
| 29 | Cholesterol | C27:1 (5) | Sterol | 3.99 | 34.38 | 913 |
| 30 | Cholestan-3-ol, 2-methylene-, (3α,5α)- | C28:1 (5) | Sterol | 0.16 | 35.38 | 797 |
| 31 | Heptacosane | C27:0 | SHC | 3.75 | 37.61 | 877 |
| 32 | Dotriacontane | C32:0 | SHC | 1.68 | 43.35 | 851 |
| 33 | Cholesterol margarate | C44:1 (5) | Sterol | 0.30 | 43.83 | 812 |
| 34 | Cholesta-4,6-dien-3-ol, (3α)- | C27:1 (5) | Sterol | 0.51 | 43.92 | 890 |
| 35 | Cholesta-3,5-diene | C27:2 (3,5) | Sterol | 1.15 | 44.08 | 889 |
| SFA | 25.33% | |||||
| MUFA | 12.16% | |||||
| PUFA | 22.08% | |||||
| SHC | 5.43% | |||||
| Sterol | 6.11 % | |||||
| Total | 71.11% | |||||
| Compound | COX-2 (µM) | COX-1 (µM) | COX-2/COX-1 |
|---|---|---|---|
| 1 | >100 | >100 | - |
| 2 | >100 | >100 | - |
| 3 | >100 | >100 | - |
| 4 | 2.41 ± 0.2 * | 18.5 ± 0.4 * | 0.13 |
| 5 | >100 | >100 | - |
| 6 | >100 | >100 | - |
| Crude oil | 15.27 ± 0.3 * | 46.33 ± 0.2 * | 0.33 |
| Celecoxib® | 0.125 ± 0.2 * | 2.53 ± 0.3 * | 0.05 |
| Gene Name | GenBank Accession | ||
|---|---|---|---|
| IL-β1 | NC_013670.1 | Forward | 5′-AGCTTCTCCAGAGCCACAAC-3′ |
| Reverse | 5′-CCTGACTACCCTCACGCACC-3′ | ||
| GAPDH | NC_013676.1 | Forward | 5′-GTCAAGGCTGAGAACGGGAA-3′ |
| Reverse | 5′-ACAAGAGAGTTGGCTGGGTG-3′ | ||
| TGF-β1 | NC_013672.1 | Forward | 5′-GACTGTGCGTTTTGGGTTCC-3′ |
| Reverse | 5′-CCTGGGCTCCTCCTAGAGTT-3′ | ||
| TNF-α | NC_013680.1 | Forward | 5′-GAGAACCCCACGGCTAGATG-3′ |
| Reverse | 5′-TTCTCCAACTGGAAGACGCC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsenani, F.; Ashour, A.M.; Alzubaidi, M.A.; Azmy, A.F.; Hetta, M.H.; Abu-Baih, D.H.; Elrehany, M.A.; Zayed, A.; Sayed, A.M.; Abdelmohsen, U.R.; et al. Wound Healing Metabolites from Peters’ Elephant-Nose Fish Oil: An In Vivo Investigation Supported by In Vitro and In Silico Studies. Mar. Drugs 2021, 19, 605. https://doi.org/10.3390/md19110605
Alsenani F, Ashour AM, Alzubaidi MA, Azmy AF, Hetta MH, Abu-Baih DH, Elrehany MA, Zayed A, Sayed AM, Abdelmohsen UR, et al. Wound Healing Metabolites from Peters’ Elephant-Nose Fish Oil: An In Vivo Investigation Supported by In Vitro and In Silico Studies. Marine Drugs. 2021; 19(11):605. https://doi.org/10.3390/md19110605
Chicago/Turabian StyleAlsenani, Faisal, Ahmed M. Ashour, Mubarak A. Alzubaidi, Ahmed F. Azmy, Mona H. Hetta, Dalia H. Abu-Baih, Mahmoud A. Elrehany, Ahmed Zayed, Ahmed M. Sayed, Usama Ramadan Abdelmohsen, and et al. 2021. "Wound Healing Metabolites from Peters’ Elephant-Nose Fish Oil: An In Vivo Investigation Supported by In Vitro and In Silico Studies" Marine Drugs 19, no. 11: 605. https://doi.org/10.3390/md19110605
APA StyleAlsenani, F., Ashour, A. M., Alzubaidi, M. A., Azmy, A. F., Hetta, M. H., Abu-Baih, D. H., Elrehany, M. A., Zayed, A., Sayed, A. M., Abdelmohsen, U. R., & Elmaidomy, A. H. (2021). Wound Healing Metabolites from Peters’ Elephant-Nose Fish Oil: An In Vivo Investigation Supported by In Vitro and In Silico Studies. Marine Drugs, 19(11), 605. https://doi.org/10.3390/md19110605