
Deep-Sea Anemones Are Prospective Source of New Antimicrobial and Cytotoxic Compounds

 , ,
, ,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Results
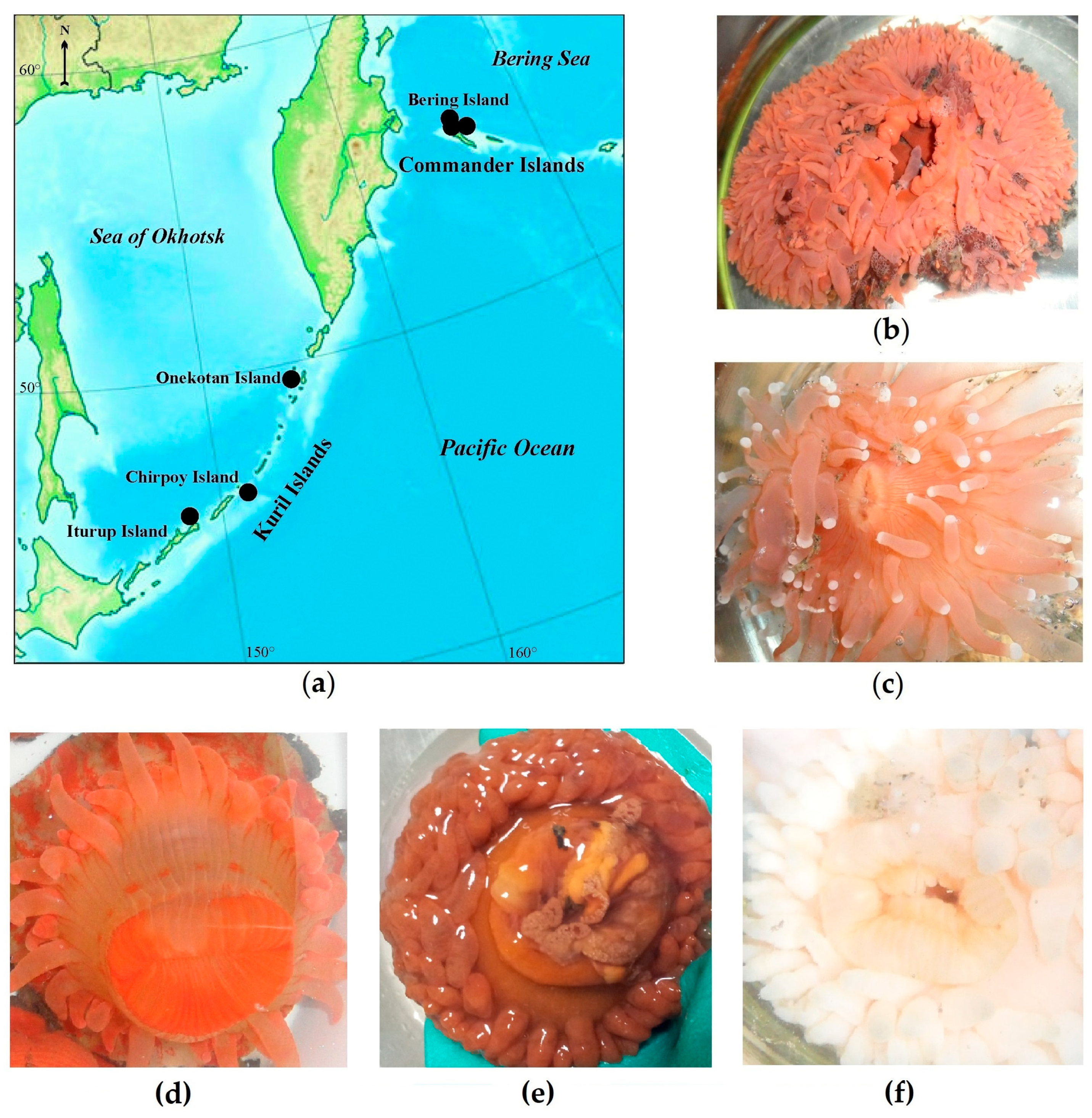
2.1. Identification of Deep-Sea Anemone Species
2.2. Sea Anemone Tentacle Extraction
2.3. Determination of Biological Activity of Sea Anemone Extracts
2.3.1. Hemolytic Activity
2.3.2. Cytotoxic Activity
2.3.3. Antimicrobial Activity
2.3.4. Enzyme-Inhibiting Activity
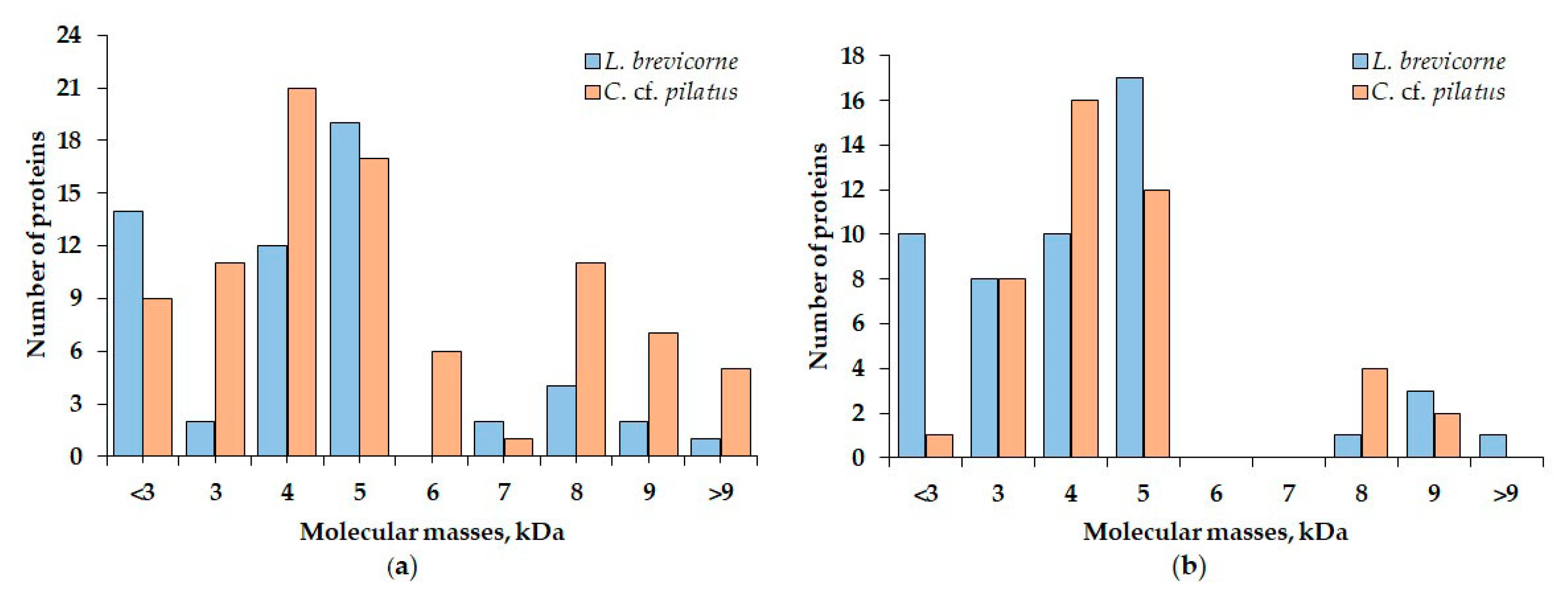
2.4. Identification of Peptide Compositions in L. brevicorne and C. cf. pilatus Extracts
3. Discussion
4. Materials and Methods
4.1. Sea Anemone Collection
4.2. DNA Extraction, Amplification, and Sequencing
4.3. Phylogenetic Analysis
4.4. Preparation of Aqueous and Ethanol Extracts
4.5. Determination of Protein Concentration
4.6. Trypsin Inhibition Assay
4.7. Assay of Enzymatic Activity of Recombinant α-Galactosidase and Its Mutant Form from Marine Bacterium Pseudoalteromonas KMM 701
4.8. Hemolytic Activity Assay
4.9. Assay of Cytotoxicity Effect against Murine Ascite Ehrlich Carcinoma Cells In Vitro
4.10. Assay of Cytotoxic Effect against Murine Spleen Cells
4.11. Antimicrobial Assay
4.12. Size-Exclusion Chromatography
4.13. Mass Spectrometry Analysis
4.14. Sample Preparation for Proteomic Analysis
4.15. Tandem Mass Spectrometry Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Bull, A.T.; Ward, A.C.; Goodfellow, M. Search and discovery strategies for biotechnology: The paradigm shift. Microbiol. Mol. Biol. Rev. 2000, 64, 573–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, P.C.; Westacott, R.E.; Burja, A.M. Piezotolerance as a metabolic engineering tool for the biosynthesis of natural products. Biomol. Eng. 2003, 20, 325–331. [Google Scholar] [CrossRef]
- Beliaev, G.M. Deep-Sea Ocean Trenches and Their Fauna; Vinogradov, M.Y., Ed.; Nauka Publishing House: Moscow, Russia, 1989; ISBN 5020052760. [Google Scholar]
- Jungo, F.; Bougueleret, L.; Xenarios, I.; Poux, S. The UniProtKB/Swiss-Prot Tox-Prot program: A central hub of integrated venom protein data. Toxicon 2012, 60, 551–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maček, P. Polypeptide cytolytic toxins from sea anemones (Actiniaria). FEMS Microbiol. Immunol. 1992, 105, 121–130. [Google Scholar] [CrossRef]
- Klyshko, E.V.; Issaeva, M.P.; Monastyrnaya, M.M.; Il’yna, A.P.; Guzev, K.V.; Vakorina, T.I.; Dmitrenok, P.S.; Zykova, T.A.; Kozlovskaya, E.P. Isolation, properties and partial amino acid sequence of a new actinoporin from the sea anemone Radianthus macrodactylus. Toxicon 2004, 44, 315–324. [Google Scholar] [CrossRef]
- Monastyrnaya, M.; Leychenko, E.; Isaeva, M.; Likhatskaya, G.; Zelepuga, E.; Kostina, E.; Trifonov, E.; Nurminski, E.; Kozlovskaya, E. Actinoporins from the sea anemones, tropical Radianthus macrodactylus and northern Oulactis orientalis: Comparative analysis of structure-function relationships. Toxicon 2010, 56, 1299–1314. [Google Scholar] [CrossRef]
- Rojko, N.; Dalla Serra, M.; MačEk, P.; Anderluh, G. Pore formation by actinoporins, cytolysins from sea anemones. Biochim. Biophys. Acta Biomembr. 2016, 1858, 446–456. [Google Scholar] [CrossRef]
- Leychenko, E.; Isaeva, M.; Tkacheva, E.; Zelepuga, E.; Kvetkina, A.; Guzev, K.; Margarita, M.; Kozlovskaya, E. Multigene family of pore-forming toxins from sea anemone Heteractis crispa. Mar. Drugs 2018, 16, 183. [Google Scholar] [CrossRef] [Green Version]
- Grotendorst, G.R.; Hessinger, D.A. Enzymatic characterization of the major phospholipase A2 component of sea anemone (Aiptasia pallida) nematocyst venom. Toxicon 2000, 38, 931–943. [Google Scholar] [CrossRef]
- Razpotnik, A.; Križaj, I.; Šribar, J.; Kordiš, D.; Maček, P.; Frangež, R.; Kem, W.R.; Turk, T. A new phospholipase A2 isolated from the sea anemone Urticina crassicornis—Its primary structure and phylogenetic classification. FEBS J. 2010, 277, 2641–2653. [Google Scholar] [CrossRef]
- Madio, B.; King, G.F.; Undheim, E.A.B. Sea anemone toxins: A structural overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef] [Green Version]
- Klyshko, E.V.; Il’ina, A.P.; Monastyrnaya, M.M.; Burtseva, Y.V.; Kostina, E.E.; Zykova, T.A.; Menzorova, N.I.; Kozlovskaya, E.P. Biologically active polypeptides and hydrolytic enzymes in sea anemones of northern temperate waters. Russ. J. Mar. Biol. 2003, 29, 161–166. [Google Scholar] [CrossRef]
- Mourão, C.B.F.; Schwartz, E.F. Protease inhibitors from marine venomous animals and their counterparts in terrestrial venomous animals. Mar. Drugs 2013, 11, 2069–2112. [Google Scholar] [CrossRef] [Green Version]
- Anderluh, G.; Maček, P. Cytolytic peptide and protein toxins from sea anemones (Anthozoa: Actinaria). Toxicon 2002, 40, 111–124. [Google Scholar] [CrossRef]
- Monastyrnaya, M.M.; Zykova, T.A.; Apalikova, O.V.; Shwets, T.V.; Kozlovskaya, E.P. Biologically active polypeptides from the tropical sea anemone Radianthus macrodactylus. Toxicon 2002, 40, 1197–1217. [Google Scholar] [CrossRef]
- Stabili, L.; Schirosi, R.; Parisi, M.G.; Piraino, S.; Cammarata, M. The mucus of Actinia equina (Anthozoa, Cnidaria): An unexplored resource for potential applicative purposes. Mar. Drugs 2015, 13, 5276–5296. [Google Scholar] [CrossRef] [Green Version]
- Kvetkina, A.; Malyarenko, O.; Pavlenko, A.; Dyshlovoy, S.; von Amsberg, G.; Ermakova, S.; Leychenko, E. Sea anemone Heteractis crispa actinoporin demonstrates in vitro anticancer activities and prevents HT-29 colorectal cancer cell migration. Molecules 2020, 25, 5979. [Google Scholar] [CrossRef]
- Avila, A.D.; Mateo de Acosta, C.; Lage, A. A carcinoembryonic antigen-directed immunotoxin built by linking a monoclonal antibody to a hemolytic toxin. Int. J. Cancer 1989, 43, 926–929. [Google Scholar] [CrossRef]
- Pederzolli, C.; Belmonte, G.; Serra, M.D.; Menestrina, G.; Maček, P. Biochemical and cytotoxic properties of conjugates of transferrin with equinatoxin II, a cytolysin from a sea anemone. Bioconjug. Chem. 1995, 6, 166–173. [Google Scholar] [CrossRef]
- Potrich, C.; Anderluh, G.; Macek, P. Construction of new immunotoxins by linking equinatoxin II to monoclonal antibodies via the biotin-avidin interaction. Cytotoxic effects on human tumor cells. Acta Biol. Slov. 2000, 43, 47–51. [Google Scholar]
- Álvarez, C.; Mancheño, J.M.; Martínez, D.; Tejuca, M.; Pazos, F.; Lanio, M.E. Sticholysins, two pore-forming toxins produced by the Caribbean sea anemone Stichodactyla helianthus: Their interaction with membranes. Toxicon 2009, 54, 1135–1147. [Google Scholar] [CrossRef]
- Lv, X.; Zhang, J.; Xu, R.; Dong, Y.; Sun, A.; Shen, Y.; Wei, D. Gigantoxin-4-4D5 scFv is a novel recombinant immunotoxin with specific toxicity against HER2/neu-positive ovarian carcinoma cells. Appl. Microbiol. Biotechnol. 2016, 100, 6403–6413. [Google Scholar] [CrossRef]
- Mutter, N.L.; Soskine, M.; Huang, G.; Albuquerque, I.S.; Bernardes, G.J.L.; Maglia, G. Modular pore-forming immunotoxins with caged cytotoxicity tailored by directed evolution. ACS Chem. Biol. 2018, 13, 3153–3160. [Google Scholar] [CrossRef]
- Norton, R.; Pennington, M.; Wulff, H. Potassium channel blockade by the sea anemone toxin ShK for the treatment of multiple sclerosis and other autoimmune diseases. Curr. Med. Chem. 2004, 11, 3041–3052. [Google Scholar] [CrossRef]
- Honma, T.; Shiomi, K. Peptide toxins in sea anemones: Structural and functional aspects. Mar. Biotechnol. 2006, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Schweitz, H.; Bruhn, T.; Guillemare, E.; Moinier, D.; Lancelin, J.; Lazdunski, M. Kalicludines and Kaliseptine. Two different classes of sea anemone toxins for voltage-sensitive K+ channels. J. Biol. Chem. 1995, 270, 25121–25126. [Google Scholar] [CrossRef] [Green Version]
- García-Fernández, R.; Peigneur, S.; Pons, T.; Alvarez, C.; González, L.; Chávez, M.A.; Tytgat, J. The Kunitz-type protein ShPI-1 inhibits serine proteases and voltage-gated potassium channels. Toxins 2016, 8, 110. [Google Scholar] [CrossRef] [Green Version]
- Gladkikh, I.; Peigneur, S.; Sintsova, O.; Pinheiro-Junior, E.L.; Klimovich, A.; Menshov, A.; Kalinovsky, A.; Isaeva, M.; Monastyrnaya, M.; Kozlovskaya, E.; et al. Kunitz-type peptides from the sea anemone Heteractis crispa demonstrate potassium channel blocking and anti-inflammatory activities. Biomedicines 2020, 8, 473. [Google Scholar] [CrossRef]
- Andreev, Y.A.; Kozlov, S.A.; Koshelev, S.G.; Ivanova, E.A.; Monastyrnaya, M.M.; Kozlovskaya, E.P.; Grishin, E.V. Analgesic compound from sea anemone Heteractis crispa is the first polypeptide inhibitor of vanilloid receptor 1 (TRPV1). J. Biol. Chem. 2008, 283, 23914–23921. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, S.A.; Andreev, Y.A.; Murashev, A.N.; Skobtsov, D.I.; D’iachenko, I.A.; Grishin, E.V. New polypeptide components from the Heteractis crispa sea anemone with analgesic activity. Russ. J. Bioorganic Chem. 2009, 35, 789–798. [Google Scholar] [CrossRef]
- Monastyrnaya, M.; Peigneur, S.; Zelepuga, E.; Sintsova, O.; Gladkikh, I.; Leychenko, E.; Isaeva, M.; Tytgat, J.; Kozlovskaya, E. Kunitz-Type peptide HCRG21 from the sea anemone Heteractis crispa is a full antagonist of the TRPV1 receptor. Mar. Drugs 2016, 14, 229. [Google Scholar] [CrossRef] [PubMed]
- Nikolaev, M.V.; Dorofeeva, N.A.; Komarova, M.S.; Korolkova, Y.V.; Andreev, Y.A.; Mosharova, I.V.; Grishin, E.V.; Tikhonov, D.B.; Kozlov, S.A. TRPV1 activation power can switch an action mode for its polypeptide ligands. PLoS ONE 2017, 12, e0177077. [Google Scholar] [CrossRef] [PubMed]
- Sintsova, O.V.; Monastyrnaya, M.M.; Pislyagin, E.A.; Menchinskaya, E.S.; Leychenko, E.V.; Aminin, D.L.; Kozlovskaya, E.P. Anti-inflammatory activity of a polypeptide from the Heteractis crispa sea anemone. Russ. J. Bioorganic Chem. 2015, 41, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Sintsova, O.V.; Pislyagin, E.A.; Gladkikh, I.N.; Monastyrnaya, M.M.; Menchinskaya, E.S.; Leychenko, E.V.; Aminin, D.L.; Kozlovskaya, E.P. Kunitz-type peptides of the sea anemone Heteractis crispa: Potential anti-inflammatory compounds. Russ. J. Bioorganic Chem. 2017, 43, 91–97. [Google Scholar] [CrossRef]
- Gladkikh, I.; Monastyrnaya, M.; Zelepuga, E.; Sintsova, O.; Tabakmakher, V.; Gnedenko, O.; Ivanov, A.; Hua, K.F.; Kozlovskaya, E.; Jacobson, P.B. New Kunitz-type HCRG polypeptides from the sea anemone Heteractis crispa. Mar. Drugs 2015, 13, 6038–6063. [Google Scholar] [CrossRef] [Green Version]
- Andreev, Y.A.; Kozlov, S.A.; Korolkova, Y.V.; Dyachenko, I.A.; Bondarenko, D.A.; Skobtsov, D.I.; Murashev, A.N.; Kotova, P.D.; Rogachevskaja, O.A.; Kabanova, N.V.; et al. Polypeptide modulators of TRPV1 produce analgesia without hyperthermia. Mar. Drugs 2013, 11, 5100–5115. [Google Scholar] [CrossRef] [Green Version]
- Tabakmakher, V.M.; Sintsova, O.V.; Krivoshapko, O.N.; Zelepuga, E.A.; Monastyrnaya, M.M.; Kozlovskaya, E.P. Analgesic effect of novel Kunitz-type polypeptides of the sea anemone Heteractis crispa. Dokl. Biochem. Biophys. 2015, 461, 80–83. [Google Scholar] [CrossRef]
- Sintsova, O.V.; Palikov, V.A.; Palikova, Y.A.; Klimovich, A.A.; Gladkikh, I.N.; Andreev, Y.A.; Monastyrnaya, M.M.; Kozlovskaya, E.P.; Dyachenko, I.A.; Kozlov, S.A.; et al. Peptide blocker of ion channel TRPV1 exhibits a long analgesic effect in the heat stimulation model. Dokl. Biochem. Biophys. 2020, 493, 215–217. [Google Scholar] [CrossRef]
- Kvetkina, A.N.; Leychenko, E.V.; Yurchenko, E.A.; Pislyagin, E.A.; Peigneur, S.; Tytgat, Y.; Isaeva, M.P.; Aminin, D.L.; Kozlovskaya, E.P. A New IQ-peptide of the Kunitz-type from the Heteractis magnifica sea anemone exhibits neuroprotective activity in a model of Alzheimer’s Disease. Russ. J. Bioorganic Chem. 2018, 44, 416–423. [Google Scholar] [CrossRef]
- Kvetkina, A.; Leychenko, E.; Chausova, V.; Zelepuga, E.; Chernysheva, N.; Guzev, K.; Pislyagin, E.; Yurchenko, E.; Menchinskaya, E.; Aminin, D.; et al. A new multigene HCIQ subfamily from the sea anemone Heteractis crispa encodes Kunitz-peptides exhibiting neuroprotective activity against 6-hydroxydopamine. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sintsova, O.; Gladkikh, I.; Monastyrnaya, M.; Tabakmakher, V.; Yurchenko, E.; Menchinskaya, E.; Pislyagin, E.; Andreev, Y.; Kozlov, S.; Peigneur, S.; et al. Sea anemone Kunitz-type peptides demonstrate neuroprotective activity in the 6-hydroxydopamine induced neurotoxicity model. Biomedicines 2021, 9, 283. [Google Scholar] [CrossRef]
- Diochot, S.; Baron, A.; Rash, L.D.; Deval, E.; Escoubas, P.; Scarzello, S.; Salinas, M.; Lazdunski, M. A new sea anemone peptide, APETx2, inhibits ASIC3, a major acid-sensitive channel in sensory neurons. EMBO J. 2004, 23, 1516–1525. [Google Scholar] [CrossRef]
- Osmakov, D.I.; Kozlov, S.A.; Andreev, Y.A.; Koshelev, S.G.; Sanamyan, N.P.; Sanamyan, K.E.; Dyachenko, I.A.; Bondarenko, D.A.; Murashev, A.N.; Mineev, K.S.; et al. Sea anemone peptide with uncommon β-hairpin structure inhibits acid-sensing ion channel 3 (ASIC3) and reveals analgesic activity. J. Biol. Chem. 2013, 288, 23116–23127. [Google Scholar] [CrossRef] [Green Version]
- Cristofori-Armstrong, B.; Rash, L.D. Acid-sensing ion channel (ASIC) structure and function: Insights from spider, snake and sea anemone venoms. Neuropharmacology 2017, 127, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Kalina, R.; Gladkikh, I.; Dmitrenok, P.; Chernikov, O.; Koshelev, S.; Kvetkina, A.; Kozlov, S.; Kozlovskaya, E.; Monastyrnaya, M. New APETx-like peptides from sea anemone Heteractis crispa modulate ASIC1a channels. Peptides 2018, 104, 41–49. [Google Scholar] [CrossRef]
- Logashina, Y.A.; Mosharova, I.V.; Korolkova, Y.V.; Shelukhina, I.V.; Dyachenko, I.A.; Palikov, V.A.; Palikova, Y.A.; Murashev, A.N.; Kozlov, S.A.; Stensvåg, K.; et al. Peptide from sea anemone Metridium senile affects transient receptor potential ankyrin-repeat 1 (TRPA1) function and produces analgesic effect. J. Biol. Chem. 2017, 292, 2992–3004. [Google Scholar] [CrossRef] [Green Version]
- Logashina, Y.A.; Solstad, R.G.; Mineev, K.S.; Korolkova, Y.V.; Mosharova, I.V.; Dyachenko, I.A.; Palikov, V.A.; Palikova, Y.A.; Murashev, A.N.; Arseniev, A.S.; et al. New disulfide-stabilized fold provides sea anemone peptide to exhibit both antimicrobial and TRPA1 potentiating properties. Toxins 2017, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Kim, J. Channelopathies. Korean J. Pediatr. 2014, 57, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Moran, Y.; Praher, D.; Schlesinger, A.; Ayalon, A.; Tal, Y.; Technau, U. Analysis of soluble protein contents from the nematocysts of a model sea anemone sheds light on venom evolution. Mar. Biotechnol. 2013, 15, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, M.L.; Hamilton, B.R.; Madio, B.; Morales, R.A.V.; Tonkin-Hill, G.Q.; Papenfuss, A.T.; Purcell, A.W.; King, G.F.; Undheim, E.A.B.; Norton, R.S. The use of imaging mass spectrometry to study peptide toxin distribution in Australian sea anemones. Aust. J. Chem. 2017, 70, 1235–1237. [Google Scholar] [CrossRef] [Green Version]
- Zaharenko, A.J.; Ferreira, W.A.; Oliveira, J.S.; Richardson, M.; Pimenta, D.C.; Konno, K.; Portaro, F.C.V.; de Freitas, J.C. Proteomics of the neurotoxic fraction from the sea anemone Bunodosoma cangicum venom: Novel peptides belonging to new classes of toxins. Comp. Biochem. Physiol.-Part D Genom. Proteom. 2008, 3, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.A.; Cassoli, J.S.; Sa, F.; Dong, Z.Q.; De Freitas, J.C.; Pimenta, A.M.C.; De Lima, M.E.; Konno, K.; Lee, S.M.Y.; Garateix, A.; et al. Peptide fingerprinting of the neurotoxic fractions isolated from the secretions of sea anemones Stichodactyla helianthus and Bunodosoma granulifera. New members of the APETx-like family identified by a 454 pyrosequencing approach. Peptides 2012, 34, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Rachamim, T.; Morgenstern, D.; Aharonovich, D.; Brekhman, V.; Lotan, T.; Sher, D. The dynamically evolving nematocyst content of an Anthozoan, a Scyphozoan, and a Hydrozoan. Mol. Biol. Evol. 2015, 32, 740–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sintsova, O.; Gladkikh, I.; Chausova, V.; Monastyrnaya, M.; Anastyuk, S.; Chernikov, O.; Yurchenko, E.; Aminin, D.; Isaeva, M.; Leychenko, E.; et al. Peptide fingerprinting of the sea anemone Heteractis magnifica mucus revealed neurotoxins, Kunitz-type proteinase inhibitors and a new β-defensin α-amylase inhibitor. J. Proteom. 2018, 173, 12–21. [Google Scholar] [CrossRef]
- Cassoli, J.S.; Verano-Braga, T.; Oliveira, J.S.; Montandon, G.G.; Cologna, C.T.; Peigneur, S.; Pimenta, A.M.; Kjeldsen, F.; Roepstorff, P.; Tytgat, J.; et al. The proteomic profile of Stichodactyla duerdeni secretion reveals the presence of a novel O-linked glycopeptide. J. Proteom. 2013, 87, 89–102. [Google Scholar] [CrossRef]
- D’Ambra, I.; Lauritano, C. A Review of toxins from Cnidaria. Mar. Drugs 2020, 18, 507. [Google Scholar] [CrossRef]
- Delfin, J.; Martinez, I.; Antuch, W.; Morera, V.; Gonzalez, Y.; Rodriguez, R.; Marquez, M.; Larionova, N.; Diaz, J.; Chavez, M.; et al. Purification, characterization and of proteinase inhibitors from Stichodactyla helianthus. Toxicon 1996, 34, 1367–1376. [Google Scholar] [CrossRef]
- Sokotun, I.N.; Il’ina, A.P.; Monastyrnaya, M.M.; Leychenko, E.V.; Es’kov, A.A.; Anastuk, S.D.; Kozlovskaya, E.P. Proteinase inhibitors from the tropical sea anemone Radianthus macrodactylus: Isolation and characteristic. Biochemistry 2007, 72, 301–306. [Google Scholar] [CrossRef]
- Bakunina, I.; Likhatskaya, G.; Slepchenko, L.; Balabanova, L.; Tekutyeva, L.; Son, O.; Shubina, L.; Makarieva, T. Effect of pentacyclic guanidine alkaloids from the sponge Monanchora pulchra on activity of α-glycosidases from marine bacteria. Mar. Drugs 2019, 17, 22. [Google Scholar] [CrossRef] [Green Version]
- Utkina, N.K.; Likhatskaya, G.N.; Balabanova, L.A.; Bakunina, I.Y. Sponge-derived polybrominated diphenyl ethers and dibenzo-p -dioxins, irreversible inhibitors of the bacterial α-d-galactosidase. Environ. Sci. Process. Impacts 2019, 21, 1754–1763. [Google Scholar] [CrossRef]
- Bakunina, I.; Slepchenko, L.; Anastyuk, S.; Isakov, V.; Likhatskaya, G.; Kim, N.; Tekutyeva, L.; Son, O.; Balabanova, L. Characterization of properties and transglycosylation abilities of recombinant α-galactosidase from cold-adapted marine bacterium Pseudoalteromonas KMM 701 and its C494N and D451A mutants. Mar. Drugs 2018, 16, 349. [Google Scholar] [CrossRef] [Green Version]
- Grebelnyi, S.D. Order Actiniaria-sea anemones. In List of Species of Free-Living Invertebrates of Eurasian Arctic Seas and Adjacent Deep Waters. Cnidaria and Ctenophora; Sirenko, B.I., Stepanjants, S.D., Eds.; Zoological Institute RAS: St. Petersburg, Russia, 2001; pp. 151–175. [Google Scholar]
- Eash-Loucks, W.E.; Fautin, D.G. Taxonomy and distribution of sea anemones (Cnidaria: Actiniaria and Corallimorpharia) from deep water of the northeastern pacific. Zootaxa 2012, 3375, 1–80. [Google Scholar] [CrossRef]
- Worthington Wilmer, J.; Mitchell, M.L. A preliminary investigation of the utility of ribosomal genes for species identification of sea anemones (Cnidaria: Actiniaria). In Proceedings of the Thirteenth International Marine Biological Workshop, The Marine Fauna and Flora of Moreton Bay, Brisbane, Australia, 1 December 2008; Volume 54, pp. 65–73. [Google Scholar]
- Il’ina, A.P.; Monastyrsnaia, M.M.; Sokotun, I.N.; Egorov, T.A.; Nazarenko, I.A.; Likhatskaia, G.N.; Kozlovskaia, E.P. Actinoporins from the Sea of Japan anemone Oulactis orientalis: Isolation and partial characterization. Russ. J. Bioorganic Chem. 2005, 31, 34–42. [Google Scholar] [CrossRef]
- Fedorov, S.; Dyshlovoy, S.; Monastyrnaya, M.; Shubina, L.; Leychenko, E.; Jin, J.O.; Kwak, J.Y.; Bode, A.M.; Dong, Z.; Stonik, V. The anticancer effects of actinoporin RTX-A from the sea anemone Heteractis crispa (=Radianthus macrodactylus). Toxicon 2010, 5, 811–817. [Google Scholar] [CrossRef] [Green Version]
- Bosch, T.C.G. The path less explored: Innate immune reactions in cnidarians. In Innate Immunity of Plants, Animals, and Humans. Nucleic Acids and Molecular Biology; Heine, H., Ed.; Springer: Heidelberg/Berlin, Germany, 2008; pp. 27–42. ISBN 9783540739302. [Google Scholar]
- Augustin, R.; Bosch, T. Cnidarian immunity: A tale of two barriers. In Invertebrate Immunity. Advances in Experimental Medicine and Biology; Söderhäll, K., Ed.; Springer: Boston, MA, USA, 2010; Volume 708, pp. 1–16. [Google Scholar]
- Trapani, M.R.; Parisi, M.G.; Toubiana, M.; Coquet, L.; Jouenne, T.; Roch, P.; Cammarata, M. First evidence of antimicrobial activity of neurotoxin 2 from Anemonia sulcata (Cnidaria). Invertebr. Surviv. J. 2014, 11, 182–191. [Google Scholar]
- Kim, C.H.; Lee, Y.J.; Go, H.J.; Oh, H.Y.; Lee, T.K.; Park, J.B.; Park, N.G. Defensin-neurotoxin dyad in a basally branching metazoan sea anemone. FEBS J. 2017, 284, 3320–3338. [Google Scholar] [CrossRef]
- Asano, N. Glycosidase inhibitors: Update and perspectives on practical use. Glycobiology 2003, 13, 93–104. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Kassell, B. Bovine trypsin-kallikrein inhibitor (Kunitz inhibitor, Basic Pancreatic Trypsin Inhibitor, polyvalent inhibitor from bovine organs). Methods Enzymol. 1970, 19, 844–852. [Google Scholar]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Andryjaschenko, P.V.; Dmitrenok, P.S.; Martyyas, E.A.; Kalinin, V.I. Triterpene glycosides from the sea cucumber Eupentacta fraudatrix. Structure and cytotoxic action of cucumariosides A2, A7, A9, A10, A11, A13 and A 14, sevennew minor non-sulfated tetraosides and an aglycone with an uncommon 18-hydroxy group. Nat. Prod. Commun. 2012, 7, 845–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heatley, N.G. A method for the assay of penicillin. Biochem. J. 1944, 38, 61–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geer, L.Y.; Markey, S.P.; Kowalak, J.A.; Wagner, L.; Xu, M.; Maynard, D.M.; Yang, X.; Shi, W.; Bryant, S.H. Open mass spectrometry search algorithm. J. Proteome Res. 2004, 3, 958–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine integrated into the MaxQuant environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef]
- Dorfer, V.; Strobl, M.; Winkler, S.; Mechtler, K. MS Amanda 2.0: Advancements in the standalone implementation. Rapid Commun. Mass Spectrom. 2021, 35, e9088. [Google Scholar] [CrossRef]
- Kim, S.; Pevzner, P.A. Universal database search tool for proteomics. Nat. Commun. 2016, 5, 5277. [Google Scholar] [CrossRef] [Green Version]
- Eng, J.K.; Jahan, T.A.; Hoopmann, M.R. Comet: An open-source MS/MS sequence database search tool. Proteomics 2013, 13, 22–24. [Google Scholar] [CrossRef]
- Barsnes, H.; Vaudel, M. SearchGUI: A highly adaptable common interface for proteomics search and de novo engines. J. Proteome Res. 2018, 17, 2552–2555. [Google Scholar] [CrossRef]
- Elias, J.E.; Gygi, S.P. Target-Decoy search strategy for mass spectrometry-based proteomics. Methods Mol. Biol. 2010, 604, 55–71. [Google Scholar] [CrossRef] [Green Version]
- Vaudel, M.; Burkhart, J.M.; Zahedi, R.P.; Oveland, E.; Berven, F.S.; Sickmann, A.; Martens, L.; Barsnes, H. PeptideShaker enables reanalysis of MS-derived proteomics data sets. Nat. Biotechnol. 2015, 33, 22–24. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sea Anemone | Habitat | Protein, mg | |
|---|---|---|---|
| Aqueous Extract | Ethanol Extract | ||
| Order Actiniaria Family Actinostolidae | |||
| Stomphia coccinea | The Sea of Okhotsk, Iturup Island, 45°44.4′ N, 148°33.4′ E, 263 m, gravel | 8.5 | 14.5 |
| Actinostola callosa | The Bering Sea, Bering Island, 55°25.2′ N, 165°49.8′ E, 207 m, silt | 7.6 | 2.6 |
| Actinostola faeculenta | The Bering Sea, Bering Island, 55°25.2′ N, 165°49.8′ E, 207 m, silt | 8.3 | 9.3 |
| Family Liponematidae | |||
| Liponema brevicorne | The Sea of Okhotsk, Onekotan Island, 49°24.1′ N, 154°16.1′ E, 146 m, sand, shells | 4.6 | 4.0 |
| Liponema brevicorne | The Bering Sea, Bering Island, 55°18.5′ N, 166°31.4′ E, 153 m, silt | 12.3 | 5.1 |
| Order Corallimorpharia Family Corallimorphidae | |||
| Corallimorphus cf. pilatus | The Pacific Ocean, Chirpoy Island, 46°21.1′ N, 150°59.0′ E, 455 m, gravel | 10.0 | 9.0 |
| Corallimorphus cf. pilatus | The Bering Sea, Bering Island, 55°25.7′ N, 165°49.4′ E, 289 m, silt over rocky ground | 8.9 | 3.1 |
| Species | Hemolytic Activity, MC100 ± SE | Cytotoxic Activity | Antimicrobial Activity, MC10 ± SE | |||
|---|---|---|---|---|---|---|
| Murine Splenocytes, MC70 ± SE, μg | Ehrlich Carcinoma Cells, MC70 ± SE, μg | B | S | C | ||
| Stomphia coccinea | 15.3 ± 0.05/– | –/– | 15.3 ± 0.09/14.5 ± 0.10 * | 8.5 ± 0.03/– | 8.5 ± 0.06/– | 8.5 ± 0.12/– |
| Actinostola callosa | 7.6 ± 0.12/– | –/– | 7.6 ± 0.04 **/7.5 ± 0.06 * | –/– | –/12.5 ± 0.09 | –/– |
| Actinostola faeculenta | 35.5 ± 0.01/– | –/9.3 ± 0.02 | 35.5 ± 0.8 */9.3 ± 0.05 * | –/– | –/– | –/– |
| Liponema brevicorne (Kuril Islands) | 8.8 ± 0.04/– | 8.8 ± 0.09 */– | –/– | 4.6 ± 0.08/– | –/– | –/– |
| Liponema brevicorne (Commander Islands) | –/7.6 ± 0.12 | –/– | –/– | –/– | –/– | –/– |
| Corallimorphus cf. pilatus (Kuril Islands) | 15.0 ± 0.04/45.0 ± 0.08 | –/– | 15.0 ± 0.25 **/9.0 ± 0.08 | 10.0 ± 0.04/– | 10.0 ± 0.08/– | 10.0 ± 0.03/– |
| Corallimorphus cf. pilatus (Commander Islands) | –/– | –/– | –/31.0 ± 0.06 | –/– | –/– | –/– |
| Species | Residual Activity, % ± SE | |||
|---|---|---|---|---|
| Ethanol Extract | Aqueous Extract | |||
| α-galactosidase | α-galactosidase C494N | α-galactosidase | α-galactosidase C494N | |
| Stomphia coccinea | 42 ± 1.3 | 100 ± 0.03 | 147 ± 0.65 | N.d. |
| Actinostola callosa | 45 ± 0.8 | 50 ± 1.8 | 174 ± 0.9 | N.d. |
| Actinostola faeculenta | 20 ± 1.12 | 30 ± 1.04 | 148 ± 1.1 | N.d. |
| Liponema brevicorne (Kuril Islands) | 6 ± 1.0 | N.d. | 126 ± 0.95 | N.d. |
| Liponema brevicorne (Commander Islands) | 113 ± 0.93 | N.d. | 115 ± 0.78 | N.d. |
| Corallimorphus cf. pilatus (Kuril Islands) | 37 ± 1.6 | 50 ± 0.18 | 170 ± 0.92 | N.d. |
| Corallimorphus cf. pilatus (Commander Islands) | 61 ± 1.37 | N.d. | 117 ± 1.2 | N.d. |
| Fractions | Peptide Molecular Masses Found in Aqueous Extracts, Da | Peptide Molecular Masses Found in Ethanol Extracts, Da | ||
|---|---|---|---|---|
| L. brevicorne | C. cf. pilatus | L. brevicorne | C. cf. pilatus | |
| 1 | 4050, 4221, 4504 5545 7864 8006 9884 10,395 | 2277 3264, 3391, 3582, 3835 4102, 4336, 4542, 4882 5153, 5539 6135, 6763, 6973 8241, 8393, 8548, 8896 9023, 9095, 9805 | 1527 2982 3052, 3182, 3403, 3947 4034, 4477, 4850 5053, 5274, 5381, 5522, 5934 8603 9785 | 2984 3075, 3586, 3780, 3865 4101, 4186, 4314, 4430, 4542, 4615, 4770, 4882 5151, 5410, 5769 8486, 8564, 8779, 8897 9121, 9808 |
| 2 | 2054, 2868, 2982 3198, 3663 4122, 4253, 4501 5111, 5415, 5970 7468 8322, 8469, 8600 | 3585, 3781, 3888 4030, 4337, 4444, 4543, 4882 5153, 5774, 5890 6136, 6370 8177, 8288, 8470, 8577, 8690, 8779, 8898 9100, 9812 | 2885, 2890 3829 4044, 4725, 4955, 4960 5091, 5182, 5267, 5271, 5406, 5547, 5549, 5752 9124 10,607, 10,846 | 3053, 3273, 3570, 3781 4021, 4086, 4187, 4306, 4457, 4560, 4600, 4743 5317, 5378, 5463, 5555, 5659, 5796, 5886 |
| 3 | 1458, 1687, 1950 2186, 2402, 2618, 2639, 27,132,789 3257, 3681, 3783, 3787, 3957 4078, 4132, 4242, 4725, 4780, 4806, 4956, 4970 5188, 5190, 5275, 5309, 5316, 5403, 5415, 5547, 5554, 5749, 5757, 5969 9768 | 1298, 1491 2123, 2978 3026, 3144, 3342, 3695, 3779 4020, 4023, 4186, 4302, 4376, 4383, 4539, 4542, 4873 5184, 5272, 5275, 5552, 5773, 5787, 5888, 5948 5959 | 1517, 1737, 1969 2215, 2492, 2970 3196, 3449, 3841 4039, 4727, 4944 5307, 5550, 5774 | 5546, 5760 |
| 4 | 1482, 1723, 1986 2214, 2455, 2719, 2971 3191 4784 5548, 5765, 5884 | 1520, 1720, 1993 2525 3974 4125, 4542, 4738 5231, 5374 6025, 6946 7608 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kvetkina, A.; Kostina, E.; Gladkikh, I.; Chausova, V.; Yurchenko, E.; Bakunina, I.; Pivkin, M.; Anastyuk, S.; Popov, R.; Monastyrnaya, M.; et al. Deep-Sea Anemones Are Prospective Source of New Antimicrobial and Cytotoxic Compounds. Mar. Drugs 2021, 19, 654. https://doi.org/10.3390/md19120654
Kvetkina A, Kostina E, Gladkikh I, Chausova V, Yurchenko E, Bakunina I, Pivkin M, Anastyuk S, Popov R, Monastyrnaya M, et al. Deep-Sea Anemones Are Prospective Source of New Antimicrobial and Cytotoxic Compounds. Marine Drugs. 2021; 19(12):654. https://doi.org/10.3390/md19120654
Chicago/Turabian StyleKvetkina, Aleksandra, Elena Kostina, Irina Gladkikh, Victoria Chausova, Ekaterina Yurchenko, Irina Bakunina, Mikhail Pivkin, Stanislav Anastyuk, Roman Popov, Margarita Monastyrnaya, and et al. 2021. "Deep-Sea Anemones Are Prospective Source of New Antimicrobial and Cytotoxic Compounds" Marine Drugs 19, no. 12: 654. https://doi.org/10.3390/md19120654
APA StyleKvetkina, A., Kostina, E., Gladkikh, I., Chausova, V., Yurchenko, E., Bakunina, I., Pivkin, M., Anastyuk, S., Popov, R., Monastyrnaya, M., Kozlovskaya, E., Isaeva, M., Dmitrenok, P., & Leychenko, E. (2021). Deep-Sea Anemones Are Prospective Source of New Antimicrobial and Cytotoxic Compounds. Marine Drugs, 19(12), 654. https://doi.org/10.3390/md19120654