The Marine-Derived Triterpenoid Frondoside A Inhibits Thrombus Formation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
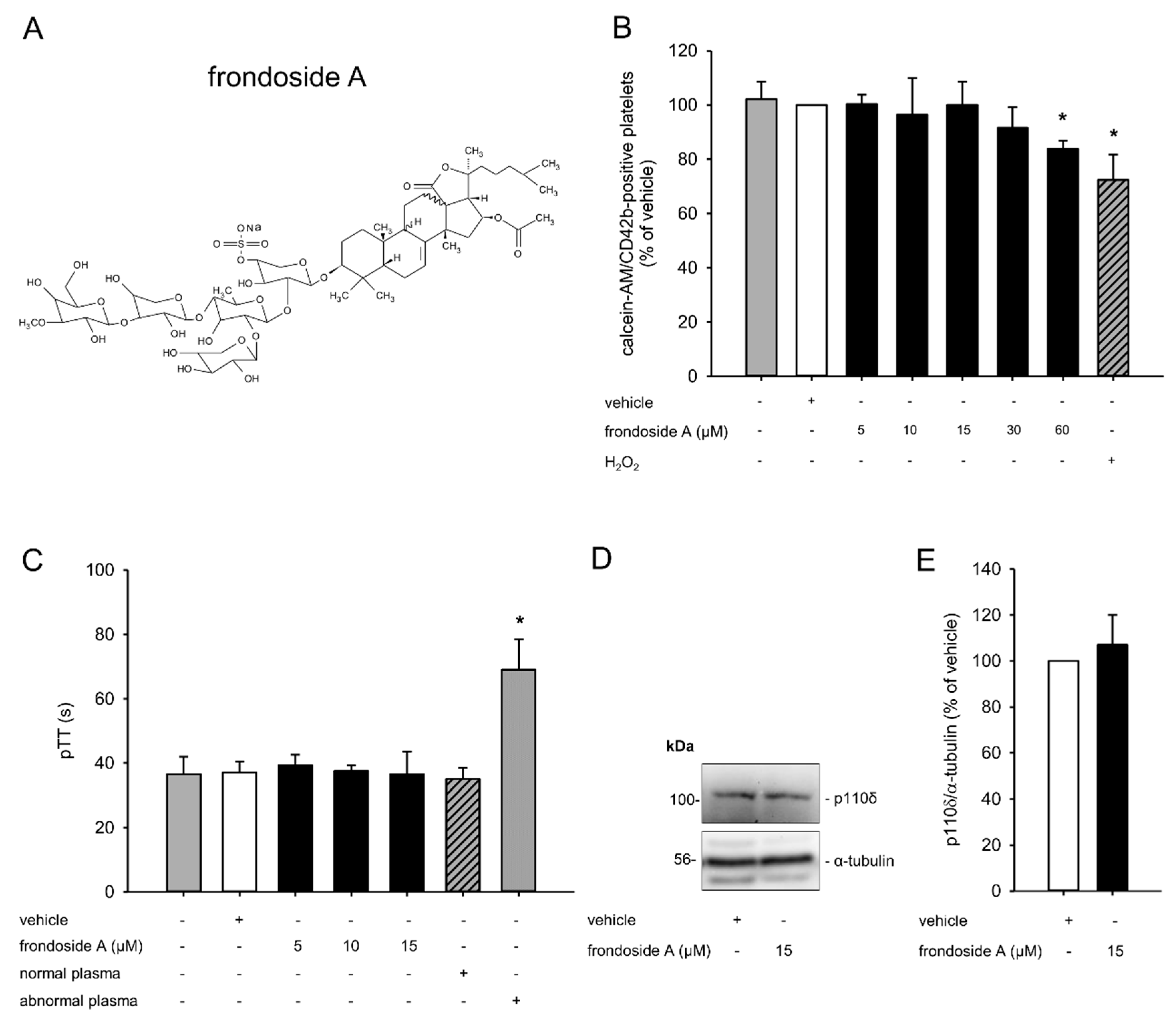
2.1. Effect of Frondoside A on Platelet Viability
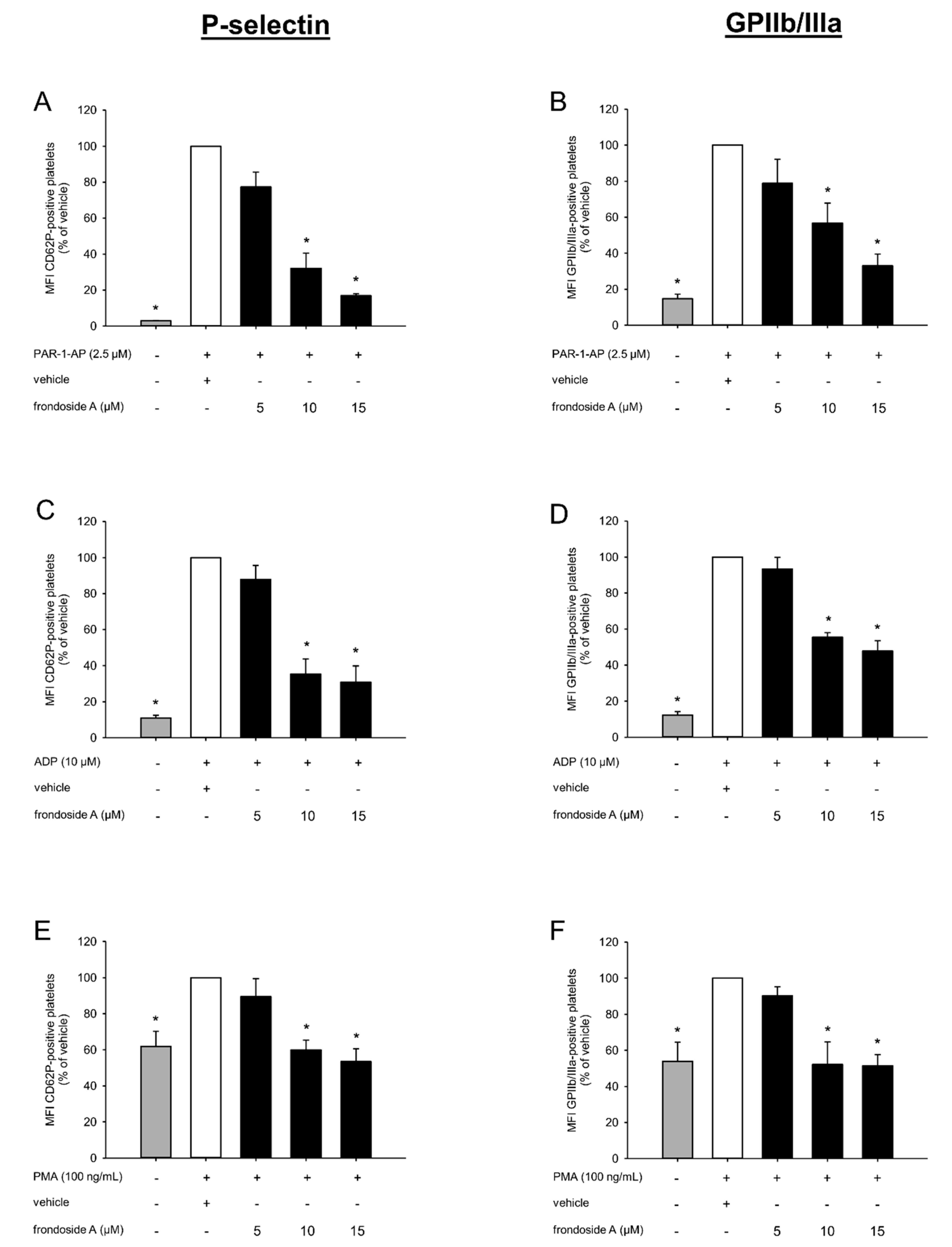
2.2. Effect of Frondoside A on Platelet Activation
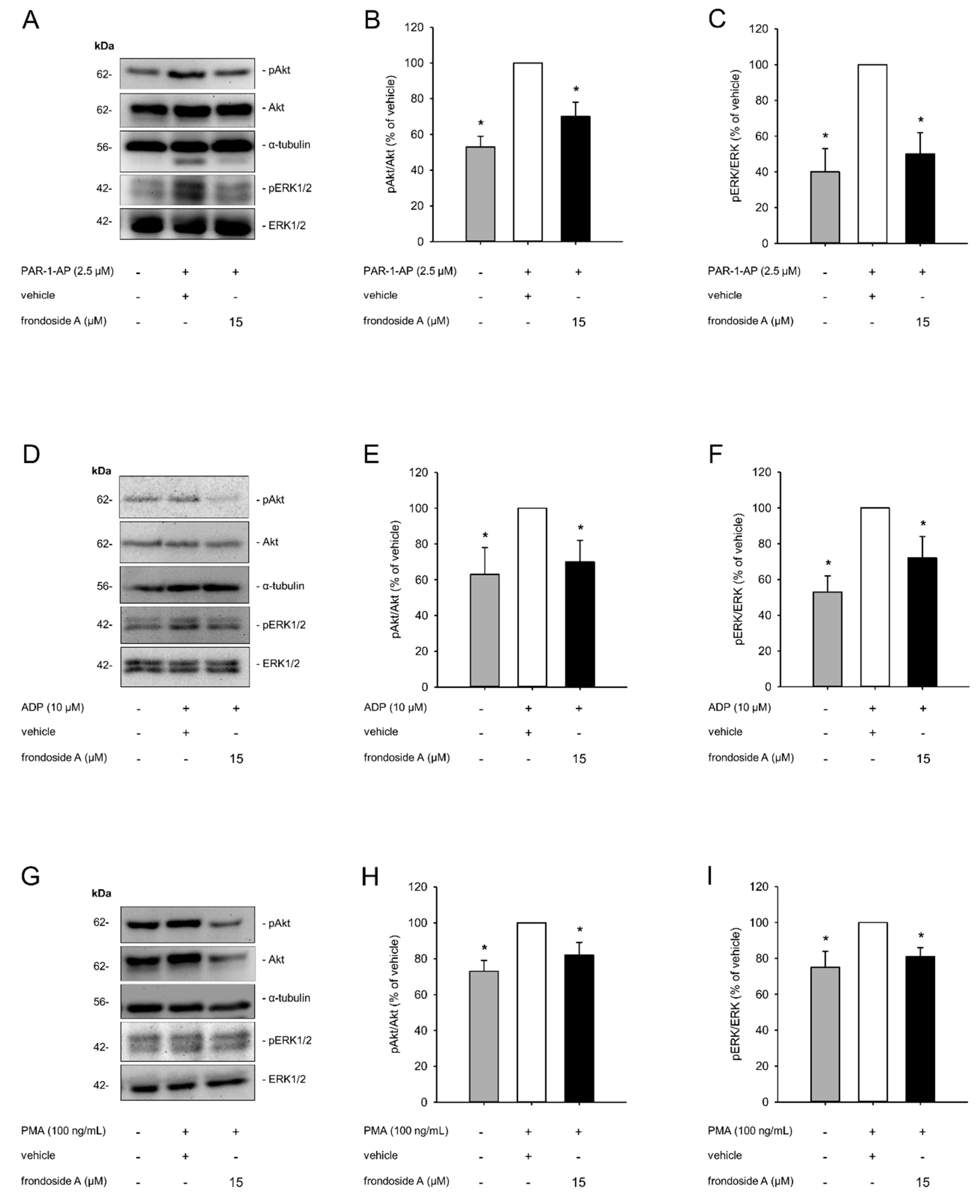
2.3. Effect of Frondoside A on PI3K Signaling
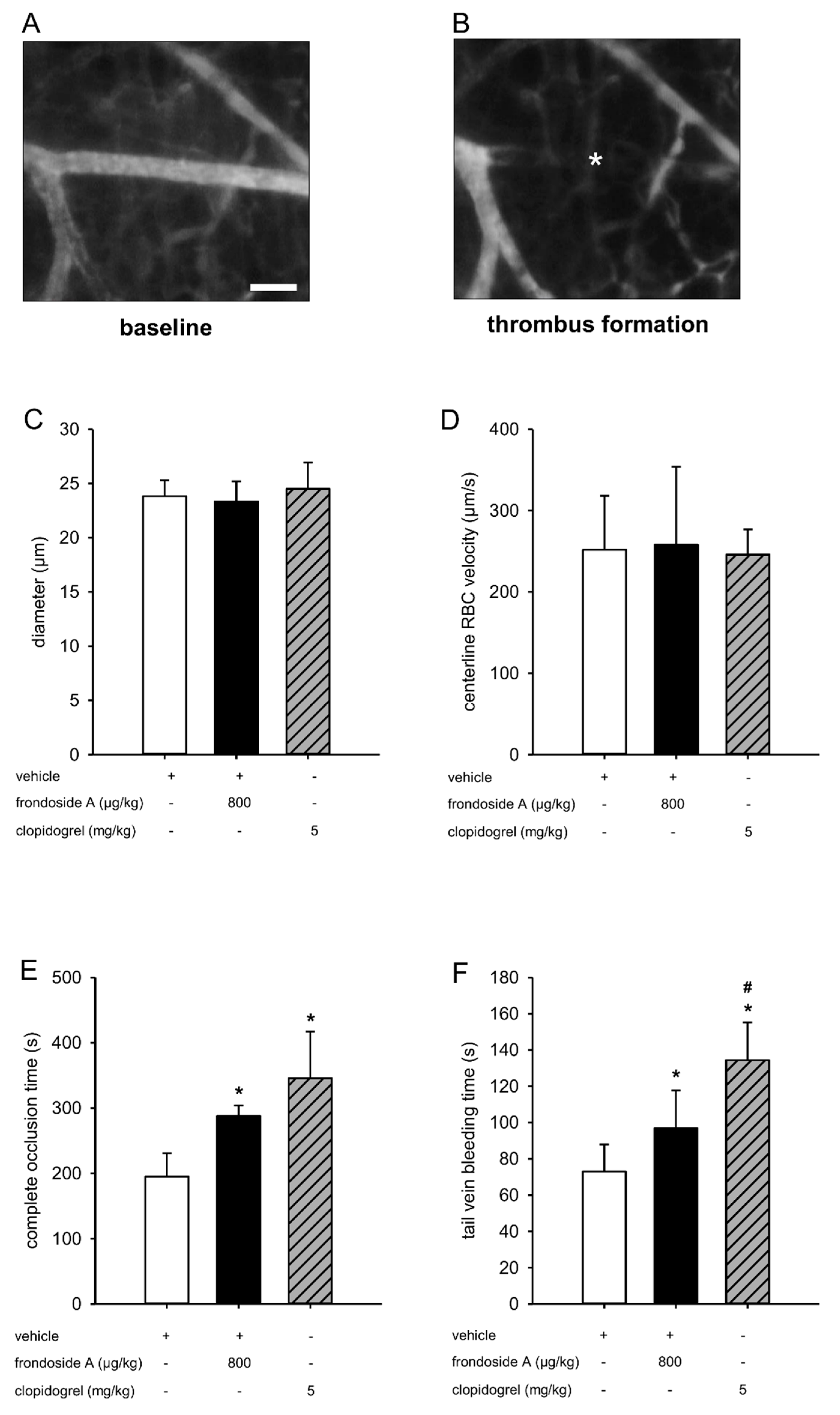
2.4. Effect of Frondoside A on Thrombus Formation In Vivo
3. Discussion
4. Materials and Methods
4.1. Chemicals, Reagents, and Antibodies
4.2. Ethics Statement
4.3. Preparation of PRP and Platelet Poor Plasma (PPP)
4.4. Viability Tests
4.5. Assessment of pTT
4.6. Western Blot Analysis
4.7. Analysis of Surface Adhesion Protein Expression
4.8. Animals
4.9. Photochemically Induced Thrombus Formation
4.10. Tail Vein Bleeding Time
4.11. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kapil, N.; Datta, Y.H.; Alakbarova, N.; Bershad, E.; Selim, M.; Liebeskind, D.S.; Bachour, O.; Rao, G.H.R.; Divani, A.A. Antiplatelet and Anticoagulant Therapies for Prevention of Ischemic Stroke. Clin. Appl. Thromb. Hemost. 2017, 23, 301–318. [Google Scholar] [CrossRef] [PubMed]
- Eikelboom, J.W.; Hirsh, J.; Spencer, F.A.; Baglin, T.P.; Weitz, J.I. Antiplatelet drugs: Antithrombotic Therapy and Prevention of Thrombosis, 9th ed: American College of Chest Physicians Evidence-Based Clinical Practice Guidelines. Chest 2012, 141, e89S–e119S. [Google Scholar] [CrossRef]
- Guidetti, G.F.; Canobbio, I.; Torti, M. PI3K/Akt in platelet integrin signaling and implications in thrombosis. Adv. Biol. Regul. 2015, 59, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Gratacap, M.P.; Guillermet-Guibert, J.; Martin, V.; Chicanne, G.; Tronchère, H.; Gaits-Iacovoni, F.; Payrastre, B. Regulation and roles of PI3Kβ, a major actor in platelet signaling and functions. Adv. Enzyme Regul. 2011, 51, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Wu, S.Y.; Liao, C.Y.; Teng, C.M.; Wu, Y.C.; Kuo, S.C. The roles and mechanisms of PAR4 and P2Y12/phosphatidylinositol 3-kinase pathway in maintaining thrombin-induced platelet aggregation. Br. J. Pharmacol. 2010, 161, 643–658. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.J. Mechanisms of Platelet Activation and Integrin αIIβ3. Korean Circ. J. 2012, 42, 295–301. [Google Scholar] [CrossRef]
- Nachman, R.L.; Leung, L.L. Complex formation of platelet membrane glycoproteins IIb and IIIa with fibrinogen. J. Clin. Investig. 1982, 69, 263–269. [Google Scholar] [CrossRef]
- Plow, E.F.; D’Souza, S.E.; Ginsberg, M.H. Ligand binding to GPIIb-IIIa: A status report. Semin. Thromb. Hemost. 1992, 18, 324–332. [Google Scholar] [CrossRef]
- Davenpeck, K.L.; Brummet, M.E.; Hudson, S.A.; Mayer, R.J.; Bochner, B.S. Activation of human leukocytes reduces surface P-selectin glycoprotein ligand-1 (PSGL-1, CD162) and adhesion to P-selectin in vitro. J. Immunol. 2000, 165, 2764–2772. [Google Scholar] [CrossRef]
- Li, Z.; Delaney, M.K.; O’Brien, K.A.; Du, X. Signaling during platelet adhesion and activation. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2341–2349. [Google Scholar] [CrossRef]
- Girard, M.; Bélanger, J.; ApSimon, J.W.; Garneau, F.X.; Harvey, C.; Brisson, J.R.; Frondoside, A. A novel triterpene glycoside from the holothurian Cucumariafrondosa. Can. J. Chem. 1990, 68, 11–18. [Google Scholar] [CrossRef]
- Aminin, D.L.; Menchinskaya, E.S.; Pisliagin, E.A.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I. Anticancer activity of sea cucumber triterpene glycosides. Mar. Drugs. 2015, 13, 1202–1223. [Google Scholar] [CrossRef] [PubMed]
- Al Shemaili, J.; Mensah-Brown, E.; Parekh, K.; Thomas, S.A.; Attoub, S.; Hellman, B.; Nyberg, F.; Adem, A.; Collin, P.; Adrian, T.E. Frondoside A enhances the antiproliferative effects of gemcitabine in pancreatic cancer. Eur. J. Cancer 2014, 50, 1391–1398. [Google Scholar] [CrossRef] [PubMed]
- Dyshlovoy, S.A.; Menchinskaya, E.S.; Venz, S.; Rast, S.; Amann, K.; Hauschild, J.; Otte, K.; Kalinin, V.I.; Silchenko, A.S.; Avilov, S.A.; et al. The marine triterpene glycoside frondoside A exhibits activity in vitro and in vivo in prostate cancer. Int. J. Cancer 2016, 138, 2450–2465. [Google Scholar] [CrossRef] [PubMed]
- Dyshlovoy, S.A.; Rast, S.; Hauschild, J.; Otte, K.; Alsdorf, W.H.; Madanchi, R.; Kalinin, V.I.; Silchenko, A.S.; Avilov, S.A.; Dierlamm, J.; et al. Frondoside A induces AIF-associated caspase-independent apoptosis in Burkitt lymphoma cells. Leuk. Lymphoma 2017, 58, 2905–2915. [Google Scholar] [CrossRef]
- Attoub, S.; Arafat, K.; Khalaf, T.; Sulaiman, S.; Iratni, R. Frondoside A Enhances the Anti-Cancer Effects of Oxaliplatin and 5-Fluorouracil on Colon Cancer Cells. Nutrients 2018, 10, 560. [Google Scholar] [CrossRef]
- Li, X.; Roginsky, A.B.; Ding, X.Z.; Woodward, C.; Collin, P.; Newman, R.A.; Bell, R.H., Jr.; Adrian, T.E. Review of the apoptosis pathways in pancreatic cancer and the anti-apoptotic effects of the novel sea cucumber compound, Frondoside, A. Ann. N. Y. Acad. Sci. 2008, 1138, 181–198. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, Y.H.; Kim, Y.; Lee, S.J. Frondoside A has an anti-invasive effect by inhibiting TPA-induced MMP-9 activation via NF-κB and AP-1 signaling in human breast cancer cells. Int. J. Oncol. 2012, 41, 933–940. [Google Scholar] [CrossRef]
- Sajwani, F.H.; Collin, P.; Adrian, T.E. Frondoside A potentiates the effects of conventional therapeutic agents in acute leukemia. Leuk. Res. 2017, 63, 98–108. [Google Scholar] [CrossRef]
- Lin, O.A.; Karim, Z.A.; Vemana, H.P.; Espinosa, E.V.; Khasawneh, F.T. The antidepressant 5-HT2A receptor antagonists pizotifen and cyproheptadine inhibit serotonin-enhanced platelet function. PLoS ONE 2014, 23, e87026. [Google Scholar] [CrossRef]
- Lindequist, U. Marine-Derived Pharmaceuticals—Challenges and Opportunities. Biomol. Ther. 2016, 24, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Dubey, S.; Kallubai, M.; Sarkar, A.; Subramanyam, R. Elucidating the active interaction mechanism of phytochemicals withanolide and withanoside derivatives with human serum albumin. PLoS ONE 2018, 13, e0200053. [Google Scholar] [CrossRef] [PubMed]
- Gale, A.J. Continuing education course #2: Current understanding of hemostasis. Toxicol. Pathol. 2011, 39, 273–280. [Google Scholar] [PubMed]
- Kim, S.; Foster, C.; Lecchi, A.; Quinton, T.M.; Prosser, D.M.; Jin, J.; Cattaneo, M.; Kunapuli, S.P. Protease-activated receptors 1 and 4 do not stimulate G(i) signaling pathways in the absence of secreted ADP and cause human platelet aggregation independently of G(i) signaling. Blood J. Am. Soc. Hematol. 2002, 99, 3629–3636. [Google Scholar] [CrossRef]
- Gurbel, P.A.; Kuliopulos., A.; Tantry, U.S. G-protein-coupled receptors signaling pathways in new antiplatelet drug development. Arterioscler Thromb. Vasc. Biol. 2015, 35, 500–512. [Google Scholar] [CrossRef]
- Pulcinelli, F.M.; Ciampa, M.T.; Favilla, M.; Pignatelli, P.; Riondino, S.; Gazzaniga, P.P. Concomitant activation of Gi protein-coupled receptor and protein kinase C or phospholipase C is required for platelet aggregation. FEBS Lett. 1999, 22, 37–40. [Google Scholar] [CrossRef]
- Guidetti, G.F.; Lova, P.; Bernardi, B.; Campus, F.; Baldanzi, G.; Graziani, A.; Balduini, C.; Torti, M. The Gi-coupled P2Y12 receptor regulates diacylglycerol-mediated signaling in human platelets. J. Biol. Chem. 2008, 24, 28795–28805. [Google Scholar] [CrossRef]
- Goggs, R.; Poole, A.W. Platelet signaling—A primer. J. Vet. Emerg. Crit. Care (San Antonio) 2012, 22, 5–29. [Google Scholar] [CrossRef]
- Stalker, T.J.; Newman, D.K.; Ma, P.; Wannemacher, K.M.; Brass, L.F. Platelet signaling. Handb. Exp. Pharmacol. 2012, 210, 59–85. [Google Scholar]
- Ampofo, E.; Später, T.; Müller, I.; Eichler, H.; Menger, M.D.; Laschke, M.W. The Marine-Derived Kinase Inhibitor Fascaplysin Exerts Anti-Thrombotic Activity. Mar. Drugs 2015, 13, 6774–6791. [Google Scholar] [CrossRef]
- Später, T.; Müller, I.; Eichler, H.; Menger, M.D.; Laschke, M.W.; Ampofo, E. Dual inhibition of PI3K and mTOR by VS-5584 suppresses thrombus formation. Platelets 2018, 29, 277–287. [Google Scholar] [CrossRef]
- Ni, R.; Vaezzadeh, N.; Zhou, J.; Weitz, J.I.; Cattaneo, M.; Gross, P.L. Effect of Different Doses of Acetylsalicylic Acid on the Antithrombotic Activity of Clopidogrel in a Mouse Arterial Thrombosis Model. Arterioscler Thromb. Vasc. Biol. 2018, 38, 2338–2344. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Wang, Y.; Lei, D.; Huang, L.; Wang, G.; Chi, Q.; Zheng, Y.; Gachet, C.; Mangin, P.H.; Zhu, L. Standardization of a well-controlled in vivo mouse model of thrombus formation induced by mechanical injury. Thromb. Res. 2016, 141, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Vanhaesebroeck, B.; Stephens, L.; Hawkins, P. PI3K signalling: The path to discovery and understanding. Nat. Rev. Mol. Cell Biol. 2012, 13, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Adrian, T.E.; Collin, P. The Anti-Cancer Effects of Frondoside, A. Mar Drugs 2018, 16, 64. [Google Scholar] [CrossRef] [PubMed]
- Al Marzouqi, N.; Iratni, R.; Nemmar, A.; Arafat, K.; Ahmed Al Sultan, M.; Yasin, J.; Collin, P.; Mester, J.; Adrian, T.E.; Attoub, S. Frondoside A inhibits human breast cancer cell survival, migration, invasion and the growth of breast tumor xenografts. Eur. J. Pharmacol. 2011, 668, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Menchinskaya, E.S.; Aminin, D.L.; Avilov, S.A.; Silchenko, A.S.; Andryjashchenko, P.V.; Kalinin, V.I.; Stonik, V.A. Inhibition of tumor cells multidrug resistance by cucumarioside A2-2, frondoside A and their complexes with cholesterol. Nat. Prod. Commun. 2013, 8, 1377–1380. [Google Scholar] [CrossRef]
- Maccarrone, M.; Bari, M.; Battista, N.; Finazzi-Agro, A. Estrogen stimulates arachidonoylethanolamide release from human endothelial cells and platelet activation. Blood J. Am. Soc. Hematol. 2002, 100, 4040–4048. [Google Scholar] [CrossRef]
- Selles, J.; Polini, N.; Alvarez, C.; Massheimer, V. Progesterone and 17 beta-estradiol acutely stimulate nitric oxide synthase activity in rat aorta and inhibit platelet aggregation. Life Sci. 2001, 69, 815–827. [Google Scholar] [CrossRef]
- Lindenblatt, N.; Platz, U.; Hameister, J.; Klar, E.; Menger, M.D.; Vollmar, B. Distinct effects of acute and chronic nicotine application on microvascular thrombus formation and endothelial function in male and female mice. Langenbecks Arch. Surg. 2007, 392, 285–295. [Google Scholar] [CrossRef]
- Laschke, M.W.; Menger, M.D. The dorsal skinfold chamber: A versatile tool for preclinical research in tissue engineering and regenerative medicine. Eur. Cell. Mater. 2016, 32, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Roesken, F.; Ruecker, M.; Vollmar, B.; Boeckel, N.; Morgenstern, E.; Menger, M.D. A new model for quantitative in vivo microscopic analysis of thrombus formation and vascular recanalisation: The ear of the hairless (hr/hr) mouse. Thromb. Haemost. 1997, 78, 1408–1414. [Google Scholar] [CrossRef] [PubMed]
- Vollmar, B.; Schmits, R.; Kunz, D.; Menger, M.D. Lack of in vivo function of CD31 in vascular thrombosis. Thromb. Haemost. 2001, 85, 160–164. [Google Scholar] [PubMed]
- Broze, G.J., Jr.; Yin, Z.F.; Lasky, N. A tail vein bleeding time model and delayed bleeding in hemophiliac mice. Thromb. Haemost. 2001, 85, 747–748. [Google Scholar] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ampofo, E.; Später, T.; Nalbach, L.; Menger, M.D.; Laschke, M.W. The Marine-Derived Triterpenoid Frondoside A Inhibits Thrombus Formation. Mar. Drugs 2020, 18, 111. https://doi.org/10.3390/md18020111
Ampofo E, Später T, Nalbach L, Menger MD, Laschke MW. The Marine-Derived Triterpenoid Frondoside A Inhibits Thrombus Formation. Marine Drugs. 2020; 18(2):111. https://doi.org/10.3390/md18020111
Chicago/Turabian StyleAmpofo, Emmanuel, Thomas Später, Lisa Nalbach, Michael D. Menger, and Matthias W. Laschke. 2020. "The Marine-Derived Triterpenoid Frondoside A Inhibits Thrombus Formation" Marine Drugs 18, no. 2: 111. https://doi.org/10.3390/md18020111
APA StyleAmpofo, E., Später, T., Nalbach, L., Menger, M. D., & Laschke, M. W. (2020). The Marine-Derived Triterpenoid Frondoside A Inhibits Thrombus Formation. Marine Drugs, 18(2), 111. https://doi.org/10.3390/md18020111