The Phylum Bryozoa as a Promising Source of Anticancer Drugs



Abstract
1. Introduction
2. Cytotoxic Compounds from Marine Bryozoans with Activity against Cancer Cell Lines
2.1. Alkaloids
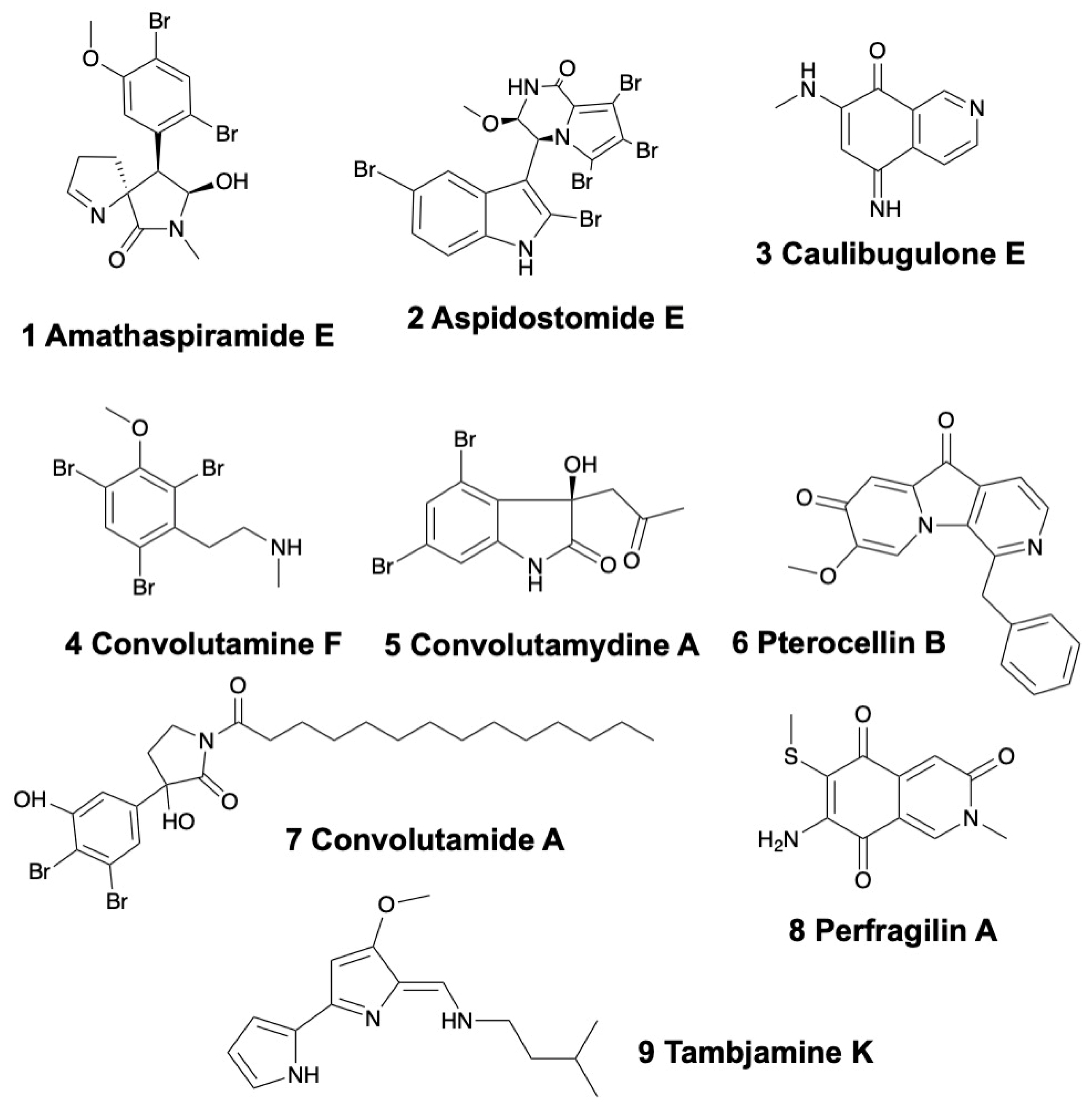
2.1.1. Amathaspiramides
2.1.2. Aspidostomides
2.1.3. Brominated Alkaloids
2.1.4. β-Carboline Alkaloids
2.1.5. Caulamidines
2.1.6. Caulibugulones
2.1.7. Convolutamides, Convolutamydines, and Convolutamines
2.1.8. Eusynstyelamides
2.1.9. Perfragilins
2.1.10. Polycyclic Indole Alkaloids
2.1.11. Pterocellins
2.1.12. Tambjamines
2.1.13. Terminoflustrindoles
2.2. Lactones
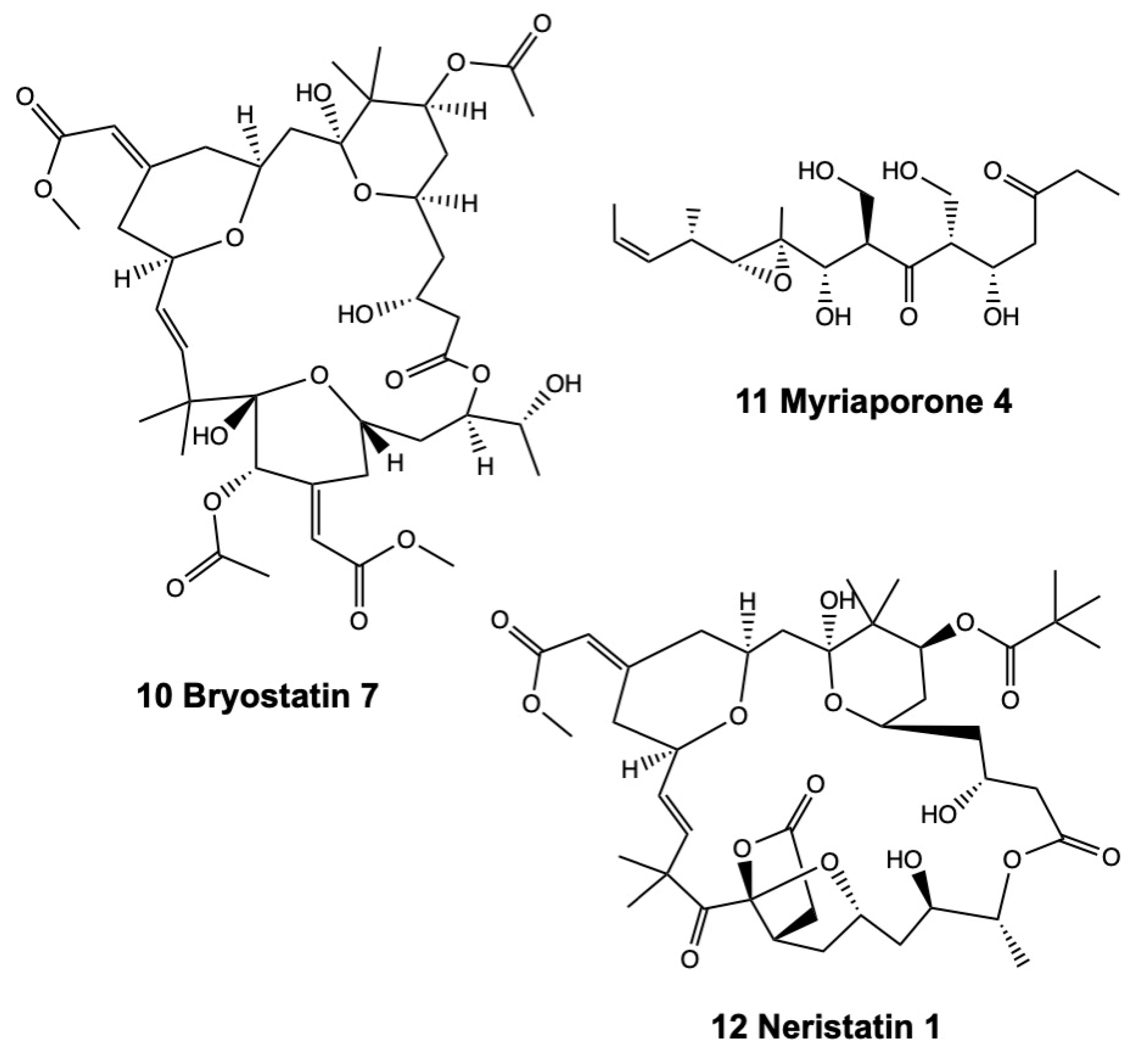
2.2.1. Bryostatins
2.2.2. Neristatin 1
2.2.3. Myriaporones
2.2.4. Other Lactones
2.3. Ceramides
2.4. Sterols
3. Other Compounds
4. Active Extracts
5. Future Research Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3T3 | normal mice fibroblasts |
| 3T3-SV40 | transformed mice fibroblasts |
| 768-O | renal carcinoma |
| ADM | adriamycin |
| ALCL | anaplastic large cell lymphoma |
| A-2058 | human melanoma cancer |
| BAEC | bovine aortic endothelial cell |
| bFGF | basic fibroblast growth factor |
| B-16 | mice melanoma |
| CaCo-2 | human epithelial colorectal adenocarcinoma |
| CCRF-CEM | human leukemia |
| C6 | rat histioblastome |
| HCT-116 | human colon cancer cell line |
| Hela | human cervical cancer cell line |
| Hep-G2 | human hepatocellular carcinoma |
| HL-60 | human plomyelocytic leukemia |
| HT-29 | human colon carcinoma |
| IC-2wt | murine tumor |
| PKC | protein kinase C |
| K1735-M2 | murine melanoma |
| KB | human epidermoid carcinoma |
| KB/VJ-300 | vincristine-resistant human cancer KB cell line |
| L-1210 | murine leukemia |
| LNcaP | human prostate cancer |
| M-14 | human melanoma |
| M-5076 | reticulum cell sarcoma |
| MALME-3M | melanoma |
| MB-231 | breast carcinoma |
| MCF-7 | human breast cancer |
| MDA-MB-435 | human breast cancer |
| MDA-N | melanoma |
| MiaPaCa-2 | human pancreatic cancer |
| MNP | marine natural products |
| MRC-5 | human fibroblasts |
| MV4-11 | acute myeloid leukemia |
| NCI-60 | a panel of 60 diverse human cancer cell lines of the U.S. National Cancer Institute |
| NCI-H23 | human non-small cell lung cancer |
| NCI-H460 | human non-small cell lung cancer |
| P-388 | murine lymphocytic leukemia |
| PC-3 | human prostate cancer |
| SF-268 | central nervous system cancer |
| SGC-7901 | human gastric carcinoma |
| SK-MEL-5 | human melanoma |
| SK-N-SH | human neuroblastome |
| U-937 | histiocytic lymphoma |
| VCR | vincristine |
References
- Cragg, G.M.; Newman, D.J. Natural product drug discovery in the next millennium. Pharm. Biol. 2001, 39, 8–17. [Google Scholar] [PubMed]
- Munro, M.H.G.; Blunt, J.W.; Dumdei, E.J.; Hickford, S.J.H.; Lill, R.E.; Li, S.; Battershill, C.N.; Duckworth, A.R. The discovery and development of marine compounds with pharmaceutical potential. J. Biotechnol. 1999, 70, 15–25. [Google Scholar] [CrossRef]
- Puglisi, M.P.; Sneed, J.M.; Ritson-Williams, R.; Young, R. Marine chemical ecology in benthic environments. Nat. Prod. Rep. 2019, 36, 410–429. [Google Scholar] [CrossRef] [PubMed]
- Lahlou, M. The success of natural products in drug discovery. Pharmacol. Pharm. 2013, 4, 17–31. [Google Scholar] [CrossRef]
- Schwartsmann, G.; Brondani, A.; Berlinck, R.G.S.; Jimeno, J. Marine organisms as a source of new anticancer agents. Lancet Oncol. 2001, 2, 221–225. [Google Scholar] [CrossRef]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Chung-Davidson, Y.W.; Bussy, U.; Li, W. Recent advances and applications of experimental technologies in marine natural product research. Mar. Drugs 2015, 13, 2694–2713. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Torres, V.; Encinar, J.A.; Herranz-López, M.; Pérez-Sánchez, A.; Galiano, V.; Barrajón-Catalán, E.; Micol, V. An updated review on marine anticancer compounds: The use of virtual screening for the discovery of small-molecule cancer drugs. Molecules 2017, 22, 1037. [Google Scholar] [CrossRef]
- Cortes, J.; O’Shaughnessy, J.; Loesch, D.; Blum, J.L.; Vahdat, L.T.; Petrakova, K.; Chollet, P.; Manikas, A.; Diéras, V.; Delozier, T.; et al. Eribulin monotherapy versus treatment of physician’s choice in patients with metastatic breast cancer (EMBRACE): A phase 3 open-label randomised study. Lancet 2011, 377, 914–923. [Google Scholar] [CrossRef]
- Schöffski, P.; Chawla, S.; Maki, R.G.; Italiano, A.; Gelderblom, H.; Choy, E.; Grignani, G.; Camargo, V.; Bauer, S.; Rha, S.Y.; et al. Eribulin versus dacarbazine in previously treated patients with advanced liposarcoma or leiomyosarcoma: A randomised, open-label, multicentre, phase 3 trial. Lancet 2016, 387, 1629–1637. [Google Scholar] [CrossRef]
- Olivera, B.M.; Cruz, L.J.; De Santos, V.; LeCheminant, G.; Griffin, D.; Zeikus, R.; McIntosh, J.M.; Galyean, R.; Varga, J. Neuronal calcium channel antagonists. Discrimination between Calcium channel subtypes using omega-Conotoxin from Conus magus venom. Biochemistry 2005, 26, 2086–2090. [Google Scholar] [CrossRef]
- Gopal, A.K.; Chen, R.; Smith, S.E.; Ansell, S.M.; Rosenblatt, J.D.; Savage, K.J.; Connors, J.M.; Engert, A.; Larsen, E.K.; Chi, X.; et al. Durable remissions in a pivotal phase 2 study of brentuximab vedotin in relapsed or refractory Hodgkin lymphoma. Blood 2015, 125, 1236–1243. [Google Scholar] [CrossRef]
- Newland, A.M.; Li, J.X.; Wasco, L.E.; Aziz, M.T.; Lowe, D.K. Brentuximab vedotin: A CD30-Directed antibody-cytotoxic drug conjugate. Pharmacotherapy 2013, 33, 93–104. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, D.; He, W.; Zhang, H.; Li, Z.; Luan, Y. Nanoassemblies from amphiphilic cytarabine prodrug for leukemia targeted therapy. J. Colloid Interface Sci. 2017, 487, 239–249. [Google Scholar] [CrossRef]
- D’Incalci, M.; Badri, N.; Galmarini, C.M.; Allavena, P. Trabectedin, a drug acting on both cancer cells and the tumour microenvironment. Br. J. Cancer 2014, 111, 646–650. [Google Scholar] [CrossRef]
- Atmaca, H.; Bozkurt, E.; Uzunoglu, S.; Uslu, R.; Karaca, B. A diverse induction of apoptosis by trabectedin in MCF-7 (HER2-/ER+) and MDA-MB-453 (HER2+/ER-) breast cancer cells. Toxicol. Lett. 2013, 221, 128–136. [Google Scholar] [CrossRef]
- Anjum, K.; Abbas, S.Q.; Asmat, S.; Shah, A.; Akhter, N.; Batool, S. Marine sponges as a drug treasure. Biomol. Ther. (Seoul). 2016, 24, 347–362. [Google Scholar] [CrossRef]
- Cooper, E.L.; Hirabayashi, K.; Strychar, K.B.; Sammarco, P.W. Corals and their potential applications to integrative medicine. Evid.-Based Complement. Altern. Med. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Tian, X.R.; Tang, H.F.; Tian, X.L.; Hu, J.J.; Huang, L.L.; Gustafson, K.R. Review of bioactive secondary metabolites from marine bryozoans in the progress of new drugs discovery. Future Med. Chem. 2018, 10, 1497–1514. [Google Scholar] [CrossRef]
- Sharp, J.H.; Winson, M.K.; Porter, J.S. Bryozoan metabolites: An ecological perspective. Nat. Prod. Rep. 2007, 24, 659–673. [Google Scholar] [CrossRef]
- Hayward, P.J. Antarctic Cheilostomatous Bryozoa; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Figuerola, B.; Gordon, D.P.; Cristobo, J. New deep Cheilostomata (Bryozoa) species from the Southwestern Atlantic: Shedding light in the dark. Zootaxa 2018, 4375, 211–249. [Google Scholar] [CrossRef]
- Figuerola, B.; Ballesteros, M.; Avila, C. Description of a new species of Reteporella (Bryozoa: Phidoloporidae) from the Weddell Sea (Antarctica) and the possible functional morphology of avicularia. Acta Zool. 2013, 73, 66–73. [Google Scholar] [CrossRef]
- Blauwe, H.D.E.; Gordon, D.P. New bryozoan taxa from a biodiversity hotspot in the Eastern Weddell Sea. Stud. Trent. Sci. Nat. 2014, 94, 53–78. [Google Scholar]
- Souto, J.; Albuquerque, M. Diversity and community structure of Cheilostomata (Bryozoa) from the Hayes Fracture Zone, Mid-Atlantic ridge. Deep. Res. Part I 2019, 147, 32–53. [Google Scholar] [CrossRef]
- Bock, P.E.; Gordon, D.P. Phylum Bryozoa Ehrenberg, 1831. Zootaxa 2013, 3703, 67–74. [Google Scholar] [CrossRef]
- Waeschenbach, A.; Taylor, P.D.; Littlewood, D.T.J. Molecular phylogenetics and evolution of bryozoans. Mol. Phylogenet. Evol. 2012, 62, 718–735. [Google Scholar] [CrossRef]
- Lebar, M.D.; Heimbegner, J.L.; Baker, B.J.; Baker, B. Cold-water marine natural products. Nat. Prod. Rep. 2007, 24, 774–797. [Google Scholar] [CrossRef]
- Figuerola, B.; Sala-Comorera, L.; Angulo-Preckler, C.; Vázquez, J.; Blanch, A.R.; Montes, M.J.; García-Aljaro, C.; Mercadé, E.; Avila, C. Antimicrobial activity of Antarctic bryozoans: An ecological perspective with potential for clinical applications. Mar. Environ. Res. 2014, 101, 52–59. [Google Scholar] [CrossRef]
- Figuerola, B.; Angulo-Preckler, C.; Núñez-Pons, L.; Moles, J.; Sala-Comorera, L.; García-Aljaro, C.; Blanch, A.R.; Avila, C. Experimental evidence of chemical defence mechanisms in Antarctic bryozoans. Mar. Environ. Res. 2017, 129, 68–75. [Google Scholar] [CrossRef]
- Taboada, S.; Núñez-Pons, L.; Avila, C. Feeding repellence of Antarctic and sub-Antarctic benthic invertebrates against the omnivorous sea star Odontaster validus. Polar Biol. 2013, 36, 13–25. [Google Scholar] [CrossRef]
- Figuerola, B.; Núñez-Pons, L.; Moles, J.; Avila, C. Feeding repellence in Antarctic bryozoans. Naturwissenschaften 2013, 100, 1069–1081. [Google Scholar] [CrossRef]
- Figuerola, B.; Taboada, S.; Monleón-getino, T.; Vázquez, J.; Àvila, C. Cytotoxic activity of Antarctic benthic organisms against the common sea urchin Sterechinus neumayeri. Oceanography 2013, 1, 1–9. [Google Scholar]
- Figuerola, B.; Núñez-Pons, L.; Monleón-Getino, T.; Avila, C. Chemo–ecological interactions in Antarctic bryozoans. Polar Biol. 2014, 37, 1017–1030. [Google Scholar] [CrossRef]
- Prinsep, M.R.; Yao, B.; Nicholson, B.K.; Gordon, D.P. The pterocellins, bioactive alkaloids from the marine bryozoan Pterocella vesiculosa. Phytochem. Rev. 2005, 325–331. [Google Scholar]
- Lopanik, N.B. Chemical defensive symbioses in the marine environment. Funct. Ecol. 2014, 28, 328–340. [Google Scholar] [CrossRef]
- Davidson, S.K.; Allen, S.W.; Lim, G.E.; Anderson, C.M.; Haygood, M.G. Evidence for the biosynthesis of bryostatins by the bacterial symbiont “Candidatus Endobugula sertula” of the bryozoan Bugula neritina. Appl. Environ. Microbiol. 2001, 67, 4531–4537. [Google Scholar] [CrossRef]
- Lopanik, N.; Lindquist, N.; Targett, N. Potent cytotoxins produced by a microbial symbiont protect host larvae from predation. Oecologia 2004, 139, 131–139. [Google Scholar] [CrossRef]
- Hughes, J.P.; Rees, S.S.; Kalindjian, S.B.; Philpott, K.L. Principles of early drug discovery. Br. J. Pharmacol. 2011, 162, 1239–1249. [Google Scholar] [CrossRef]
- Lefranc, F.; Koutsaviti, A.; Ioannou, E.; Kornienko, A.; Roussis, V.; Kiss, R.; Newman, D. Algae metabolites: From in vitro growth inhibitory effects to promising anticancer activity. Nat. Prod. Rep. 2019, 36, 810–841. [Google Scholar] [CrossRef]
- Morris, B.D.; Prinsep, M.R. Amathaspiramides A-F, novel brominated alkaloids from the marine bryozoan Amathia wilsoni. J. Nat. Prod. 1999, 62, 688–693. [Google Scholar] [CrossRef]
- Shimokawa, J.; Chiyoda, K.; Umihara, H.; Fukuyama, T. Antiproliferative activity of Amathaspiramide alkaloids and analogs. Chem. Pharm. Bull. 2016, 64, 1239–1241. [Google Scholar] [CrossRef]
- Patiño, L.P.; Muniain, C.; Knott, M.E.; Puricelli, L.; Palermo, J.A. Bromopyrrole alkaloids isolated from the Patagonian bryozoan Aspidostoma giganteum. J. Nat. Prod. 2014, 77, 1170–1178. [Google Scholar] [CrossRef]
- Tian, X.; Tang, H.; Li, Y.; Lin, H.; Zhang, X.; Feng, J.; Zhang, X. Studies on the chemical constituents from marine bryozoan Cryptosula pallasiana. Rec. Nat. Prod. 2014, 9, 628–632. [Google Scholar]
- Beutler, J.; Cardellina, J.; Prather, T.; Shoemaker, R.; Boyd, M.; Snader, K.; Snader, M. A cytotoxic beta-carboline from the bryozoan Catenicella cribraria. J. Nat. Prod. 1993, 56, 1825–1826. [Google Scholar] [CrossRef]
- Prinsep, R.; Munro, H.G. New cytotoxic p-carboline alkaloids from the marine bryozoan, Cribricellina cribraria. J. Nat. Prod. 1991, 5, 1068–1076. [Google Scholar] [CrossRef]
- Harwood, D.T.; Urban, S.; Blunt, J.W.; Munro, M.H.G. β-Carboline alkaloids from a New Zealand marine bryozoan, Cribricellina cribraria. Nat. Prod. Res. 2003, 17, 15–19. [Google Scholar] [CrossRef]
- Till, M.; Prinsep, M.R. 5-Bromo-8-methoxy-1-methyl-β-carboline, an alkaloid from the New Zealand marine bryozoan Pterocella vesiculosa. J. Nat. Prod. 2009, 72, 796–798. [Google Scholar] [CrossRef]
- Milanowski, D.J.; Oku, N.; Cartner, L.K.; Bokesch, H.R.; Williamson, R.T.; Saurí, J.; Liu, Y.; Blinov, K.A.; Ding, Y.; Li, X.C.; et al. Unequivocal determination of caulamidines A and B: Application and validation of new tools in the structure elucidation tool box. Chem. Sci. 2018, 9, 307–314. [Google Scholar] [CrossRef]
- Milanowski, D.J.; Gustafson, K.R.; Kelley, J.A.; Mcmahon, J.B. Caulibugulones A~F, novel cytotoxic isoquinoline quinones and iminoquinones from the marine bryozoan Caulibugula intermis. J. Nat. Prod. 2004, 67, 70–73. [Google Scholar] [CrossRef]
- Zhang, H.; Shigemori, H.; Ishibashi, M.; Kosaka, T.; Pettit, G.R.; Kamano, Y.; Kobayashi, J. Convolutamides A~F, novel γ-lactam alkaloids from the marine bryozoan Amathia convoluta. Tetrahedron 1994, 50, 10201–10206. [Google Scholar] [CrossRef]
- Kamano, Y.; Zhang, H.; Ichihara, Y.; Kizu, H.; Komiyama, K.; Pettit, G.R. Convolutamydine A, a novel bioactive hydroxyoxindole alkaloid from marine bryozoan Amathia convoluta. Tetrahedron Lett. 1995, 36, 2783–2784. [Google Scholar] [CrossRef]
- Zhang, H.; Kamano, Y.; Ichihara, Y.; Kizu, H.; Komiyama, K.; Itokawa, H.; Pettit, G.R. Isolation and structure of convolutamydines B~D from marine bryozoan Amathia convoluta. Tetrahedron 1995, 51, 5523–5528. [Google Scholar] [CrossRef]
- Hashima, H.; Hayashi, M.; Kamano, Y.; Sato, N. Synthesis and biological activities of the marine bryozoan alkaloids convolutamines A, C and F, and lutamides A and C. Bioorganic. Med. Chem. 2000, 8, 1757–1766. [Google Scholar] [CrossRef]
- Zhang, H.; Kamano, Y.; Kizu, H.; Itokawa, H.; Pettit, G.; Herald, C. Convolutamines A-E, novel β-phenylethylamine alkaloids from marine bryozoan Amathia convoluta. Chem. Lett. 1994, 26, 2271–2274. [Google Scholar] [CrossRef]
- Kamano, Y.; Kotake, A.; Hashima, H.; Hayakawa, I.; Hiraide, H.; Zhang, H.; Kizu, H.; Komiyana, K.; Hayashi, M.; Pettit, G.R. Three new alkaloids, convolutamines F and G, and convolutamydine E, from the Floridian marine bryozoan Amathia convoluta. Collect. Czechoslov. Chem. Commun. 1999, 64, 1147–1153. [Google Scholar] [CrossRef]
- Llorach-Pares, L.; Nonell-Canals, A.; Avila, C.; Sanchez-Martinez, M. Kororamides, Convolutamines, and Indole derivatives as possible Tau and Dual-specificity kinase inhibitors for Alzheimer’s disease: A computational study. Mar. Drugs 2018, 16, 386. [Google Scholar] [CrossRef]
- Tadesse, M.; Tabudravu, J.N.; Jaspars, M.; Strøm, M.B.; Hansen, E.; Andersen, J.H.; Kristiansen, P.E.; Haug, T. The antibacterial ent-Eusynstyelamide B and Eusynstyelamides D, E, and F from the Arctic bryozoan Tegella cf. spitzbergensis. J. Nat. Prod. 2011, 74, 837–841. [Google Scholar] [CrossRef]
- Tapiolas, D.M.; Bowden, B.F.; Abou-Mansour, E.; Willis, R.H.; Doyle, J.R.; Muirhead, A.N.; Liptrot, C.; Llewellyn, L.E.; Wolff, C.W.W.; Wright, A.D.; et al. Eusynstyelamides A, B, and C, nNOS inhibitors, from the ascidian Eusynstyela latericius. J. Nat. Prod. 2009, 72, 1115–1120. [Google Scholar] [CrossRef]
- Swersey, J.C.; Ireland, C.M. Eusynstyelamide, a highly modified dimer peptide from the ascidian Eusynstyela misakiensis. J. Nat. Prod. 1994, 57, 842–845. [Google Scholar] [CrossRef]
- Choi, Y.-H.; Park, A.; Schmitz, F.J. Perfragilins A and B, cytotoxic Isoquinolinequinones from the bryozoan Membranipora perfragilis. J. Nat. Prod. 1993, 56, 1431–1433. [Google Scholar] [CrossRef]
- Arai, T.; Yazawa, K.; Mikami, Y. Isolation and characterization of satellite antibiotics, mimosamycin and chlorocarcins from Streptomyces lavendulae, streptothricin source. J. Antibiot. (Tokyo). 1976, XXIX, 398–407. [Google Scholar] [CrossRef]
- He, H.; Faulkner, D.J. Renieramycins E and F from the sponge Reniera sp.: Reassignment of the stereochemistry of the Renieramycins. Am. Chem. Soc. 1989, 54, 5822–5824. [Google Scholar] [CrossRef]
- Anthoni, U.; Bock, K.; Chevolot, L.; Larsen, C.; Nielsen, P.H.; Christophersen, C. Chartellamide A and B, halogenated β-lactam indole-imidazole alkaloids from the marine bryozoan Chartella papyraceala. J. Org. Chem. 1987, 52, 5638–5639. [Google Scholar] [CrossRef]
- Anthoni, U.; Chevolot, L.; Larsen, C.; Nielsen, P.H.; Christophersen, C. Marine alkaloids. 12. Chartellines, halogenated β-lactam alkaloids from the marine bryozoan Chartella papyraceala. J. Org. Chem. 1987, 52, 4709–4712. [Google Scholar] [CrossRef]
- Hansen, K.; Isaksson, J.; Bayer, A.; Johansen, J.A.; Andersen, J.H.; Hansen, E. Securamine derivatives from the Arctic bryozoan Securiflustra securifrons. J. Nat. Prod. 2017, 80, 3276–3283. [Google Scholar] [CrossRef]
- Lysek, N.; Rachor, E.; Lindel, T. Isolation and structure elucidation of deformylflustrabromine from the North Sea bryozoans Flustra foliacea. Verlag der Zeitschrift für Naturforsch. 2002, 57, 1056–1061. [Google Scholar] [CrossRef]
- Yao, B.; Prinsep, R.; Nicholson, B.K.; Gordon, D.P. The Pterocellins, novel bioactive alkaloids from the marine bryozoan Pterocella vesiculosa. J. Nat. Prod. 2003, 66, 1074–1077. [Google Scholar] [CrossRef]
- Prinsep, M.R. Further Pterocellins from the New Zealand marine bryozoan Pterocella vesiculosa. Notes J. Nat. Prod. 2008, 71, 134–136. [Google Scholar] [CrossRef]
- Wang, A.T.; Prinsep, M.R.; Martinus, R.D. Pterocellin A isolated from marine bryozoan Pterocella vesiculosa is cytotoxic to human HeLa cells via mitochondrial apoptotic processes. Springerplus 2016, 5, 742. [Google Scholar] [CrossRef]
- Carte, B.; Faulkner, D.J. Defensive metabolites from three nembrothid nudibranchs. J. Org. Chem. 1983, 48, 2314–2318. [Google Scholar] [CrossRef]
- Lindquist, N.; Fenical, W. New tambjamine class alkaloids from the marine ascidian Atapozoa sp. and its nudibranch predators. Origin of the tambjamines in Atapozoa. Experientia 1991, 47, 503–504. [Google Scholar] [CrossRef]
- Blackman, J.; Li, C. New Tambjamine alkaloids from the marine bryozoan Bugula dentata. Aust. J. Chem. 1994, 47, 1625–1629. [Google Scholar] [CrossRef]
- Franks, A.; Haywood, P.; Holmstrom, C.; Egan, S.; Kjelleverg, S.; Kumar, N. Isolation and structure elucidation of a novel yellow pigment from the marine bacterium Pseudoalteromonas tunicata. Molecules 2005, 10, 1286–1291. [Google Scholar] [CrossRef]
- Carté, B.; Faulkner, D.J. Role of secondary metabolites in feeding associations between a predatory nudibranch, two grazing nudibranchs, and a bryozoan. J. Chem. Ecol. 1986, 12, 795–804. [Google Scholar] [CrossRef]
- Carbone, M.; Irace, C.; Costagliola, F.; Castelluccio, F.; Villani, G.; Calado, G.; Padula, V.; Cimino, G.; Cervera, J.L.; Santamaria, R.; et al. A new cytotoxic tambjamine alkaloid from the Azorean nudibranch Tambja ceutae. Bioorg. Med. Chem. Lett. 2010, 20, 2668–2670. [Google Scholar] [CrossRef]
- Aldrich, L.N.; Stoops, S.L.; Crews, B.C.; Marnett, L.J.; Lindsley, C.W. Total synthesis and biological evaluation of tambjamine K and a library of unnatural analogs. Bioorg. Med. Chem. Lett. 2010, 20, 5207–5211. [Google Scholar] [CrossRef]
- Maltseva, A.; Kotenko, O.N.; Shabalin, K. Novel brominated fungicidal alkaloid isolated from the marine bryozoan Chartella membranaceatruncata (Smitt, 1868). Stud. Trentini di Sci. Nat. 2014, 94, 163–168. [Google Scholar]
- Maltseva, A.L.; Kotenko, O.N.; Kutyumov, V.A.; Darya, A.; Shavarda, A.L.; Winson, M.K.; Ostrovsky, A.N. Novel brominated metabolites from Bryozoa: A functional analysis. Nat. Prod. Res. 2017, 31, 1840–1848. [Google Scholar] [CrossRef]
- Pettit, G.R.; Herald, C.L.; Doubek, D.L.; Herald, D.L. Isolation and structure of bryostatin 1. J. Am. Chem. Soc. 1982, 6846–6848. [Google Scholar] [CrossRef]
- Pettit, G.R.; Kamano, Y.; Aoyagi, R.; Herald, C.L.; Doubek, D.L.; Schmidt, J.M.; Rudloe, J.J. Antineoplastic agents 100. The marine bryozoan Amathia convoluta. Tetrahedron 1985, 41, 985–994. [Google Scholar] [CrossRef]
- Pettit, G.R.; Kamano, Y.; Herald, C.L.; Tozawa, M. Isolation and structure of bryostatins 5–7. Can. J. Chem. 1985, 63, 1204–1208. [Google Scholar] [CrossRef]
- Pettit, G.R.; Kamano, Y.; Herald, C.L. Antineoplastic agents 119. Isolation and structure of bryostatins 10 and 11. J. Org. Chem. 1987, 52, 2848–2854. [Google Scholar] [CrossRef]
- Pettit, G.R.; Leet, J.E.; Herald, C.L.; Kamano, Y.; Boettner, F.E.; Baczynskyj, L.; Nieman, R.A. Antineoplastic agents 126. Isolation and structure of bryostatins 12 and 13. J. Org. Chem. 1987, 52, 2854–2860. [Google Scholar] [CrossRef]
- Pettit, G.R.; Gao, F.; Sengupta, D.; Coll, J.C.; Herald, C.L.; Doubek, D.L.; Schmidt, J.M.; van Camp, J.R.; Rudloe, J.J.; Nieman, R.A. Isolation and structure of bryostatins 14 and 15. Tetrahedron 1991, 47, 3601–3610. [Google Scholar] [CrossRef]
- Pettit, G.R.; Herald, C.L.; Kamano, Y.; Gust, D.; Aoyagi, R. The structure of bryostatin 2 from the marine bryozoan Bugula neritina. J. Nat. Prod. 1983, 2, 528–531. [Google Scholar] [CrossRef]
- Pettit, G.R.; Herald, C.L.; Kamano, Y. Antineoplastic agents 93. Structure of the Bugula neritina (marine bryozoa) antineoplastic component bryostatin 3. J. Org. Chem. 1983, 48, 5354–5356. [Google Scholar] [CrossRef]
- Pettit, G.R.; Kamano, Y.; Herald, C.L. Antineoplastic agents, 118. Isolation and structure of bryostatin 9. J. Nat. Prod. 1986, 49, 661–664. [Google Scholar] [CrossRef]
- Pettit, G.R.; Kamano, Y.; Herald, C.L.; Tozawa, M. Structure of Bryostatin 4. An important antineoplastic constituent of geographically diverse Bugula neritina (Bryozoa). J. Am. Chem. Soc. 1984, 106, 6768–6771. [Google Scholar] [CrossRef]
- Pettit, G.R.; Gao, F.; Blumberg, P.M.; Herald, C.L.; Coll, J.C.; Kamano, Y.; Lewin, N.E.; Schmidt, J.M.; Chapuis, J.C. Antineoplastic agents. 340. Isolation and structural elucidation of bryostatins 16-18. J. Nat. Prod. 1996, 59, 286–289. [Google Scholar] [CrossRef]
- Mutter, R.; Wills, M. Chemistry and clinical biology of the bryostatins. Bioorg. Med. Chem. 2000, 8, 1841–1860. [Google Scholar] [CrossRef]
- Ruan, B.; Zhu, H. The chemistry and biology of the bryostatins: Potential PKC inhibitors in clinical development. Curr. Med. Chem. 2012, 19, 2652–2664. [Google Scholar] [CrossRef]
- Kollár, P.; Rajchard, J.; Balounová, Z.; Pazourek, J. Marine natural products: Bryostatins in preclinical and clinical studies. Pharm. Biol. 2014, 52, 237–242. [Google Scholar] [CrossRef]
- Nelson, T.J.; Sun, M.K.; Lim, C.; Sen, A.; Khan, T.; Chirila, F.V.; Alkon, D.L. Bryostatin effects on cognitive function and PKCϵ in Alzheimer’s disease Phase IIa and expanded access trials. J. Alzheimer’s Dis. 2017, 58, 521–535. [Google Scholar] [CrossRef]
- Kageyama, M.; Tamura, T.; Nantz, M.H.; Roberts, J.C.; Somfai, P.; Whritenour, D.C.; Masamune, S. Synthesis of Bryostatin 7. J. Am. Chem. Soc. 1990, 112, 7407–7408. [Google Scholar] [CrossRef]
- Kortmansky, J.; Schwartz, G.K. Bryostatin-1: A novel PKC inhibitor in clinical development. Cancer Investig. 2003, 21, 924–936. [Google Scholar] [CrossRef]
- Hornung, R.L.; Pearson, J.W.; Beckwith, M.; Longo, D.L. Preclinical evaluation of bryostatin as an anticancer agent against several murine tumor cell lines: In vitro versus in vivo activity. Cancer Res. 1992, 52, 101–107. [Google Scholar]
- Trenn, G.; Pettit, G.; Takayama, H.; Hu-Li, J.; Sitkovskf, M. V Immunomodulating properties of a novel series of protein kinase C activators. The bryostatins. J. Immunol. 2018, 140, 433–439. [Google Scholar]
- Kraft, A.S.; Smith, J.B.; Berkow, R.L. Bryostatin, an activator of the calcium phospholipid-dependent protein kinase, blocks phorbol ester-induced differentiation of human Promyelocytic leukemia cells HL-60. Proc. Natl. Acad. Sci. USA 1986, 83, 1334–1338. [Google Scholar] [CrossRef]
- van der Hem, K.G.; Drager, A.M.; Huijgens, P.C.; Tol, C.; Deville, W.; Langenhuijsen, M.M. The differentiation inducing effect of bryostatin 5 on human myeloid blast cells is potentiated by vitamin D3. Leukemia 1994, 8, 266–273. [Google Scholar]
- Kraft, A.S.; Woodley, S.; Pettit, G.R.; Gao, F.; Coll, J.C.; Wagner, F. Comparison of the antitumor activity of bryostatins 1, 5, and 8. Cancer Chemother. Pharmacol. 1995, 37, 271–278. [Google Scholar] [CrossRef]
- van der Hem, K.G.; Dräger, A.M.; Odding, J.H.; Langenhuijsen, M.M.A.C.; Huijgens, P.C. Bryostatin-5 stimulates normal human hematopoiesis and inhibits proliferation of HL60 leukemic cells. Leuk. Res. 1995, 19, 7–13. [Google Scholar] [CrossRef]
- Houwen, L.; Gaolin, L.; Yanghua, Y.; Xinsheng, Y.; Houming, W. Studies on antineoplastic constituents from marine bryozoan Bugula neritina inhabiting South China Sea: Isolation and structural elucidation of a novel macrolide. Acad. J. Second Mil. Med. Coll. 2004, 25, 473–478. [Google Scholar]
- Manning, T.J.; Land, M.; Rhodes, E.; Rudloe, J.; Phillips, D.; Lam, T.T.; Purcell, J.; Cooper, H.J.; Emmett, M.R.; Marshall, G. Identifying bryostatins and potential precursors from the bryozoan Bugula neritina. Nat. Prod. Res. 2005, 19, 467–491. [Google Scholar] [CrossRef]
- Sudek, S.; Lopanik, N.B.; Waggoner, L.E.; Hildebrand, M.; Anderson, C.; Liu, H.; Patel, A.; Sherman, D.H.; Haygood, M.G. Identification of the putative bryostatin polyketide synthase gene cluster from “Candidatus Endobugula sertula”, the uncultivated microbial symbiont of the marine bryozoan Bugula neritina. J. Nat. Prod. 2007, 70, 67–74. [Google Scholar] [CrossRef]
- Haygood, M.G.; Davidson, S.K. Small-subunit rRNA genes and in situ hybridization with oligonucleotides specific for the bacterial symbionts in the larvae of the bryozoan Bugula neritina and proposal of “Candidatus Endobugula sertula”. Appl. Environ. Microbiol. 1997, 63, 4612–4616. [Google Scholar]
- DeChristopher, B.A.; Fan, A.C.; Felsher, D.W.; Wender, P.A. “Picolog,” a synthetically-available bryostatin analog, inhibits growth of MYC-induced lymphoma in vivo. Oncotarget 2012, 3, 58–66. [Google Scholar] [CrossRef]
- Kedei, N.; Kraft, M.B.; Keck, G.E.; Herald, C.L.; Melody, N.; Pettit, G.R.; Blumberg, P.M. Neristatin 1 provides critical insight into bryostatin 1 structure-function relationships. J. Nat. Prod. 2015, 78, 896–900. [Google Scholar] [CrossRef]
- Pettit, G.R.; Gao, F.; Herald, D.L.; Blumberg, P.M.; Lewin, N.E.; Nieman, R.A. Antineoplastic agents. 224. Isolation and structure of neristatin 1. J. Am. Chem. Soc. 1991, 113, 6693–6695. [Google Scholar] [CrossRef]
- Cheng, J.-F.; Lee, J.; Sakai, R.; Jares-Erijman, E.A.; Silva, M.V.; Rinehart, K.L. Myriaporones 1-4, cytotoxic metabolites from the mediterranean bryozoan Myriapora truncata. J. Nat. Prod. 2007, 70, 332–336. [Google Scholar] [CrossRef]
- Tian, X.R.; Gao, Y.Q.; Tian, X.L.; Li, J.; Tang, H.F.; Li, Y.S.; Lin, H.W.; Ma, Z.Q. New cytotoxic secondary metabolites from marine bryozoan Cryptosula pallasiana. Mar. Drugs 2017, 15, 120. [Google Scholar] [CrossRef]
- Ojika, M.; Yoshino, G.; Sakagami, Y. Novel Ceramide 1-sulfates, potent DNA Topoisomerase I inhibitors isolated from the bryozoa Watersipora cucullata. Tetrahedron Lett. 1997, 38, 4235–4238. [Google Scholar] [CrossRef]
- Tian, X.; Tang, H.; Feng, J.; Li, Y.; Lin, H.; Fan, X.; Zhang, X. Neritinaceramides A–E, new ceramides from the marine bryozoan Bugula neritina inhabiting South China Sea and their cytotoxicity. Mar. Drugs 2014, 12, 1987–2003. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, H.; Chen, J.; Tang, H. New cytotoxic oxygenated sterols from marine bryozoan Bugula neritina. Nat. Prod. Res. Former. Nat. Prod. Lett. 2011, 25, 1505–1511. [Google Scholar]
- Tian, X.R.; Tang, H.F.; Li, Y.S.; Lin, H.W.; Fan, X.P.; Feng, J.T.; Zhang, X. New 3β, 6β-dihydroxy and 3β, 5α, 6β-trihydroxy sterols from marine bryozoan Bugula neritina in South China Sea and their cytotoxicity. Phytochem. Lett. 2014, 9, 1–6. [Google Scholar] [CrossRef]
- Tian, X.; Tang, H.; Li, Y.; Lin, H.; Chen, X.; Ma, N.; Yao, M.; Zhang, P. New cytotoxic oxygenated sterols from the marine bryozoan Cryptosula pallasiana. Mar. Drugs 2011, 9, 162–183. [Google Scholar] [CrossRef]
- Notaro, G.; Piccialli, V.; Sica, D. New steroidal hydroxyketones and closely related diols from the marine sponge Cliona copiosa. J. Nat. Prod. 1992, 55, 1588–1594. [Google Scholar] [CrossRef]
- Riccardis, F.; Minale, L.; Iorizzi, M.; Debitus, C.; Lévi, C. Marine sterols. Side-chain-oxygenated sterols. Possibly of abiotic origin, from the New Caledonian sponge Stelodoryx chlorophylla. J. Nat. Prod. 1993, 56, 282–287. [Google Scholar] [CrossRef]
- Jeong, S.J.; Higuchi, R.; Miyamoto, T.; Ono, M.; Kuwano, M.; Mawatari, S.F. Bryoanthrathiophene, a new antiangiogenic constituent from the bryozoan Watersipora subtorquata (d’Orbigny, 1852). J. Nat. Prod. 2002, 65, 1344–1345. [Google Scholar] [CrossRef]
- Dobretsov, S.; Tamimi, Y.; Al-kindi, M.A.; Burney, I. Screening for anti-cancer compounds in marine organisms in Oman. Sultan Qaboos Univ Med. J. 2016, 16, 168–174. [Google Scholar] [CrossRef]
- Hansen, K.Ø.; Isaksson, J.; Glomsaker, E.; Andersen, J.H.; Hansen, E. Ponasterone A and F, Ecdysteroids from the arctic bryozoan Alcyonidium gelatinosum. Molecules 2018, 23, 1481. [Google Scholar] [CrossRef]
- Winston, J.E.; Hakansson, E. Molting by Cupuladria doma, a free-living bryozoan. Bull. Mar. Sci. 1989, 44, 1152–1158. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef]
- Rahman, M.A. An overview of the medical applications of marine skeletal matrix proteins. Mar. Drugs 2016, 14, 167. [Google Scholar] [CrossRef]
- Schaufelberger, D.E.; Koleck, M.P.; Beutler, J.A.; Vatakis, A.M.; Belinda Alvarado, A.; Andrews, P.; Marzo, L.V.; Muschik, G.M.; Roach, J.; Ross, J.T.; et al. The large-scale isolation of bryostatin 1 from Bugula neritina following current good manufacturing practices. J. Nat. Prod. 1991, 54, 1265–1270. [Google Scholar] [CrossRef]
- Figuerola, B.; Gore, D.B.; Johnstone, G.; Stark, J.S. Spatio-temporal variation of skeletal Mg-calcite in Antarctic marine calcifiers. PLoS ONE 2019, 14, e0210231. [Google Scholar] [CrossRef]
- Chown, S.L.; Brooks, C.M.; Terauds, A.; Le Bohec, C.; van Klaveren-Impagliazzo, C.; Whittington, J.D.; Butchart, S.H.M.; Coetzee, B.W.T.; Collen, B.; Convey, P.; et al. Antarctica and the strategic plan for biodiversity. PLoS Biol. 2017, 15, 1–10. [Google Scholar] [CrossRef]
- Marsden, M.D.; Wu, X.; Navab, S.M.; Loy, B.A.; Schrier, A.J.; DeChristopher, B.A.; Shimizu, A.J.; Hardman, C.T.; Ho, S.; Ramirez, C.M.; et al. Characterization of designed, synthetically accessible bryostatin analog HIV latency reversing agents. Virology 2018, 520, 83–93. [Google Scholar] [CrossRef]
- Esquenazi, E.; Coates, C.; Simmons, L.; Gonzalez, D.; Gerwick, W.H.; Dorrestein, P.C. Visualizing the spatial distribution of secondary metabolites produced by marine cyanobacteria and sponges via MALDI-TOF imaging. Mol. Biosyst. 2008, 4, 562–570. [Google Scholar] [CrossRef]
- Kaeberlein, T.; Lewis, K.; Epstein, S. Isolating “uncultivabte” microorganisms in pure culture in a simulated natural environment. Science 2002, 296, 1127–1129. [Google Scholar] [CrossRef]
- Piel, J.; Hui, D.; Wen, G.; Butzke, D.; Platzer, M.; Fusetani, N.; Matsunaga, S. Antitumor polyketide biosynthesis by an uncultivated bacterial symbiont of the marine sponge Theonella swinhoei. Proc. Natl. Acad. Sci. USA 2004, 101, 16222–16227. [Google Scholar] [CrossRef]
- Wilson, M.C.; Mori, T.; Rückert, C.; Uria, A.R.; Helf, M.J.; Takada, K.; Gernert, C.; Steffens, U.A.E.; Heycke, N.; Schmitt, S.; et al. An environmental bacterial taxon with a large and distinct metabolic repertoire. Nature 2014, 506, 58–62. [Google Scholar] [CrossRef]
- Schofield, M.M.; Jain, S.; Porat, D.; Dick, G.J.; Sherman, D.H. Identification and analysis of the bacterial endosymbiont specialized for production of the chemotherapeutic natural product ET-743. Environ. Microbiol. 2015, 17, 3964–3975. [Google Scholar] [CrossRef]
- Russo, P.; Del Bufalo, A.; Fini, M. Deep sea as a source of novel-anticancer drugs: Update on discovery and preclinical/clinical evaluation in a systems medicine perspective. EXCLI J. 2015, 14, 228–236. [Google Scholar]



{kind=link}
{kind=link}
| O. Cheilostomatida | Geographical Area | Compounds | Activity against Cell Lines | References |
|---|---|---|---|---|
| Fam. Aspidostomatidae | ||||
| Aspidostoma giganteum (Busk, 1854) | Patagonia | Alkaloids: Aspidostomides (Figure 1.2) | 768-O | [45] |
| Fam. Bugulidae | ||||
| Bugula neritina (Linnaeus, 1758) | California, China, Gulf of Mexico | Lactones: Bryostatins (Figure 2.10) | U-937, HL-60, P-388, B-16, K1735-M2, LNcaP, M-5076 | [80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,107] |
| Neristatins (Figure 2.12) | P-388 | [108] | ||
| Caulibugula intermis Harmer, 1926 | Palau | Alkaloids: Caulibugulones (Figure 1.3) | IC-2wt | [50] |
| Virididentula (Bugula) dentata (Lamouroux, 1816) | North Atlantic | Alkaloids: Tambjamines (Figure 1.9) | CaCo-2 | [76,77] |
| Fam. Catenicellidae | ||||
| Paracribricellina (Cribricellina) cribraria (Busk, 1852) | Australia, New Zealand | Alkaloids: β-Carboline alkaloids | NCI-60, P-388 | [45,46,47] |
| Pterocella vesiculosa (Lamarck, 1816) | New Zealand | Alkaloids: β-Carboline alkaloids | P-388 | [48] |
| Pterocellins (Figure 1.6) | P-388, CCRF-CEM, MALME-3M, NCI-H23, M-14, SK-MEL-5, MDA-MB-435, MDA-N, Hela, and others | [68,69,70] | ||
| Fam. Cryptosulidae | ||||
| Cryptosula pallasiana (Moll, 1803) | China | Lactones: Other lactones | HL-60, Hep-G2, SGC-7901 | [111] |
| Fam. Flustridae | ||||
| Flustra foliacea (Linnaeus, 1758) | North Sea | Alkaloids: Deformylflustrabromine | HCT-116 | [67] |
| Securiflustra securifrons (Pallas, 1766) | Norway | Alkaloids: Securamines | A-2058, HT-29, MCF-7, MRC-5 | [66] |
| Terminoflustra (Chartella) membranaceotruncata (Smitt, 1868) | White Sea | Alkaloids: Terminoflustrindoles | 3T3, 3T3-SV40, SK-N-SH, C6, B-16, U-937 | [78,79] |
| Fam. Membraniporidae | ||||
| Biflustra perfragilis MacGillivray, 1881 | South Australia | Alkaloids: Perfragilins (Figure 1.8) | P-388 | [61] |
| Fam. Myriaporidae | ||||
| Myriapora truncata (Pallas, 1766) | Mediterranean | Lactones: Myriaporones (Figure 2.11) | L-1210 | [110] |
| Fam. Watersiporidae | ||||
| Watersipora cucullata (Busk, 1854) | Japan | Ceramides | DNA topoisomerase I enzyme | [112] |
| Watersipora subtorquata (d’Orbigny, 1852) | Japan | Other compounds | BAEC | [119] |
| O. Ctenostomatida | ||||
| Fam. Vesiculariidae | ||||
| Amathia convoluta (Lamarck, 1816) | Gulf of Mexico | Alkaloids: Convolutamides, convolutamydines and convolutamines (Figure 1.4, Figure 1.5, Figure 1.7) | L-1210, KB, KB/VJ-300, P-388, HL-60, and other resistant lines | [51,52,53,54,55,56] |
| Amathia wilsoni Kirkpatrick, 1888 | New Zealand | Alkaloids: Amathaspiramides (Figure 1.1) | MiaPaCa-2 | [41,42] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figuerola, B.; Avila, C. The Phylum Bryozoa as a Promising Source of Anticancer Drugs. Mar. Drugs 2019, 17, 477. https://doi.org/10.3390/md17080477
Figuerola B, Avila C. The Phylum Bryozoa as a Promising Source of Anticancer Drugs. Marine Drugs. 2019; 17(8):477. https://doi.org/10.3390/md17080477
Chicago/Turabian StyleFiguerola, Blanca, and Conxita Avila. 2019. "The Phylum Bryozoa as a Promising Source of Anticancer Drugs" Marine Drugs 17, no. 8: 477. https://doi.org/10.3390/md17080477
APA StyleFiguerola, B., & Avila, C. (2019). The Phylum Bryozoa as a Promising Source of Anticancer Drugs. Marine Drugs, 17(8), 477. https://doi.org/10.3390/md17080477