A Novel Peptide from Abalone (Haliotis discus hannai) to Suppress Metastasis and Vasculogenic Mimicry of Tumor Cells and Enhance Anti-Tumor Effect In Vitro

 ,
,
Abstract
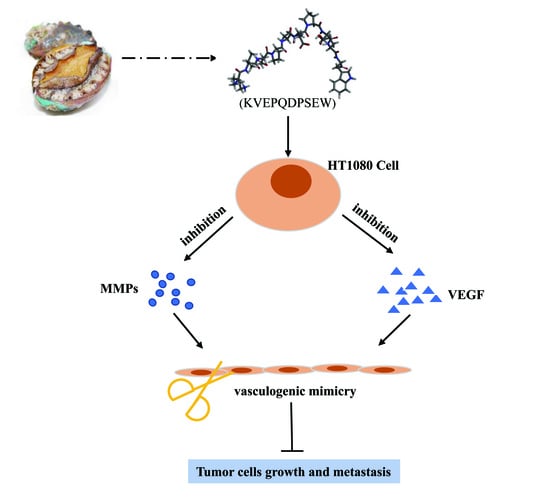
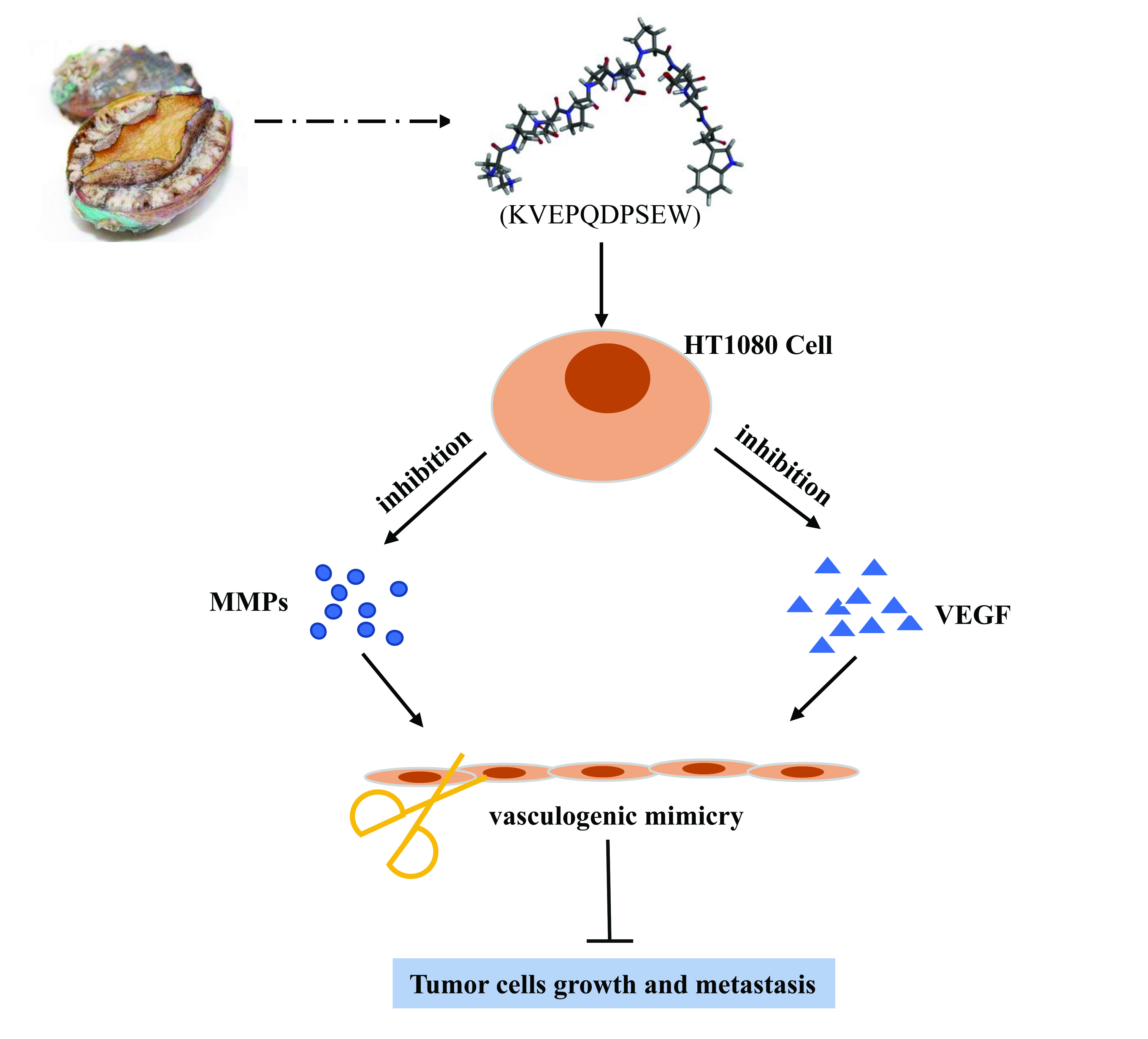
1. Introduction
2. Results
2.1. Cell Viability and Colony Formation Assay
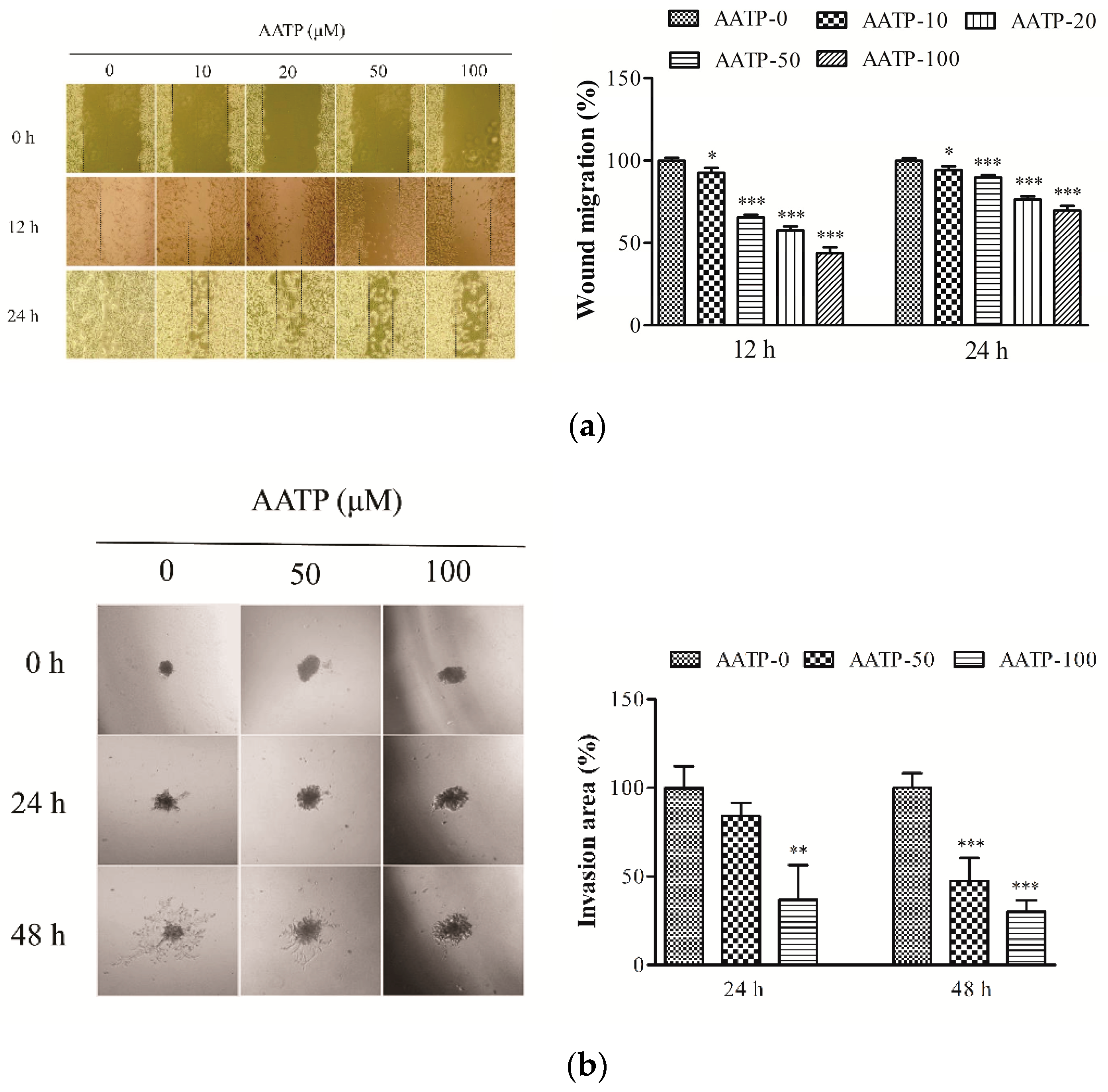
2.2. AATP Inhibits Tumor Cells Migration and Invasion
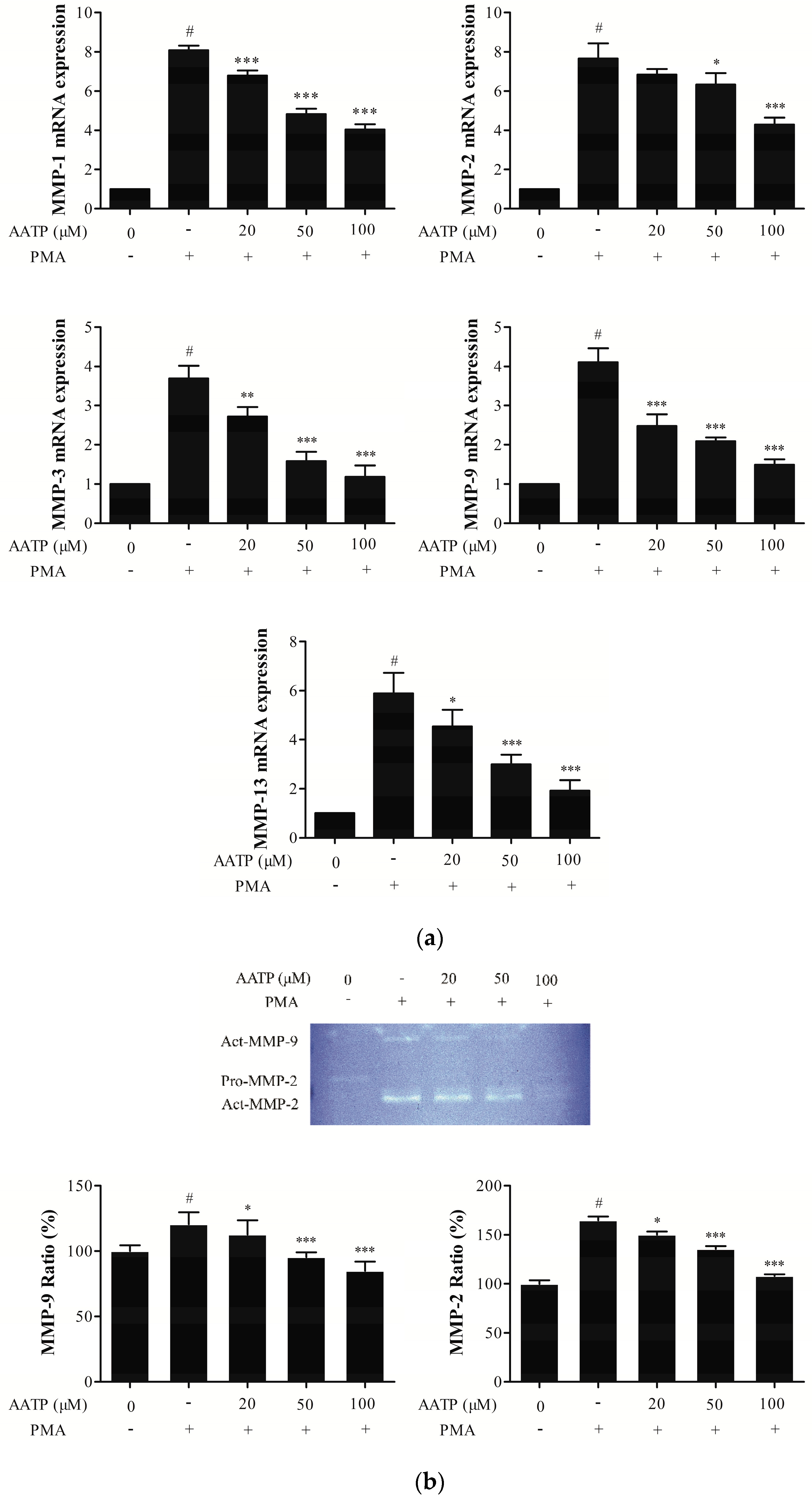
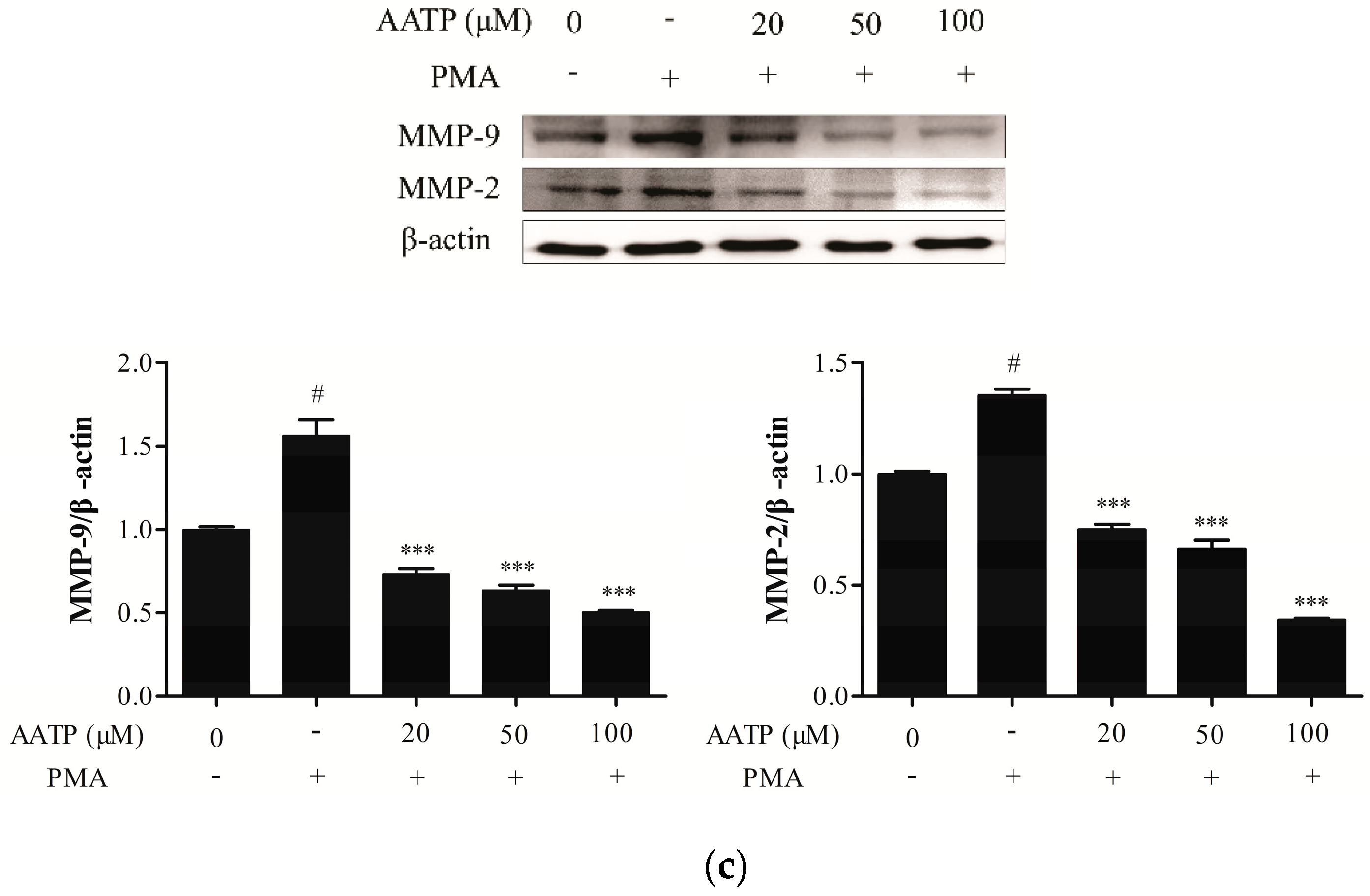
2.3. AATP Reduces PMA-induced MMPs Expression and Suppresses Proteolytic Activities in HT1080 Cells
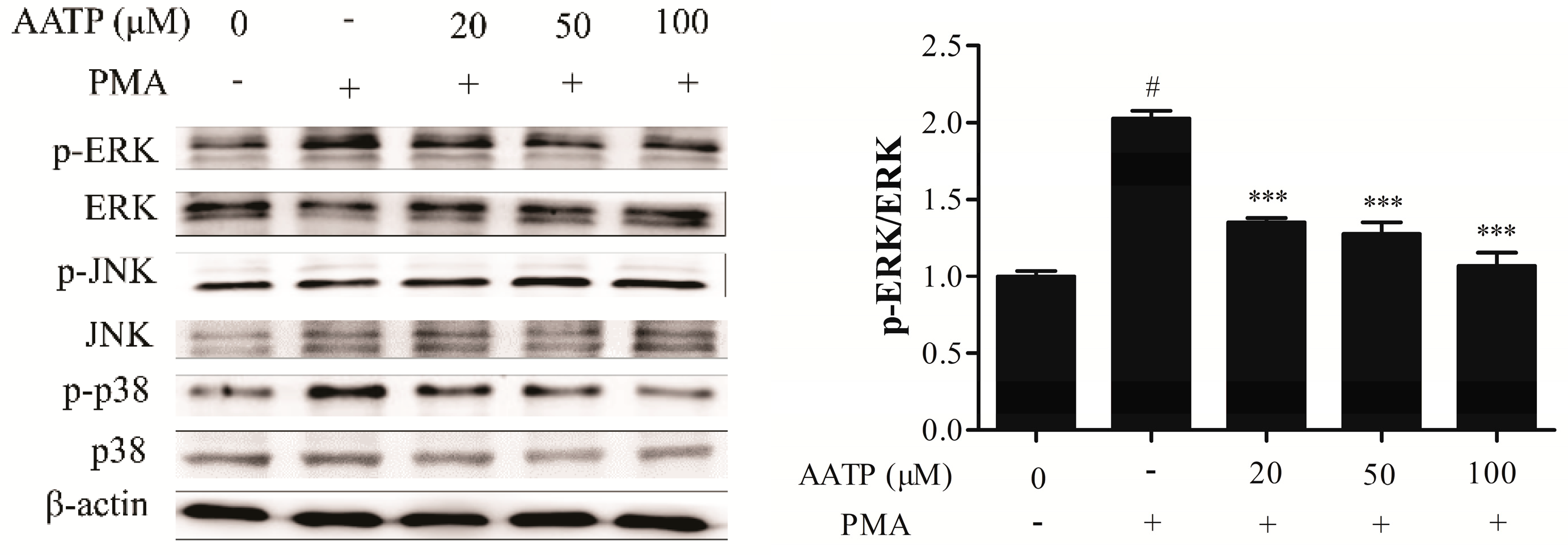
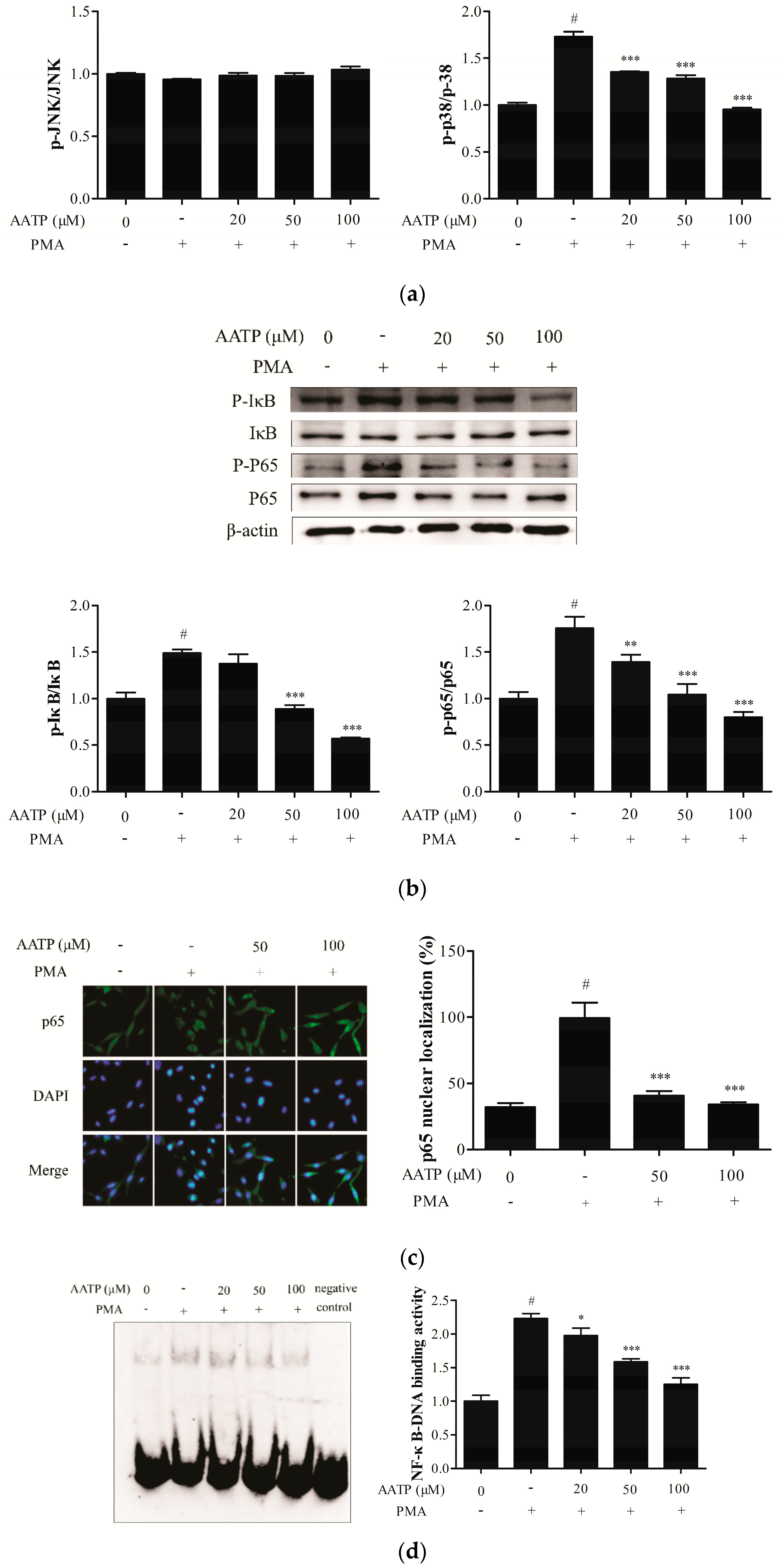
2.4. AATP Inhibits PMA-induced ERK and JNK Phosphorylation and NF-κB Activation in HT1080 Cells
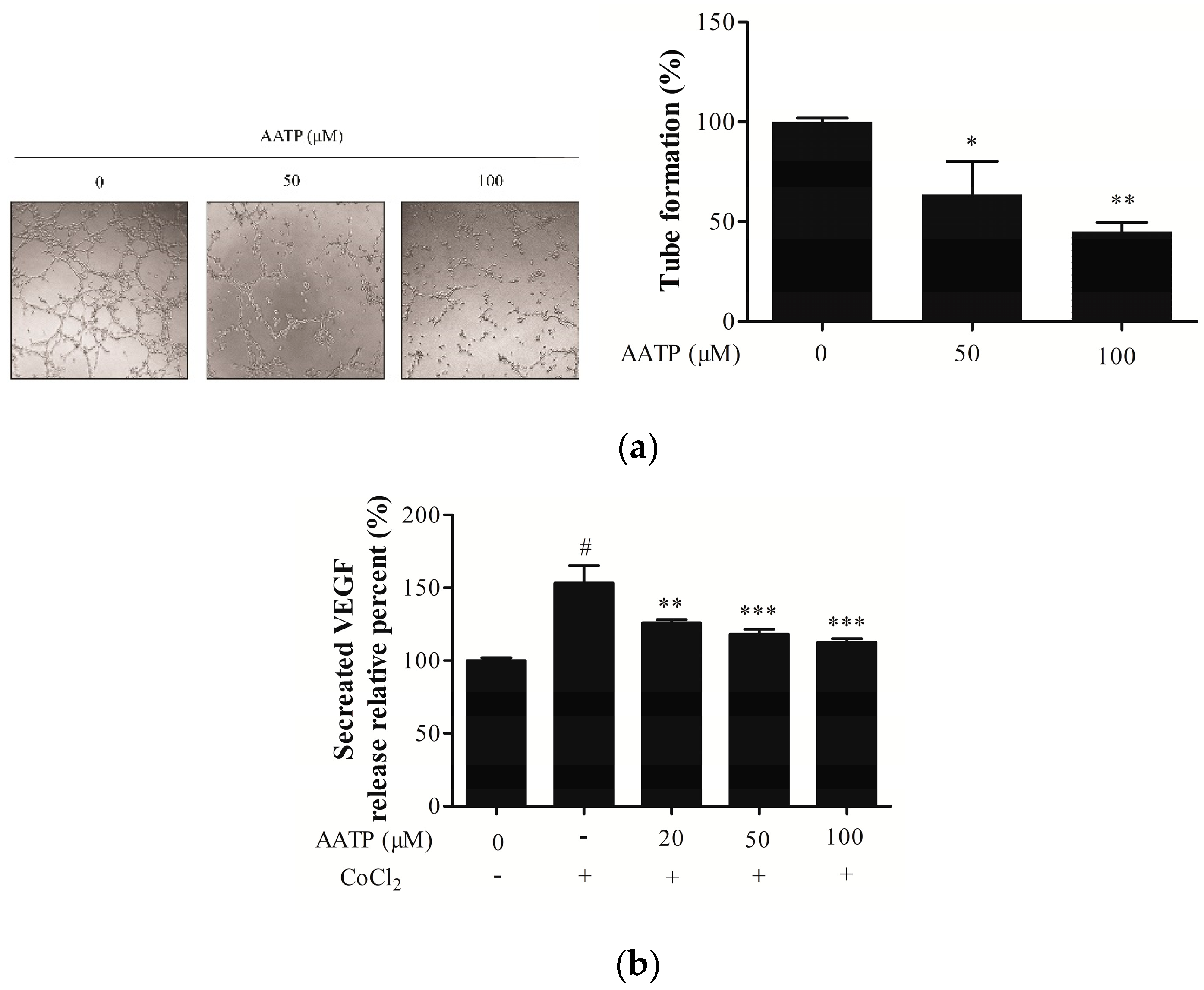
2.5. AATP Abolishes VM Formation and Inhibits Secretion of VEGF and Related Protein of Angiogenesis by Suppressing Hypoxia Inducible Factor (HIF)-1α Signal Pathway Under Hypoxic Conditions
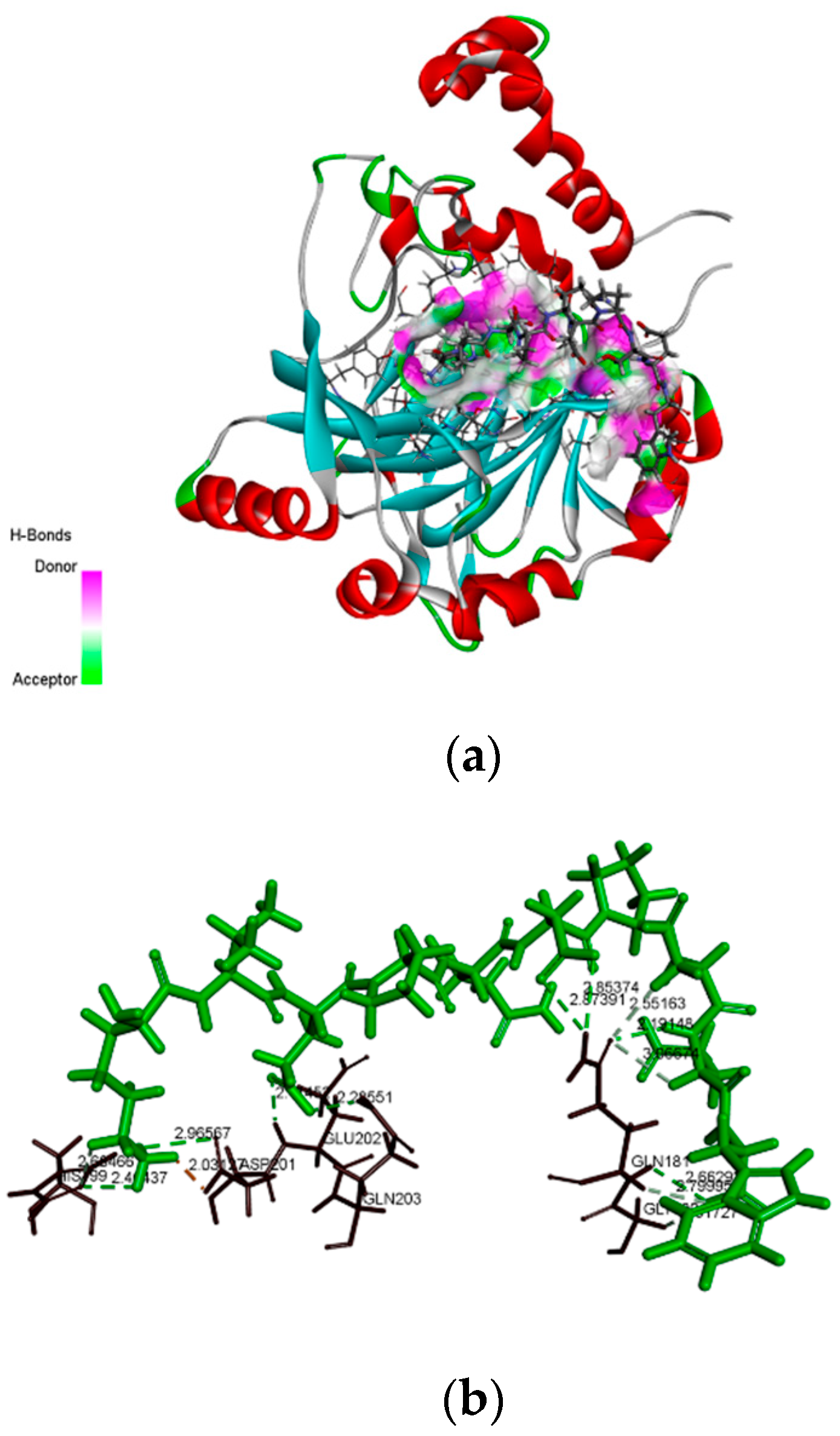
2.6. Molecular Docking Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Viability Assay
4.3. Colony Formation Assay
4.4. Cells Migration Assay
4.5. Cancer Cell Spheroid Invasion Assay
4.6. RNA Extraction and Quantitative Real Time PCR (qPCR)
4.7. Gelatinolytic Activity
4.8. Western Blotting
4.9. Immunofluorescence Staining
4.10. NF-κB Activation Assay (Electrophoretic Mobility Shift Assays, EMSA)
4.11. Assaying the Release of VEGF
4.12. Molecular Docking Analysis
4.13. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Kohandel, M.; Kardar, M.; Milosevic, M.; Sivaloganathan, S. Dynamics of tumor growth and combination of anti-angiogenic and cytotoxic therapies. Phys. Med. Biol. 2007, 52, 3665–3677. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Ma, C.; Zhang, Y.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Chen, T. A novel antimicrobial peptide, Ranatuerin-2PLx, showing therapeutic potential in inhibiting proliferation of cancer cells. Biosci. Rep. 2018, 38, BSR20180710. [Google Scholar] [CrossRef]
- Maheswaran, S.; Haber, D.A. Circulating tumor cells: A window into cancer biology and metastasis. Curr. Opin. Genet. Dev. 2010, 20, 96–99. [Google Scholar] [CrossRef]
- Sahai, E. Mechanisms of cancer cell invasion. Curr. Opin. Genet. Dev. 2005, 15, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Mook, O.R.F.; Frederiks, W.M.; Van Noorden, C.J.F. The role of gelatinases in colorectal cancer progression and metastasis. Biochim. Biophys. Acta BBA Rev. Cancer 2004, 1705, 69–89. [Google Scholar] [CrossRef]
- Li, S.; Zhang, Q.; Zhou, L.; Guan, Y.; Chen, S.; Zhang, Y.; Han, X. Inhibitory effects of compound DMBT on hypoxia-induced vasculogenic mimicry in human breast cancer. Biomed. Pharmacother. 2017, 96, 982–992. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, R.; Niwa, Y.; Simizu, S. Integrin β1 is an essential factor in vasculogenic mimicry of human cancer cells. Cancer Sci. 2018, 109, 2490–2496. [Google Scholar] [CrossRef]
- Liu, T.J.; Sun, B.C.; Zhao, X.L.; Zhao, X.M.; Sun, T.; Gu, Q.; Yao, Z.; Dong, X.Y.; Zhao, N.; Liu, N. CD133+ cells with cancer stem cell characteristics associates with vasculogenic mimicry in triple-negative breast cancer. Oncogene 2013, 32, 544–553. [Google Scholar] [CrossRef]
- Niland, S.; Komljenovic, D.; Macas, J.; Bracht, T.; Bäuerle, T.; Liebner, S.; Eble, J.A. Rhodocetin-αβ selectively breaks the endothelial barrier of the tumor vasculature in HT1080 fibrosarcoma and A431 epidermoid carcinoma tumor models. Oncotarget 2018, 9. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Semenza, G.L. HIF-1 mediates metabolic responses to intratumoral hypoxia and oncogenic mutations. J. Clin. Invest. 2013, 123, 3664–3671. [Google Scholar] [CrossRef]
- Keith, B.; Johnson, R.S.; Simon, M.C. HIF1α and HIF2α: Sibling rivalry in hypoxic tumour growth and progression. Nat. Rev. Cancer 2012, 12, 9–22. [Google Scholar] [CrossRef]
- Ghattass, K.; Assah, R.; El-Sabban, M.; Gali-Muhtasib, H. Targeting Hypoxia for Sensitization of Tumors to Radio- and Chemotherapy. Curr. Cancer Drug Targets 2013, 13, 670–685. [Google Scholar] [CrossRef]
- Lv, Y.; Zhao, S.; Han, J.; Zheng, L.; Yang, Z.; Zhao, L. Hypoxia-inducible factor-1α induces multidrug resistance protein in colon cancer. OncoTargets Ther. 2015, 1941. [Google Scholar] [CrossRef]
- Olenyuk, B.Z.; Zhang, G.-J.; Klco, J.M.; Nickols, N.G.; Kaelin, W.G.; Dervan, P.B. Inhibition of Vascular Endothelial Growth Factor with a Sequence-Specific Hypoxia Response Element Antagonist. Proc. Natl. Acad. Sci. USA 2004, 101, 16768–16773. [Google Scholar] [CrossRef]
- Seftor, R.E.B.; Seftor, E.A.; Koshikawa, N.; Meltzer, P.S.; Gardner, L.M.G.; Bilban, M.; Stetler-Stevenson, W.G.; Quaranta, V.; Hendrix, M.J.C. Cooperative Interactions of Laminin 5 γ2 Chain, Matrix Metalloproteinase-2, and Membrane Type-1-Matrix/Metalloproteinase Are Required for Mimicry of Embryonic Vasculogenesis by Aggressive Melanoma. Cancer Res. 2001, 61, 6322–6327. [Google Scholar] [CrossRef] [PubMed]
- Heitzer, E.; Haque, I.S.; Roberts, C.E.S.; Speicher, M.R. Current and future perspectives of liquid biopsies in genomics-driven oncology. Nat. Rev. Genet. 2019, 20, 71–88. [Google Scholar] [CrossRef]
- De Zoysa, M.; Nikapitiya, C.; Whang, I.; Lee, J.-S.; Lee, J. Abhisin: A potential antimicrobial peptide derived from histone H2A of disk abalone (Haliotis discus discus). Fish Shellfish Immunol. 2009, 27, 639–646. [Google Scholar] [CrossRef]
- Li, J.; Tong, T.; Ko, D.-O.; Kang, S.-G. Antithrombotic potential of extracts from abalone, Haliotis Discus Hannai Ino: In vitro and animal studies. Food Sci. Biotechnol. 2013, 22, 471–476. [Google Scholar] [CrossRef]
- Nguyen, V.-T.; Qian, Z.-J.; Jung, W.-K. Abalone Haliotis discus hannai Intestine Digests with Different Molecule Weights Inhibit MMP-2 and MMP-9 Expression in Human Fibrosarcoma Cells. Fish. Aquat. Sci. 2012, 15, 137–143. [Google Scholar] [CrossRef][Green Version]
- Qian, Z.-J.; Ryu, B.; Park, W.S.; Choi, I.-W.; Jung, W.-K. Inhibitory Effects and Molecular Mechanism of an Anti-inflammatory Peptide Isolated from Intestine of Abalone, Haliotis Discus Hannai on LPS-Induced Cytokine Production via the p-p38/p-JNK Pathways in RAW264.7 Macrophages. J. Food Nutr. Res. 2016, 4, 690–698. [Google Scholar] [CrossRef]
- Lee, C.-G.; Kwon, H.-K.; Ryu, J.H.; Kang, S.J.; Im, C.-R.; II Kim, J.; Im, S.-H. Abalone visceral extract inhibit tumor growth and metastasis by modulating Cox-2 levels and CD8+ T cell activity. BMC Complement. Altern. Med. 2010, 10. [Google Scholar] [CrossRef]
- Uchida, H.; Sasaki, T.; Uchida, N.; Takasuka, N.; Endo, Y.; Kamiya, H. Oncostatic and immunomodulatory effects of a glycoprotein fraction from water extract of abalone, Haliotis discus hannai. Cancer Immunol. Immunother. 1987, 24. [Google Scholar] [CrossRef]
- Zhu, B.-W.; Zhou, D.-Y.; Yang, J.-F.; Li, D.-M.; Yin, H.-L.; Tada, M. A neutral polysaccharide from the abalone pleopod, Haliotis discus hannai Ino. Eur. Food Res. Technol. 2009, 228, 591–595. [Google Scholar] [CrossRef]
- Sable, R.; Parajuli, P.; Jois, S. Peptides, Peptidomimetics, and Polypeptides from Marine Sources: A Wealth of Natural Sources for Pharmaceutical Applications. Mar. Drugs 2017, 15, 124. [Google Scholar] [CrossRef]
- Jayatunga, M.K.P.; Thompson, S.; Hamilton, A.D. α-Helix mimetics: Outwards and upwards. Bioorg. Med. Chem. Lett. 2014, 24, 717–724. [Google Scholar] [CrossRef]
- Hirschmann, R.F.; Nicolaou, K.C.; Angeles, A.R.; Chen, J.S.; Smith, A.B. The β- d -Glucose Scaffold as a β-Turn Mimetic. Acc. Chem. Res. 2009, 42, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Maniotis, A.J.; Folberg, R.; Hess, A.; Seftor, E.A.; Gardner, L.M.G.; Pe’er, J.; Trent, J.M.; Meltzer, P.S.; Hendrix, M.J.C. Vascular Channel Formation by Human Melanoma Cells in Vivo and in Vitro: Vasculogenic Mimicry. Am. J. Pathol. 1999, 155, 739–752. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, D.; Sun, B. Vasculogenic mimicry: Current status and future prospects. Cancer Lett. 2007, 254, 157–164. [Google Scholar] [CrossRef]
- Yan, C.; Boyd, D.D. Regulation of matrix metalloproteinase gene expression. J. Cell. Physiol. 2007, 211, 19–26. [Google Scholar] [CrossRef]
- Michel, G.; Minet, E.; Ernest, I.; Roland, I.; Durant, F.; Remacle, J.; Michiels, C. A Model for the Complex Between the Hypoxia-Inducible Factor-1 (HIF-1) and its Consensus DNA Sequence. J. Biomol. Struct. Dyn. 2000, 18, 169–179. [Google Scholar] [CrossRef]
- Madanecki, P.; Kapoor, N.; Bebok, Z.; Ochocka, R.; Collawn, J.; Bartoszewski, R. Regulation of angiogenesis by hypoxia: The role of microRNA. Cell. Mol. Biol. Lett. 2013, 18, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Folberg, R.; Hendrix, M.J.C.; Maniotis, A.J. Vasculogenic Mimicry and Tumor Angiogenesis. Am. J. Pathol. 2000, 156, 361–381. [Google Scholar] [CrossRef]
- Kim, A.; Im, M.; Gu, M.J.; Ma, J.Y. Ethanol extract of Lophatheri Herba exhibits anti-cancer activity in human cancer cells by suppression of metastatic and angiogenic potential. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Shukla, K.; Sonowal, H.; Saxena, A.; Ramana, K.V. Didymin by suppressing NF-κB activation prevents VEGF-induced angiogenesis in vitro and in vivo. Vascul. Pharmacol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Yan, J.; Fu, X.; Pan, Q.; Sun, D.; Xu, Y.; Wang, J.; Nie, L.; Tong, L.; Shen, A.; et al. Aspirin Inhibits Cancer Metastasis and Angiogenesis via Targeting Heparanase. Clin. Cancer Res. 2017, 23, 6267–6278. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.A.; Zheng, Q.; Liu, Z.; Thompson, J.E. Role of p38 and JNK MAPK signaling pathways and tumor suppressor p53 on induction of apoptosis in response to Ad-eIF5A1 in A549 lung cancer cells. Mol. Cancer. 2013, 12, 35. [Google Scholar] [CrossRef]
- Song, F.-N.; Duan, M.; Liu, L.-Z.; Wang, Z.-C.; Shi, J.-Y.; Yang, L.-X.; Zhou, J.; Fan, J.; Gao, Q.; Wang, X.-Y. RANKL Promotes Migration and Invasion of Hepatocellular Carcinoma Cells via NF-κB-Mediated Epithelial-Mesenchymal Transition. PLoS ONE. 2014, 9, e108507. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Koh, D.; Kim, G.S.; Lee, S.E.; Noh, H.J.; Kim, S.Y.; Lee, Y.H.; Lim, Y.; Shin, S.Y. 2-Hydroxy-3,4-naphthochalcone (2H-NC) inhibits TNFα-induced tumor invasion through the downregulation of NF-κB-mediated MMP-9 gene expression. Bioorg. Med. Chem. Lett. 2015, 25, 128–132. [Google Scholar] [CrossRef]
- Lu, Y.; Jeong, Y.-T.; Li, X.; Kim, M.J.; Park, P.-H.; Hwang, S.-L.; Son, J.K.; Chang, H.W. Emodin Isolated from Polygoni cuspidati Radix Inhibits TNF-α and IL-6 Release by Blockading NF-κB and MAP Kinase Pathways in Mast Cells Stimulated with PMA Plus A23187. Biomol. Ther. 2013, 21, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Tong, C.; Liu, Y.; Wang, J.; Ni, X.; Xiong, M. Ginkgetin inhibits growth of breast carcinoma via regulating MAPKs pathway. Biomed. Pharmacother. 2017, 96, 450–458. [Google Scholar] [CrossRef]
- Nguyen, V.-T.; Qian, Z.-J.; Lee, B.; Heo, S.-J.; Kim, K.-N.; Jeon, Y.-J.; Park, W.S.; Choi, I.-W.; Jang, C.H.; Ko, S.-C.; et al. Fucoxanthin derivatives from Sargassum siliquastrum inhibit matrix metalloproteinases by suppressing NF-κB and MAPKs in human fibrosarcoma cells. ALGAE 2014, 29, 355–366. [Google Scholar] [CrossRef]
- Zhu, B.-W.; Wang, L.-S.; Zhou, D.-Y.; Li, D.-M.; Sun, L.-M.; Yang, J.-F.; Wu, H.-T.; Zhou, X.-Q.; Tada, M. Antioxidant activity of sulphated polysaccharide conjugates from abalone (Haliotis discus hannai Ino). Eur. Food Res. Technol. 2008, 227, 1663–1668. [Google Scholar] [CrossRef]
- Wu, Z.-Z.; Ding, G.-F.; Huang, F.-F.; Yang, Z.-S.; Yu, F.-M.; Tang, Y.-P.; Jia, Y.-L.; Zheng, Y.-Y.; Chen, R. Anticancer Activity of Anthopleura anjunae Oligopeptides in Prostate Cancer DU-145 Cells. Mar. Drugs 2018, 16, 125. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Funct. Foods. 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Huang, F.; Yang, Z.; Yu, D.; Wang, J.; Li, R.; Ding, G. Sepia Ink Oligopeptide Induces Apoptosis in Prostate Cancer Cell Lines via Caspase-3 Activation and Elevation of Bax/Bcl-2 Ratio. Mar. Drugs 2012, 10, 2153–2165. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer ID | Primer Sequence (5′→3′) |
|---|---|
| MMP-1 | F: 5′-AGCCATCACTTACCTTGCACTGAG-3′ |
| R: 5′-RCCACATCAGGCACTCCACATCTG-3′ | |
| MMP-2 | F: 5′-AGCCAAGCGGTCTAAGTCCAGAG-3′ |
| R: 5′-GGAATGAAGCACAGCAGGTCTCAG-3′ | |
| MMP-3 | F: 5′-ACGCACAGCAACAGTAGGATTGG-3′ |
| R: 5′-GAGGCAGGCAAGACAGCAAGG-3′ | |
| MMP-9 | F: 5′-TCCTGGTGCTCCTGGTGCTG-3′ |
| R: 5′-CTGCCTGTCGGTGAGATTGGTTC-3′ | |
| MMP-13 | F: 5′-AGTCATGGAGCTTGCTGCATTCTC-3′ |
| R: 5′-TCCTGGCTGCCTTCCTCTTCTTG-3′ | |
| β-actin | F: 5′-CCTGGCACCCAGCACAAT-3′ |
| R: 5′-GGGCCGGACTCGTCATAC-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, F.; Chen, M.-F.; Zhang, Y.-Y.; Li, C.-Y.; Zhou, C.-X.; Hong, P.-Z.; Sun, S.-L.; Qian, Z.-J. A Novel Peptide from Abalone (Haliotis discus hannai) to Suppress Metastasis and Vasculogenic Mimicry of Tumor Cells and Enhance Anti-Tumor Effect In Vitro. Mar. Drugs 2019, 17, 244. https://doi.org/10.3390/md17040244
Gong F, Chen M-F, Zhang Y-Y, Li C-Y, Zhou C-X, Hong P-Z, Sun S-L, Qian Z-J. A Novel Peptide from Abalone (Haliotis discus hannai) to Suppress Metastasis and Vasculogenic Mimicry of Tumor Cells and Enhance Anti-Tumor Effect In Vitro. Marine Drugs. 2019; 17(4):244. https://doi.org/10.3390/md17040244
Chicago/Turabian StyleGong, Fang, Mei-Fang Chen, Yuan-Yuan Zhang, Cheng-Yong Li, Chun-Xia Zhou, Peng-Zhi Hong, Sheng-Li Sun, and Zhong-Ji Qian. 2019. "A Novel Peptide from Abalone (Haliotis discus hannai) to Suppress Metastasis and Vasculogenic Mimicry of Tumor Cells and Enhance Anti-Tumor Effect In Vitro" Marine Drugs 17, no. 4: 244. https://doi.org/10.3390/md17040244
APA StyleGong, F., Chen, M.-F., Zhang, Y.-Y., Li, C.-Y., Zhou, C.-X., Hong, P.-Z., Sun, S.-L., & Qian, Z.-J. (2019). A Novel Peptide from Abalone (Haliotis discus hannai) to Suppress Metastasis and Vasculogenic Mimicry of Tumor Cells and Enhance Anti-Tumor Effect In Vitro. Marine Drugs, 17(4), 244. https://doi.org/10.3390/md17040244