Antifungal and Antioxidant Properties of Chitosan Polymers Obtained from Nontraditional Polybius henslowii Sources

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Polybius henslowii Characterization
2.2. Chitin Extraction Optimization
2.3. Chitosan and Water-Soluble Chitosan Characterization
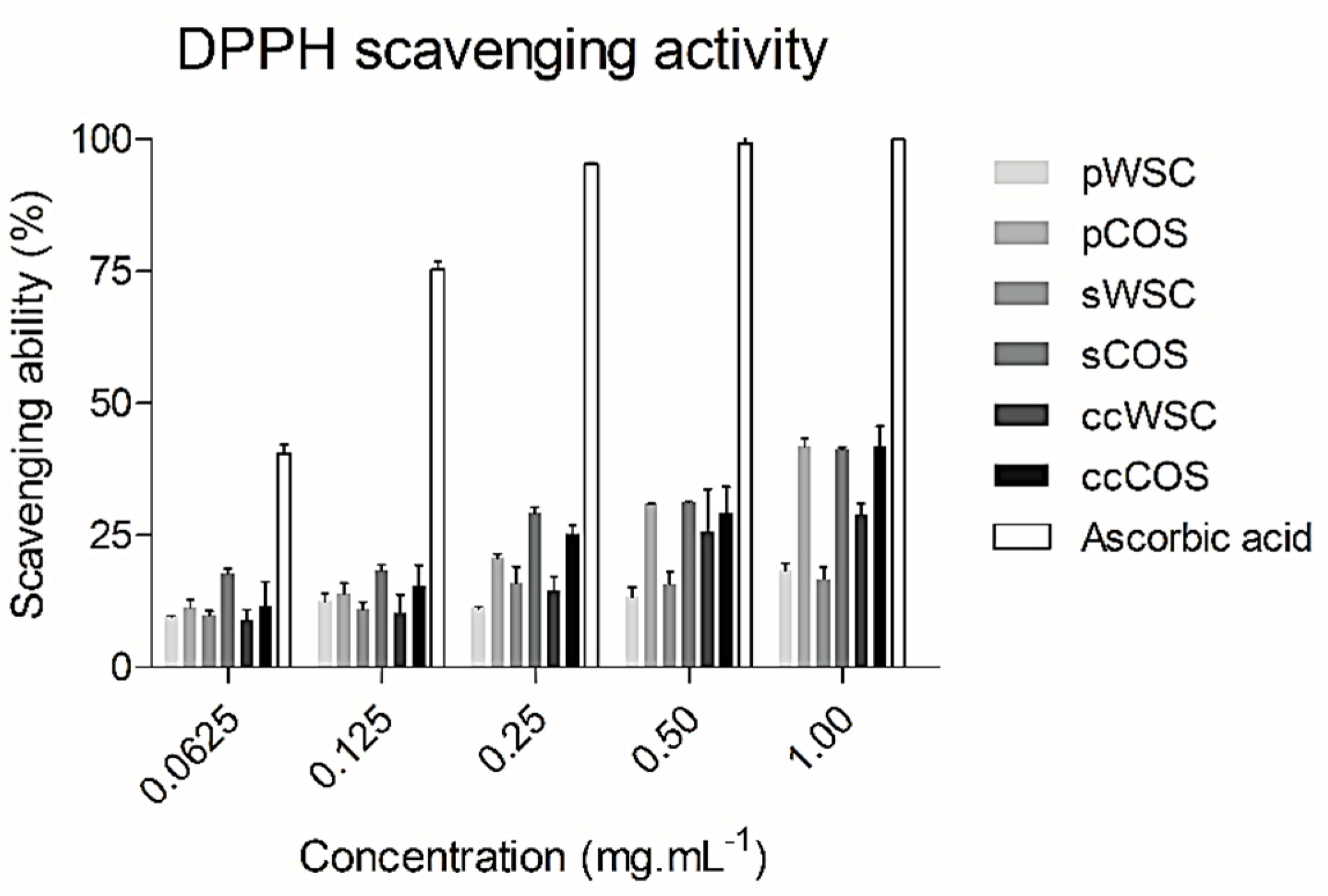
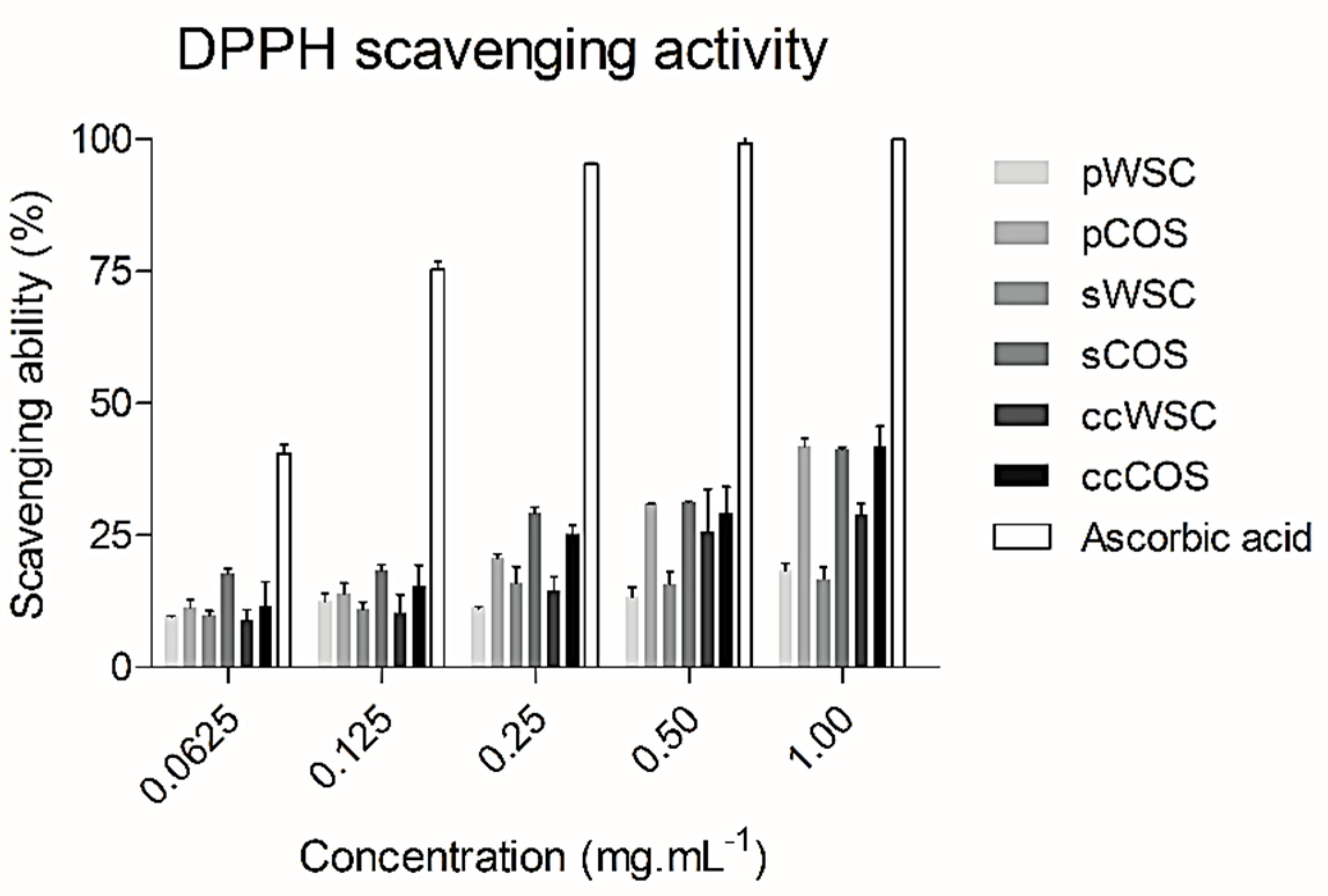
2.4. DPPH Radical Scavenging Activity
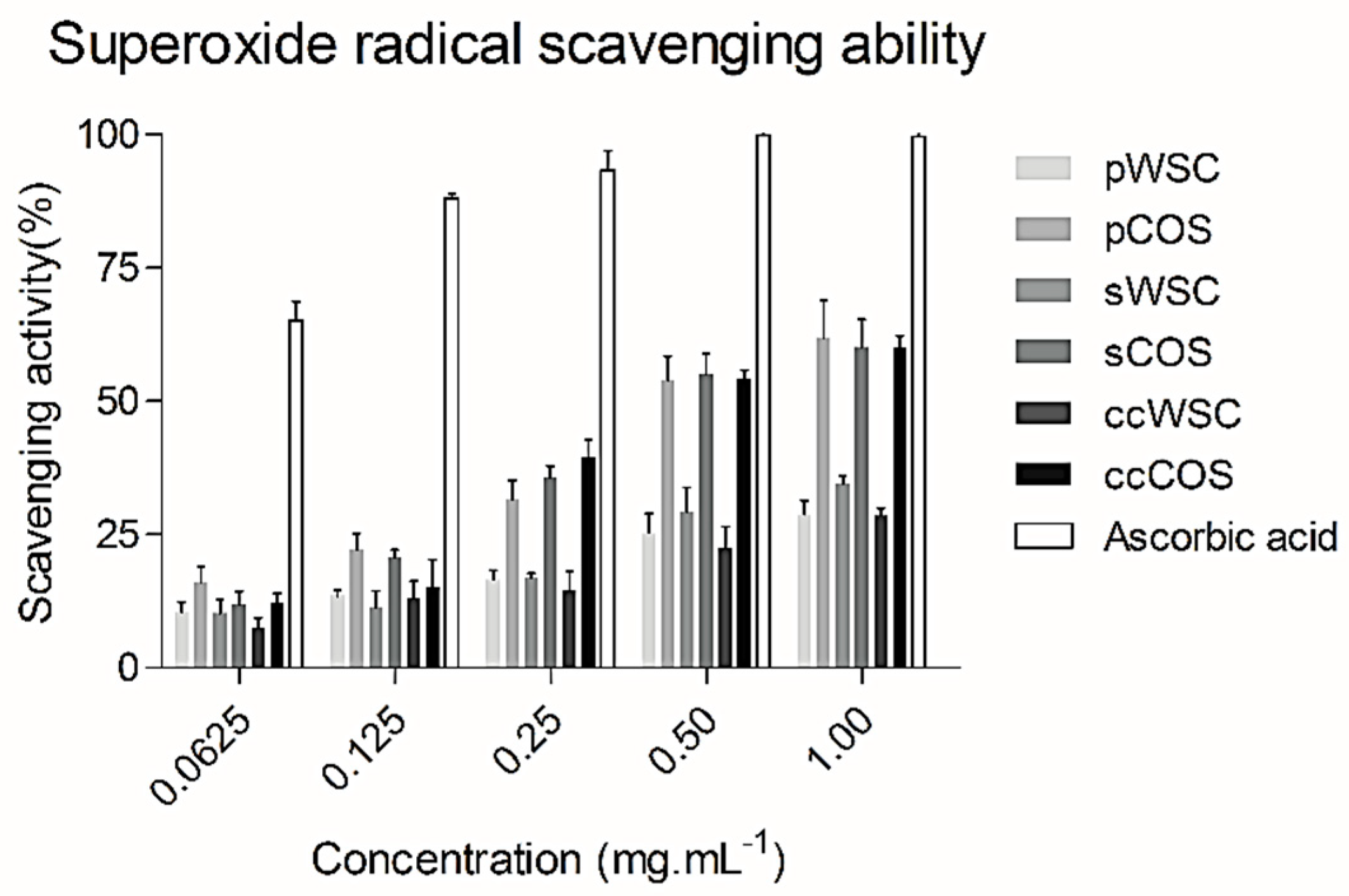
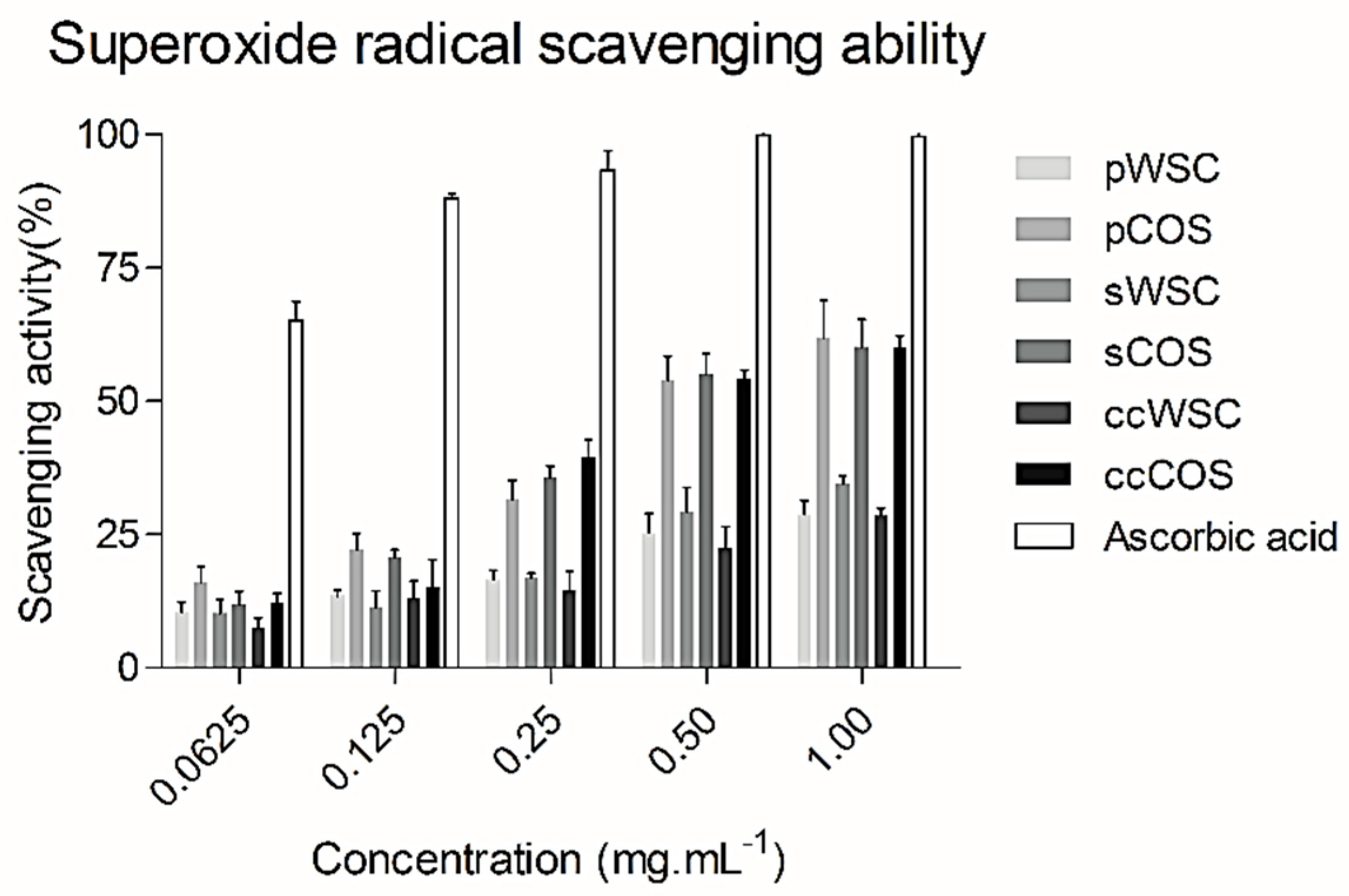
2.5. Superoxide Radical (O2−) Scavenging Ability
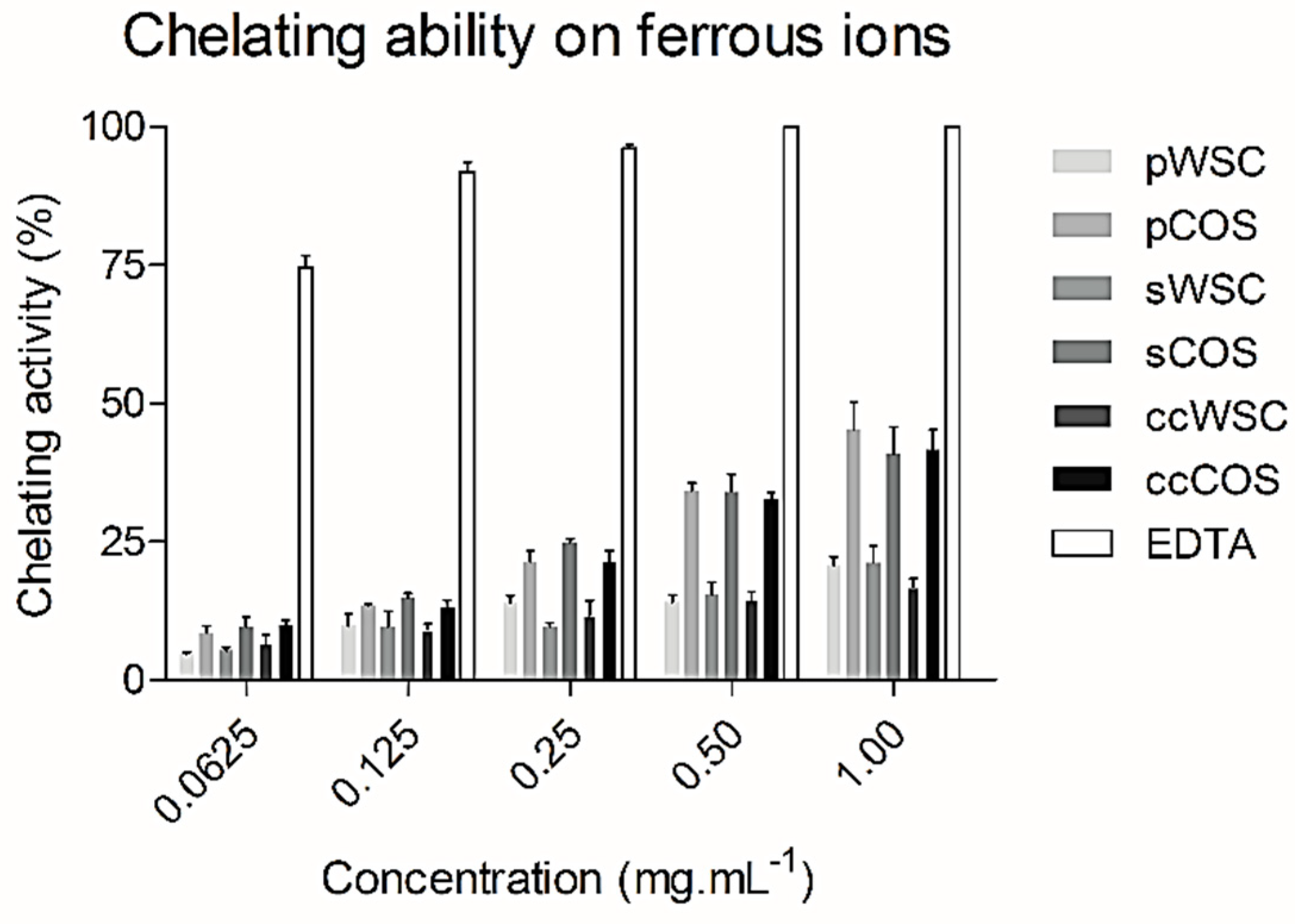
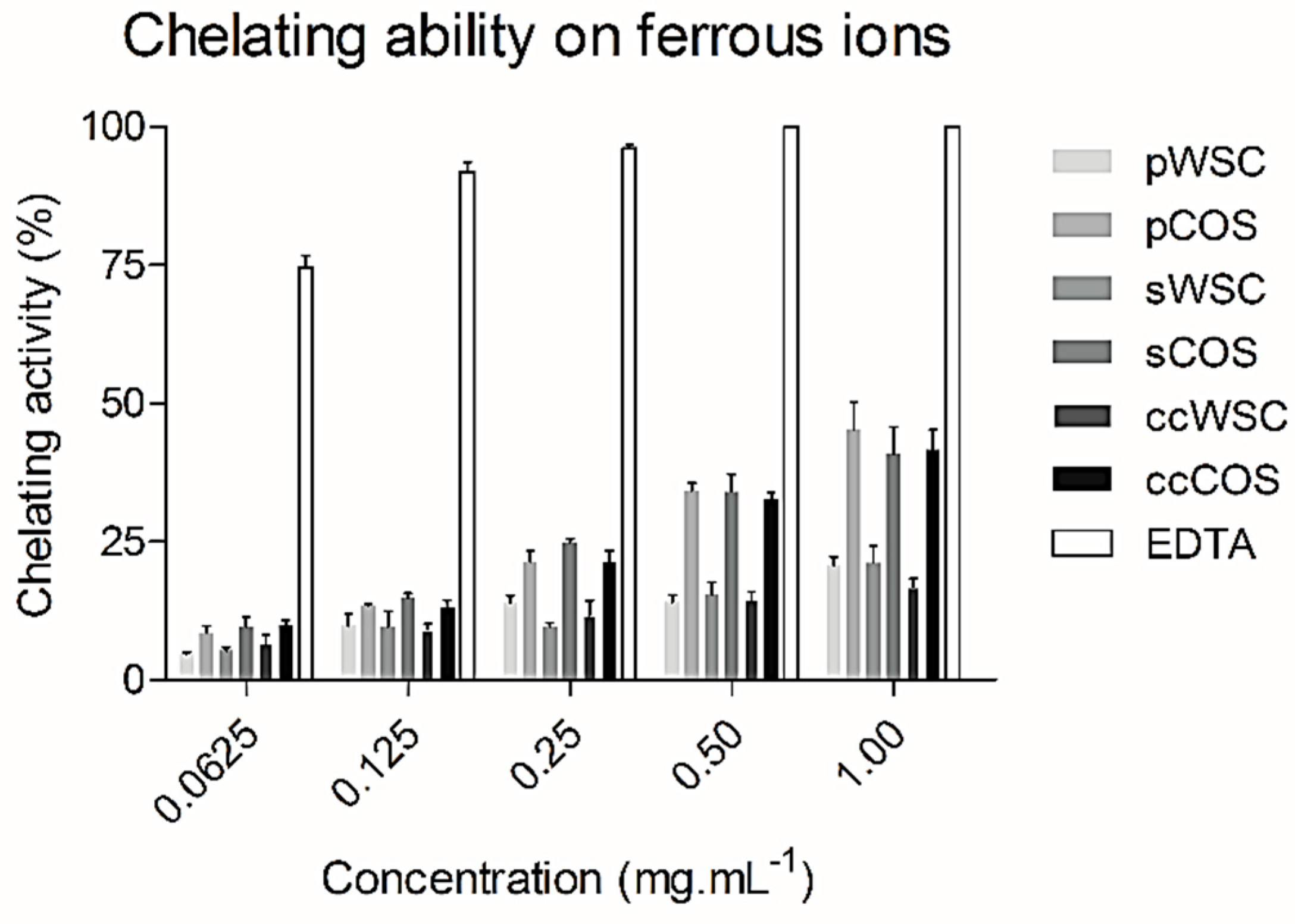
2.6. Chelating Ability on Ferrous Ions
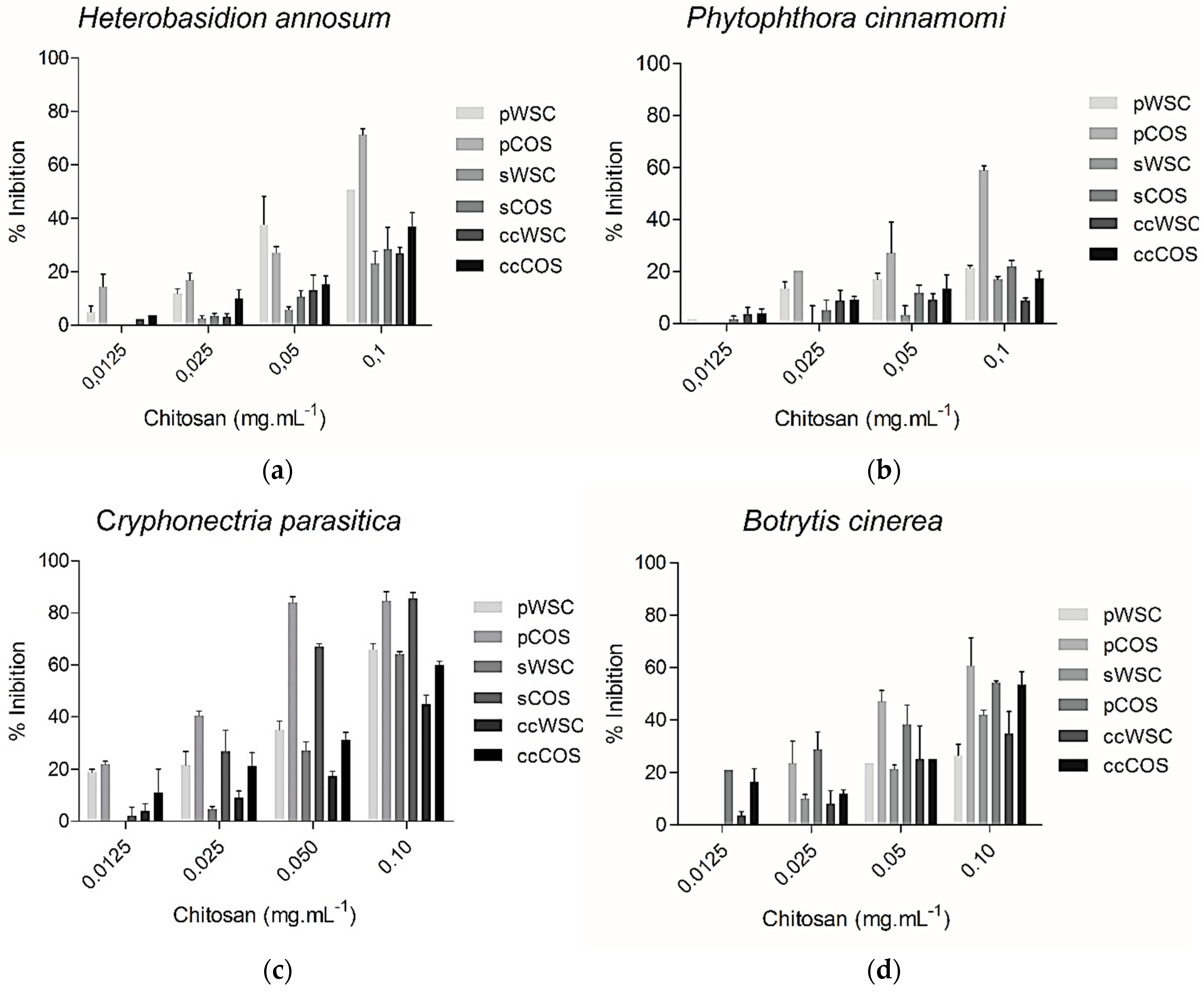
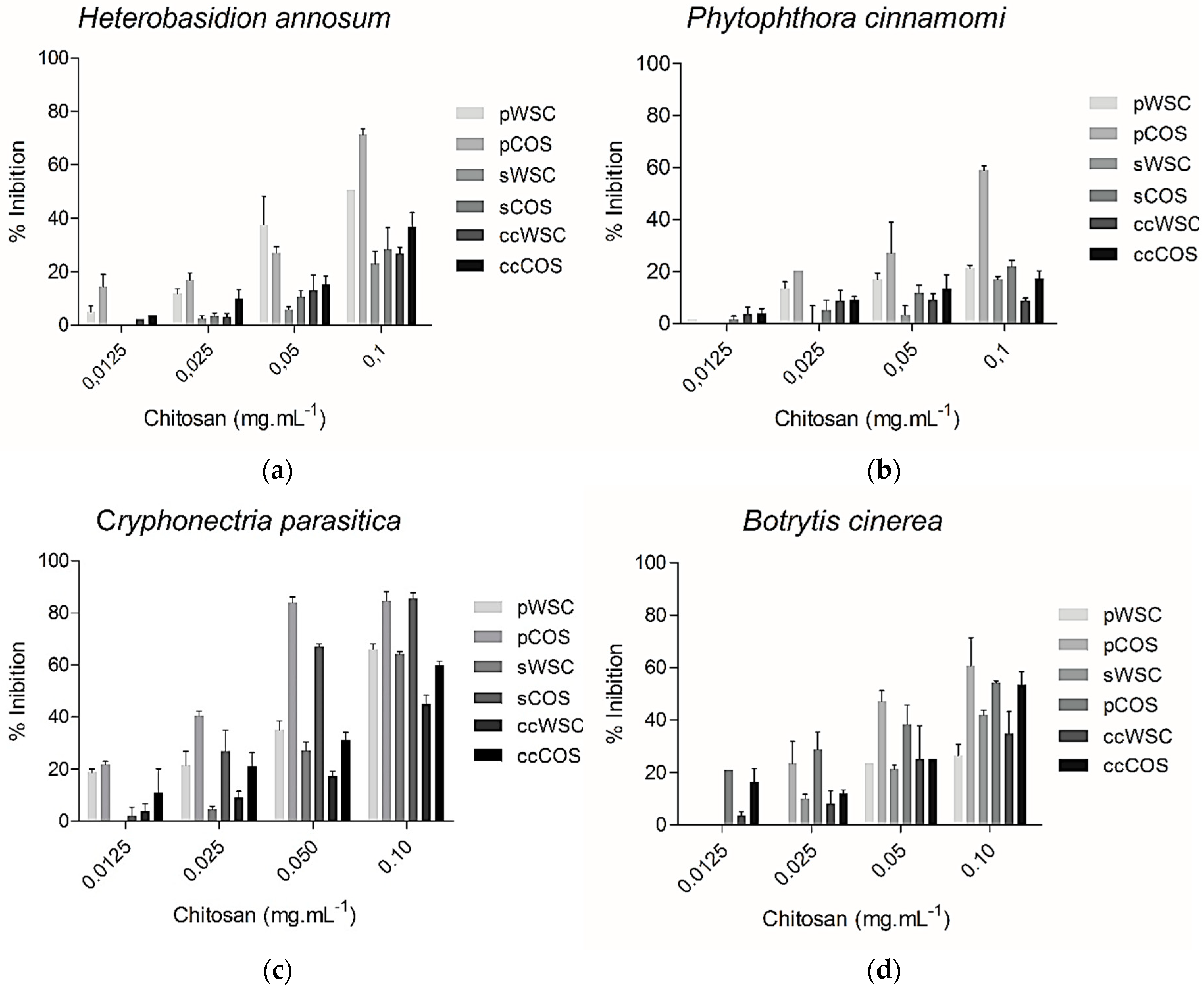
2.7. Antifungal Assay
3. Experimental Section
3.1. Biochemical Characterization of Raw Materials
3.2. Chitin Extraction and Deacetylation
3.3. Chitooligosaccharides and Water-Soluble Chitosan Production
3.4. Chitosan Product Characterization
3.5. Scavenging of 1,1-diphenyl-2-picrylhydrazyl Radicals
3.6. Superoxide Radical (O2−) Scavenging Activity
3.7. Chelating Ability on Ferrous Ions
3.8. Effect of Chitosan on Mycelial Growth
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moreira, A.C.; Caetano, P.; Correia, S.; Brasier, C.M.; Ferraz, J.F.P. Phytophthora species associated with cork oak decline in a Mediterranean forest in in southern Portugal. In Proceedings of the 6th International Congress of Plant Pathology, Montréal, QC, Canada, 28 July–6 August 1993. [Google Scholar]
- Droby, S.; Lichter, A. Post-Harvest Botrytis Infection: Etiology, Development and Management. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 349–367. ISBN 978-1-4020-2626-3. [Google Scholar]
- Nuss, D.L. Biological control of chestnut blight: An example of virus-mediated attenuation of fungal pathogenesis. Microbiol. Rev. 1992, 56, 561–576. [Google Scholar]
- Niemelä, T.; Korhonen, K. Taxonomy of the genus Heterobasidion. In Heterobasidion Annosum: Biology, Ecology, 1988, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hu¨ttermann, A., Eds.; CAB International: Wallingford, NY, USA, 2000; pp. 27–33. [Google Scholar]
- Smilanick, J.L.; Harvey, J.M.; Hartsell, P.L.; Henson, D.J.; Harris, C.M.; Fouse, D.C.; Assemi, M. Influence of sulfur dioxide fumigant dose on residues and control of postharvest decay of grapes. Plant Dis. J. 1990, 74, 418–421. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.; Rochetti, R.; Stanic, V.; Weckx, M. Methods for the determination of the degree of acetylation of chitin and chitosan. In Chitin Handbook; Muzzarelli, R.A.A., Peter, M.G., Eds.; ATEC: College Park, MD, USA, 1997; pp. 109–119. [Google Scholar]
- Dutta, P.K.; Tripathi, S.; Mehrotra, G.K.; Dutta, J. Perspectives for chitosan based antimicrobial films in food applications. Food Chem. 2009, 114, 1173–1182. [Google Scholar] [CrossRef]
- Kumar, R.M.N.V. A review of chitin and chitosan applications. Reactive and Functional. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar] [CrossRef]
- Cerdá, C.; Sánchez, C.; Climent, B.; Vázquez, A.; Iradi, A.; El Amrani, F.; Bediaga, A.; Sáez, G.T. Oxidative stress and DNA damage in obesity-related tumorigenesis. Adv. Exp. Med. Biol. 2014, 824, 5–17. [Google Scholar] [PubMed]
- Wölfle, U.; Seelinger, G.; Bauer, G.; Meinke, M.C.; Lademann, J.; Schempp, C.M. Reactive molecule species and antioxidative mechanisms in normal skin and skin aging. Ski. Pharmacol. Physiol. 2014, 27, 316–332. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Hirose, M.; Fukushima, H.; Tsuda, T.; Shirai, T.; Tatenatsu, M. Studies on antioxidants: Their carcinogenic and modifying effects on chemical carcinogens. Food Chem. Toxicol. 1986, 24, 1071–1092. [Google Scholar] [CrossRef]
- Ferraro, V.; Cruz, I.B.; Jorge, R.F.; Malcata, F.X.; Pintado, M.E.; Castro, P.M.L. Valorisation of natural extracts from marine source focused on marine by-products: A review. Food Res. Int. 2010, 43, 2221–2233. [Google Scholar] [CrossRef] [Green Version]
- Cartes, J.E.; Abelló, P.; Lloris, D.; Carbonell, A.; Torres, P.; Maynou, F.; Sola, L.G.D. Feeding guilds of western Mediterranean demersal fish and crustaceans: An analysis based on a spring survey. Sci. Mar. 2002, 590, 209–220. [Google Scholar] [CrossRef]
- Serrano, A.; Sánchez, F.; Punzón, A.; Velasco, F.; Olaso, I. Deep sea megafaunal assemblages off the northern Iberian slope related to environmental factors. Sci. Mar. 2011, 75, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Hayward, P.J.; Ryland, J.S. Handbook of the Marine Fauna of North–West Europe; Oxford University Press: Oxford, UK, 1995; p. 454. [Google Scholar]
- Signa, G.; Cartes, J.E.; Solé, M.; Serrano, A.; Sánchez, F. Trophic ecology of the swimming crab Polybius henslowii Leach, 1820 in Galician and Cantabrian Seas: Influences of natural variability and the Prestige oil spill. Cont. Shelf Res. 2008, 28, 2659–2667. [Google Scholar] [CrossRef]
- González-Gurriarán, E. El patexo, Polybius henslowii Leach (Decapoda, Brachyura), en las costas de Galicia (NW de Espanã): I. Distribución espacial y cambios temporales. Investig. Pesq. 1987, 51, 361–374. [Google Scholar]
- Zariquey, R. Crustáceos decápodos ibéricos. Investig. Pesq. 1968, 32, 83. [Google Scholar]
- Abdou, E.S.; Nagy, K.S.A.; Elsabee, M.Z. Extraction and characterization of chitin and chitosan from local sources. Bioresour. Technol. 2008, 99, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Krzeczkowski, R.A.; Tenney, R.D.; Kelley, C. Alaska king crab: Fatty acid composition, carotenoid index and proximate analysis. J. Food Sci. 2006, 36, 604–606. [Google Scholar] [CrossRef]
- King, I.; Childs, M.T.; Dorsett, C.; Ostrander, J.G.; Monsen, E.R. Shellfish: Proximate composition, minerals, fatty acids, and sterols. J. Am. Diet. Assoc. 1990, 90, 677–685. [Google Scholar]
- Nalan, G.; Dlua, K.; Yerlikayaa, P. Determination of proximate and mineral contents of blue crab (Callinectes sapidus) and swim crab (Portunus pelagicus) caught of the Gulf of Antalya. Food Chem. 2003, 80, 495–498. [Google Scholar] [CrossRef]
- Naczk, M.; Williams, J.; Brennan, K.; Liyanapathirana, C.; Shahidi, F. Compositional characteristics of green crab (Carcinus maenas). Food Chem. 2004, 88, 429–434. [Google Scholar] [CrossRef]
- Venugopal, V. Marine Polysaccharides: Food Applications; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2011; Volume 3, pp. 61–84. [Google Scholar]
- Youn, D.K.; No, H.K.; Prinyawiwatkul, W. Physicochemical and functional properties of chitosans prepared from shells of crabs harvested in three different years. Carbohydr. Polym. 2009, 78, 41–45. [Google Scholar] [CrossRef]
- Mahmoud, N.S.; Ghaly, A.E.; Arab, F. Unconventional Approach for Demineralization of Deproteinized Crustacean Shells for Chitin Production. Am. J. Biochem. Biotechnol. 2007, 3, 1–9. [Google Scholar] [CrossRef]
- No, H.K.; Meyers, S.P. Preparation and characterization of chitin and chitosan: A review. J. Aquat. Food Prod. Technol. 1995, 4, 27–52. [Google Scholar] [CrossRef]
- Benhabile, M.S.; Salah, R.; Lounici, H.; Drouiche, N.; Goosen, M.F.A.; Mameri, N. Antibacterial activity of chitin, chitosan and its oligomers prepared from shrimp shell waste. Food Hydrol. 2012, 29, 48–56. [Google Scholar] [CrossRef]
- Yen, M.-T.; Yang, J.-H.; Mau, J.-L. Physicochemical characterization of chitin and chitosan from crab shells. Carbohydr. Polym. 2009, 75, 15–21. [Google Scholar] [CrossRef]
- Al-Sagheer, F.A.; Al-Sughayer, M.A.; Muslim, S.; Elsabee, M.Z. Extraction and characterization of chitin and chitosan from marine sources in Arabian Gulf. Carbohydate. Polymerst 2009, 77, 410–419. [Google Scholar]
- Barroca-Aubry, N.; Pernet-Poil-Chevrier, A.; Domard, A.; Trombotto, S. Towards a modular synthesis of well-defined chitooligosaccharides: Synthesis of the four chitodisaccharides. Carbohydr. Res. 2010, 345, 1685–1697. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Y.; Dai, S.; Yang, B. Preparation of water-soluble chitosan from shrimp shell and its antibacterial activity. Innov. Food Sci. Emerg. Technol. 2009, 10, 103–107. [Google Scholar] [CrossRef]
- Tian, F.; Liu, Y.; Hu, K.; Zhao, B. Study of the depolymerization behavior of chitosan by hydrogen peroxide. Carbohydr. Polym. 2004, 57, 31–37. [Google Scholar] [CrossRef]
- Lu, S.; Song, X.; Cao, D.; Chen, Y.; Yao, K. Preparation of Water-Soluble Chitosan. J. Appl. Polym. Sci. 2004, 685, 3497–3503. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Takamura, H.; Matoba, T.; Terao, J. HPLC method for evaluation of the free radical-scavenging activity of foods by using 1,1,-diphenyl-2-picrylhydrazyl. Biosci. Biotechnol. Biochem. 1998, 62, 1201–1204. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W.; Thomas, R.L. Antioxidative activity of chitosan’s with varying molecular weight. Food Chem. 2007, 101, 308–313. [Google Scholar] [CrossRef]
- Samar, M.M.; El-Kalyoubi, M.H.; Khalaf, M.M.; El-Razik, M.M.A. Physicochemical, functional, antioxidant and antibacterial properties of chitosan extracted from shrimp wastes by microwave technique. Ann. Agric. Sci. 2013, 58, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Xu, P.; Liu, Q. Antioxidant activity of water-soluble chitosan derivatives. Bioorgan. Med. Chem. Lett. 2001, 11, 1699–1701. [Google Scholar] [CrossRef]
- Je, J.Y.; Kim, S.K. Reactive oxygen species scavenging activity of aminoderivatized chitosan with different 651 degree of deacetylation. Bioorgan. Med. Chem. 2006, 14, 5989–5994. [Google Scholar] [CrossRef]
- Chien, P.J.; Sheu, F.; Huang, W.T.; Su, M.S. Antioxidant Polymers: Synthesis, Properties, and Applications. Food Chem. 2007, 102, 1192. [Google Scholar] [CrossRef]
- Yen, M.-T.; Yang, J.-H.; Mau, J.-L. Antioxidant properties of chitosan from crab shells. Carbohydr. Polym. 2008, 74, 840–844. [Google Scholar] [CrossRef]
- Korycka-Dahl, M.; Richardson, T. Photogeneration of superoxide anion in serum of bovine milk and in model systems containing riboflavin and amino acids. J. Dairy Sci. 1978, 61, 400–407. [Google Scholar] [CrossRef]
- Bloknina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef]
- Meyer, A.S.; Isaksen, A. Application of enzymes as food antioxidants. Trends Food Sci. Technol. 1995, 6, 300. [Google Scholar] [CrossRef]
- Feng, T.; Du, Y.M.; Li, J.; Hu, Y.; Kennedy, J.F. Enhancement of antioxidant activity of chitosan by irradiation. Carbohydr. Polym. 2008, 73, 126–132. [Google Scholar] [CrossRef]
- Xing, R.; Liu, S.; Guo, Z.; Yu, H.; Wang, P.; Li, C.; Li, Z.; Li, P. Relevance of molecular weight of chitosan and its derivatives and their antioxidant activities in vitro. Bioorgan. Med. Chem. 2005, 13, 1573–1577. [Google Scholar] [CrossRef]
- Yang, Y.; Shu, R.; Shao, J.; Xu, G.; Gu, X. Radical scavenging activity of chitooligosaccharide with different molecular weights. Eur. Food Res. Technol. 2006, 222, 36–40. [Google Scholar] [CrossRef]
- Charernsriwilaiwat, N.; Opanasopit, P.; Rojanarata, T.; Ngawhirunpat, T. In Vitro Antioxidant Activity of Chitosan Aqueous Solution: Effect of Salt Form. Trop. J. Pharm. Res. 2012, 11, 235–242. [Google Scholar] [CrossRef]
- Halliwell, B.; Aeschbach, R.; Löliger, J.; Aruoma, O.I. The characterization of antioxidants. Food Chem. Toxicol. 1995, 33, 601–617. [Google Scholar] [CrossRef]
- Tikhonov, V.E.; Radigina, L.A.; Yamskov, Y.A. Metal-chelating chitin derivatives via reaction of chitosan with nitrilotriacetic acid. Carbohydr. Res. 1996, 290, 33–41. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Chou, C.-C. Antioxidative activities of water-soluble disaccharide chitosan derivatives. Food Res. Int. 2004, 37, 883–889. [Google Scholar] [CrossRef]
- Milosavljevic, N.B.; Ristica, M.D.; Peric-Grujic, A.A.; Filipovic, J.M.; Strbacb, S.B.; Rakocevic, Z.L.; Krusic, M.T.K. Sorption of zinc by novel pH-sensitive hydrogels based on chitosan, itaconic acid and methacrylic acid. J. Hazard. Mater. 2011, 192, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Kurita, K. Chitin and Chitosan: Functional biopolymers from marine crustaceans. Mar. Biotechnol. 2006, 8, 203–226. [Google Scholar] [CrossRef]
- Qin, Y. The chelating properties of chitosan fibers. J. Appl. Polym. Sci. 1993, 49, 727–731. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef]
- Liu, J.; Tian, S.; Meng, X.; Xu, Y. Effects of chitosan on control of postharvest diseases and physiological responses of tomato fruit. Postharvest Biol. Technol. 2007, 44, 300–306. [Google Scholar] [CrossRef]
- Skjak-Braek, G.; Anthonsen, T.; Sandford, P.A. Chitin and Chitosan: Sources, Chemistry, Biochemistry, Physical Properties and Applications; Springer: Dordrecht, The Netherlands, 1989; ISBN 978-1-85166-395-8. [Google Scholar]
- Jeon, Y. Antimicrobial effect of chitooligosaccharides produced by bioreactor. Carbohydr. Polym. 2001, 44, 71–76. [Google Scholar] [CrossRef]
- No, H.K.; Young Park, N.; Ho Lee, S.; Meyers, S.P. Antibacterial activity of chitosans and chitosan oligomers with different molecular weights. Int. J. Food Microbiol. 2002, 74, 65–72. [Google Scholar] [CrossRef]
- Singh, T.; Vesentini, D.; Singh, A.P.; Daniel, G. Effect of chitosan on physiological, morphological, and ultrastructural characteristics of wood-degrading fungi. Int. Biodeterior. Biodegrad. 2008, 62, 116–124. [Google Scholar] [CrossRef]
- Ing, L.Y.; Zin, N.M.; Sarwar, A.; Katas, H. Antifungal Activity of Chitosan Nanoparticles and Correlation with Their Physical Properties. J. Appl. Polym. Sci. 2012, 2012, 632698. [Google Scholar] [CrossRef] [PubMed]
- Vallapa, N.; Wiarachai, O.; Thongchul, N.; Pan, J.; Tangpasuthadol, V.; Kiatkamjornwong, S.; Hoven, V.P. Enhancing antibacterial activity of chitosan surface by heterogeneous quaternization. Carbohydr. Polym. 2011, 83, 868–875. [Google Scholar] [CrossRef]
- Wiarachai, O.; Thongchul, N.; Kiatkamjornwong, S.; Hoven, V.P. Surface-quaternized chitosan particles as an alternative and effective organic antibacterial material. Colloids Surf. 2012, 92, 121–129. [Google Scholar] [CrossRef]
- Guo, Z.; Xing, R.; Liu, S.; Zhong, Z.; Ji, X.; Wang, L.; Li, P. Antifungal properties of Schiff bases of chitosan, 629 N-substituted chitosan and quaternized chitosan. Carbohydr. Res. 2007, 342, 1329–1332. [Google Scholar] [CrossRef]
- Galván Márquez, I.; Akuaku, J.; Cruz, I.; Cheetham, J.; Golshani, A.; Smith, M.L. Disruption of protein synthesis as antifungal mode of action by chitosan. Int. J. Food Microbiol. 2013, 164, 108–112. [Google Scholar] [CrossRef]
- Johnson, M.K. Variable Sensitivity in Microbiuret Assay of Protein. Anal. Biochem. 1978, 86, 320–323. [Google Scholar] [CrossRef]
- ISO 1444 (1996); International Organisation for Standardisation: Geneva, Switzerland, 1996.
- Haastert-Talini, K.; Geuna, S.; Dahlin, L.B.; Meyer, C.; Stenberg, L.; Freier, T.; Heimann, C.; Barwig, C.; Pinto, L.F.V.; Raimondo, S.; et al. Chitosan tubes of varying degrees of acetylation for bridging peripheral nerve defects. Biomaterials 2013, 34, 9886–9904. [Google Scholar] [CrossRef] [Green Version]
- Brugnerotto, J.; Lizardi, J.; Goycoolea, F.M.; Argüelles-Monal, W.; Desbrières, J.; Rinaudo, M. An infrared investigation in relation with chitin and chitosan characterization. Polymer 2001, 42, 3569–3580. [Google Scholar] [CrossRef]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Nishikimi, M. Oxidation of ascorbic acid with superoxide anion generated by the xanthine-xanthine oxidase system. Biochem. Biophys. Res. Commun. 1975, 63, 463–468. [Google Scholar] [CrossRef]
- Decker, E.A.; Welch, B. Role of ferritin as a lipid oxidation catalyst in muscle food. J. Agric. Food Chem. (USA) 1990, 38, 674–677. [Google Scholar] [CrossRef]
- El Ghaouth, A.; Arul, J.; Grenier, J.; Asselin, A. Antifungal activity of chitosan on two post-harvest 618 pathogens of strawberry fruits. Phytopathology 1992, 82, 398–402. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Material | Protein (%) | Ash (%) | Lipids (%) | Chitin (%) |
|---|---|---|---|---|
| Shell | 32.1 ± 6.68 | 44.5 ± 0.57 | 13.2 ± 0.25 | 9.7 ± 0.57 |
| Pereopods | 16.6 ± 1.21 | 49.3 ± 5.86 | 1.6 ± 0.14 | 11.4 ± 0.19 |
| NaOH/HCl | Shells Samples | |||
| Protein Content (%) | Protein Removal (%) | Ash Content (%) | Ash Removal (%) | |
| 1 M | 2.0 ± 0.12 | 96.1 ± 0.25 | 0.8 ± 0.01 | 98.2 ± 0.02 |
| 0.75 M | 2.3 ± 0.14 | 95.35 ± 0.28 | 1.0 ± 0.05 | 97.8 ± 0.11 |
| 0.5 M | 2.29 ± 0.15 | 95.39 ± 0.31 | 1.2 ± 0.17 | 97.3 ± 0.38 |
| NaOH/HCl | Pereopods Samples | |||
| Protein Content (%) | Protein Removal (%) | Ash Content (%) | Ash Removal (%) | |
| 1 M | 1.2 ± 0.12 | 92.2 ± 0.78 | 0.4 ± 0.19 | 99.1 ± 0.4 |
| 0.75 M | 1.5 ± 0.07 | 90.5 ± 0.48 | 0.5 ± 0.01 | 98.9 ± 0.8 |
| 0.5 M | 1.8 ± 0.06 | 88.2 ± 0.43 | 0.7 ± 0.05 | 98.6 ± 0.1 |
| Chitosan Products | Yield (%) | Dynamic Viscosity (cP) | DD (%) | Mw (kg·mol−1) |
|---|---|---|---|---|
| Pereopods chitosan | 9.7 ± 0.62 | 749.2 ± 62.69 | 94.3 ± 0.04 | 378.2 ± 78.00 |
| pWSC | - | - | 62 ± 0.53 | 404.0 ± 45.00 |
| pCOS | - | - | 93.3 ± 0.04 | 7.4 ± 1.20 |
| Shells chitosan | 8.0 ± 0.24 | 417.2 ± 94.99 | 95.1 ± 0.01 | 247.0 ± 31.20 |
| sWSC | - | - | 55.0 ± 3.21 | 279.0 ± 33.00 |
| sCOS | - | - | 95.0 ± 0.62 | 2.7 ± 0.40 |
| Commercial chitosan | - | - | 87.0 | 780.0 |
| ccWSC | - | - | 57.0 ± 0.83 | 775.0 ± 42.00 |
| ccCOS | - | - | 86.0 ± 1.4 | 10.4 ± 0.70 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avelelas, F.; Horta, A.; Pinto, L.F.V.; Cotrim Marques, S.; Marques Nunes, P.; Pedrosa, R.; Leandro, S.M. Antifungal and Antioxidant Properties of Chitosan Polymers Obtained from Nontraditional Polybius henslowii Sources. Mar. Drugs 2019, 17, 239. https://doi.org/10.3390/md17040239
Avelelas F, Horta A, Pinto LFV, Cotrim Marques S, Marques Nunes P, Pedrosa R, Leandro SM. Antifungal and Antioxidant Properties of Chitosan Polymers Obtained from Nontraditional Polybius henslowii Sources. Marine Drugs. 2019; 17(4):239. https://doi.org/10.3390/md17040239
Chicago/Turabian StyleAvelelas, Francisco, André Horta, Luís F.V. Pinto, Sónia Cotrim Marques, Paulo Marques Nunes, Rui Pedrosa, and Sérgio Miguel Leandro. 2019. "Antifungal and Antioxidant Properties of Chitosan Polymers Obtained from Nontraditional Polybius henslowii Sources" Marine Drugs 17, no. 4: 239. https://doi.org/10.3390/md17040239
APA StyleAvelelas, F., Horta, A., Pinto, L. F. V., Cotrim Marques, S., Marques Nunes, P., Pedrosa, R., & Leandro, S. M. (2019). Antifungal and Antioxidant Properties of Chitosan Polymers Obtained from Nontraditional Polybius henslowii Sources. Marine Drugs, 17(4), 239. https://doi.org/10.3390/md17040239