Sargassum muticum and Osmundea pinnatifida Enzymatic Extracts: Chemical, Structural, and Cytotoxic Characterization

 ,
,  ,
, 
 ,
,  ,
,  ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Results and Discussion
2.1. Elemental Inorganic and Organic Composition of Seaweeds’ Enzymatic Extracts
2.2. Monosaccharides and Free Amino Acids
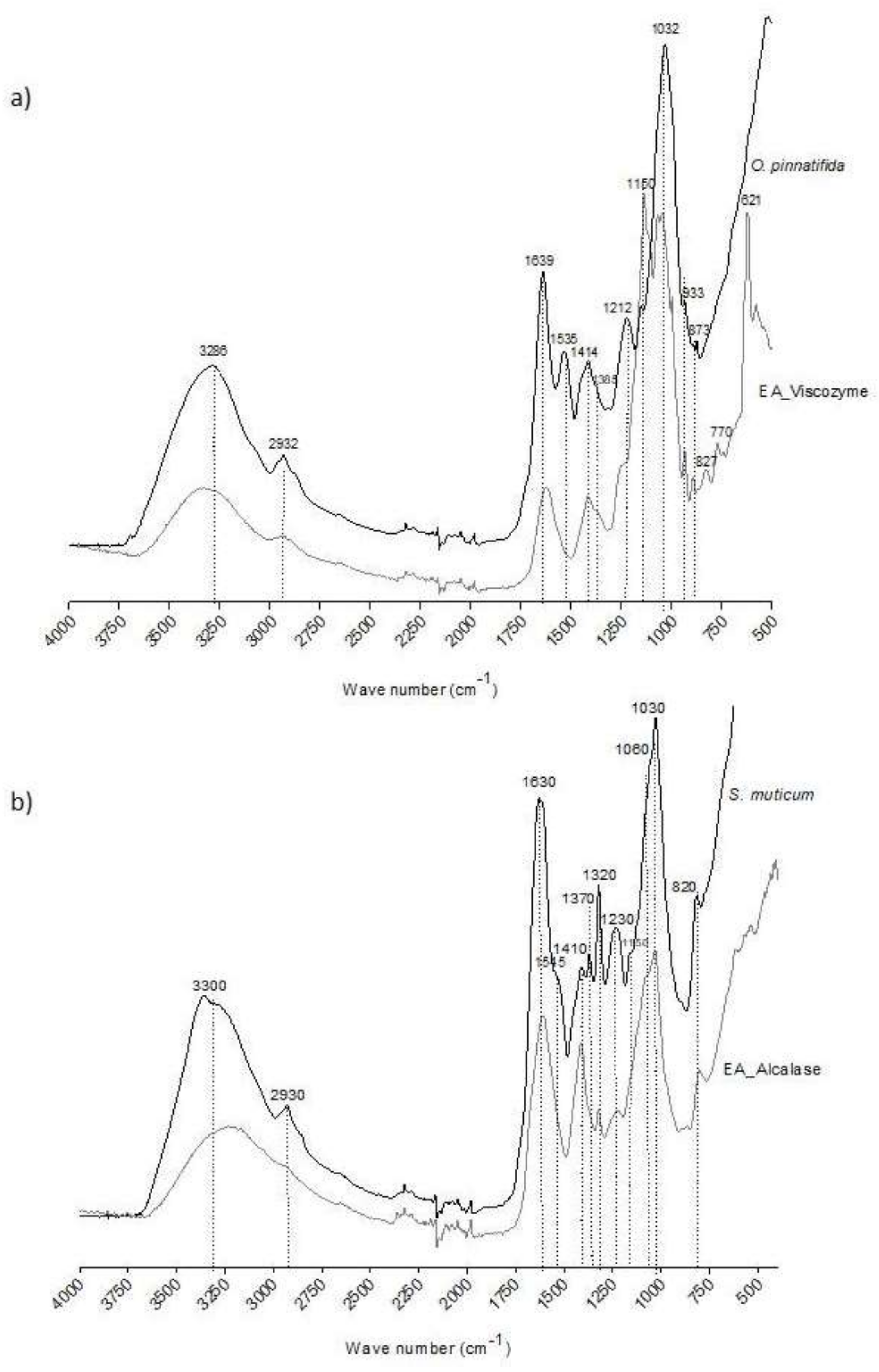
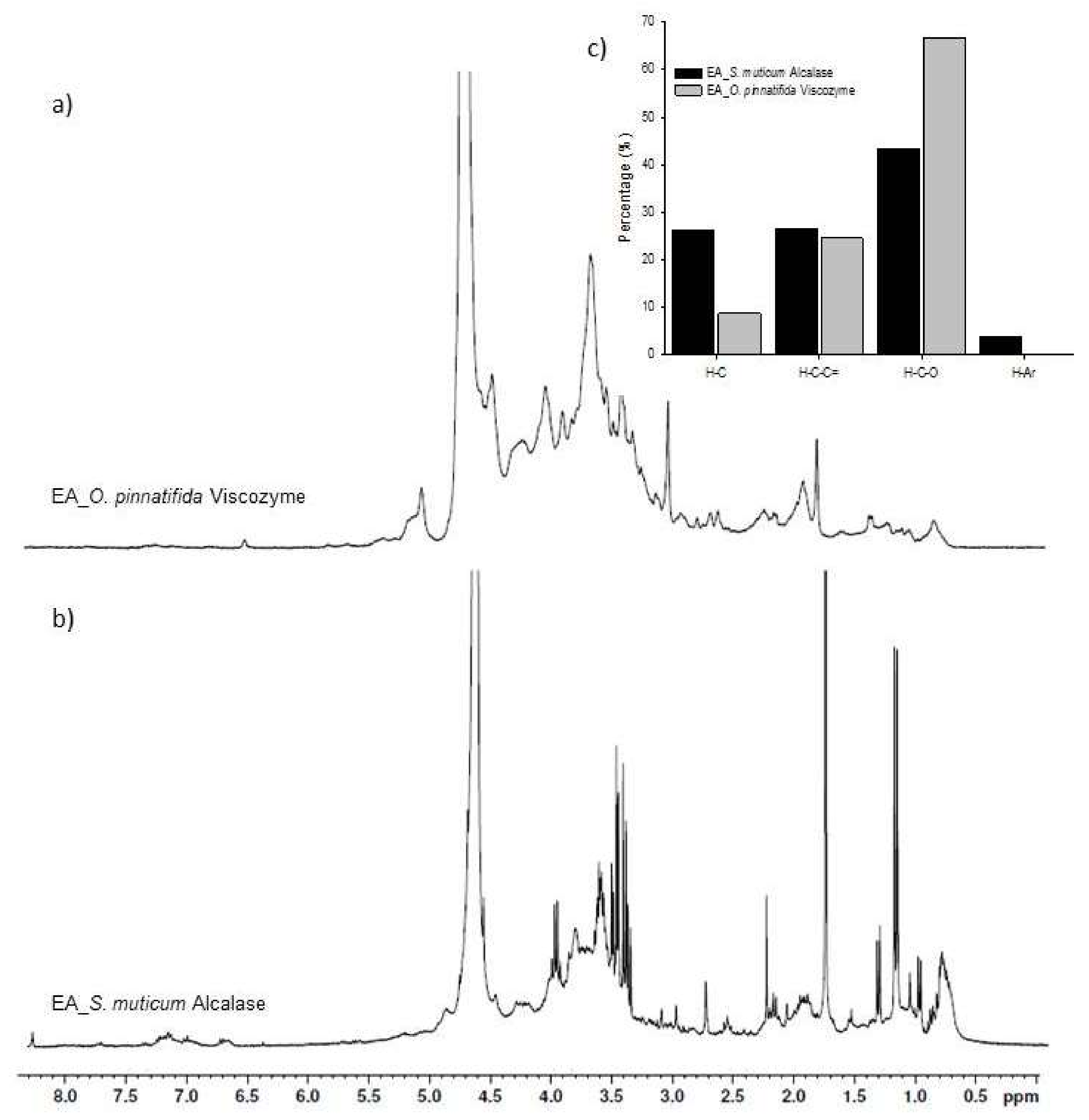
2.3. Structural Characterization of Seaweed Extracts
- 1)
- δH = 0.6–1.8 ppm: Aliphatic protons, H–C; -CH > -CH2 > -CH3.
- 2)
- δH = 1.8–3.2 ppm: Protons bound to carbon atoms in the alpha position to unsaturated groups in allylic (H-Cα-C=), carbonyl, or imino (H-Cα-C=O or H-Cα-C=N) groups, and protons in secondary and tertiary amines (H-C-NR2 and NR3, respectively).
- 3)
- δH = 3.2–4.1 ppm: Aliphatic protons on carbon atoms singly bound to oxygen atoms (H-C-O-CO-R > H-C-OH or H-C-O-C) in alcohols, polyols, ethers, and esters.
- 4)
- δH = 6.5–8.5 ppm (aromatic protons).
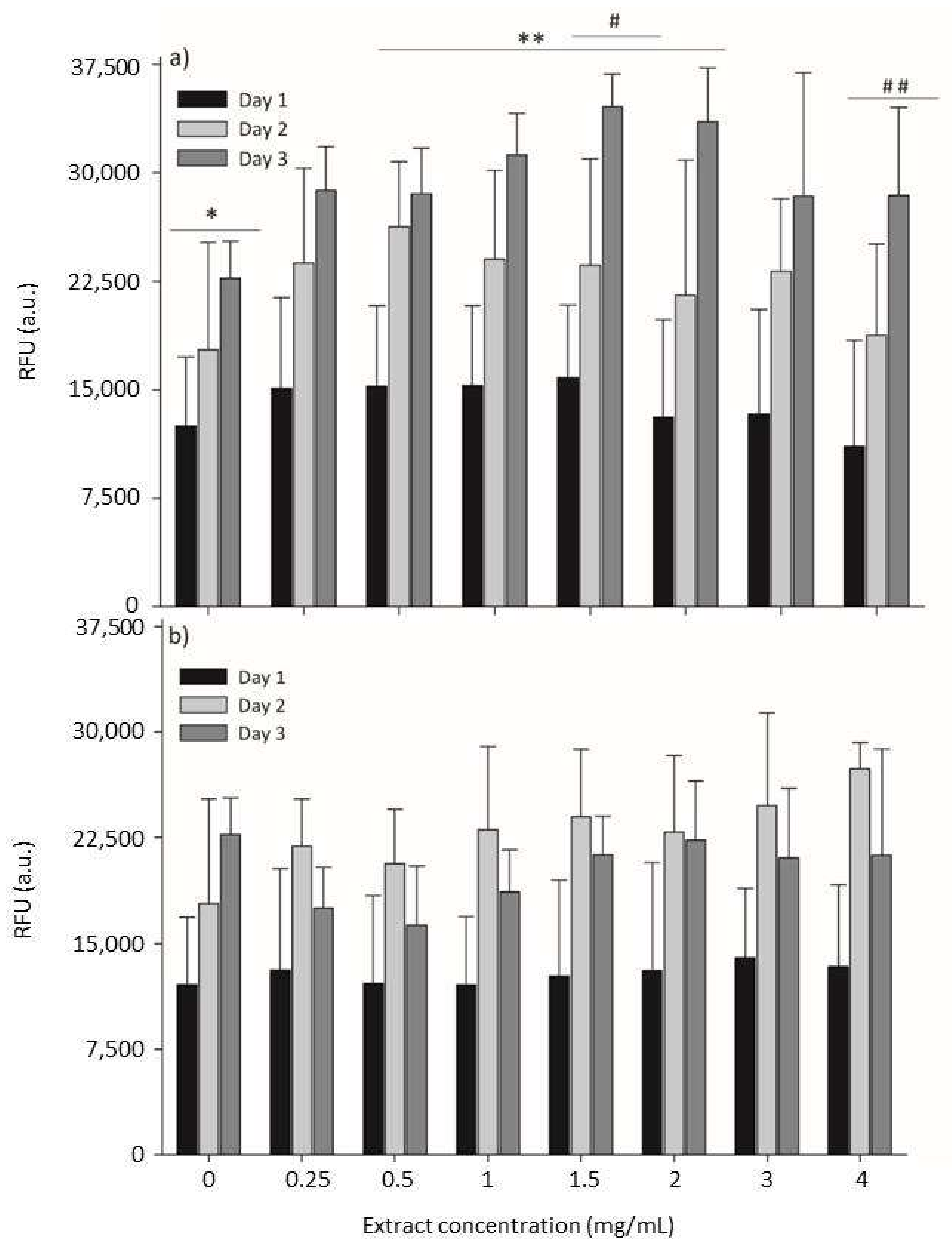
2.4. Cytotoxicity Evaluation
3. Materials and Methods
3.1. Seaweed Extracts
3.2. Chemical Characterization
3.2.1. Elemental Analysis
3.2.2. Analysis of Monosaccharide Composition
3.2.3. Analysis of Amino Acids
3.2.4. FTIR-ATR Analysis
3.2.5. 1H NMR Analysis
3.3. Cytotoxicity Assessment
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Singh, R.P.; Reddy, C.R.K. Unraveling the functions of the macroalgal microbiome. Front. Microbiol. 2016, 6, 1488. [Google Scholar] [CrossRef] [PubMed]
- Gil-Chávez, G.J.; Villa, J.A.; Ayala-Zavala, J.F.; Heredia, J.B.; Sepulveda, D.; Yahia, E.M.; González-Aguilar, G.A. Technologies for extraction and production of bioactive compounds to be used as nutraceuticals and food ingredients: An Overview. Compr. Rev. Food Sci. Food Saf. 2013, 12, 5–23. [Google Scholar] [CrossRef]
- Rodrigues, D.; Sousa, S.; Silva, A.G.; Amorim, M.; Pereira, L.; Rocha-Santos, T.A.P.; Gomes, A.M.; Duarte, A.C.; Freitas, A.C. Impact of Enzyme- and Ultrasound-Assisted Extraction Methods on Biological Properties of Red, Brown, and Green Seaweeds from the Central West Coast of Portugal. J. Agric. Food Chem. 2015, 63, 3177–3188. [Google Scholar] [CrossRef]
- Milledge, J.J.; Nielsen, B.V.; Bailey, D. High-value products from macroalgae: The potencial uses of invasive brown seaweed, Sargassum muticum. Rev. Environ. Sci. Biotechnol. 2016, 15, 67–88. [Google Scholar] [CrossRef]
- Rodrigues, D.; Freitas, A.C.; Pereira, L.; Rocha-Santos, T.A.P.; Vasconcelos, M.W.; Roriz, M.; Rodríguez-Alcalá, L.M.; Gomes, A.M.P.; Duarte, A.C. Chemical composition of red, brown and green macroalgae from Buarcos bay in Central West Coast of Portugal. Food Chem. 2015, 183, 197–207. [Google Scholar] [CrossRef]
- Pérez, F.; Garaulet, M.; Gil, A.; Zamora, S. Calcio, fósforo, magnesio y flúor. Metabolismo óseo y su regulación. In Tratado de Nutrición, Vol. I; Gil, A., Ed.; Grupo Acción Médica: Madrid, Spain, 2005; pp. 897–925. [Google Scholar]
- Allen, L.; de Benoist, B.; Dary, O.; Hurrel, R. Guidelines on Food Fortification with Micronutrientes; World Health Organization and Food and Agriculture Organization of the United Nations: Geneva, Switzerland, 2006; ISBN 92-4-159401-2. [Google Scholar]
- Balboa, E.M.; Conde, E.; Moure, A.; Falque, E.; Dominguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef] [PubMed]
- Dore, C.M.; Alves, M.G.F.; Will, L.S.; Costa, T.G.; Sabry, D.A.; de Souza, R.L.A.; Accardo, C.M.; Rocha, H.A.; Filgueira, L.G.; Leite, E.L. A sulfated polysaccharide, fucans, isolated from brown algae Sargassum vulgare with anticoagulant, antithrombotic, antioxidant and anti-inflammatory effects. Carbohydr. Polym. 2013, 91, 467–475. [Google Scholar] [CrossRef]
- Bilan, M.I.; Usov, A.I. Structural analysis of fucoidans. Nat. Prod. Commun. 2008, 3, 1639–1648. [Google Scholar] [CrossRef]
- Duarte, M.E.R.; Cardoso, M.A.; Noseda, M.D.; Cerezo, A.S. Structural studies on fucoidans from the brown seaweed Sargassum stenophyllum. Carbohydr. Res. 2001, 333, 281–293. [Google Scholar] [CrossRef]
- García-Ríos, V.; Ríos-Leal, E.; Robledo, D.; Freile-Pelegrin, Y. Polysaccharides composition from tropical brown seaweeds. Phycol. Res. 2012, 60, 305–315. [Google Scholar] [CrossRef]
- Henrotin, Y.; Chevalier, X.; Herrero-Beaumont, G.; McAlindon, T.; Mobasheri, A.; Pavelka, K.; Biesalski, H. Physiological effects of oral glucosamine on joint health: Current status and consensus on future research priorities. BMC Res. Notes 2013, 6, 115. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Waszkuc, T.; Mohammed, F. Determination of glucosamine in raw materials and dietary supplements containing glucosamine sulfate and/or glucosamine hydrochloride by high-performance liquid chromatography with FMOC-Su derivatization: Collaborative Study. J. AOAC Int. 2005, 88, 1048–1058. [Google Scholar]
- Canelón, D.J.; Ciancia, M.; Suárez, A.I.; Compagnone, R.S.; Matulewicz, M.C. Structure of highly substituted agarans from the red seaweeds Laurencia obtusa and Laurencia filiformis. Carbohydr. Polym. 2014, 30, 705–713. [Google Scholar] [CrossRef]
- Usov, A.I. Polysaccharides of the red algae. Adv. Carbohydr. Chem. Biochem. 2015, 65, 115–217. [Google Scholar]
- Gómez-Ordóñez, E.; Rupérez, P. FTIR-ATR spectroscopy as a tool for polysaccharide identification in edible brown and red seaweeds. Food Hydrocoll. 2011, 25, 1514–1520. [Google Scholar] [CrossRef]
- Zheng, L.; Zhai, G.; Zhang, J.; Wang, L.; Ma, Z.; Jia, M.; Jia, L. Antihyperlipidemic and hepatoprotective activities of mycelia zinc polysaccharide from Pholiota nameko. Int. J. Biol. Macromol. 2014, 70, 523–529. [Google Scholar] [CrossRef]
- Pereira, L.; Gheda, S.F.; Ribeiro-Claro, P.J.A. Analysis by vibrational spectroscopy of seaweed polysaccharides with potential use in food, pharmaceutical and cosmetic industries. Int. J. Carbohydr. Chem. 2013, 2013, 537202. [Google Scholar] [CrossRef]
- Hardoko, S.T.; Eveline, Y.M.; Olivia, S. An in vitro of antidiabetic activity of Sargassum duplicatum and Turbinaria decurens seaweed. Int. J. Pharm. Sci. Invent. 2014, 3, 13–18. [Google Scholar]
- Yu, Q.; Yan, J.; Wang, S.; Ji, L.; Ding, K.; Vella, C.; Wang, Z.; Hu, Z. Antiangiogenic effects of GFP08, an agaran-type polysaccharide isolated from Grateloupia filicina. Glycobiology 2012, 22, 1343–1352. [Google Scholar] [CrossRef]
- Pereira, L. Identification of phycocolloids by vibrational spectroscopy. In World Seaweed Resources—An Authoritative Reference System; Critchley, A.T., Ohno, M., Largo, D.B., Eds.; Hybrid Windows and Mac DVD-ROM; ETI Information Services Ltd.: Amsterdam, The Netherlands, 2006; ISBN 90-75000-80-4. [Google Scholar]
- Santos, P.S.M.; Santos, E.B.H.; Duarte, A.C. First spectroscopic study on the structural features of dissolved organic matter isolated from rainwater in different seasons. Sci. Total Environ. 2012, 426, 172–179. [Google Scholar] [CrossRef]
- Clarke, C.J.; Haselden, J.N. Metabolic profiling as a tool for understanding mechanisms of toxicity. Toxicol. Pathol. 2008, 36, 140–147. [Google Scholar] [CrossRef]
- Gonzaga, M.L.C.; Ricardo, N.M.P.S.; Heatley, F.; Soares, S.A. Isolation and characterization of polysaccharides from Agaricus blazei Murill. Carbohydr. Polym. 2005, 60, 43–49. [Google Scholar] [CrossRef]
- Bubb, W.A. NMR spectroscopy in the study of carbohydrates; Characterizing the structural complexity. Concepts Magn. Reson. Part A 2003, 19, 1–19. [Google Scholar] [CrossRef]
- Tanniou, A.; Vandanjon, L.; Gonçalves, O.; Kercvarec, N.; Stiger, P.V. Rapid geographical differentiation of the European spread brown macroalga Sargassum muticum using HRMAS NMR and Fourier-Transform Infrared spectroscopy. Talanta 2015, 132, 451–456. [Google Scholar] [CrossRef]
- Robic, A.; Rondeau-Mouro, C.; Sassi, J.-F.; Lerat, Y.; Lahaye, M. Structure and interactions of ulvan in the cell wall of the marine green algae Ulva rotundata (Ulvales, Chlorophyceae). Carbohydr. Polym. 2009, 77, 206–216. [Google Scholar] [CrossRef]
- Barros, F.C.N.; Silva, D.C.; Sombra, V.G.; Maciel, J.S.; Feitosa, J.P.A.; Freitas, A.L.P.; de Paula, R.C. Structural characterization of polysaccharide obtained from red seaweed Gracilaria caudata (J Agardh). Carbohydr. Polym. 2013, 92, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Llanes, F.; Sauriol, F.; Morin, F.G.; Perlin, A.S. An examination of sodium alginate from Sargassum by NMR spectroscopy. Can. J. Chem. 1997, 75, 585–590. [Google Scholar] [CrossRef]
- O’Brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef] [PubMed]
- Krugera, C.L.; Mann, S.W. Safety evaluation of functional ingredients. Food Chem. Toxicol. 2003, 41, 793–805. [Google Scholar] [CrossRef]
- Alves, A.; Sousa, R.A.; Reis, R.L. In Vitro Cytotoxicity Assessment of Ulvan, a Polysaccharide Extracted from Green Algae. Phytother. Res. 2013, 27, 1143–1148. [Google Scholar] [CrossRef]
- Heussner, A.H.; Mazija, L.; Fastner, J.; Dietrich, D.R. Toxin content and cytotoxicity of algal dietary supplements. Toxicol. Appl. Pharmacol. 2012, 265, 263–271. [Google Scholar] [CrossRef]
- Süzgeç-Selçuk, S.; Meriçli, A.H.; Güven, K.C.; Kaiser, M.; Casey, R.; Hingley-Wilson, S.; Lalvani, A.; Tasdemir, D. Evaluation of Turkish Seaweeds for Antiprotozoal, Antimycobacterial and Cytotoxic Activities. Phytother. Res. 2011, 25, 778–783. [Google Scholar] [CrossRef]
- Ayyad, S.N.; Ezmirly, S.T.; Basaif, S.A.; Alarif, W.M.; Badria, A.F.; Badria, F.A. Antioxidant, cytotoxic, antitumor, and protective DNA damage metabolites from the red sea brown alga Sargassum sp. Pharmacogn. Res. 2011, 3, 160–165. [Google Scholar] [CrossRef]
- Khanavi, M.; Gheidarloo, R.; Sadati, N.; Ardekani, M.R.S.; Nabavi, S.M.B.; Tavajohi, S.; Ostad, S.N. Cytotoxicity of fucosterol containing fraction of marine algae against breast and colon carcinoma cell line. Pharmacogn. Mag. 2012, 8, 60–64. [Google Scholar]
- Proestos, C.; Loukatos, P.; Komaitis, M. Determination of biogenic amines in wines by HPLC with precolumn dansylation and fluorimetric detection. Food Chem. 2008, 106, 1218–1224. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| EA_S. muticum Alcalase | EA_O. pinnatifida Viscozyme | |||||
|---|---|---|---|---|---|---|
| (mg/mglyophilized extract) | Ratio 1 | (mg/mglyophilized extract) | Ratio 1 | |||
| Inorganic | Macro elements | K | 407 ± 13 | 7.1 | 174 ± 3 | 6.7 |
| Na | 66 ± 2 | 1.8 | 222 ± 3 | 2.4 | ||
| Ca | 2.31 ± 0.06 | 0.3 | 8.34 ± 0.04 | 1.5 | ||
| Mg | 29.3 ± 0.3 | 2.0 | 36.8 ± 0.8 | 7.7 | ||
| P | 11.6 ± 0.5 | 5.1 | 6.30 ± 0.09 | 3.6 | ||
| Micro elements | Zn | 0.033 ± 0.002 | 1.3 | 0.34 ± 0.02 | 5.9 | |
| B | 0.319 ± 0.004 | 3.0 | 0.28 ± 0.01 | 2.2 | ||
| Mn | 0.045 ± 0.004 | 4.1 | 0.34 ± 0.06 | 29.2 | ||
| Fe | 0.17 ± 0.02 | 0.9 | 1.8 ± 0.1 | 4.9 | ||
| Al | <LOD | - | 0.085 ± 0.07 | 0.6 | ||
| Cu | 0.065 ± 0.002 | 14.4 | 0.026 ± 0.001 | 5.3 | ||
| Ni | <LOD | - | 0.20 ± 0.01 | - | ||
| Pb | 0.040 ± 0.001 | - | 0.020 ± 0.001 | - | ||
| Organic | %N | 2.5 | - | 1.3 | - | |
| %C | 17.1 | - | 16.8 | - | ||
| %H | 2.5 | - | 3.5 | - | ||
| %S | 0.7 | - | 1.7 | - | ||
| S. muticum_Alcalase (mg/glyophyzed extract) | O. pinnatifida_Viscozyme (mg/glyophyzed extract) | ||
|---|---|---|---|
| Monosaccharides | Glucose | <LOD | <LOD |
| Galactose | 19.1 ± 0.3 | 25.3 ± 0.2 | |
| Mannose | 7.8 ± 0.2 | 11.4 ± 0.2 | |
| Arabinose | 0.10 ± 0.01 | 0.16 ± 0.01 | |
| Xylose | 3.23 ± 0.02 | 4.8 ± 0.1 | |
| Rhamnose | 0.27 ± 0.01 | 0.52 ± 0.01 | |
| Fucose | 4.3 ± 0.1 | 5.60 ± 0.09 | |
| Uronic acids | Glucuronic acid | 17.4 ± 0.3 | 27.3 ± 0.2 |
| Galacturonic acid | 1.07 ± 0.01 | 1.50 ± 0.02 | |
| Amino-mon. | Glucosamine | 7.9 ± 0.1 | 12.7 ± 0.4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, D.; Costa-Pinto, A.R.; Sousa, S.; Vasconcelos, M.W.; Pintado, M.M.; Pereira, L.; Rocha-Santos, T.A.P.; Costa, J.P.d.; Silva, A.M.S.; Duarte, A.C.; et al. Sargassum muticum and Osmundea pinnatifida Enzymatic Extracts: Chemical, Structural, and Cytotoxic Characterization. Mar. Drugs 2019, 17, 209. https://doi.org/10.3390/md17040209
Rodrigues D, Costa-Pinto AR, Sousa S, Vasconcelos MW, Pintado MM, Pereira L, Rocha-Santos TAP, Costa JPd, Silva AMS, Duarte AC, et al. Sargassum muticum and Osmundea pinnatifida Enzymatic Extracts: Chemical, Structural, and Cytotoxic Characterization. Marine Drugs. 2019; 17(4):209. https://doi.org/10.3390/md17040209
Chicago/Turabian StyleRodrigues, Dina, Ana R. Costa-Pinto, Sérgio Sousa, Marta W. Vasconcelos, Manuela M. Pintado, Leonel Pereira, Teresa A.P. Rocha-Santos, João P. da Costa, Artur M.S. Silva, Armando C. Duarte, and et al. 2019. "Sargassum muticum and Osmundea pinnatifida Enzymatic Extracts: Chemical, Structural, and Cytotoxic Characterization" Marine Drugs 17, no. 4: 209. https://doi.org/10.3390/md17040209
APA StyleRodrigues, D., Costa-Pinto, A. R., Sousa, S., Vasconcelos, M. W., Pintado, M. M., Pereira, L., Rocha-Santos, T. A. P., Costa, J. P. d., Silva, A. M. S., Duarte, A. C., Gomes, A. M. P., & Freitas, A. C. (2019). Sargassum muticum and Osmundea pinnatifida Enzymatic Extracts: Chemical, Structural, and Cytotoxic Characterization. Marine Drugs, 17(4), 209. https://doi.org/10.3390/md17040209