Microbial Population Changes in Decaying Ascophyllum nodosum Result in Macroalgal-Polysaccharide-Degrading Bacteria with Potential Applicability in Enzyme-Assisted Extraction Technologies

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
2.1. MiSeq Sequencing and Data Processing
2.2. Metagenomic Communities Associated with Intact Ascophyllum nodosum
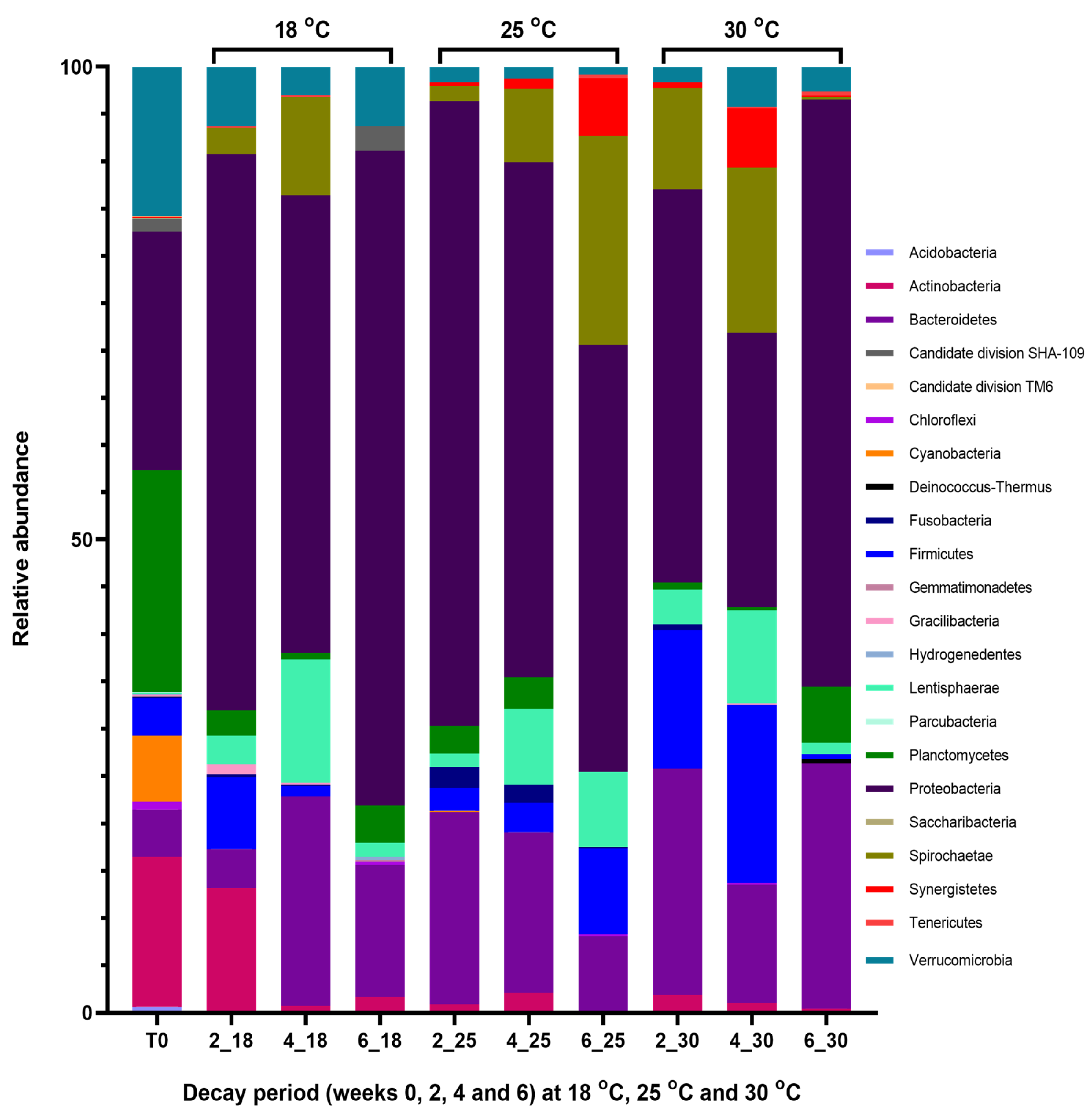
2.3. Metagenomic Communities Associated with Decaying Ascophyllum nodosum
2.3.1. Population Changes in the Seaweed Decaying at 18 °C
2.3.2. Population Changes in the Seaweed Decaying at 25 °C
2.3.3. Population Changes in the Seaweed Decaying at 30 °C
2.4. Cultivable Surface Microbiota Associated with Intact A. nodosum
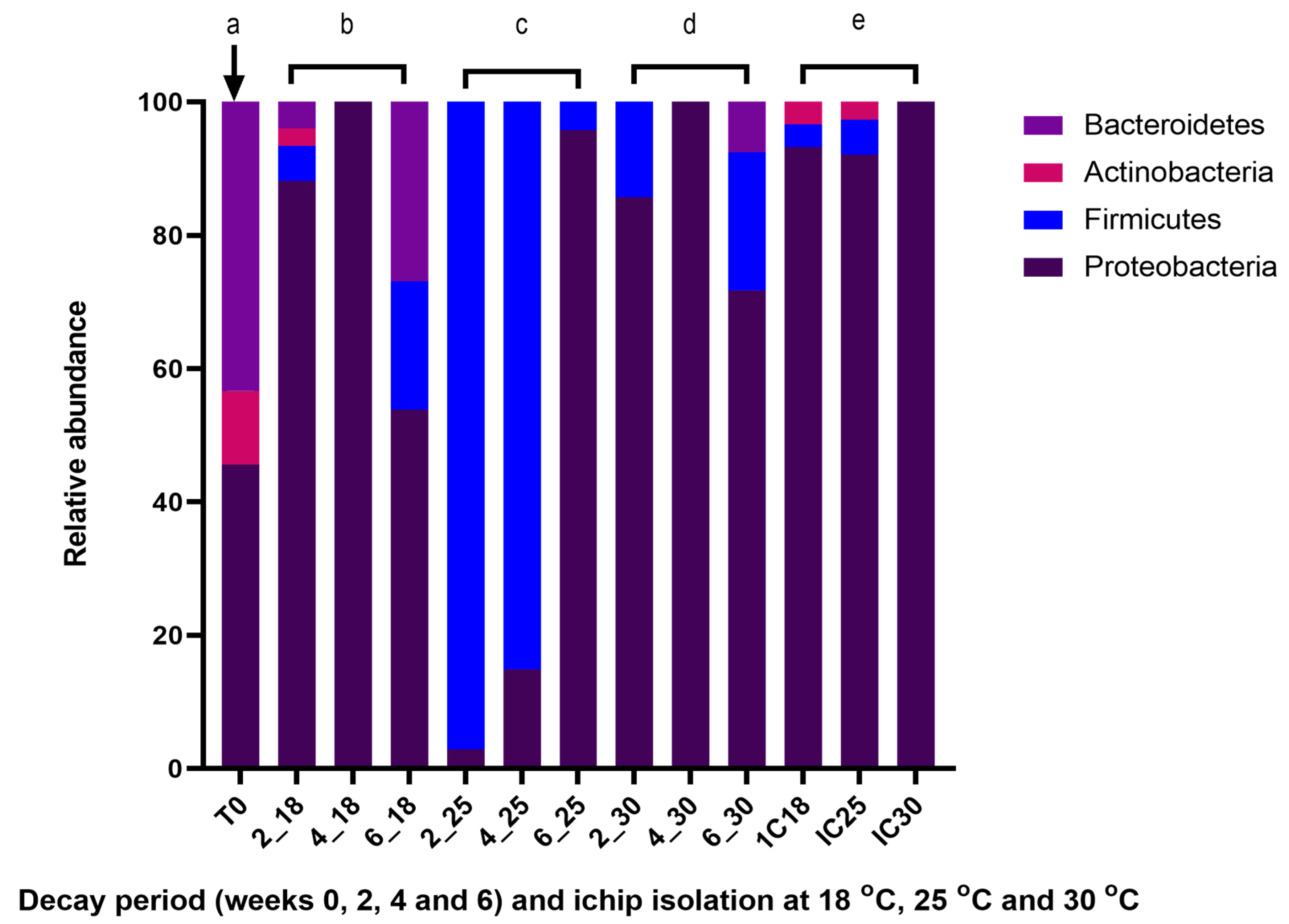
2.5. Cultivable Surface Microbiota Found on Decaying A. nodosum
2.6. ichip Bacterial Isolation Method Applied to Decaying A. nodosum
2.7. Enzymatic Activities of A. nodosum Cultivable Surface Microbiota
2.7.1. Intact Ascophyllum nodosum Isolated Using the Maceration Method
2.7.2. Decaying Ascophyllum nodosum Isolated Using the Maceration Method
2.7.3. Decaying Ascophyllum nodosum Isolated Using the ichip Method
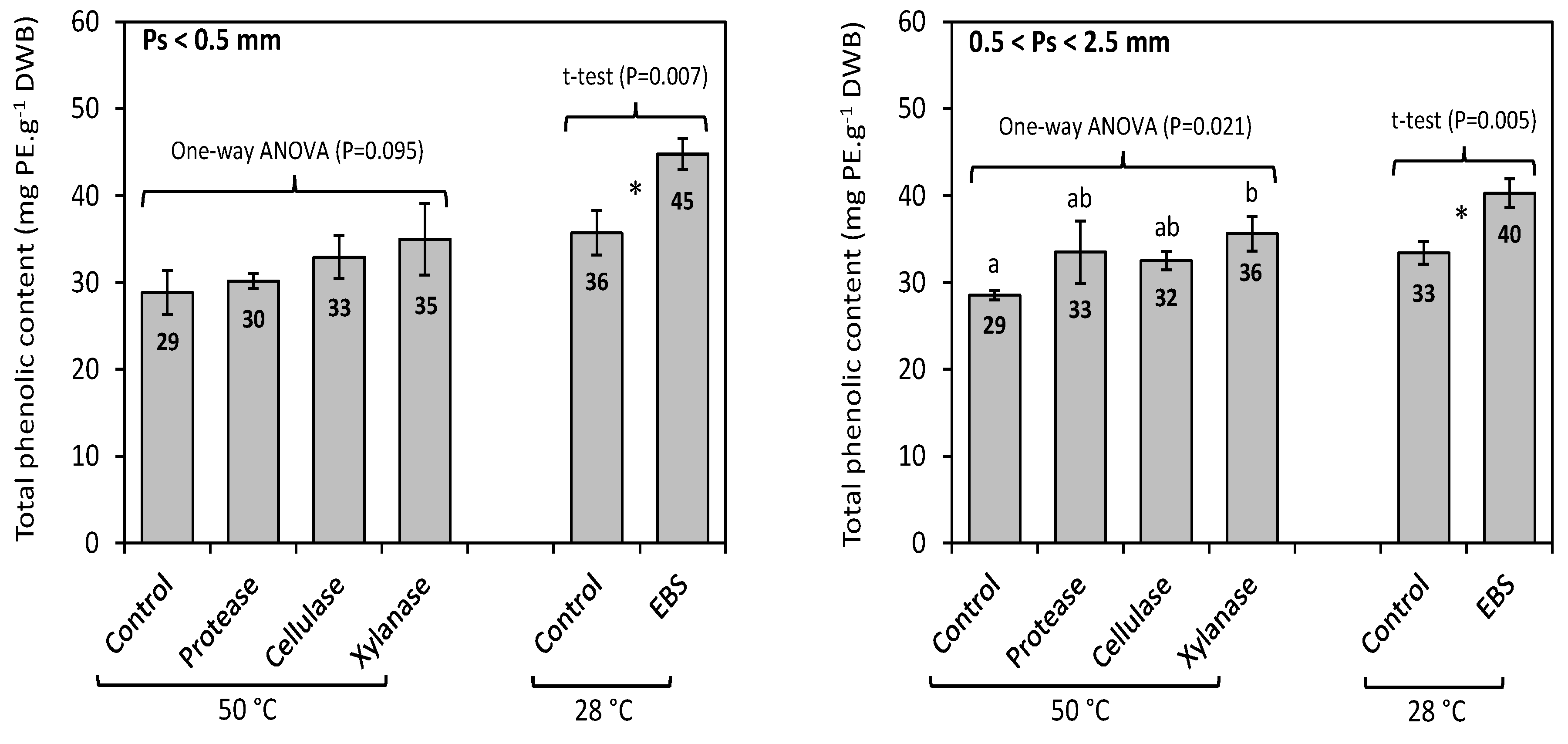
2.8. Enzyme-Assisted Extraction (EAE) of Total Phenolics from F. vesiculosus
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Experimental Design
4.3. 16S rRNA Gene Amplicon Library Preparation and MiSeq Sequencing
4.4. Bacterial Isolation from Intact and Decaying A. nodosum Using Maceration Method
4.5. Bacterial Isolation from Decaying A. nodosum Using ichip Device
4.6. Taxonomic Identification of A. nodosum Cultivable Surface Microbiota Populations
4.7. Enzyme Screens
4.8. Enzyme-Assisted Extraction
4.9. Determination of Total Phenolic Content (TPC)
4.10. Accession Numbers
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Olsen, J.L.; Zechman, F.W.; Hoarau, G.; Coyer, J.A.; Stam, W.T.; Valero, M.; Åberg, P. The phylogeographic architecture of the fucoid seaweed Ascophyllum nodosum: An intertidal ‘marine tree’and survivor of more than one glacial-interglacial cycle. J. Biogeogr. 2010, 37, 842–856. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.; de Morais, A.; de Morais, R. Emergent sources of prebiotics: Seaweeds and microalgae. Mar. Drugs 2016, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Okimura, T.; Yokose, T.; Yamasaki, Y.; Yamaguchi, K.; Oda, T. Effects of sulfated fucan, ascophyllan, from the brown Alga Ascophyllum nodosum on various cell lines: A comparative study on ascophyllan and fucoidan. J. Biosci. Bioeng. 2010, 110, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Nakayasu, S.; Soegima, R.; Yamaguchi, K.; Oda, T. Biological activities of fucose-containing polysaccharide ascophyllan isolated from the brown alga Ascophyllum nodosum. Biosci. Biotechnol. Biochem. 2009, 73, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Suleria, H.A.R.; Osborne, S.; Masci, P.; Gobe, G. Marine-based nutraceuticals: An innovative trend in the food and supplement industries. Mar. Drugs 2015, 13, 6336–6351. [Google Scholar] [CrossRef] [PubMed]
- Adrien, A.; Dufour, D.; Baudouin, S.; Maugard, T.; Bridiau, N. Evaluation of the anticoagulant potential of polysaccharide-rich fractions extracted from macroalgae. Nat. Prod. Res. 2017, 31, 2126–2136. [Google Scholar] [CrossRef] [PubMed]
- Abu, R.; Jiang, Z.; Ueno, M.; Isaka, S.; Nakazono, S.; Okimura, T.; Cho, K.; Yamaguchi, K.; Kim, D.; Oda, T. Anti-metastatic effects of the sulfated polysaccharide ascophyllan isolated from Ascophyllum nodosum on B16 melanoma. Biochem. Biophys. Res. Commun. 2015, 458, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Du, J.-Y.; Jiang, Z.; Okimura, T.; Oda, T.; Yu, Q.; Jin, J.-O. Ascophyllan purified from Ascophyllum nodosum induces Th1 and Tc1 immune responses by promoting dendritic cell maturation. Mar. Drugs 2014, 12, 4148–4164. [Google Scholar] [CrossRef]
- Wang, W.; Wang, S.-X.; Guan, H.-S. The antiviral activities and mechanisms of marine polysaccharides: An overview. Mar. Drugs 2012, 10, 2795–2816. [Google Scholar] [CrossRef]
- Belanche, A.; Ramos-Morales, E.; Newbold, C.J. In vitro screening of natural feed additives from crustaceans, diatoms, seaweeds and plant extracts to manipulate rumen fermentation. J. Sci. Food Agric. 2016, 96, 3069–3078. [Google Scholar] [CrossRef]
- Goñi, O.; Fort, A.; Quille, P.; McKeown, P.C.; Spillane, C.; O’Connell, S. Comparative transcriptome analysis of two Ascophyllum nodosum extract biostimulants: Same seaweed but different. J. Agric. Food Chem. 2016, 64, 2980–2989. [Google Scholar] [CrossRef]
- Santaniello, A.; Scartazza, A.; Gresta, F.; Loreti, E.; Biasone, A.; Di Tommaso, D.; Piaggesi, A.; Perata, P. Ascophyllum nodosum seaweed extract alleviates drought stress in Arabidopsis by affecting photosynthetic performance and related gene expression. Front. Plant Sci. 2017, 8, 1362. [Google Scholar] [CrossRef]
- Bengtsson, M.M.; Sjøtun, K.; Øvreås, L. Seasonal dynamics of bacterial biofilms on the kelp Laminaria hyperborea. Aquat. Microb. Ecol. 2010, 60, 71–83. [Google Scholar] [CrossRef]
- Egan, S.; Harder, T.; Burke, C.; Steinberg, P.; Kjelleberg, S.; Thomas, T. The seaweed holobiont: Understanding seaweed-bacteria interactions. FEMS Microbiol. Rev. 2013, 37, 462–476. [Google Scholar] [CrossRef]
- Singh, R.P.; Shukla, M.K.; Mishra, A.; Reddy, C.; Jha, B. Bacterial extracellular polymeric substances and their effect on settlement of zoospore of Ulva fasciata. Colloids Surf. B Biointerfaces 2013, 103, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, P.D.; De Nys, R.; Kjelleberg, S. Chemical cues for surface colonization. J. Chem. Ecol. 2002, 28, 1935–1951. [Google Scholar] [CrossRef] [PubMed]
- Goecke, F.; Labes, A.; Wiese, J.; Imhoff, J.F. Chemical interactions between marine macroalgae and bacteria. Mar. Ecol. Prog. Ser. 2010, 409, 267–299. [Google Scholar] [CrossRef]
- Singh, R.P.; Reddy, C. Seaweed-microbial interactions: Key functions of seaweed-associated bacteria. FEMS Microbiol. Ecol. 2014, 88, 213–230. [Google Scholar] [CrossRef] [PubMed]
- Lachnit, T.; Fischer, M.; Künzel, S.; Baines, J.F.; Harder, T. Compounds associated with algal surfaces mediate epiphytic colonization of the marine macroalga Fucus vesiculosus. FEMS Microbiol. Ecol. 2013, 84, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Chun, C.L.; Peller, J.R.; Shively, D.; Byappanahalli, M.N.; Whitman, R.L.; Staley, C.; Zhang, Q.; Ishii, S.; Sadowsky, M.J. Virulence and biodegradation potential of dynamic microbial communities associated with decaying Cladophora in Great Lakes. Sci. Total Environ. 2017, 574, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Barbeyron, T.; Martin, R.; Portetelle, D.; Michel, G.; Vandenbol, M. The cultivable surface microbiota of the brown alga Ascophyllum nodosum is enriched in macroalgal-polysaccharide-degrading bacteria. Front. Microbiol. 2015, 6, 1487. [Google Scholar] [CrossRef]
- Martin, M.; Vandermies, M.; Joyeux, C.; Martin, R.; Barbeyron, T.; Michel, G.; Vandenbol, M. Discovering novel enzymes by functional screening of plurigenomic libraries from alga-associated Flavobacteria and Gammaproteobacteria. Microbiol. Res. 2016, 186, 52–61. [Google Scholar] [CrossRef]
- Hardouin, K.; Bedoux, G.; Burlot, A.-S.; Donnay-Moreno, C.; Bergé, J.-P.; Nyvall-Collén, P.; Bourgougnon, N. Enzyme-assisted extraction (EAE) for the production of antiviral and antioxidant extracts from the green seaweed Ulva armoricana (Ulvales, Ulvophyceae). Algal Res. 2016, 16, 233–239. [Google Scholar] [CrossRef]
- Kulshreshtha, G.; Burlot, A.-S.; Marty, C.; Critchley, A.; Hafting, J.; Bedoux, G.; Bourgougnon, N.; Prithiviraj, B. Enzyme-assisted extraction of bioactive material from Chondrus crispus and Codium fragile and its effect on herpes simplex virus (HSV-1). Mar. Drugs 2015, 13, 558–580. [Google Scholar] [CrossRef] [PubMed]
- Leal, M.C.; Munro, M.H.; Blunt, J.W.; Puga, J.; Jesus, B.; Calado, R.; Rosa, R.; Madeira, C. Biogeography and biodiscovery hotspots of macroalgal marine natural products. Nat. Prod. Rep. 2013, 30, 1380–1390. [Google Scholar] [CrossRef] [PubMed]
- Deniaud-Bouët, E.; Kervarec, N.; Michel, G.; Tonon, T.; Kloareg, B.; Hervé, C. Chemical and enzymatic fractionation of cell walls from Fucales: Insights into the structure of the extracellular matrix of brown algae. Ann. Bot. 2014, 114, 1203–1216. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Application of novel extraction technologies for bioactives from marine algae. J. Agric. Food Chem. 2013, 61, 4667–4675. [Google Scholar] [CrossRef] [PubMed]
- Joana Gil-Chávez, G.; Villa, J.A.; Fernando Ayala-Zavala, J.; Basilio Heredia, J.; Sepulveda, D.; Yahia, E.M.; González-Aguilar, G.A. Technologies for extraction and production of bioactive compounds to be used as nutraceuticals and food ingredients: An overview. Compr. Rev. Food Sci. Food Saf. 2013, 12, 5–23. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Wijesinghe, W.P.; Kim, S.K. Enzyme-assisted extraction and recovery of bioactive components from seaweeds. In Handbook of Marine Macroalgae: Biotechnology and Applied Phycology; John Wiley & Sons, Ltd.: Chichester, UK, 2011; pp. 221–228. [Google Scholar]
- Nadar, S.S.; Rao, P.; Rathod, V.K. Enzyme assisted extraction of biomolecules as an approach to novel extraction technology: A review. Food Res. Int. 2018, 108, 309–330. [Google Scholar] [CrossRef] [PubMed]
- Charoensiddhi, S.; Lorbeer, A.J.; Lahnstein, J.; Bulone, V.; Franco, C.M.; Zhang, W. Enzyme-assisted extraction of carbohydrates from the brown alga Ecklonia radiata: Effect of enzyme type, pH and buffer on sugar yield and molecular weight profiles. Process Biochem. 2016, 51, 1503–1510. [Google Scholar] [CrossRef]
- Nichols, D.; Cahoon, N.; Trakhtenberg, E.; Pham, L.; Mehta, A.; Belanger, A.; Kanigan, T.; Lewis, K.; Epstein, S. Use of ichip for high-throughput in situ cultivation of “uncultivable” microbial species. Appl. Environ. Microbiol. 2010, 76, 2445–2450. [Google Scholar] [CrossRef] [PubMed]
- Boyd, K.G.; Adams, D.R.; Burgess, J.G. Antibacterial and repellent activities of marine bacteria associated with algal surfaces. Biofouling 1999, 14, 227–236. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Kenny, O.; Brunton, N.P.; Smyth, T.J. In vitro protocols for measuring the antioxidant capacity of algal extracts. In Natural Products from Marine Algae; Springer, Humana Press: New York, NY, USA, 2015; pp. 375–402. [Google Scholar]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Lee, S.-H.; Jeon, Y.-J. Anti-diabetic effects of brown algae derived phlorotannins, marine polyphenols through diverse mechanisms. Fitoterapia 2013, 86, 129–136. [Google Scholar] [CrossRef]
- Shibata, T.; Ishimaru, K.; Kawaguchi, S.; Yoshikawa, H.; Hama, Y. Antioxidant activities of phlorotannins isolated from Japanese Laminariaceae. In Nineteenth International Seaweed Symposium; Springer: New York, NY, USA, 2007; pp. 255–261. [Google Scholar]
- Nwosu, F.; Morris, J.; Lund, V.A.; Stewart, D.; Ross, H.A.; McDougall, G.J. Anti-proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem. 2011, 126, 1006–1012. [Google Scholar] [CrossRef]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Angell, A.R.; Angell, S.F.; de Nys, R.; Paul, N.A. Seaweed as a protein source for mono-gastric livestock. Trends Food Sci. Technol. 2016, 54, 74–84. [Google Scholar] [CrossRef]
- Martin, M.; Biver, S.; Steels, S.; Barbeyron, T.; Jam, M.; Portetelle, D.; Michel, G.; Vandenbol, M. Identification and characterization of a halotolerant, cold-active marine endo-β-1, 4-glucanase by using functional metagenomics of seaweed-associated microbiota. Appl. Environ. Microbiol. 2014, 80, 4958–4967. [Google Scholar] [CrossRef]
- Dubinina, G.; Grabovich, M.; Leshcheva, N.; Gronow, S.; Gavrish, E.; Akimov, V. Spirochaeta sinaica sp. nov. a halophilic spirochaete isolated from a cyanobacterial mat. Int. J. Syst. Evol. Microbiol. 2015, 65, 3872–3877. [Google Scholar] [CrossRef]
- Jeske, O.; Schüler, M.; Schumann, P.; Schneider, A.; Boedeker, C.; Jogler, M.; Bollschweiler, D.; Rohde, M.; Mayer, C.; Engelhardt, H. Planctomycetes do possess a peptidoglycan cell wall. Nat. Commun. 2015, 6, 7116. [Google Scholar] [CrossRef]
- Lage, O.M.; Bondoso, J. Bringing Planctomycetes into pure culture. Front. Microbiol. 2012, 3, 405. [Google Scholar] [CrossRef]
- Tanaka, Y.; Matsuzawa, H.; Tamaki, H.; Tagawa, M.; Toyama, T.; Kamagata, Y.; Mori, K. Isolation of novel bacteria including rarely cultivated Phyla, Acidobacteria and Verrucomicrobia, from the roots of emergent plants by simple culturing method. Microbes Environ. 2017, 32, 288–292. [Google Scholar] [CrossRef]
- Berdy, B.; Spoering, A.L.; Ling, L.L.; Epstein, S.S. In situ cultivation of previously uncultivable microorganisms using the ichip. Nat. Protoc. 2017, 12, 2232. [Google Scholar] [CrossRef]
- Kealey, C.; Creaven, C.; Murphy, C.; Brady, C. New approaches to antibiotic discovery. Biotechnol. Lett. 2017, 39, 805–817. [Google Scholar] [CrossRef]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schäberle, T.F.; Hughes, D.E.; Epstein, S. A new antibiotic kills pathogens without detectable resistance. Nature 2015, 517, 455. [Google Scholar] [CrossRef] [PubMed]
- Marzinelli, E.M.; Campbell, A.H.; Zozaya Valdes, E.; Vergés, A.; Nielsen, S.; Wernberg, T.; de Bettignies, T.; Bennett, S.; Caporaso, J.G.; Thomas, T. Continental-scale variation in seaweed host-associated bacterial communities is a function of host condition, not geography. Environ. Microbiol. 2015, 17, 4078–4088. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E.; Koren, O.; Reshef, L.; Efrony, R.; Zilber-Rosenberg, I. The role of microorganisms in coral health, disease and evolution. Nat. Rev. Microbiol. 2007, 5, 355. [Google Scholar] [CrossRef] [PubMed]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbial. 2010, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Evans, F.F.; Schleheck, D.; Mai-Prochnow, A.; Burke, C.; Penesyan, A.; Dalisay, D.S.; Stelzer-Braid, S.; Saunders, N.; Johnson, J. Analysis of the Pseudoalteromonas tunicata genome reveals properties of a surface-associated life style in the marine environment. PLoS ONE 2008, 3, e3252. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.-L.; Li, Y.; Chi, Z.; Chi, Z.-M. Purification and characterization of κ-carrageenase from the marine bacterium Pseudoalteromonas porphyrae for hydrolysis of κ-carrageenan. Process Biochem. 2011, 46, 265–271. [Google Scholar] [CrossRef]
- Zhu, Y.; Thomas, F.; Larocque, R.; Li, N.; Duffieux, D.; Cladière, L.; Souchaud, F.; Michel, G.; Mcbride, M.J. Genetic analyses unravel the crucial role of a horizontally acquired alginate lyase for brown algal biomass degradation by Zobellia galactanivorans. Environ. Microbiol. 2017, 19, 2164–2181. [Google Scholar] [CrossRef]
- Varela-Álvarez, E.; Andreakis, N.; Lago-Lestón, A.; Pearson, G.A.; Serrao, E.A.; Procaccini, G.; Duarte, C.M.; Marba, N. Genomic DNA isolation from green and brown algae (caulerpales and fucales) for microsatellite library construction 1. J. Phycol. 2006, 42, 741–745. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Logue, J.B.; Langenheder, S.; Andersson, A.F.; Bertilsson, S.; Drakare, S.; Lanzén, A.; Lindström, E.S. Freshwater bacterioplankton richness in oligotrophic lakes depends on nutrient availability rather than on species–area relationships. ISME J. 2012, 6, 1127. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsson, T.R.; Jakobsson, H.E.; Andersson, A.F.; Björkstén, B.; Engstrand, L.; Jenmalm, M.C. Low diversity of the gut microbiota in infants with atopic eczema. J. Allergy Clin. Immunol. 2012, 129, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571. [Google Scholar] [CrossRef]
- Buffalo, V. Scythe—A Bayesian Adapter Trimmer (version 0.994 BETA). 2014. Unpublished. Available online: https://github.com/vsbuffalo/scythe (accessed on 11 May 2017).
- Joshi, N.; Fass, J. Sickle—A Windowed Adaptive Trimming Tool for FASTQ Files Using Quality. Available online: https://github.com/najoshi/sickle (accessed on 11 May 2017).
- Aronesty, E. ea-utils: Command-Line Tools for Processing Biological Sequencing Data. 2011. Available online: https://expressionanalysis.github.io/ea-utils/ (accessed on 10 October 2017).
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Paulson, J.N.; Stine, O.C.; Bravo, H.C.; Pop, M. Differential abundance analysis for microbial marker-gene surveys. Nat. Methods 2013, 10, 1200. [Google Scholar] [CrossRef] [PubMed]
- Santavy, D.; Willenz, P.; Colwell, R. Phenotypic study of bacteria associated with the caribbean sclerosponge, Ceratoporella nicholsoni. Appl. Environ. Microbiol. 1990, 56, 1750–1762. [Google Scholar]
- Atkinson, M.; Bingman, C. Elemental composition of commercial seasalts. J. Aquaricult. Aquat. Sci. 1997, 8, 39–43. [Google Scholar]
- Li, M.; Gong, J.; Cottrill, M.; Yu, H.; de Lange, C.; Burton, J.; Topp, E. Evaluation of QIAamp® DNA Stool Mini Kit for ecological studies of gut microbiota. J. Microbiol. Methods 2003, 54, 13–20. [Google Scholar] [CrossRef]
- Turner, S.; Pryer, K.M.; Miao, V.P.; Palmer, J.D. Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis 1. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef]
- Lane, D. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Seguritan, V.; Rohwer, F. FastGroup: A program to dereplicate libraries of 16S rDNA sequences. BMC Bioinform. 2001, 2, 9. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Walter, J.; Mangold, M.; Tannock, G.W. Construction, analysis, and β-glucanase screening of a bacterial artificial chromosome library from the large-bowel microbiota of mice. Appl. Environ. Microbiol. 2005, 71, 2347–2354. [Google Scholar] [CrossRef]
- Wolf, M.; Geczi, A.; Simon, O.; Borriss, R. Genes encoding xylan and β-glucan hydrolysing enzymes in Bacillus subtilis: Characterization, mapping and construction of strains deficient in lichenase, cellulase and xylanase. Microbiology 1995, 141, 281–290. [Google Scholar] [CrossRef]
- Soares, M.M.; Silva, R.D.; Gomes, E. Screening of bacterial strains for pectinolytic activity: Characterization of the polygalacturonase produced by Bacillus sp. Rev. Microbiol. 1999, 30, 299–303. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Kenny, O.; Smyth, T.; Hewage, C.; Brunton, N. Antioxidant properties and quantitative UPLC-MS analysis of phenolic compounds from extracts of fenugreek (Trigonella foenum-graecum) seeds and bitter melon (Momordica charantia) fruit. Food Chem. 2013, 141, 4295–4302. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Decay Period | Sample | No. of Reads after Quality Filtering | No. of OTUs (at 97% Sequence Identity) | Chao1 Richness | Shannon Index |
|---|---|---|---|---|---|
| Week 0 intact seaweed | T0 | 178,699 | 1467 | 2293.5 | 10.1 |
| Week 2 early decay phase | 2_18 | 123,551 | 854 | 2072.1 | 9.5 |
| 2_25 | 350,135 | 1476 | 3327.1 | 10.2 | |
| 2_30 | 138,445 | 724 | 1822.7 | 9.2 | |
| Week 4 mid decay phase | 4_18 | 148,904 | 749 | 1901.7 | 9.3 |
| 4_25 | 151,285 | 1130 | 2061.6 | 9.8 | |
| 4_30 | 139,737 | 1633 | 3490.4 | 10.4 | |
| Week 6 late decay phase | 6_18 | 120,679 | 443 | 1148 | 8.6 |
| 6_25 | 138,659 | 1202 | 2884.5 | 10.0 | |
| 6_30 | 165,816 | 1250 | 3062.8 | 10.0 |
| Week 0 | Week 2 | Week 4 | Week 6 | ichip Isolation | Incubating Temperature |
|---|---|---|---|---|---|
| 90 | 76 | 63 | 52 | 59 | 18 °C |
| 35 | 47 | 70 | 76 | 25 °C | |
| 63 | 67 | 53 | 89 | 30 °C |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ihua, M.W.; Guihéneuf, F.; Mohammed, H.; Margassery, L.M.; Jackson, S.A.; Stengel, D.B.; Clarke, D.J.; Dobson, A.D.W. Microbial Population Changes in Decaying Ascophyllum nodosum Result in Macroalgal-Polysaccharide-Degrading Bacteria with Potential Applicability in Enzyme-Assisted Extraction Technologies. Mar. Drugs 2019, 17, 200. https://doi.org/10.3390/md17040200
Ihua MW, Guihéneuf F, Mohammed H, Margassery LM, Jackson SA, Stengel DB, Clarke DJ, Dobson ADW. Microbial Population Changes in Decaying Ascophyllum nodosum Result in Macroalgal-Polysaccharide-Degrading Bacteria with Potential Applicability in Enzyme-Assisted Extraction Technologies. Marine Drugs. 2019; 17(4):200. https://doi.org/10.3390/md17040200
Chicago/Turabian StyleIhua, Maureen W., Freddy Guihéneuf, Halimah Mohammed, Lekha M. Margassery, Stephen A. Jackson, Dagmar B. Stengel, David J. Clarke, and Alan D. W. Dobson. 2019. "Microbial Population Changes in Decaying Ascophyllum nodosum Result in Macroalgal-Polysaccharide-Degrading Bacteria with Potential Applicability in Enzyme-Assisted Extraction Technologies" Marine Drugs 17, no. 4: 200. https://doi.org/10.3390/md17040200
APA StyleIhua, M. W., Guihéneuf, F., Mohammed, H., Margassery, L. M., Jackson, S. A., Stengel, D. B., Clarke, D. J., & Dobson, A. D. W. (2019). Microbial Population Changes in Decaying Ascophyllum nodosum Result in Macroalgal-Polysaccharide-Degrading Bacteria with Potential Applicability in Enzyme-Assisted Extraction Technologies. Marine Drugs, 17(4), 200. https://doi.org/10.3390/md17040200