Antithrombotics from the Sea: Polysaccharides and Beyond



 ,
,  ,
,  and
and 
Abstract
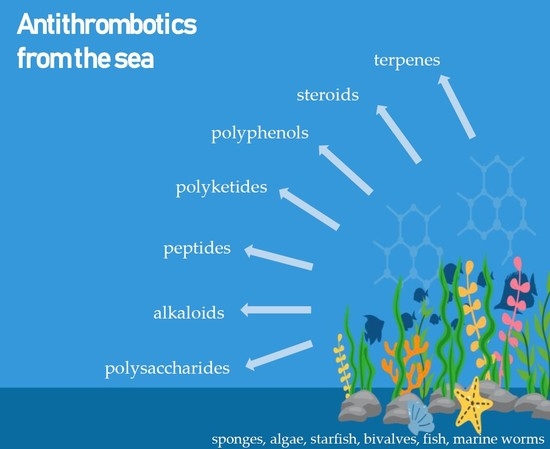
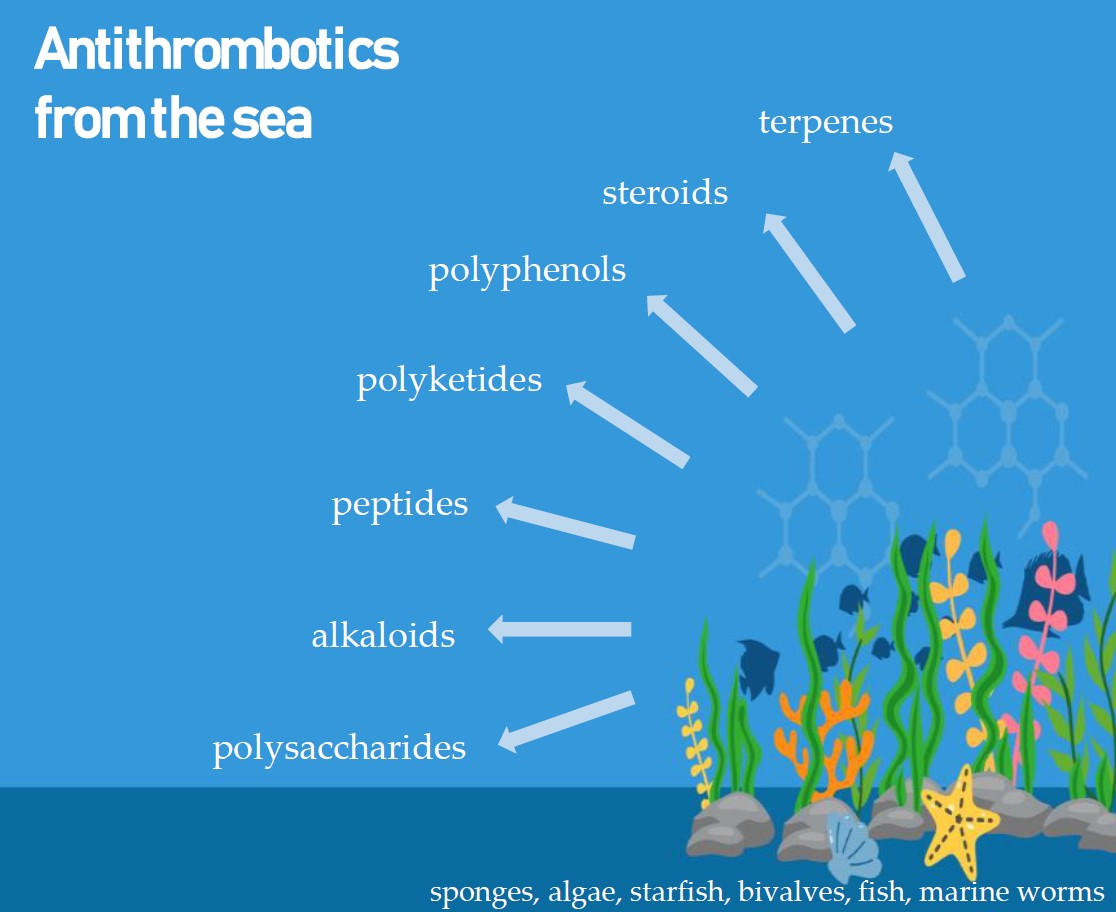
1. Introduction
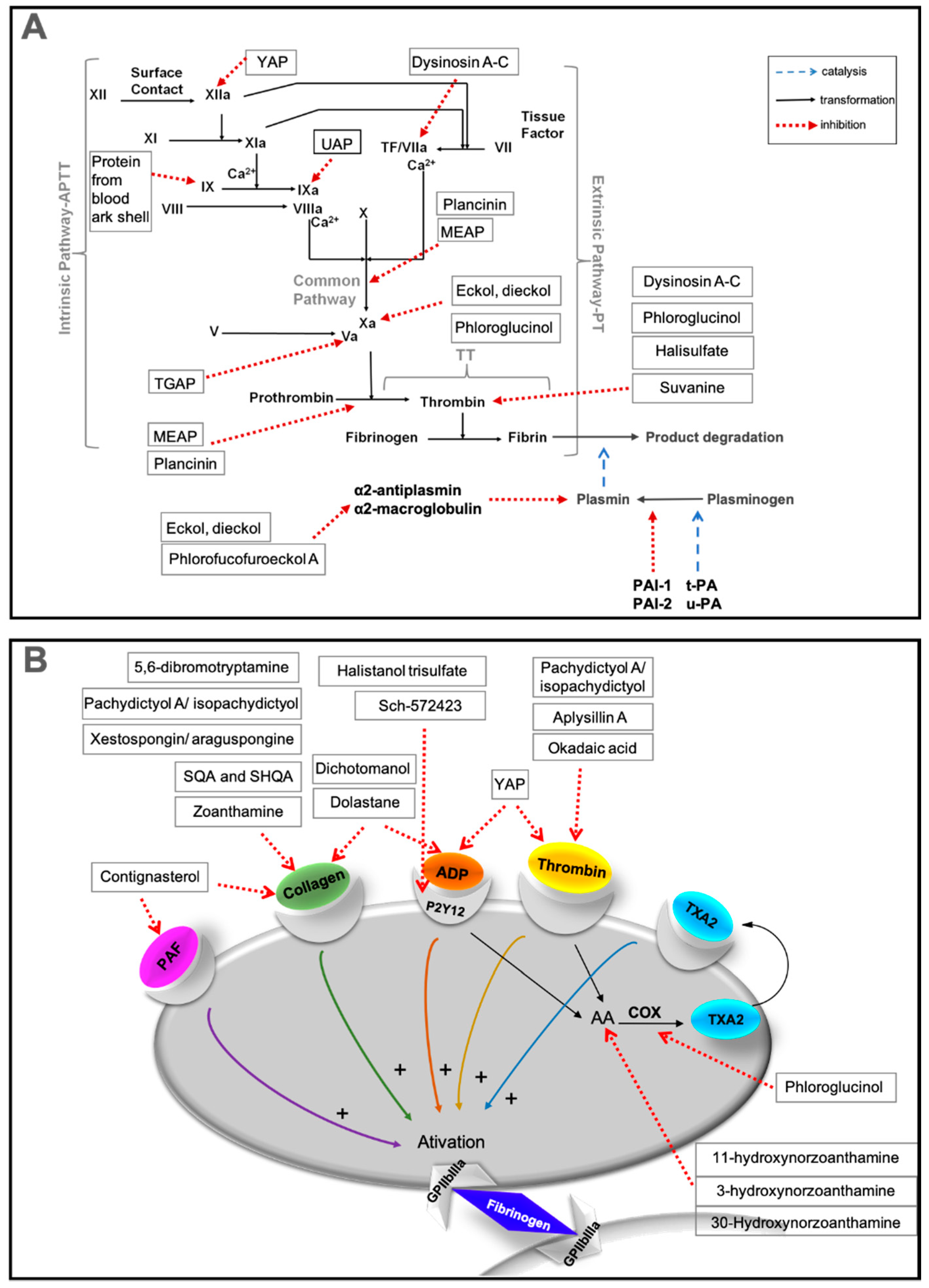
2. Methods Used to Evaluate Antithrombotic Activities of Marine Compounds
2.1. Evaluation of Coagulation
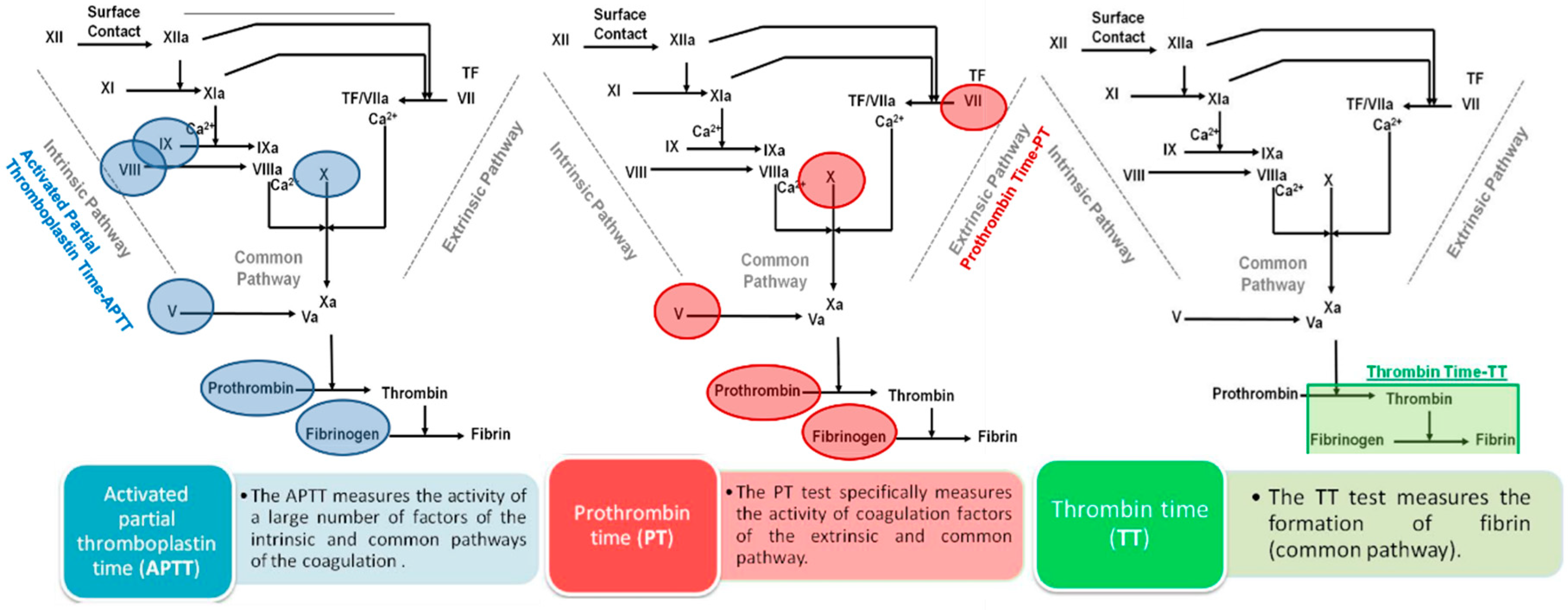
2.1.1. Classical Coagulation Times
2.1.2. Effects on Coagulation Factors
Chromogenic Assays
Modified Clotting Assays
Surface Plasmon Resonance Spectroscopy
PAGE Analysis of Clotting Factor Inhibition
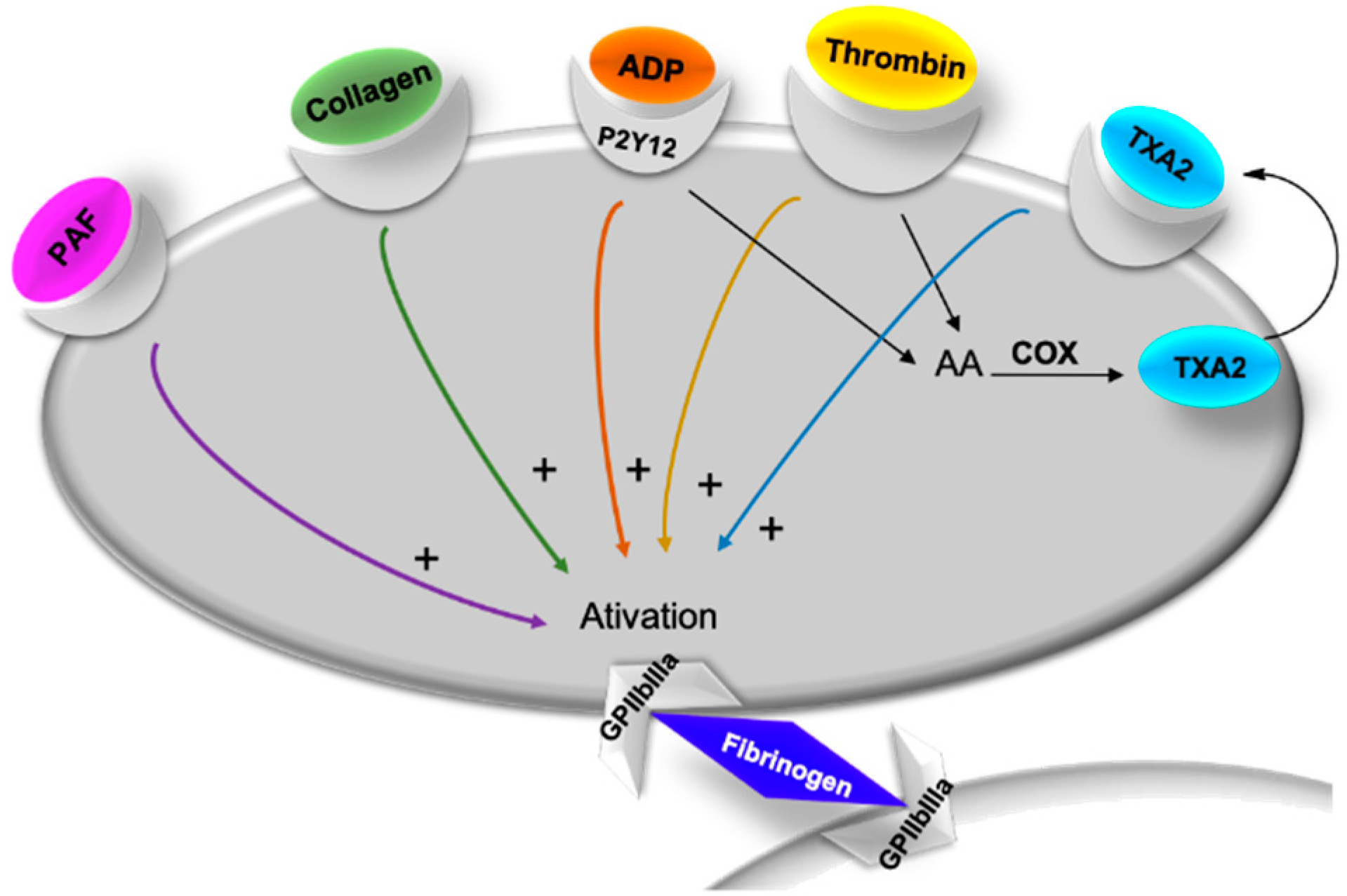
2.2. Evaluation of Platelet Aggregation
2.3. Evaluation of Fibrinolysis
2.4. In Vivo Antithrombotic Activity
2.4.1. Recovery Time from Paralysis
2.4.2. Bleeding Time
2.4.3. Venous Thrombosis
3. Marine Compounds with Antithrombotic Activity
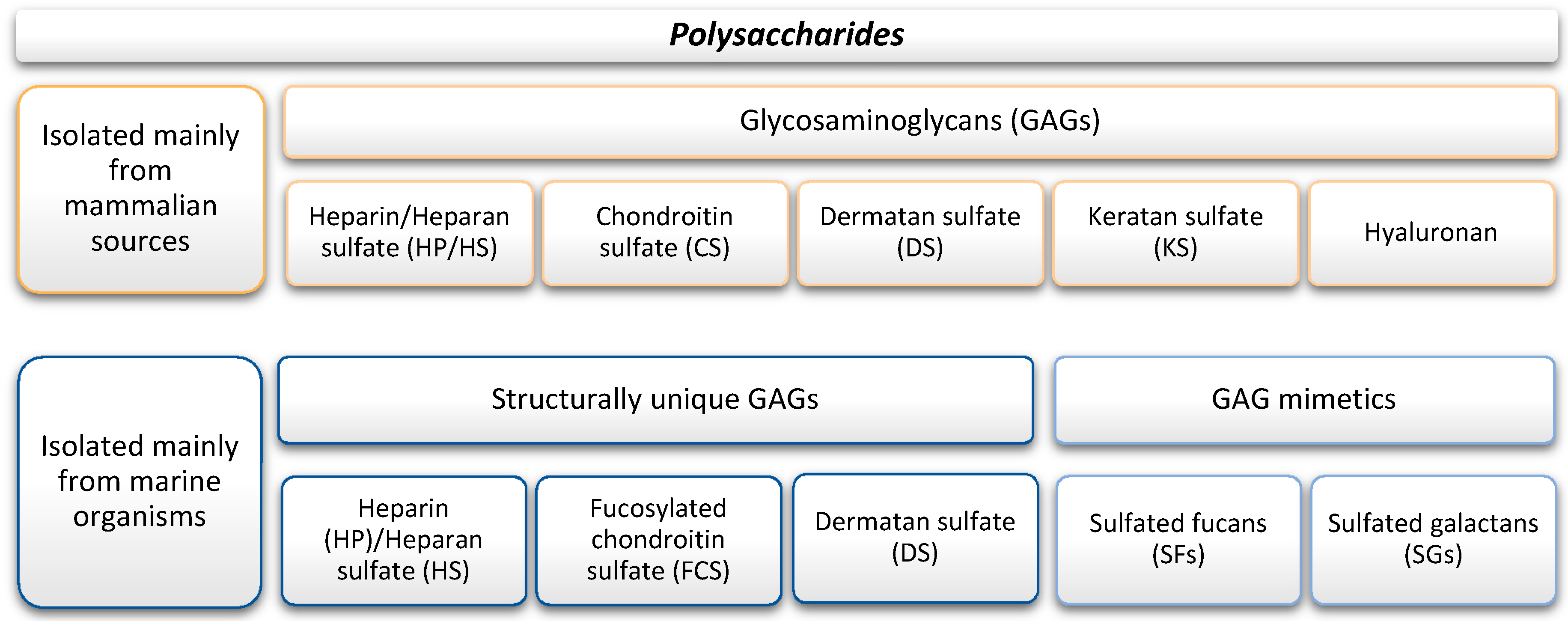
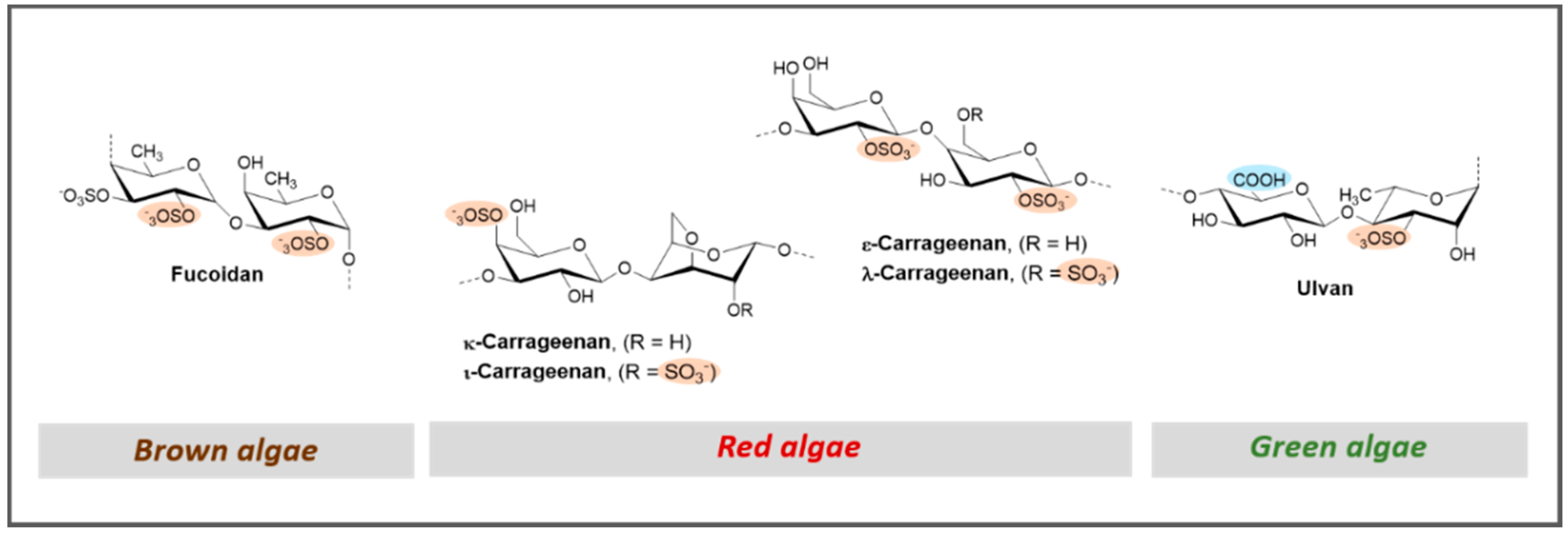
3.1. Polysaccharides
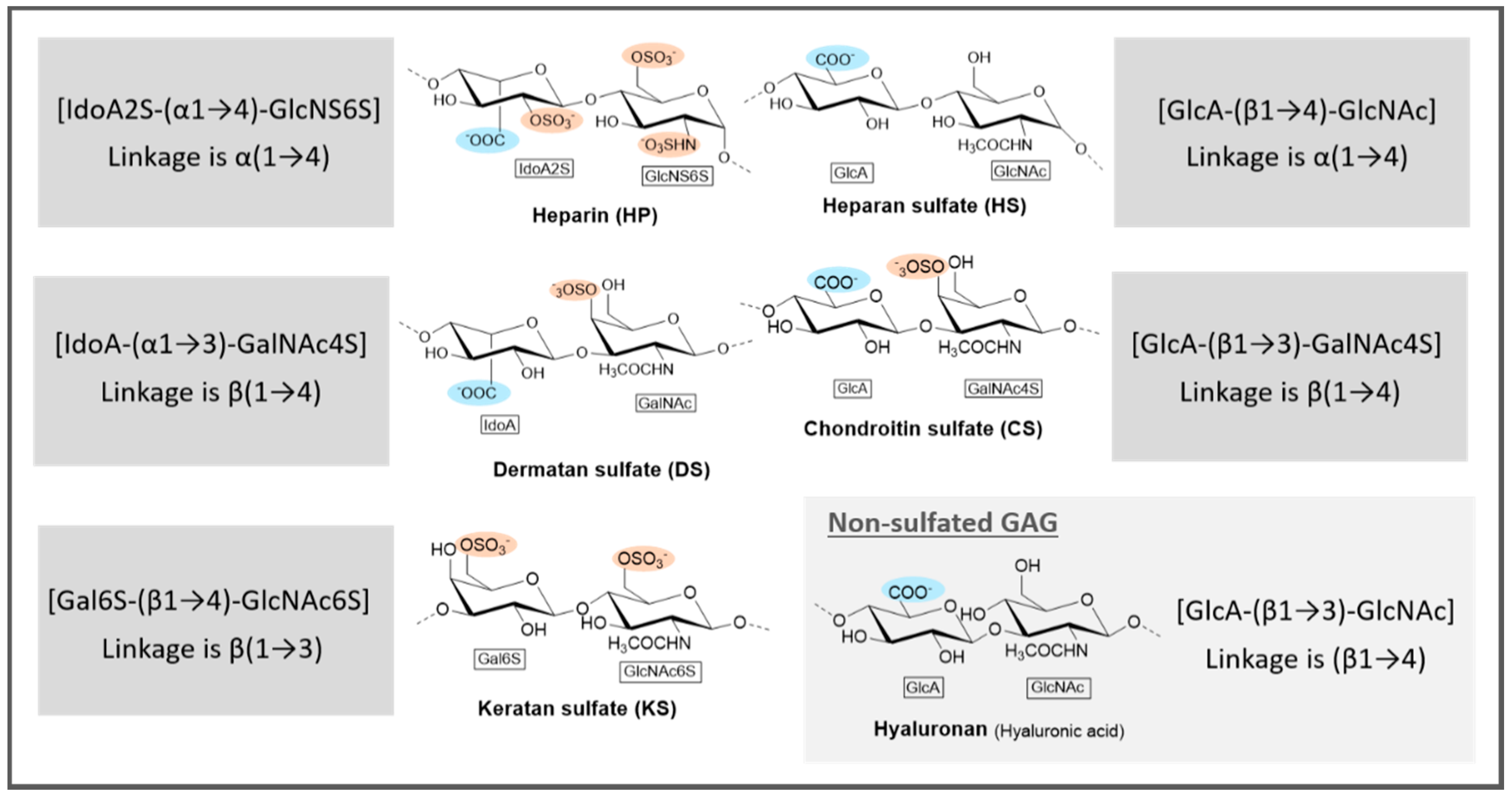
3.1.1. Marine GAGs
3.1.2. Marine GAG Mimetics
3.2. Marine Antithrombotics Other than Polysaccharides
3.2.1. Peptides
3.2.2. Terpenes
3.2.3. Alkaloids
3.2.4. Polyphenols
3.2.5. Steroids
3.2.6. Polyketides
4. Conclusions
5. Future of Marine Antithrombotic Molecules
Author Contributions
Funding
Conflicts of Interest
Abbreviation List
| 2APTT—Two-fold delay of cloth formation | HsG—Fucosylated chondroitin sulfate from Holothuria scabra |
| AA—Arachidonic acid | IC50—Half maximal inhibitory concentration |
| ADP—Adenosine diphosphate | IdoA—Iduronic acid |
| APTT—Activated partial thromboplastin time | IdoA2S—2-Sulfated iduronic acid |
| AT—Antithrombin III | KS—Keratan sulfate |
| CS—Chondroitin sulfate | LMW—Low molecular weight |
| COX—Cyclooxygenase | LMWH—Low molecular weight heparin |
| DS—Dermatan sulfate | L.v.—Sulfated fucan from Lytechinus variegatus |
| E.l.—Sulfated galactan from Echinometra lucunter | MEAP—M. edulis anticoagulant peptide |
| F—Clotting factor | MM—Fucosylated chondroitin sulfate from Massinium magnum |
| Fa—Activated clotting factor | MSP—Sulfated polysaccharide from Monostroma angicava |
| FCS—Fucosylated chondroitin sulfate | PAF—Platelet activating factor |
| FCSc—Fucosylated chondroitin sulfate from Cucumaria frondosa | PAGE—Polyacrylamide gel electrophoresis |
| FCShm—Fucosylated chondroitin sulfate from Holothuria Mexicana | PAI-1 or -2—Plasminogen activator inhibitor 1 or 2 |
| FCSt—Fucosylated chondroitin sulfate from Thelenota ananas | PRP—Platelet-rich plasma |
| Fcup—Fucopyranose | PS—Polysaccharides |
| FHC—Fucogalactan from Hormophysa cuneiformis | PT—Prothrombin time |
| FS—Sulfated fucan from Holothuria albiventer | S.f.—Sulfated fucan from Strongylocentrotus franciscanus |
| FSA—Fucoidan from Sargassum aquifolium | SFs—Sulfated fucans |
| FXase—factor Xase complex | SGs—Sulfated galactans |
| GAGs—Glycosaminoglycans | SHQA—Sargahydroquinoic acid |
| Gal6S—6-Sulfated galactose | SPS-CF—Ulvan from Capsosiphon fulvescens |
| GalNAc4S—4-Sulfated N-acetylgalactosamine | SQA—Sargaquinoic acid |
| GlcA—Glucuronic acid | TGAP—Tegillarca granosa anticoagulant protein |
| GlcN—Glucosamine | TT—Thrombin time |
| GlcNAc—N-Acetylglucosamine | TXA2—Thromboxane A2 |
| GlcNAc6S—6-Sulfated N-acetylglucosamine | UAP—Urechis unicinctus anticoagulant peptide |
| GlcNS6S—N,6-Disulfated glucosamine | u-/ t-PA—Tissue-type plasminogen activator |
| GP—Sulfated polysaccharide from Grateloupia livida | UFH—Unfractionated heparin |
| HP—Heparin | WP—Washed platelets |
| HS—Heparan sulfate | YAP—Yellowfin sole anticoagulant protein |
References
- World Health Organization. The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 22 January 2019).
- World Heath Organization. Cardiovascular Diseases (cvds). Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 22 January 2019).
- Oduah, E.I.; Linhardt, R.J.; Sharfstein, S.T. Heparin: Past, present, and future. Pharmaceuticals 2016, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Ralf, J.L. Therapeutic use of heparin beyond anticoagulation. Curr. Drug Discov. Technol. 2009, 6, 281–289. [Google Scholar] [CrossRef]
- Varki, N.M.; Varki, A. Heparin inhibition of selectin-mediated interactions during the hematogenous phase of carcinoma metastasis: Rationale for clinical studies in humans. Semin. Thromb. Hemost. 2002, 28, 53–66. [Google Scholar] [CrossRef]
- Wang, L.; Brown, J.R.; Varki, A.; Esko, J.D. Heparin’s anti-inflammatory effects require glucosamine 6-o-sulfation and are mediated by blockade of l- and p-selectins. J. Clin. Investig. 2002, 110, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Correia-da-Silva, M.; Sousa, E.; Pinto, M. Strategies to overcome heparins’ low oral bioavailability. Pharmaceuticals 2016, 9, 37. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed marine natural products in the pharmaceutical and cosmeceutical industries: Tips for success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, A.A.; Pomin, V.H. Marine carbohydrate-based compounds with medicinal properties. Mar. Drugs 2018, 16, 233. [Google Scholar] [CrossRef]
- Khotimchenko, Y. Pharmacological potential of sea cucumbers. Int. J. Mol. Sci. 2018, 19, 1342. [Google Scholar] [CrossRef]
- Neves, A.R.; Correia-da-Silva, M.; Sousa, E.; Pinto, M. Structure-activity relationship studies for multitarget antithrombotic drugs. Future Med. Chem. 2016, 8, 2305–2355. [Google Scholar] [CrossRef]
- Cunha, L.; Grenha, A. Sulfated seaweed polysaccharides as multifunctional materials in drug delivery applications. Mar. Drugs 2016, 14, 42. [Google Scholar] [CrossRef]
- Pomin, V.H. Marine non-glycosaminoglycan sulfated glycans as potential pharmaceuticals. Pharmaceuticals 2015, 8, 848–864. [Google Scholar] [CrossRef] [PubMed]
- Mourao, P.A. Perspective on the use of sulfated polysaccharides from marine organisms as a source of new antithrombotic drugs. Mar. Drugs 2015, 13, 2770–2784. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Seo, C.H.; Park, Y. The effects of marine carbohydrates and glycosylated compounds on human health. Int. J. Mol. Sci. 2015, 16, 6018–6056. [Google Scholar] [CrossRef]
- Ustyuzhanina Nadezhda, E.; Ushakova Natalia, A.; Preobrazhenskaya Marina, E.; Bilan Maria, I.; Tsvetkova Eugenia, A.; Krylov Vadim, B.; Anisimova Natalia, A.; Kiselevskiy Mikhail, V.; Krukovskaya Nadezhda, V.; Li, C.; et al. Fucoidans as a platform for new anticoagulant drugs discovery. Pure Appl. Chem. 2014, 86, 1365. [Google Scholar] [CrossRef]
- Pomin, V.H. Anticoagulant motifs of marine sulfated glycans. Glycoconj. J. 2014, 31, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Nasri, R.; Nasri, M. Marine-derived bioactive peptides as new anticoagulant agents: A review. Curr. Protein Pept. Sci. 2013, 14, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H.; Mourão, P.A.d.S. Structure versus anticoagulant and antithrombotic actions of marine sulfated polysaccharides. Rev. Bras. Farmacogn. 2012, 22, 921–928. [Google Scholar] [CrossRef]
- Misurcova, L.; Skrovankova, S.; Samek, D.; Ambrozova, J.; Machu, L. Health benefits of algal polysaccharides in human nutrition. Adv. Food Nutr. Res. 2012, 66, 75–145. [Google Scholar] [CrossRef] [PubMed]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef]
- Delattre, C.; Fenoradosoa, T.A.; Michaud, P. Galactans: An overview of their most important sourcing and applications as natural polysaccharides. Braz. Arch. Biol. Technol. 2011, 54, 1075–1092. [Google Scholar] [CrossRef]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H.; Mourao, P.A. Structure, biology, evolution, and medical importance of sulfated fucans and galactans. Glycobiology 2008, 18, 1016–1027. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.S. Echinoderms: Their culture and bioactive compounds. Prog. Mol. Subcell. Biol. 2005, 39, 139–165. [Google Scholar] [PubMed]
- Vicente, C.P.; He, L.; Pavao, M.S.; Tollefsen, D.M. Antithrombotic activity of dermatan sulfate in heparin cofactor ii-deficient mice. Blood 2004, 104, 3965–3970. [Google Scholar] [CrossRef] [PubMed]
- Mourao, P.A. Use of sulfated fucans as anticoagulant and antithrombotic agents: Future perspectives. Curr. Pharm. Des. 2004, 10, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K. Recent advances in marine algal anticoagulants. Curr. Med. Chem. Cardiovasc. Hematol. Agents 2004, 2, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Berteau, O.; Mulloy, B. Sulfated fucans, fresh perspectives: Structures, functions, and biological properties of sulfated fucans and an overview of enzymes active toward this class of polysaccharide. Glycobiology 2003, 13, 29r–40r. [Google Scholar] [CrossRef]
- Shanmugam, M.; Mody, K.H. Heparinoid-active sulphated polysaccharides from marine algae as potential blood anticoagulant agents. Curr. Sci. 2000, 79, 1672–1683. [Google Scholar]
- Pereira, M.S.; Mulloy, B.; Mourao, P.A. Structure and anticoagulant activity of sulfated fucans. Comparison between the regular, repetitive, and linear fucans from echinoderms with the more heterogeneous and branched polymers from brown algae. J. Biol. Chem. 1999, 274, 7656–7667. [Google Scholar] [CrossRef]
- Kishore, K. In-vitro and in-vivo screening methods for antithrombotic agents. Am. J. Phytomed. Clin. Ther. 2013, 1, 497–506. [Google Scholar]
- Jo, H.-Y.; Jung, W.-K.; Kim, S.-K. Purification and characterization of a novel anticoagulant peptide from marine echiuroid worm, urechis unicinctus. Process Biochem. 2008, 43, 179–184. [Google Scholar] [CrossRef]
- Jung, W.K.; Jo, H.Y.; Qian, Z.J.; Jeong, Y.J.; Park, S.G.; Choi, I.W.; Kim, S.K. A novel anticoagulant protein with high affinity to blood coagulation factor va from tegillarca granosa. J. Biochem. Mol. Biol. 2007, 40, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.-K.; Kim, S.-K. Isolation and characterisation of an anticoagulant oligopeptide from blue mussel, mytilus edulis. Food Chem. 2009, 117, 687–692. [Google Scholar] [CrossRef]
- Kim, T.H.; Ku, S.K.; Bae, J.S. Antithrombotic and profibrinolytic activities of eckol and dieckol. J. Cell. Biochem. 2012, 113, 2877–2883. [Google Scholar] [CrossRef] [PubMed]
- de Andrade Moura, L.; Bianco, E.M.; Pereira, R.C.; Teixeira, V.L.; Fuly, A.L. Anticoagulation and antiplatelet effects of a dolastane diterpene isolated from the marine brown alga canistrocarpus cervicornis. J. Thromb. Thrombolysis 2011, 31, 235–240. [Google Scholar] [CrossRef]
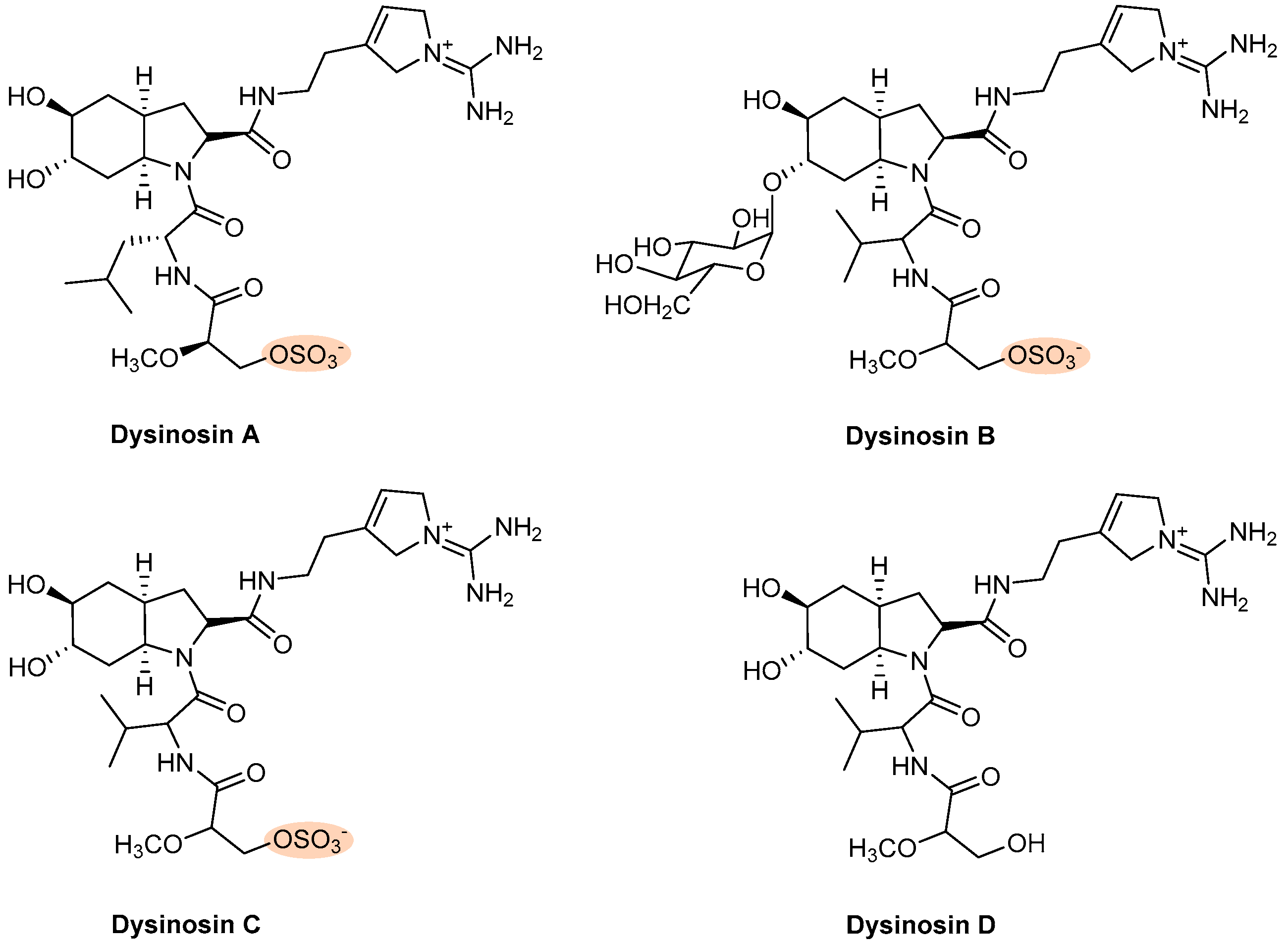
- Carroll, A.R.; Buchanan, M.S.; Edser, A.; Hyde, E.; Simpson, M.; Quinn, R.J. Dysinosins b−d, inhibitors of factor viia and thrombin from the australian sponge lamellodysidea c hlorea. J. Nat. Prod. 2004, 67, 1291–1294. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.-S. Antithrombotic and profibrinolytic activities of phloroglucinol. Food. Chem. Toxicol. 2011, 49, 1572–1577. [Google Scholar] [CrossRef]
- Rajapakse, N.; Jung, W.-K.; Mendis, E.; Moon, S.-H.; Kim, S.-K. A novel anticoagulant purified from fish protein hydrolysate inhibits factor xiia and platelet aggregation. Life Sci. 2005, 76, 2607–2619. [Google Scholar] [CrossRef]
- Jung, W.-K.; Je, J.-Y.; Kim, H.-J.; Kim, S.-K. A novel anticoagulant protein from scapharca broughtonii. J. Biochem. Mol. Biol. 2002, 35, 199–205. [Google Scholar] [CrossRef]
- Yoshiyasu, F.; Iwao, M.; Ziunei, K.; Hideo, M.; Masaru, K.; Yasuo, N.; Masayuki, T.; Masamitsu, O. Eckols, novel phlorotannins with a dibenzo-p-dioxin skeleton possessing inhibitory effects on α2-macroglobulin from the brown alga ecklonia kurome okamura. Chem. Lett. 1985, 14, 739–742. [Google Scholar] [CrossRef]
- Fukuyama, Y.; Kodama, M.; Miura, I.; Kinzyo, Z.; Mori, H.; Nakayama, Y.; Takahashi, M. Anti-plasmin inhibitor. Vi.: Structure of phlorofucofuroeckol a, a novel phlorotannin with both dibenzo-1, 4-dioxin and dibenzofuran elements, from ecklonia kurome okamura. Chem. Pharm. Bull. 1990, 38, 133–135. [Google Scholar] [CrossRef]
- Liu, X.; Du, P.; Liu, X.; Cao, S.; Qin, L.; He, M.; He, X.; Mao, W. Anticoagulant properties of a green algal rhamnan-type sulfated polysaccharide and its low-molecular-weight fragments prepared by mild acid degradation. Mar. Drugs 2018, 16, 445. [Google Scholar] [CrossRef]
- Park, B.-G.; Oh, S.; Kwon, D.; Cui, Y.; Ham, J.; Shin, W.-S.; Lee, S. Anti-platelet aggregation and anti-thrombotic effects of marine natural products sargahydroquinoic acid and sargaquinoic acid. Bull. Korean Chem. Soc. 2013, 34, 3121–3124. [Google Scholar] [CrossRef]
- Koyama, T.; Noguchi, K.; Aniya, Y.; Sakanashi, M. Analysis for sites of anticoagulant action of plancinin, a new anticoagulant peptide isolated from the starfish acanthaster planci, in the blood coagulation cascade. Gen. Pharmacol. 1998, 31, 277–282. [Google Scholar] [CrossRef]
- Vasconcelos, A.A.; Sucupira, I.D.; Guedes, A.L.; Queiroz, I.N.; Frattani, F.S.; Fonseca, R.J.; Pomin, V.H. Anticoagulant and antithrombotic properties of three structurally correlated sea urchin sulfated glycans and their low-molecular-weight derivatives. Mar. Drugs 2018, 16, 304. [Google Scholar] [CrossRef]
- Ernst, B.; Magnani, J.L. From carbohydrate leads to glycomimetic drugs. Nat. Rev. Drug Discov. 2009, 8, 661–677. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, Z.; Linhardt, R.J. Chapter 3—Glycosaminoglycans. In Handbook of Glycomics; Cummings, R.D., Pierce, J.M., Eds.; Academic Press: San Diego, CA, USA, 2010; pp. 59–80. [Google Scholar]
- Vasconcelos, A.A.; Pomin, V.H. The sea as a rich source of structurally unique glycosaminoglycans and mimetics. Microorganisms 2017, 5, 51. [Google Scholar] [CrossRef]
- Pomin, V.; Mourão, P. Specific sulfation and glycosylation—A structural combination for the anticoagulation of marine carbohydrates. Front. Cell. Infect. Microbiol. 2014, 4. [Google Scholar] [CrossRef]
- Saravanan, R.; Shanmugam, A. Isolation and characterization of low molecular weight glycosaminoglycans from marine mollusc amussium pleuronectus (linne) using chromatography. Appl. Biochem. Biotechnol. 2010, 160, 791–799. [Google Scholar] [CrossRef]
- Pavão, M.S.G.; Mourão, P.A.S.; Mulloy, B.; Tollefsen, D.M. A unique dermatan sulfate-like glycosaminoglycan from ascidian: Its structure and the effect of its unusual sulfation pattern on anticoagulant activity. J. Biol. Chem. 1995, 270, 31027–31036. [Google Scholar] [CrossRef]
- Pavao, M.S.; Aiello, K.R.; Werneck, C.C.; Silva, L.C.; Valente, A.P.; Mulloy, B.; Colwell, N.S.; Tollefsen, D.M.; Mourao, P.A. Highly sulfated dermatan sulfates from ascidians. Structure versus anticoagulant activity of these glycosaminoglycans. J. Biol. Chem. 1998, 273, 27848–27857. [Google Scholar] [CrossRef]
- Tapon-Bretaudière, J.; Chabut, D.; Zierer, M.; Matou, S.; Helley, D.; Bros, A.; Mourão, P.A.; Fischer, A.-M. A fucosylated chondroitin sulfate from echinoderm modulates in vitro fibroblast growth factor 2–dependent angiogenesis. Mol. Cancer Res. 2002, 1, 96–102. [Google Scholar]
- Santos, J.C.; Mesquita, J.M.; Belmiro, C.L.; da Silveira, C.B.; Viskov, C.; Mourier, P.A.; Pavão, M.S. Isolation and characterization of a heparin with low antithrombin activity from the body of styela plicata (chordata-tunicata). Distinct effects on venous and arterial models of thrombosis. Thromb. Res. 2007, 121, 213–223. [Google Scholar] [CrossRef]
- Pavão, M.S.G. Structure and anticoagulant properties of sulfated glycosaminoglycans from primitive chordates. An. Acad. Bras. Ciênc. 2002, 74, 105–112. [Google Scholar] [CrossRef]
- Vilela-Silva, A.-C.E.; Alves, A.-P.; Valente, A.-P.; Vacquier, V.D.; Mourão, P.A. Structure of the sulfated α-l-fucan from the egg jelly coat of the sea urchin strongylocentrotus franciscanus: Patterns of preferential 2-o-and 4-o-sulfation determine sperm cell recognition. Glycobiology 1999, 9, 927–933. [Google Scholar] [CrossRef]
- Pomin, V.H. Holothurian fucosylated chondroitin sulfate. Mar. Drugs 2014, 12, 232–254. [Google Scholar] [CrossRef]
- Luo, L.; Wu, M.; Xu, L.; Lian, W.; Xiang, J.; Lu, F.; Gao, N.; Xiao, C.; Wang, S.; Zhao, J. Comparison of physicochemical characteristics and anticoagulant activities of polysaccharides from three sea cucumbers. Mar. Drugs 2013, 11, 399. [Google Scholar] [CrossRef]
- Chen, S.; Xue, C.; Yin, L.a.; Tang, Q.; Yu, G.; Chai, W. Comparison of structures and anticoagulant activities of fucosylated chondroitin sulfates from different sea cucumbers. Carbohydr. Polym. 2011, 83, 688–696. [Google Scholar] [CrossRef]
- Gomes, A.M.; Kozlowski, E.O.; Pomin, V.H.; de Barros, C.M.; Zaganeli, J.L.; Pavão, M.S. Unique extracellular matrix heparan sulfate from the bivalve nodipecten nodosus (linnaeus, 1758) safely inhibits arterial thrombosis after photochemically induced endothelial lesion. J. Biol. Chem. 2010, 285, 7312–7323. [Google Scholar] [CrossRef]
- Brito, A.S.; Cavalcante, R.S.; Palhares, L.C.G.F.; Hughes, A.J.; Andrade, G.P.V.; Yates, E.A.; Nader, H.B.; Lima, M.A.; Chavante, S.F. A non-hemorrhagic hybrid heparin/heparan sulfate with anticoagulant potential. Carbohydr. Polym. 2014, 99, 372–378. [Google Scholar] [CrossRef]
- Chavante, S.F.; Santos, E.A.; Oliveira, F.W.; Guerrini, M.; Torri, G.; Casu, B.; Dietrich, C.P.; Nader, H.B. A novel heparan sulphate with high degree of n-sulphation and high heparin cofactor-ii activity from the brine shrimp artemia franciscana. Int. J. Biol. Macromol. 2000, 27, 49–57. [Google Scholar] [CrossRef]
- Liu, X.; Hao, J.; Shan, X.; Zhang, X.; Zhao, X.; Li, Q.; Wang, X.; Cai, C.; Li, G.; Yu, G. Antithrombotic activities of fucosylated chondroitin sulfates and their depolymerized fragments from two sea cucumbers. Carbohydr. Polym. 2016, 152, 343–350. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Shashkov, A.S.; Kusaykin, M.I.; Stonik, V.A.; Nifantiev, N.E.; Usov, A.I. Structure and biological activity of a fucosylated chondroitin sulfate from the sea cucumber cucumaria japonica. Glycobiology 2016, 26, 449–459. [Google Scholar] [CrossRef]
- Ustyuzhanina, N.E.; Bilan, M.I.; Dmitrenok, A.S.; Borodina, E.Y.; Stonik, V.A.; Nifantiev, N.E.; Usov, A.I. A highly regular fucosylated chondroitin sulfate from the sea cucumber massinium magnum: Structure and effects on coagulation. Carbohydr. Polym. 2017, 167, 20–26. [Google Scholar] [CrossRef]
- Li, Q.; Cai, C.; Chang, Y.; Zhang, F.; Linhardt, R.J.; Xue, C.; Li, G.; Yu, G. A novel structural fucosylated chondroitin sulfate from holothuria mexicana and its effects on growth factors binding and anticoagulation. Carbohydr. Polym. 2018, 181, 1160–1168. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; Yang, S.; Lv, Z. Separation, purification, structures and anticoagulant activities of fucosylated chondroitin sulfates from holothuria scabra. Int. J. Biol. Macromol. 2018, 108, 710–718. [Google Scholar] [CrossRef]
- Fonseca, R.J.; Santos, G.R.; Mourao, P.A. Effects of polysaccharides enriched in 2,4-disulfated fucose units on coagulation, thrombosis and bleeding. Practical and conceptual implications. Thromb. Haemost. 2009, 102, 829–836. [Google Scholar] [CrossRef]
- Arivuselvan, N.; Radhigaand, M.; Anantharaman, P. In vitro antioxidant and anticoagulant activities of sulphated polysaccharides from brown seaweed (turbinaria ornata) (turner) j. Agardh. Asian J. Pharm. Biol. Res. 2011, 232–239. [Google Scholar] [CrossRef]
- Athukorala, Y.; Lee, K.W.; Kim, S.K.; Jeon, Y.J. Anticoagulant activity of marine green and brown algae collected from jeju island in korea. Bioresour. Technol. 2007, 98, 1711–1716. [Google Scholar] [CrossRef]
- Azevedo, T.C.; Bezerra, M.E.; Santos Mda, G.; Souza, L.A.; Marques, C.T.; Benevides, N.M.; Leite, E.L. Heparinoids algal and their anticoagulant, hemorrhagic activities and platelet aggregation. Biomed. Pharmacother. 2009, 63, 477–483. [Google Scholar] [CrossRef]
- Bilan, M.I.; Ustyuzhanina, N.E.; Shashkov, A.S.; Thanh, T.T.T.; Bui, M.L.; Tran, T.T.V.; Bui, V.N.; Nifantiev, N.E.; Usov, A.I. A sulfated galactofucan from the brown alga hormophysa cuneiformis (fucales, sargassaceae). Carbohydr. Res. 2018, 469, 48–54. [Google Scholar] [CrossRef]
- Chevolot, L.; Foucault, A.; Chaubet, F.; Kervarec, N.; Sinquin, C.; Fisher, A.-M.; Boisson-Vidal, C. Further data on the structure of brown seaweed fucans: Relationships with anticoagulant activity. Carbohydr. Res. 1999, 319, 154–165. [Google Scholar] [CrossRef]
- de Andrade Moura, L.; Ortiz-Ramirez, F.; Cavalcanti, D.N.; Ribeiro, S.M.; Muricy, G.; Teixeira, V.L.; Fuly, A.L. Evaluation of marine brown algae and sponges from brazil as anticoagulant and antiplatelet products. Mar. Drugs 2011, 9, 1346–1358. [Google Scholar] [CrossRef]
- Dobashi, K.; Nishino, T.; Fujihara, M.; Nagumo, T. Isolation and preliminary characterization of fucose-containing sulfated polysaccharides with blood-anticoagulant activity from the brown seaweed hizikia fusiforme. Carbohydr. Res. 1989, 194, 315–320. [Google Scholar] [CrossRef]
- Dore, C.M.P.G.; Faustino Alves, M.G.d.C.; Pofírio Will, L.S.E.; Costa, T.G.; Sabry, D.A.; de Souza Rêgo, L.A.R.; Accardo, C.M.; Rocha, H.A.O.; Filgueira, L.G.A.; Leite, E.L. A sulfated polysaccharide, fucans, isolated from brown algae sargassum vulgare with anticoagulant, antithrombotic, antioxidant and anti-inflammatory effects. Carbohydr. Polym. 2013, 91, 467–475. [Google Scholar] [CrossRef]
- Grauffel, V.; Kloareg, B.; Mabeau, S.; Durand, P.; Jozefonvicz, J. New natural polysaccharides with potent antithrombic activity: Fucans from brown algae. Biomaterials 1989, 10, 363–368. [Google Scholar] [CrossRef]
- Karaki, N.; Sebaaly, C.; Chahine, N.; Faour, T.; Zinchenko, A.; Rachid, S.; Kanaan, H. The antioxidant and anticoagulant activities of polysaccharides isolated from the brown algae dictyopteris polypodioides growing on the lebanese coast. J. Appl. Pharm. Sci. 2013, 3, 43–51. [Google Scholar] [CrossRef]
- Kuznetsova, T.; Besednova, N.; Mamaev, A.; Momot, A.; Shevchenko, N.; Zvyagintseva, T. Anticoagulant activity of fucoidan from brown algae fucus evanescens of the okhotsk sea. Bull. Exp. Biol. Med. 2003, 136, 471–473. [Google Scholar] [CrossRef]
- Lee, S.-H.; Jeon, Y.-J. Effects of far infrared radiation drying on antioxidant and anticoagulant activities of ecklonia cava extracts. J. Korean Soc. Appl. Biol. Chem. 2010, 53, 175–183. [Google Scholar] [CrossRef]
- Medeiros, V.P.; Queiroz, K.C.; Cardoso, M.L.; Monteiro, G.R.; Oliveira, F.W.; Chavante, S.F.; Guimaraes, L.A.; Rocha, H.A.; Leite, E.L. Sulfated galactofucan from lobophora variegata: Anticoagulant and anti-inflammatory properties. Biochemistry 2008, 73, 1018–1024. [Google Scholar] [CrossRef]
- Millet, J.; Jouault, S.C.; Mauray, S.; Theveniaux, J.; Sternberg, C.; Boisson Vidal, C.; Fischer, A.M. Antithrombotic and anticoagulant activities of a low molecular weight fucoidan by the subcutaneous route. Thromb. Haemost. 1999, 81, 391–395. [Google Scholar] [CrossRef]
- Nishino, T.; Aizu, Y.; Nagumo, T. Antithrombin activity of a fucan sulfate from the brown seaweed ecklonia kurome. Thromb. Res. 1991, 62, 765–773. [Google Scholar] [CrossRef]
- Rocha, H.A.; Moraes, F.A.; Trindade, E.S.; Franco, C.R.; Torquato, R.J.; Veiga, S.S.; Valente, A.P.; Mourao, P.A.; Leite, E.L.; Nader, H.B.; et al. Structural and hemostatic activities of a sulfated galactofucan from the brown alga spatoglossum schroederi. An ideal antithrombotic agent? J. Biol. Chem. 2005, 280, 41278–41288. [Google Scholar] [CrossRef]
- Rupérez, P.; Ahrazem, O.; Leal, J.A. Potential antioxidant capacity of sulfated polysaccharides from the edible marine brown seaweed fucus vesiculosus. J. Agric. Food. Chem. 2002, 50, 840–845. [Google Scholar] [CrossRef]
- Trento, F.; Cattaneo, F.; Pescador, R.; Porta, R.; Ferro, L. Antithrombin activity of an algal polysaccharide. Thromb. Res. 2001, 102, 457–465. [Google Scholar] [CrossRef]
- Yoon, S.J.; Pyun, Y.R.; Hwang, J.K.; Mourao, P.A. A sulfated fucan from the brown alga laminaria cichorioides has mainly heparin cofactor ii-dependent anticoagulant activity. Carbohydr. Res. 2007, 342, 2326–2330. [Google Scholar] [CrossRef]
- Zoysa, M.D.; Nikapitiya, C.; Jeon, Y.-J.; Jee, Y.; Lee, J. Anticoagulant activity of sulfated polysaccharide isolated from fermented brown seaweed sargassum fulvellum. J. Appl. Phycol. 2008, 20, 67–74. [Google Scholar] [CrossRef]
- Carlucci, M.J.; Pujol, C.A.; Ciancia, M.; Noseda, M.D.; Matulewicz, M.C.; Damonte, E.B.; Cerezo, A.S. Antiherpetic and anticoagulant properties of carrageenans from the red seaweed gigartina skottsbergii and their cyclized derivatives: Correlation between structure and biological activity. Int. J. Biol. Macromol. 1997, 20, 97–105. [Google Scholar] [CrossRef]
- Ekanayake, P.M.; Nikapitiya, C.; De Zoysa, M.; Jeon, Y.J.; Lee, J. Isolation and purification of an anticoagulant from schizymenia dubyi by fermentation. Food Sci. Technol. Int. 2007, 13, 355–359. [Google Scholar] [CrossRef]
- Ekanayake, P.M.; Nikapitiya, C.; De Zoysa, M.; Whang, I.; Kim, S.J.; Lee, J. Novel anticoagulant compound from fermented red alga pachymeniopsis elliptica. Eur. Food Res. Technol. 2008, 227, 897–903. [Google Scholar] [CrossRef]
- Farias, W.R.; Valente, A.P.; Pereira, M.S.; Mourao, P.A. Structure and anticoagulant activity of sulfated galactans. Isolation of a unique sulfated galactan from the red algae botryocladia occidentalis and comparison of its anticoagulant action with that of sulfated galactans from invertebrates. J. Biol. Chem. 2000, 275, 29299–29307. [Google Scholar] [CrossRef]
- Gómez-Ordóñez, E.; Jiménez-Escrig, A.; Rupérez, P. Bioactivity of sulfated polysaccharides from the edible red seaweed mastocarpus stellatus. Bioact. Carbohydr. Dietary Fibre 2014, 3, 29–40. [Google Scholar] [CrossRef]
- Kolender, A.A.; Pujol, C.A.; Damonte, E.B.; Matulewicz, M.C.; Cerezo, A.S. The system of sulfated α-(1 → 3)-linked-mannans from the red seaweed nothogenia fastigiata: Structures, antiherpetic and anticoagulant properties. Carbohydr. Res. 1997, 304, 53–60. [Google Scholar] [CrossRef]
- Usov, A.I. Polysaccharides of the red algae. Adv Carbohydr Chem Biochem. 2011, 65, 115–217. [Google Scholar] [CrossRef]
- Melo, F.R.; Pereira, M.S.; Foguel, D.; Mourao, P.A. Antithrombin-mediated anticoagulant activity of sulfated polysaccharides: Different mechanisms for heparin and sulfated galactans. J. Biol. Chem. 2004, 279, 20824–20835. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.G.; Benevides, N.M.; Melo, M.R.; Valente, A.P.; Melo, F.R.; Mourao, P.A. Structure and anticoagulant activity of a sulfated galactan from the red alga, gelidium crinale. Is there a specific structural requirement for the anticoagulant action? Carbohydr. Res. 2005, 340, 2015–2023. [Google Scholar] [CrossRef]
- Pushpamali, W.A.; Nikapitiya, C.; Zoysa, M.D.; Whang, I.; Kim, S.J.; Lee, J. Isolation and purification of an anticoagulant from fermented red seaweed lomentaria catenata. Carbohydr. Polym. 2008, 73, 274–279. [Google Scholar] [CrossRef]
- Sebaaly, C.; Kassem, S.; Grishina, E.; Kanaan, H.; Sweidan, A.; Chmit, M.S.; Kanaan, H.M. Anticoagulant and antibacterial activities of polysaccharides of red algae corallina collected from lebanese coast. J. Appl. Pharm. Sci. 2014, 4, 30–37. [Google Scholar] [CrossRef]
- Sen, A.K., Sr.; Das, A.; Banerji, N.; Siddhanta, A.; Mody, K.; Ramavat, B.; Chauhan, V.; Vedasiromoni, J.; Ganguly, D. A new sulfated polysaccharide with potent blood anti-coagulant activity from the red seaweed grateloupia indica. Br. J. Pharmacol. Chemother. 1994, 16, 279–280. [Google Scholar] [CrossRef]
- Talarico, L.B.; Duarte, M.E.; Zibetti, R.G.; Noseda, M.D.; Damonte, E.B. An algal-derived dl-galactan hybrid is an efficient preventing agent for in vitro dengue virus infection. Planta Med. 2007, 73, 1464–1468. [Google Scholar] [CrossRef]
- Harada, N.; Maeda, M. Chemical structure of antithrombin-active rhamnan sulfate from monostrom nitidum. Biosci. Biotechnol. Biochem. 1998, 62, 1647–1652. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, Y.; Hayashi, T.; Lee, J.-B.; Srisomporn, P.; Maeda, M.; Ozawa, T.; Sakuragawa, N. Inhibition of thrombin by sulfated polysaccharides isolated from green algae. Biochim. Biophys. Acta 2000, 1543, 86–94. [Google Scholar] [CrossRef]
- Jurd, K.M.; Rogers, D.J.; Blunden, G.; McLellan, D.S. Anticoagulant properties of sulphated polysaccharides and a proteoglycan from codium fragile ssp. Atlanticum. J. Appl. Phycol. 1995, 7, 339. [Google Scholar] [CrossRef]
- Lee, J.-B.; Yamagaki, T.; Maeda, M.; Nakanishi, H. Rhamnan sulfate from cell walls of monostroma latissimum. Phytochemistry 1998, 48, 921–925. [Google Scholar] [CrossRef]
- Li, N.; Mao, W.; Yan, M.; Liu, X.; Xia, Z.; Wang, S.; Xiao, B.; Chen, C.; Zhang, L.; Cao, S. Structural characterization and anticoagulant activity of a sulfated polysaccharide from the green alga codium divaricatum. Carbohydr. Polym. 2015, 121, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Uehara, T.; Harada, N.; Sekiguchi, M.; Hiraoka, A. Heparinoid-active sulphated polysaccharides from monostroma nitidum and their distribution in the chlorophyta. Phytochemistry 1991, 30, 3611–3614. [Google Scholar] [CrossRef]
- Mao, W.; Li, H.; Li, Y.; Zhang, H.; Qi, X.; Sun, H.; Chen, Y.; Guo, S. Chemical characteristic and anticoagulant activity of the sulfated polysaccharide isolated from monostroma latissimum (chlorophyta). Int. J. Biol. Macromol. 2009, 44, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Zang, X.; Li, Y.; Zhang, H. Sulfated polysaccharides from marine green algae ulva conglobata and their anticoagulant activity. J. Appl. Phycol. 2006, 18, 9–14. [Google Scholar] [CrossRef]
- Mao, W.-J.; Fang, F.; Li, H.-Y.; Qi, X.-H.; Sun, H.-H.; Chen, Y.; Guo, S.-D. Heparinoid-active two sulfated polysaccharides isolated from marine green algae monostroma nitidum. Carbohydr. Polym. 2008, 74, 834–839. [Google Scholar] [CrossRef]
- Matsubara, K. An anticoagulant proteogylcan from the marine green alga, codium pungniformis. J. Appl. Phycol. 2000, 9–14. [Google Scholar] [CrossRef]
- Matsubara, K.; Hori, K.; Matsuura, Y.; Miyazawa, K. Purification and characterization of a fibrinolytic enzyme and identification of fibrinogen clotting enzyme in a marine green alga, codium divaricatum. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2000, 125, 137–143. [Google Scholar] [CrossRef]
- Qi, X.; Mao, W.; Gao, Y.; Chen, Y.; Chen, Y.; Zhao, C.; Li, N.; Wang, C.; Yan, M.; Lin, C.; et al. Chemical characteristic of an anticoagulant-active sulfated polysaccharide from enteromorpha clathrata. Carbohydr. Polym. 2012, 90, 1804–1810. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.A.; Vanderlei, E.D.; Bessa, É.F.; Magalhães, F.D.; Paula, R.C.; Lima, V.; Benevides, N.M. Anticoagulant activity of a sulfated polysaccharide isolated from the green seaweed caulerpa cupressoides. Braz. Arch. Biol. Technol. 2011, 54, 691–700. [Google Scholar] [CrossRef]
- Rogers, D.J.; Jurd, K.M.; Blunden, G.; Paoletti, S.; Zanetti, F. Anticoagulant activity of a proteoglycan in extracts ofcodium fragile ssp.Atlanticum. J. Appl. Phycol. 1990, 2, 357–361. [Google Scholar] [CrossRef]
- Siddhanta, A.K.; Shanmugam, M.; Mody, K.H.; Goswami, A.M.; Ramavat, B.K. Sulphated polysaccharides of codium dwarkense boergs. From the west coast of india: Chemical composition and blood anticoagulant activity. Int. J. Biol. Macromol. 1999, 26, 151–154. [Google Scholar] [CrossRef]
- Raposo, M.; de Morais, R.; Bernardo de Morais, A. Bioactivity and applications of sulphated polysaccharides from marine microalgae. Mar. Drugs 2013, 11, 233. [Google Scholar] [CrossRef]
- Raposo, M.F.d.J.; de Morais, A.M.M.B. Microalgae for the prevention of cardiovascular disease and stroke. Life Sciences 2015, 125, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Shen, M.; Song, Q.; Xie, J. Biological activities and pharmaceutical applications of polysaccharide from natural resources: A review. Carbohydr. Polym. 2018, 183, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Yang, W.; Yin, R.; Zhou, L.; Li, Z.; Wu, M.; Zhao, J. An anticoagulant fucan sulfate with hexasaccharide repeating units from the sea cucumber holothuria albiventer. Carbohydr. Res. 2018, 464, 12–18. [Google Scholar] [CrossRef]
- Chen, S.; Hu, Y.; Ye, X.; Li, G.; Yu, G.; Xue, C.; Chai, W. Sequence determination and anticoagulant and antithrombotic activities of a novel sulfated fucan isolated from the sea cucumber isostichopus badionotus. Biochim. Biophys. Acta 2012, 1820, 989–1000. [Google Scholar] [CrossRef]
- Mourao, P.A.; Giumaraes, B.; Mulloy, B.; Thomas, S.; Gray, E. Antithrombotic activity of a fucosylated chondroitin sulphate from echinoderm: Sulphated fucose branches on the polysaccharide account for its antithrombotic action. Br. J. Haematol. 1998, 101, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Mourao, P.A.; Pereira, M.S.; Pavao, M.S.; Mulloy, B.; Tollefsen, D.M.; Mowinckel, M.C.; Abildgaard, U. Structure and anticoagulant activity of a fucosylated chondroitin sulfate from echinoderm. Sulfated fucose branches on the polysaccharide account for its high anticoagulant action. J. Biol. Chem. 1996, 271, 23973–23984. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.S.; Vilela-Silva, A.C.; Valente, A.P.; Mourao, P.A. A 2-sulfated, 3-linked alpha-l-galactan is an anticoagulant polysaccharide. Carbohydr. Res. 2002, 337, 2231–2238. [Google Scholar] [CrossRef]
- Vilela-Silva, A.-C.E.S.; Castro, M.O.; Valente, A.-P.; Biermann, C.H.; Mourão, P.A.S. Sulfated fucans from the egg jellies of the closely related sea urchins strongylocentrotus droebachiensis and strongylocentrotus pallidus ensure species-specific fertilization. J. Biol. Chem. 2002, 277, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Xu, L.; Zhao, L.; Xiao, C.; Gao, N.; Luo, L.; Yang, L.; Li, Z.; Chen, L.; Zhao, J. Structural analysis and anticoagulant activities of the novel sulfated fucan possessing a regular well-defined repeating unit from sea cucumber. Mar. Drugs 2015, 13, 2063–2084. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, J.; Song, S.; Zhou, D.; Qiao, W.; Zhu, C.; Liu, S.; Zhu, B. Anticoagulant activity and structural characterization of polysaccharide from abalone (haliotis discus hannai ino) gonad. Molecules 2016, 21, 697. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. Fucanomics and galactanomics: Marine distribution, medicinal impact, conceptions, and challenges. Mar. Drugs 2012, 10, 793–811. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. Marine medicinal glycomics. Front. Cell. Infect. Microbiol. 2014, 4, 5. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. Fucanomics and galactanomics: Current status in drug discovery, mechanisms of action and role of the well-defined structures. Biochim. Biophys. Acta 2012, 1820, 1971–1979. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, R.J.; Oliveira, S.N.; Melo, F.R.; Pereira, M.G.; Benevides, N.M.; Mourao, P.A. Slight differences in sulfation of algal galactans account for differences in their anticoagulant and venous antithrombotic activities. Thromb. Haemost. 2008, 99, 539–545. [Google Scholar] [CrossRef]
- Pomin, V.H. Review: An overview about the structure-function relationship of marine sulfated homopolysaccharides with regular chemical structures. Biopolymers 2009, 91, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Bilan, M.I.; Ustyuzhanina, N.E.; Shashkov, A.S.; Thanh, T.T.T.; Bui, M.L.; Tran, T.T.V.; Bui, V.N.; Usov, A.I. Sulfated polysaccharides of the vietnamese brown alga sargassum aquifolium (fucales, sargassaceae). Carbohydr. Res. 2017, 449, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Morelli, A.; Puppi, D.; Chiellini, F. Chapter 16—Perspectives on biomedical applications of ulvan. In Seaweed Polysaccharides; Venkatesan, J., Anil, S., Kim, S.-K., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 305–330. [Google Scholar]
- Synytsya, A.; Choi, D.J.; Pohl, R.; Na, Y.S.; Capek, P.; Lattová, E.; Taubner, T.; Choi, J.W.; Lee, C.W.; Park, J.K.; et al. Structural features and anti-coagulant activity of the sulphated polysaccharide sps-cf from a green alga capsosiphon fulvescens. Mar. Biotechnol. 2015, 17, 718–735. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Chen, Y.; Jiang, Z.; Zhong, S.; Chen, W.; Zheng, F.; Shi, G. Purification, partial characterization and bioactivity of sulfated polysaccharides from grateloupia livida. Int. J. Biol. Macromol. 2017, 94, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.S.; Melo, F.R.; Mourão, P.A.S. Is there a correlation between structure and anticoagulant action of sulfated galactans and sulfated fucans? Glycobiology 2002, 12, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Karasudani, I.; Koyama, T.; Nakandakari, S.; Aniya, Y. Purification of anticoagulant factor from the spine venom of the crown-of-thorns starfish, acanthaster planci. Toxicon 1996, 34, 871–879. [Google Scholar] [CrossRef]
- Carroll, A.R.; Pierens, G.K.; Fechner, G.; de Almeida Leone, P.; Ngo, A.; Simpson, M.; Hyde, E.; Hooper, J.N.; Boström, S.-L.; Musil, D. Dysinosin a: A novel inhibitor of factor viia and thrombin from a new genus and species of australian sponge of the family dysideidae. J. Am. Chem. Soc. 2002, 124, 13340–13341. [Google Scholar] [CrossRef]
- Indumathi, P.; Mehta, A. A novel anticoagulant peptide from the nori hydrolysate. J. Funct. Foods 2016, 20, 606–617. [Google Scholar] [CrossRef]
- Francischetti, I.M. Platelet aggregation inhibitors from hematophagous animals. Toxicon 2010, 56, 1130–1144. [Google Scholar] [CrossRef] [PubMed]
- Markwardt, F. State-of-the-art review: Antithrombotic agents from hematophagous animals. Clin. Appl. Thromb. Hemost. 1996, 2, 75–82. [Google Scholar] [CrossRef]
- Kimura, J.; Ishizuka, E.; Nakao, Y.; Yoshida, W.Y.; Scheuer, P.J.; Kelly-Borges, M. Isolation of 1-methylherbipoline salts of halisulfate-1 and of suvanine as serine protease inhibitors from a marine sponge, coscinoderma mathewsi. J. Nat. Prod. 1998, 61, 248–250. [Google Scholar] [CrossRef]
- Park, B.G.; Shin, W.S.; Um, Y.; Cho, S.; Park, G.M.; Yeon, D.S.; Kwon, S.C.; Ham, J.; Choi, B.W.; Lee, S. Selective vasodilatation effect of sargahydroquinoic acid, an active constituent of sargassum micracanthum, on the basilar arteries of rabbits. Bioorg. Med. Chem. Lett. 2008, 18, 2624–2627. [Google Scholar] [CrossRef]
- Kim, S.N.; Choi, H.Y.; Lee, W.; Park, G.M.; Shin, W.S.; Kim, Y.K. Sargaquinoic acid and sargahydroquinoic acid from sargassum yezoense stimulate adipocyte differentiation through pparalpha/gamma activation in 3t3-l1 cells. FEBS Lett. 2008, 582, 3465–3472. [Google Scholar] [CrossRef]
- de Andrade Moura, L.; Marqui de Almeida, A.C.; Domingos, T.F.S.; Ortiz-Ramirez, F.; Cavalcanti, D.N.; Teixeira, V.L.; Fuly, A.L. Antiplatelet and anticoagulant effects of diterpenes isolated from the marine alga, dictyota menstrualis. Mar. Drugs 2014, 12, 2471–2484. [Google Scholar] [CrossRef]
- Huryn, D.M.; Wipf, P. Natural product chemistry and cancer drug discovery. In Cancer Drug Design and Discovery, 2nd ed.; Neidle, S., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 91–120. [Google Scholar]
- Villar, R.M.; Gil-Longo, J.; Daranas, A.H.; Souto, M.L.; Fernandez, J.J.; Peixinho, S.; Barral, M.A.; Santafe, G.; Rodriguez, J.; Jimenez, C. Evaluation of the effects of several zoanthamine-type alkaloids on the aggregation of human platelets. Biorg. Med. Chem. 2003, 11, 2301–2306. [Google Scholar] [CrossRef]
- Gulavita, N.K.; Pomponi, S.A.; Wright, A.E.; Garay, M.; Sills, M.A. Aplysillin a, a thrombin receptor antagonist from the marine sponge aplysina fistularis fulva. J. Nat. Prod. 1995, 58, 954–957. [Google Scholar] [CrossRef]
- Glombitza, K.W.; Hauperich, S.; Keusgen, M. Phlorotannins from the brown algae cystophora torulosa and sargassum spinuligerum. Nat. Toxins 1997, 5, 58–63. [Google Scholar] [CrossRef]
- Chang, M.-C.; Chang, H.-H.; Chan, C.-P.; Chou, H.-Y.; Chang, B.-E.; Yeung, S.-Y.; Wang, T.-M.; Jeng, J.-H. Antiplatelet effect of phloroglucinol is related to inhibition of cyclooxygenase, reactive oxygen species, erk/p38 signaling and thromboxane a2 production. Toxicol. Appl. Pharmacol. 2012, 263, 287–295. [Google Scholar] [CrossRef]
- Gould, W.R.; McClanahan, T.B.; Welch, K.M.; Baxi, S.M.; Saiya-Cork, K.; Chi, L.; Johnson, T.R.; Leadley, R.J. Inhibitors of blood coagulation factors xa and iia synergize to reduce thrombus weight and thrombin generation in vivo and in vitro. J. Thromb. Haemost. 2006, 4, 834–841. [Google Scholar] [CrossRef]
- Nar, H.; Bauer, M.; Schmid, A.; Stassen, J.M.; Wienen, W.; Priepke, H.W.; Kauffmann, I.K.; Ries, U.J.; Hauel, N.H. Structural basis for inhibition promiscuity of dual specific thrombin and factor xa blood coagulation inhibitors. Structure 2001, 9, 29–37. [Google Scholar] [CrossRef]
- Deng, J.Z.; McMasters, D.R.; Rabbat, P.M.; Williams, P.D.; Coburn, C.A.; Yan, Y.; Kuo, L.C.; Lewis, S.D.; Lucas, B.J.; Krueger, J.A.; et al. Development of an oxazolopyridine series of dual thrombin/factor xa inhibitors via structure-guided lead optimization. Bioorg. Med. Chem. Lett. 2005, 15, 4411–4416. [Google Scholar] [CrossRef] [PubMed]
- Andersen, R.J.; Allen, T.M.; Burgoyne, D.L. Contignasterol, and Related 3-alpha hydroxy-6-alpha hydroxy-7-beta hydroxy-15-keto-14-beta Steroids Useful as Anti-Inflammatory and Anti-Thrombosis Agents. U.S. Patent 5,506,221, 1996. [Google Scholar]
- Stead, P.; Hiscox, S.; Robinson, P.S.; Pike, N.B.; Sidebottom, P.J.; Roberts, A.D.; Taylor, N.L.; Wright, A.E.; Pomponi, S.A.; Langley, D. Eryloside f, a novel penasterol disaccharide possessing potent thrombin receptor antagonist activity. Bioorg. Med. Chem. Lett. 2000, 10, 661–664. [Google Scholar] [CrossRef]
- Yang, S.-W.; Buivich, A.; Chan, T.-M.; Smith, M.; Lachowicz, J.; Pomponi, S.A.; Wright, A.E.; Mierzwa, R.; Patel, M.; Gullo, V. A new sterol sulfate, sch 572423, from a marine sponge, topsentia sp. Bioorg. Med. Chem. Lett. 2003, 13, 1791–1794. [Google Scholar] [CrossRef]
- Damman, P.; Woudstra, P.; Kuijt, W.J.; de Winter, R.J.; James, S.K. P2y12 platelet inhibition in clinical practice. J. Thromb. Thrombolysis 2012, 33, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, K.; Scheuer, P.J.; Tsukitani, Y.; Kikuchi, H.; Van Engen, D.; Clardy, J.; Gopichand, Y.; Schmitz, F.J. Okadaic acid, a cytotoxic polyether from two marine sponges of the genus halichondria. J. Am. Chem. Soc. 1981, 103, 2469–2471. [Google Scholar] [CrossRef]
- Karaki, H.; Mitsui, M.; Nagase, H.; Ozaki, H.; Shibata, S.; Uemura, D. Inhibitory effect of a toxin okadaic acid, isolated from the black sponge on smooth muscle and platelets. Br. J. Pharmacol. 1989, 98, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Holmes, C.F.; Tsukitani, Y. Okadaic acid: A new probe for the study of cellular regulation. Trends Biochem. Sci. 1990, 15, 98–102. [Google Scholar] [CrossRef]
- Pimentel, S.M.; Bojo, Z.P.; Roberto, A.V.; Lazaro, J.E.; Mangalindan, G.C.; Florentino, L.M.; Lim-Navarro, P.; Tasdemir, D.; Ireland, C.M.; Concepcion, G.P. Platelet aggregation inhibitors from philippine marine invertebrate samples screened in a new microplate assay. Mar. Biotechnol. 2003, 5, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.-K.; Athukorala, Y.; Lee, Y.-J.; Cha, S.H.; Lee, C.-H.; Vasanthan, T.; Choi, K.-S.; Yoo, S.-H.; Kim, S.-K.; Jeon, Y.-J. Sulfated polysaccharide purified from ecklonia cava accelerates antithrombin iii-mediated plasma proteinase inhibition. J. Appl. Phycol. 2007, 19, 425–430. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PS Type | Molecular Type | Name(s) | Source | Structure | Antithrombotic Assays | Ref. | |
|---|---|---|---|---|---|---|---|
| Anticoagulant | Antiplatelet | ||||||
| Marine GAGs | Fucosylated chondroitin sulfates | FCSc FCSt | Sea cucumber (Cucumaria frondosa and Thelenota ananas) | NR | ↑ APTT Θ Thrombin and anti-FXa (in the presence of HP cofactor II) ↑ Bleeding time (in vivo) | NT | [65] |
| FCS | Sea cucumber (Cucumaria japonica) | NR | ↑ APTT Θ Thrombin (in the presence of ATIII) | Θ Collagen and ristocetin-induced platelets aggregation | [66] | ||
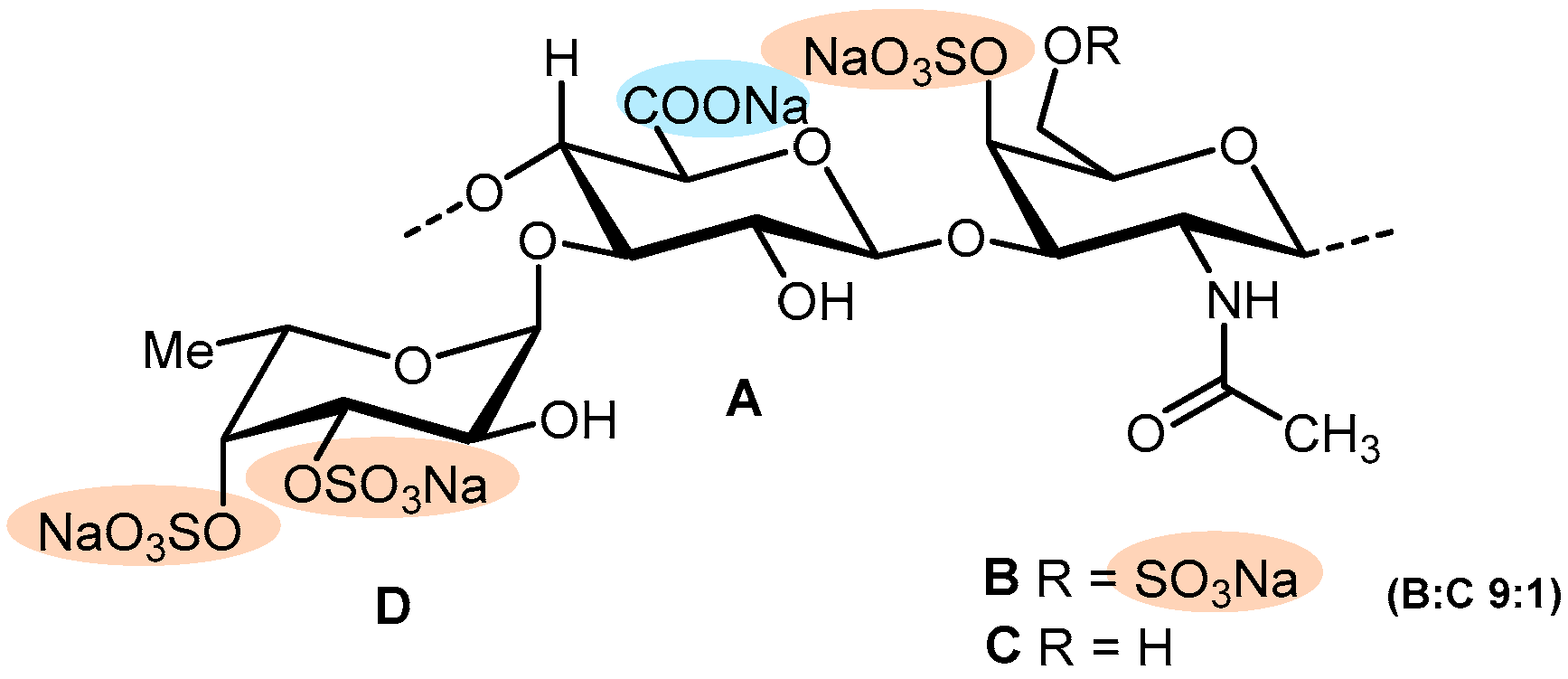
| MM | Sea cucumber (Massinium magnum) | Figure 6 | ↑ APTT, and TT Θ Thrombin (in the presence of ATIII) | NA—Platelets aggregation in PRP | [67] | ||
| FCShm | Sea cucumber (Holothuria Mexicana) | NR | ↑ APTT, and TT | NT | [68] | ||
| HsG | Sea cucumber (Holothuria scabra) | NR | ↑ APTT, and TT Θ Thrombin and FXa (in the presence of ATIII) | NT | [69] | ||
| Marine GAG mimetics | Sulfated fucans | FSA (fucoidan) | Brown algae (Sargassum aquifolium) | NR | ↑ APTT | NT | [135] |
| FHC (fucogalactan) | Brown algae (Hormophysa cuneiformis) | NR | ↑ APTT | NT | [74] | ||
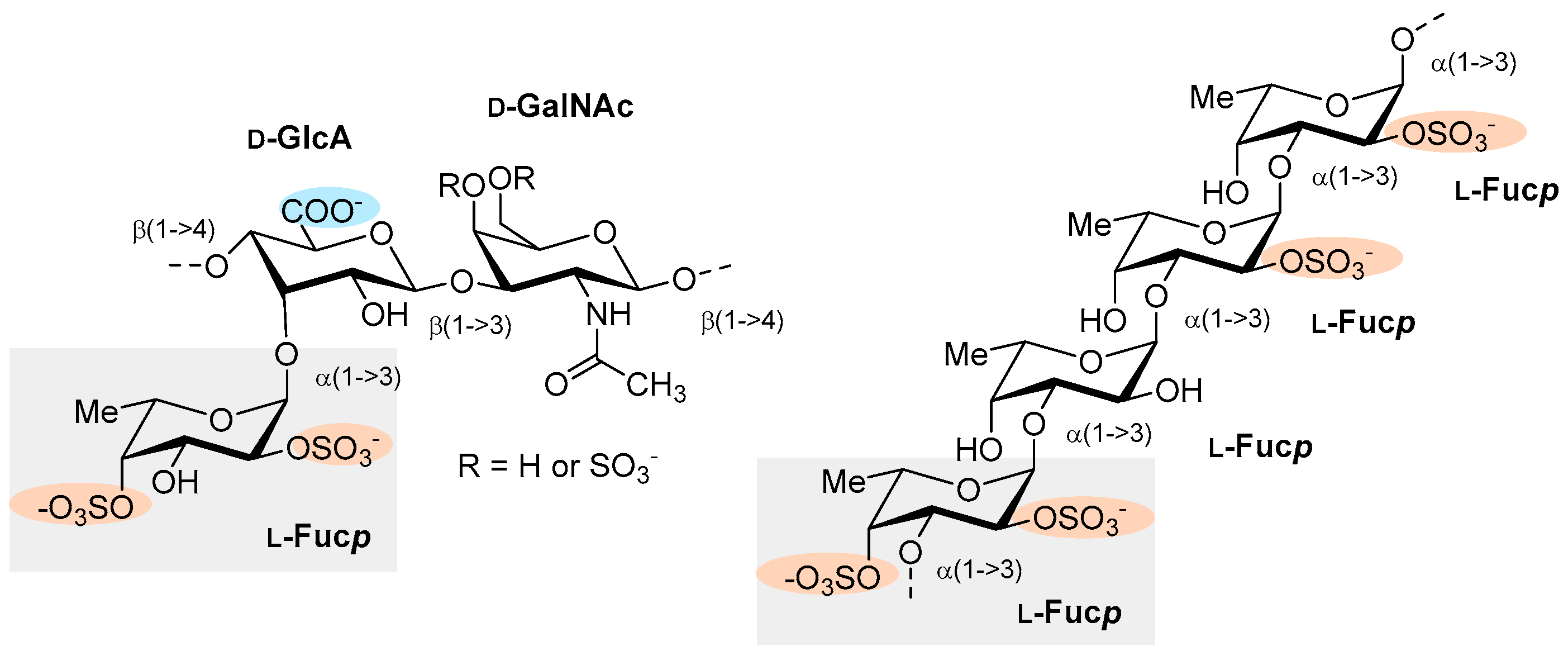
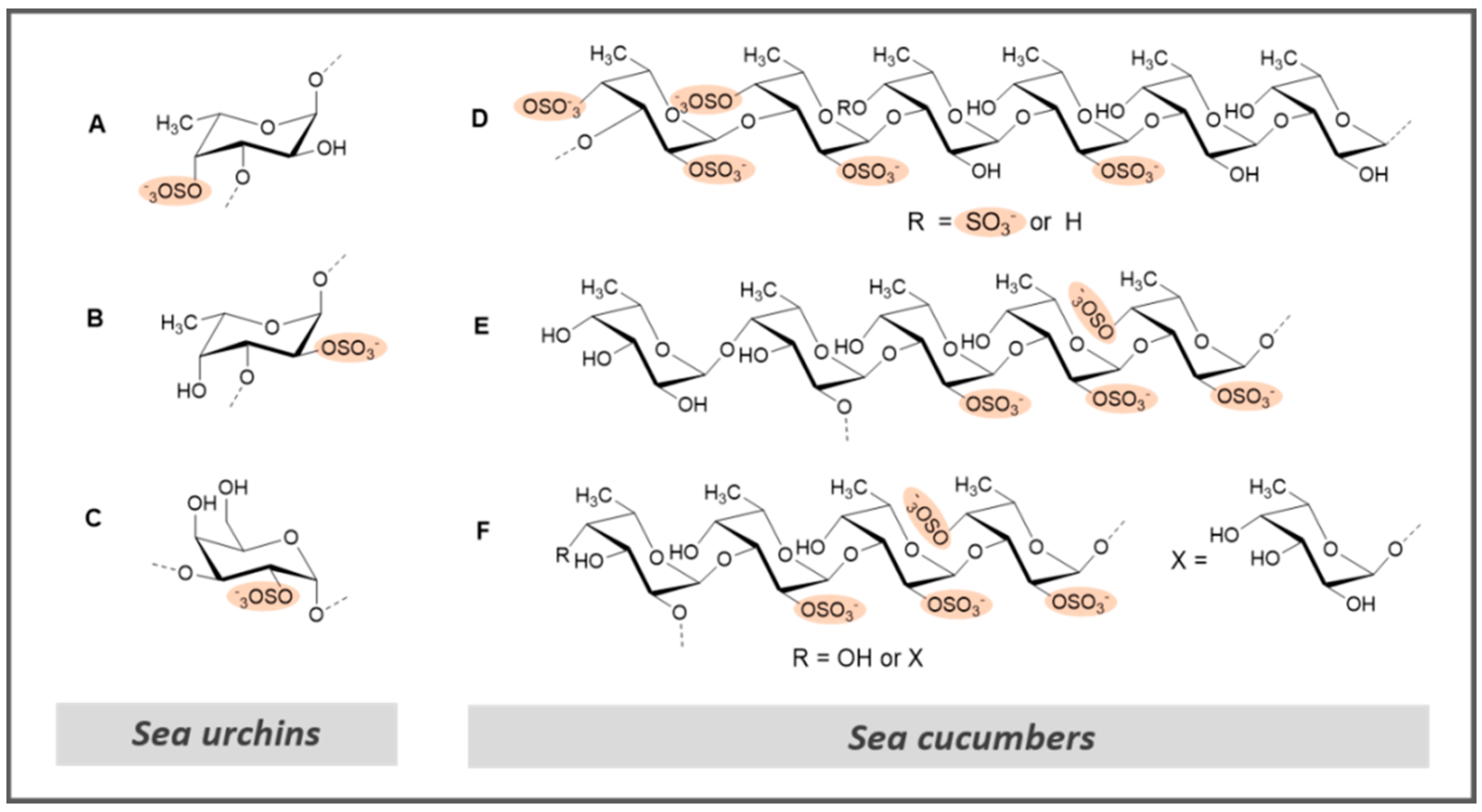
| L.v. | Sea urchin (Lytechinus variegatus) | Figure 8A | ↑ APTT ↑ FXa and thrombin (in the presence of ATIII) Θ Thrombus formation Θ Thrombus formation (in vivo) | NA | [47] | ||
| S.f. | Sea urchin (Strongylocentrotus franciscanus) | Figure 8B | ↑ APTT Θ Thrombus formation Θ Thrombus formation (in vivo) | NA | [47] | ||
| FS | Sea cucumber (Holothuria albiventer) | Figure 8D | ↑ APTT and TT Θ FXase inhibition | NT | [122] | ||
| H. edulis | Sea cucumber (Holothuria edulis) | Figure 8E | ↑ APTT Θ Thrombin (in the presence of HP cofactor II) | NT | [128] | ||
| L. grisea | Sea cucumber (Ludwigothurea grisea) | Figure 8F | ↑ APTT Θ Thrombin (in the presence of HP cofactor II) | NT | [128] | ||
| Sulfated galactans | Carageenan | Red algae (Corallina) | NR | ↑ APTT | NT | [101] | |
| E.l. | Sea urchin (Echinometra lucunter) | Figure 8C | ↑ APTT ↑ anti FXa and thrombin (in the presence of ATIII) Θ thrombus formation Θ Thrombus formation (in vivo) | Θ platelet aggregation | [47] | ||
| Other sulfated PS | SPS-CF (ulvan) | Green algae (Capsosiphon fulvescens) | NR | ↑ APTT and TT | NT | [137] | |
| MSP | Green algae (Monostroma angicava) | NR | ↑ APTT and TT ↑ APTT and TT (in vivo) ↑ Plasmin degradation products (in vivo) ↓ Level of PAI-1 (in vivo) | NT | [44] | ||
| GP | Microalgae (Grateloupia livida) | NR | ↑ APTT and TT | NT | [138] | ||
| Chemical Class | Name | Source | Structure | Antithrombotic Assays | Ref. | |
|---|---|---|---|---|---|---|
| Anticoagulant | Antiplatelet | |||||
| Peptides | Peptide from seaweed Porphyra yezoensis | Seaweed (Porphyra yezoensis) | NR | ↑ APTT | NT | [142] |
| Plancinin | Starfish (Acanthaster planci) | NR | ↑ Fibrin test ↑ APTT and PT Θ Prothrombinase complex ↑ Bleeding time (in vivo) | NA | [140] | |
| Dysinosin A–D | Sponge (Dysideidae) | Figure 10 | Θ FVIIa, thrombin | NT | [38,141] | |
| UAP | Echiuroid worm (U. unicinctus) | NR | ↑ APTT Binds to FIXa | NT | [33] | |
| MEAP | Blue mussel (Mytilus edulis) | NR | ↑ APTT and TT Θ Amidolytic activation of FX ↑ Catalytic conversion of prothrombin to thrombin | NT | [35] | |
| Protein from blood ark shell | Blood ark shell (Scapharca broughtonii) | NR | ↑ APTT Θ FIX | NT | [41] | |
| YAP | Yellowfin sole (Limanda aspera) | NR | ↑ APTT Θ FXIIa | Θ Thrombin- or ADP-induced aggregation | [40] | |
| TGAP | Granulated ark (T. granosa) | NR | ↑ TT Θ FVa Θ Interaction between prothrombin and FVa | NT | [165] | |
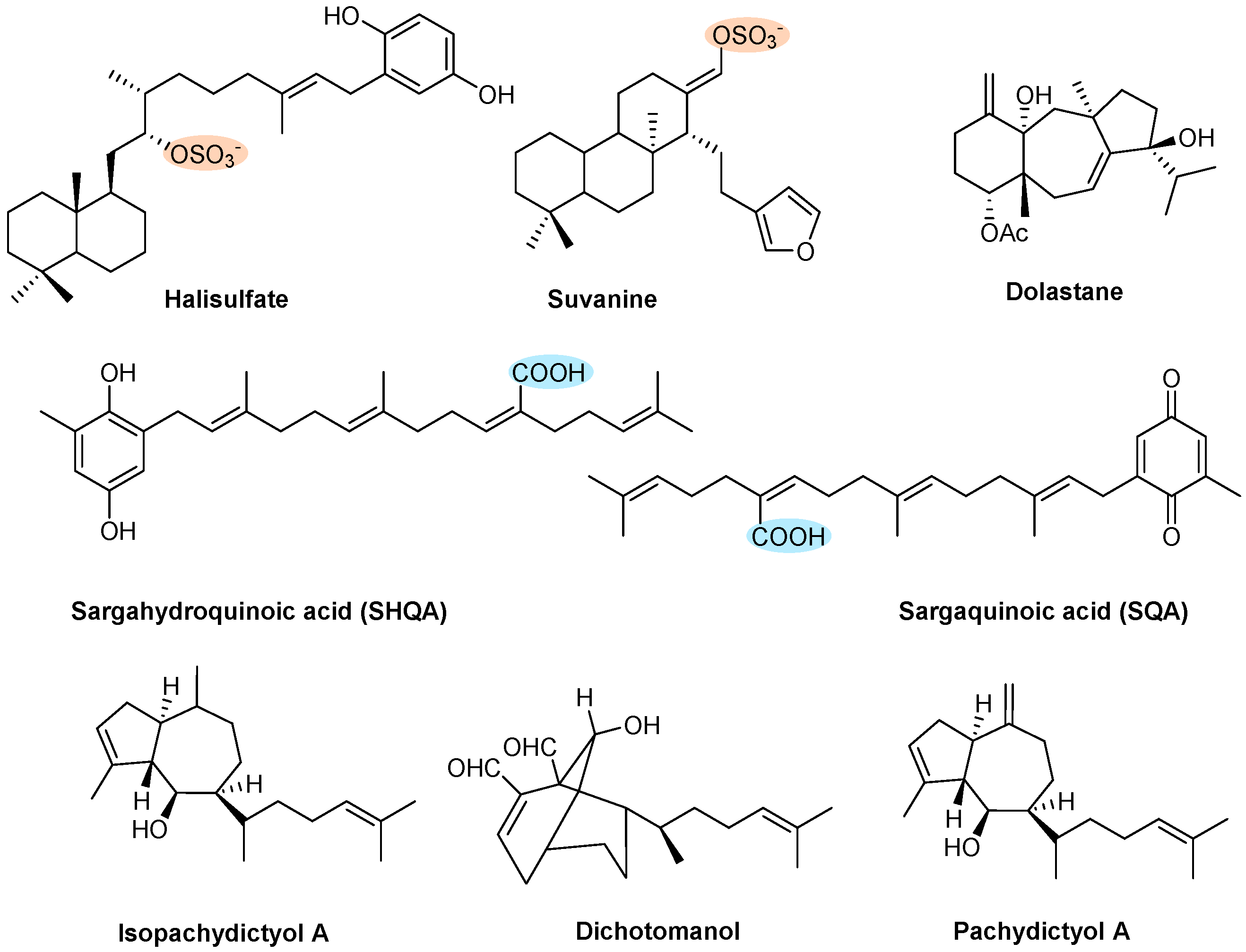
| Terpenes | Halisulfate and Suvanine | Sponge (Coscinoderma mathewsi) | Figure 11 | Θ Thrombin and trypsin | NT | [145] |
| SHQA and SQA | Algae (Sargassum micranthum and Sargassum yezoense) | Fast recovery time from paralysis (in vivo) | Θ Collagen-induced aggregation | [146,147] | ||
| Dolastane | Brown algae (Canistrocarpus cervicornis) | Θ Thrombin-induced coagulation | Θ ADP- or collagen-induced aggregation | [37] | ||
| Dichotomanol | Brown algae (Dictyota menstrualis) | ↑ APTT and PT Θ Thrombin-induced fibrinogen | Θ ADP- or collagen-induced aggregation | [148] | ||
| Pachydictyol A/Isopachydictyol | Brown algae (Dictyota menstrualis) | ↑ APTT Θ Thrombin-induced fibrinogen | Θ ADP- or thrombin-induced aggregation | [148] | ||
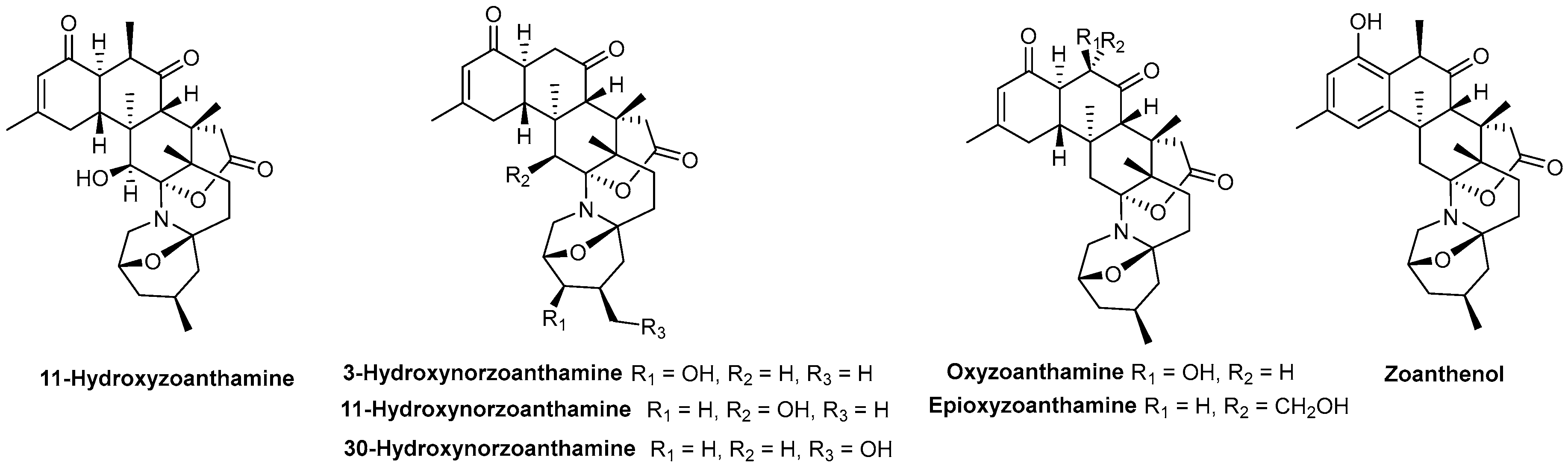
| Alkaloids | 11-Hydroxyzoanthamine | Zoanthids (Zoanthus Z. nymphaeus and an unidentified species of Zoanthus sp.) | Figure 10 | NT | Θ Thrombin-, collagen- or AA-induced aggregation | [150] |
| 3 and 11-Hydroxynorzoanthamine | NT | Θ Collagen- or AA-induced aggregation | [150] | |||
| Oxyzoanthamine, Zoethenol and Epioxyzoanthamine | NT | Θ Collagen-induced aggregation | [150] | |||
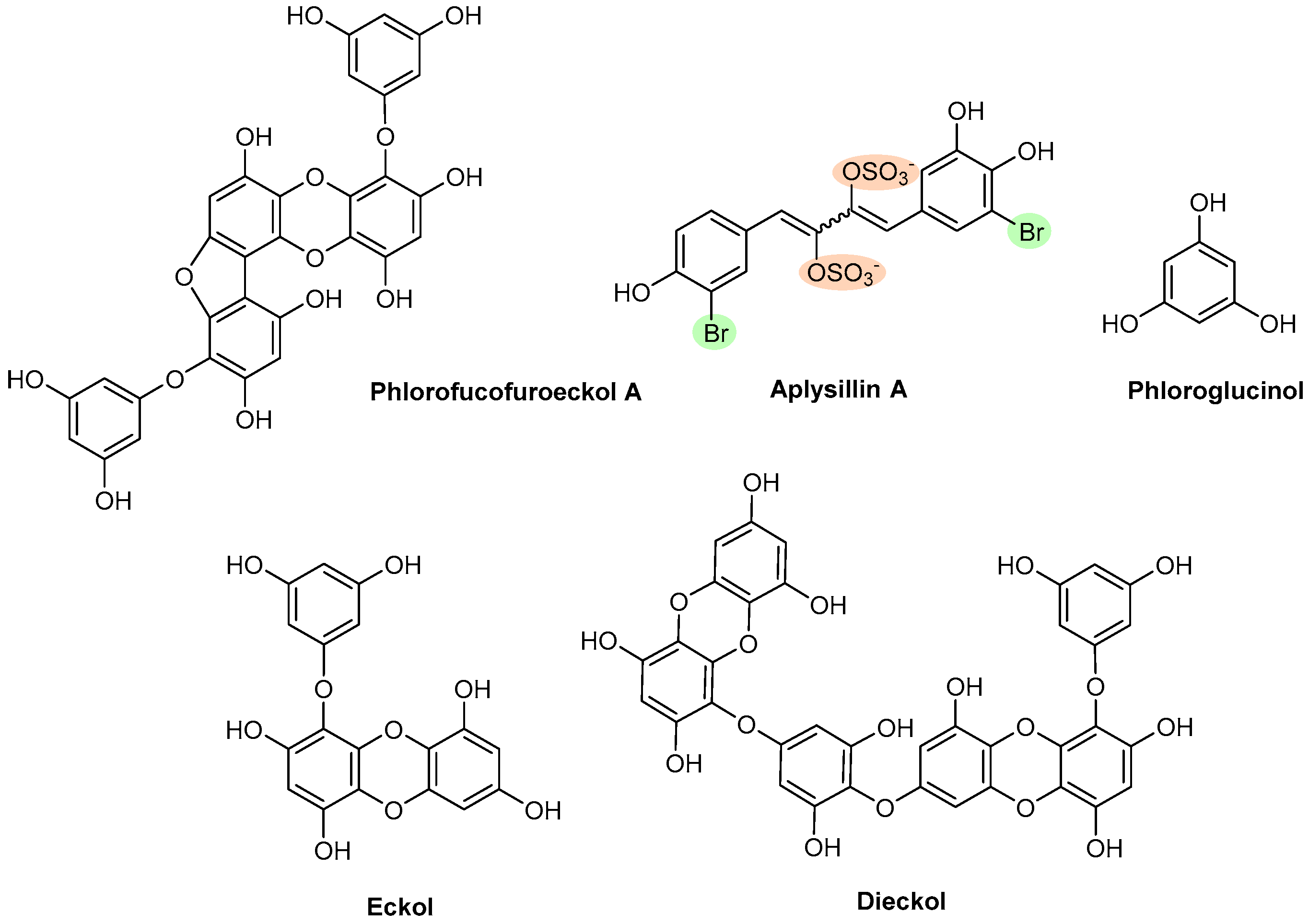
| Polyphenols | Phlorofucofuroeckol A | Brown algae (Ecklonia Kurome) | Figure 13 | Θ Plasmin inhibitors | NT | [43] |
| Aplysillin A | Sponge (Aplysina fistularis fulva) | Θ Thrombin to platelet membranes | NT | [151] | ||
| Phloroglucinol | Brown algae (Ecklonia species) | ↑ APTT and PT Θ Amidolytic activity of thrombin, in the absence of ATIII Θ FXa in the absence of ATIII ↑ Bleeding time (in vivo) | Θ AA-induced aggregation Θ COX-1 enzyme | [152,153] | ||
| Eckol and Dieckol | Brown algae (Ecklonia kurome) | ↑ APTT and PT Θ Amidolytic activity of thrombin, in the absence of ATIII Θ FXa in the absence of ATIII Θ Plasmin inhibitors | NT | [36] | ||
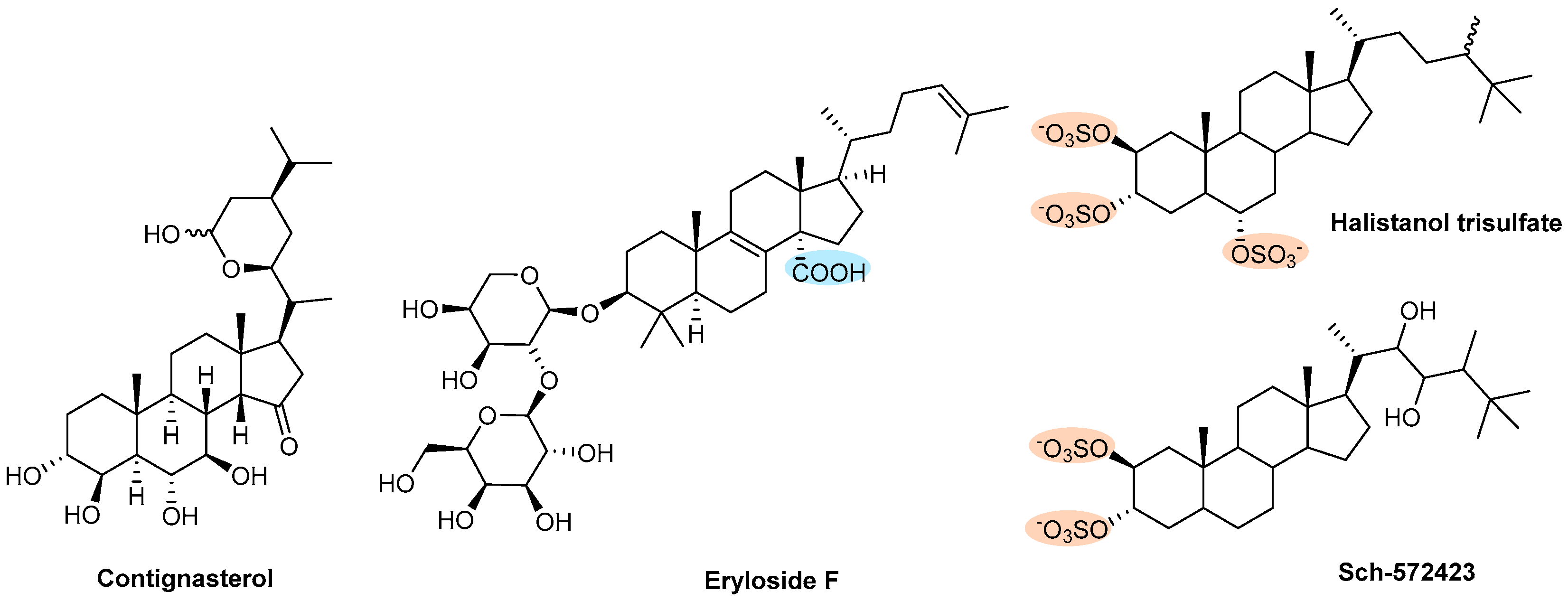
| Steroids | Contignasterol | Sponge (Petrosia contignata) | Figure 14 | NT | Θ PAF- or collagen-induced aggregation | [156] |
| Eryloside F | Sponge (Erylus formosus) | NT | Θ SFLLRN and U-46619 induced aggregation | [158] | ||
| Halistanol trisulfate and Sch-572423 | Sponge (Topsentia sp.) | NT | Θ Platelet aggregation acting through P2Y12 receptors | [159] | ||
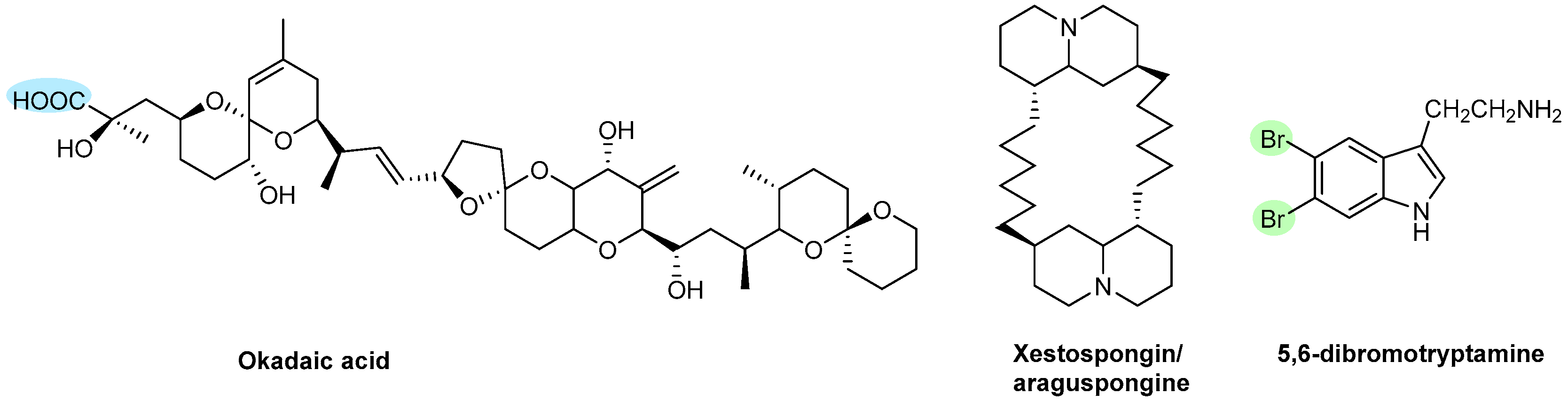
| Polyketides | Okadaic acid | Sponges (Halichondria okadai) | Figure 15 | NT | Θ Thrombin-induced aggregation Θ Protein phosphatases type 1 and type 2A Θ ATP release and increased cellular Ca2+ | [162] |
| Xestospongin/Araguspongine and 5,6-Dibromotryptamine | Sponge (Xestospongia sp.) | NT | Θ Collagen- or epinephrine-induced aggregation | [164] | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalhal, F.; Cristelo, R.R.; Resende, D.I.S.P.; Pinto, M.M.M.; Sousa, E.; Correia-da-Silva, M. Antithrombotics from the Sea: Polysaccharides and Beyond. Mar. Drugs 2019, 17, 170. https://doi.org/10.3390/md17030170
Carvalhal F, Cristelo RR, Resende DISP, Pinto MMM, Sousa E, Correia-da-Silva M. Antithrombotics from the Sea: Polysaccharides and Beyond. Marine Drugs. 2019; 17(3):170. https://doi.org/10.3390/md17030170
Chicago/Turabian StyleCarvalhal, Francisca, Ricardo R. Cristelo, Diana I. S. P. Resende, Madalena M. M. Pinto, Emília Sousa, and Marta Correia-da-Silva. 2019. "Antithrombotics from the Sea: Polysaccharides and Beyond" Marine Drugs 17, no. 3: 170. https://doi.org/10.3390/md17030170
APA StyleCarvalhal, F., Cristelo, R. R., Resende, D. I. S. P., Pinto, M. M. M., Sousa, E., & Correia-da-Silva, M. (2019). Antithrombotics from the Sea: Polysaccharides and Beyond. Marine Drugs, 17(3), 170. https://doi.org/10.3390/md17030170