Cosmetic and Therapeutic Applications of Fish Oil’s Fatty Acids on the Skin

 ,
,  ,
,
Abstract
1. Introduction
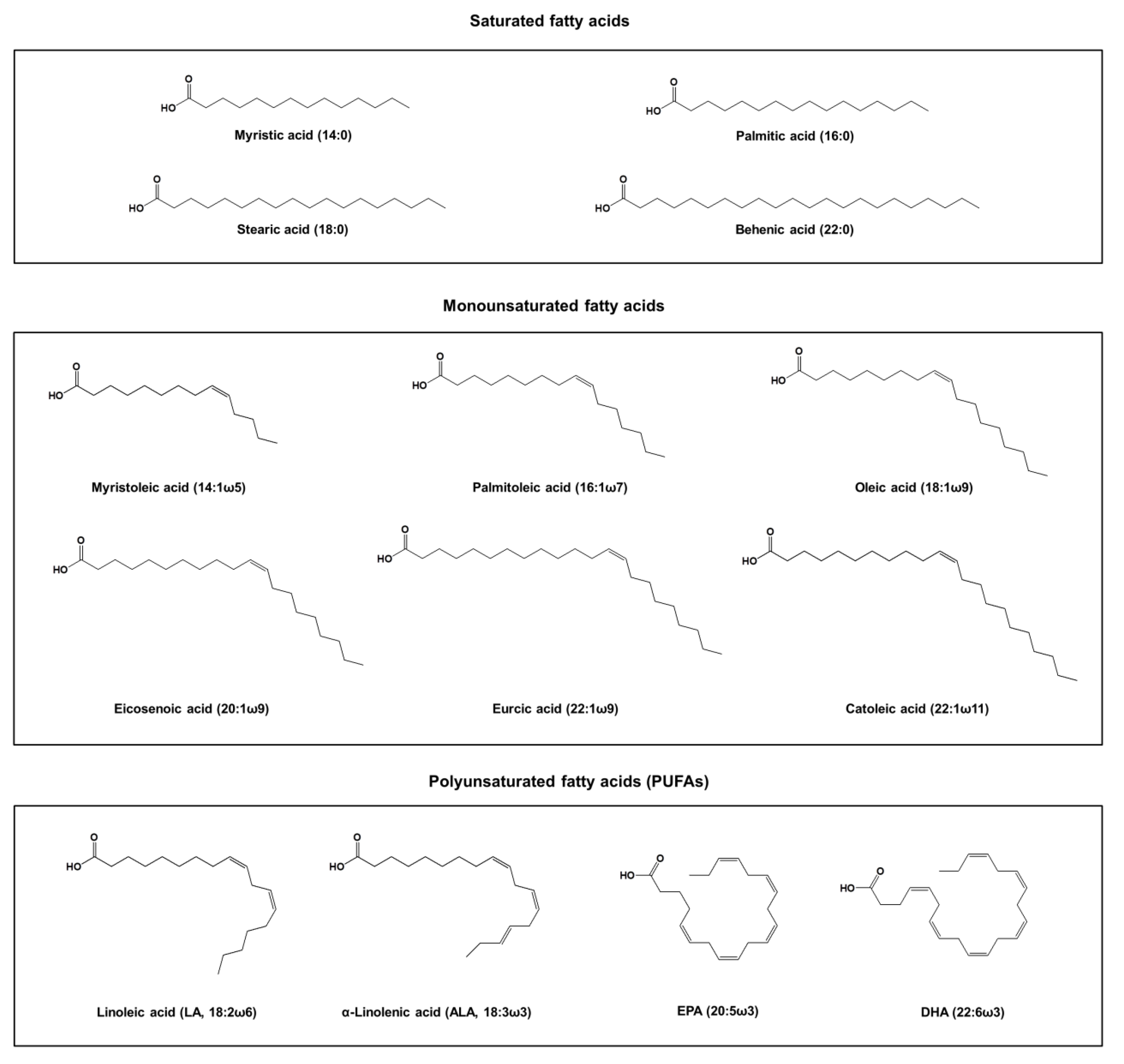
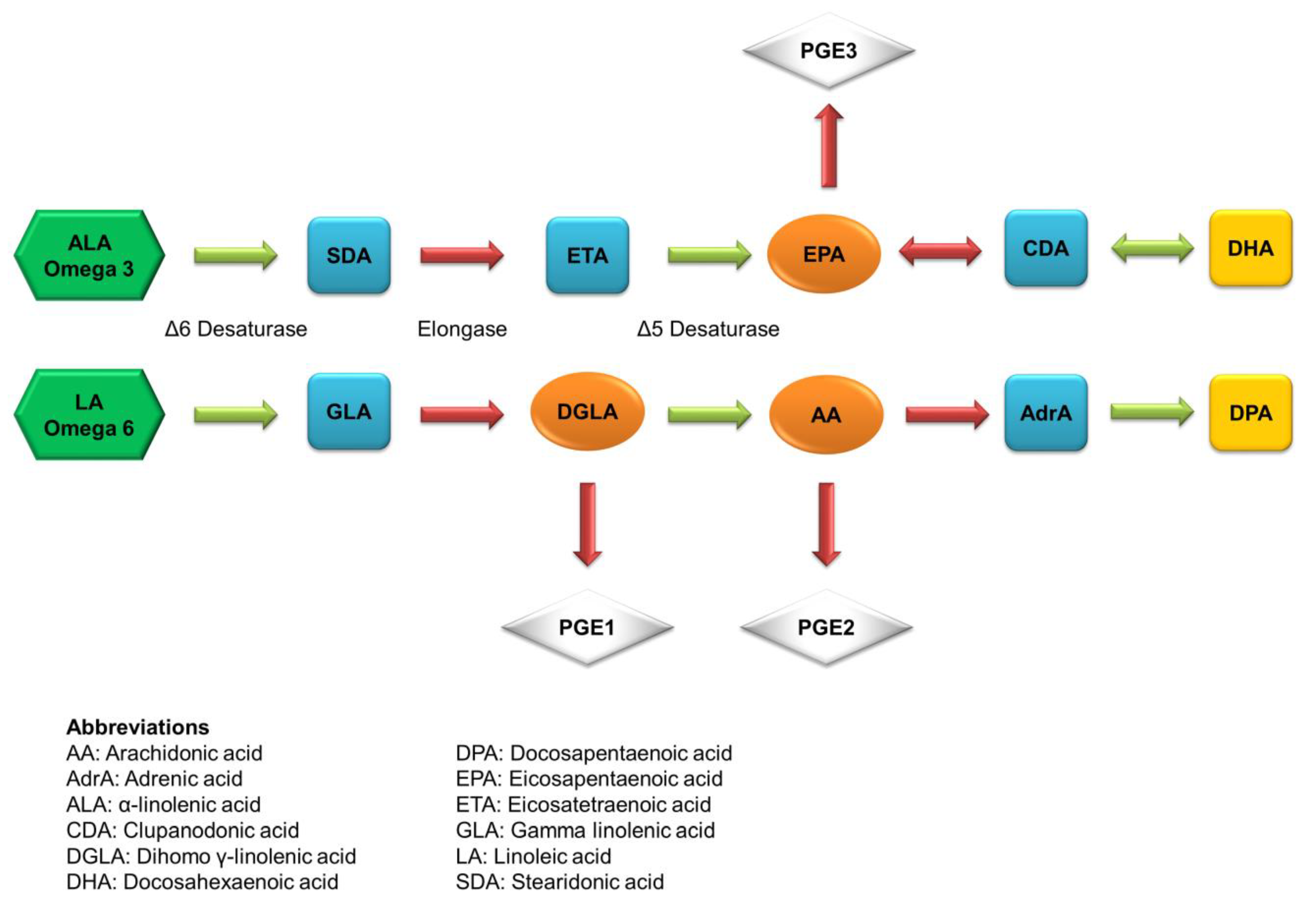
2. Fatty Acids of Fish Oil
3. Fatty Acids for Skin Disorder Prevention and Treatment
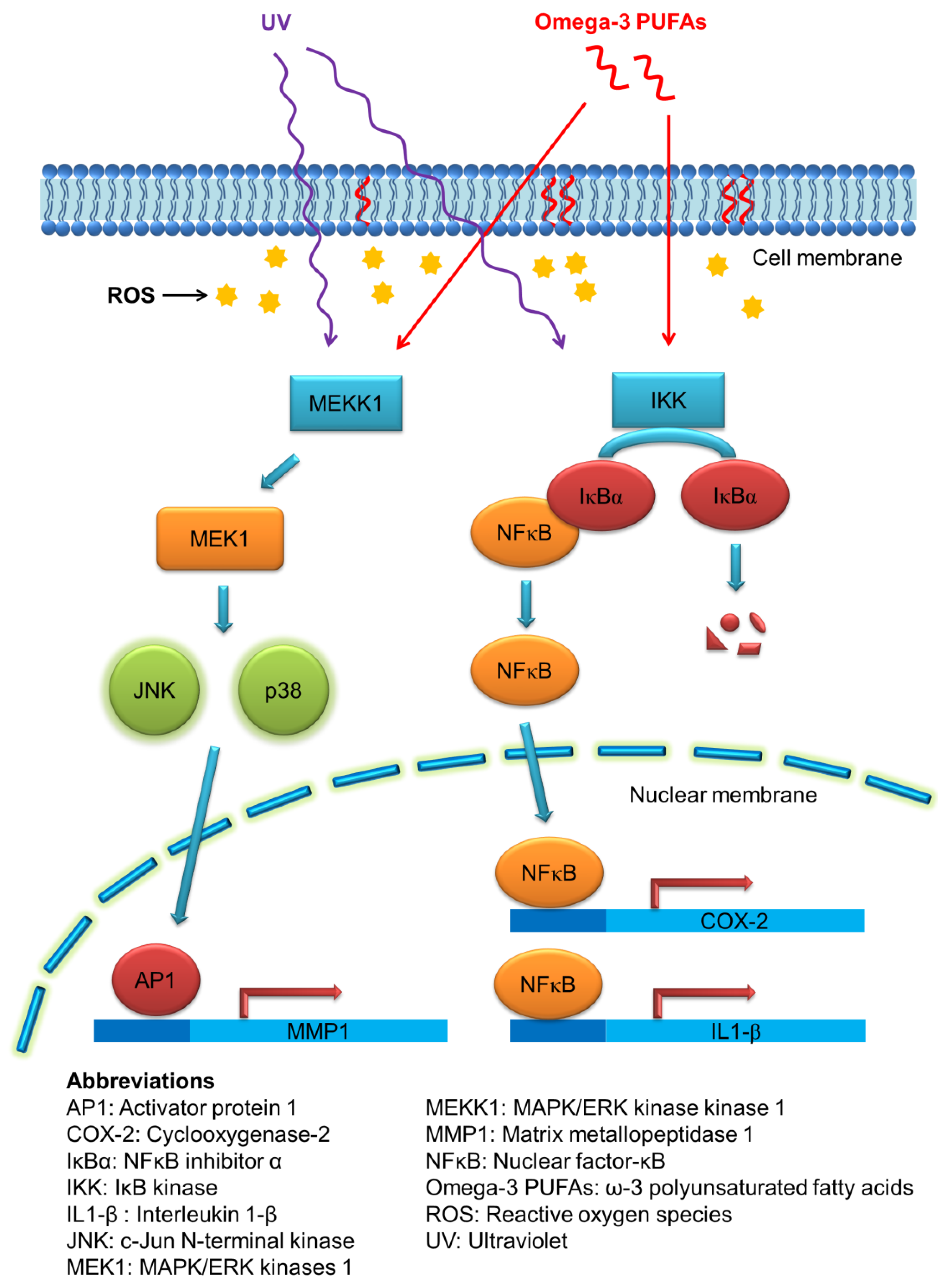
3.1. Photoaging
3.2. Cutaneous Carcinogenesis
3.3. Dermatitis
3.4. Cutaneous Wounds
3.5. Hyperpigmentation
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Rodríguez-Cruz, M.; Serna, D.S. Nutrigenomics of ω-3 fatty acids: Regulators of the master transcription factors. Nutrition 2017, 41, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Fodor, J.G.; Helis, E.; Yazdekhasti, N.; Vohnout, B. “Fishing” for the origins of the “Eskimos and heart disease” story: Facts or wishful thinking? Can. J. Cardiol. 2014, 30, 864–868. [Google Scholar] [CrossRef] [PubMed]
- Gahche, J.J.; Bailey, R.L.; Potischman, N.; Dwyer, J.T. Dietary supplement use was very high among older adults in the United States in 2011–2014. J. Nutr. 2017, 147, 1968–1976. [Google Scholar] [CrossRef] [PubMed]
- Villani, A.M.; Crotty, M.; Cleland, L.G.; James, M.J.; Fraser, R.J.; Cobiac, L.; Miller, M.D. Fish oil administration in older adults: Is there potential for adverse events? A systematic review of the literature. BMC Geriatr. 2013, 13, 41. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Chen, X.; Cao, F.; Sun, D.W.; He, Y.; Jiang, Y. Comparison of infrared spectroscopy and nuclear magnetic resonance techniques in tandem with multivariable selection for rapid determination of ω-3 polyunsaturated fatty acids in fish oil. Food Bioprocess Technol. 2014, 7, 1555–1569. [Google Scholar] [CrossRef]
- Hardy, M.S.; Kekic, A.; Graybill, N.L.; Lancaster, Z.R. A systematic review of the association between fish oil supplementation and the development of asthma exacerbations. SAGE Open Med. 2016, 4, 2050312116666216. [Google Scholar] [CrossRef] [PubMed]
- Eslick, G.D.; Howe, P.R.; Smith, C.; Priest, R.; Bensoussan, A. Benefits of fish oil supplementation in hyperlipidemia: A systematic review and meta-analysis. Int. J. Cardiol. 2009, 136, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Balk, E.M.; Lichtenstein, A.H. Omega-3 fatty acids and cardiovascular disease: Summary of the 2016 agency of healthcare research and quality evidence review. Nutrients 2017, 9, 865. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Geng, T.; Huang, T.; Zhao, Q. Fish oil supplementation and insulin sensitivity: A systematic review and meta-analysis. Lipids Health Dis. 2017, 16, 131. [Google Scholar] [CrossRef] [PubMed]
- Laviano, A.; Rianda, S.; Molfino, A.; Rossi Fanelli, F. Omega-3 fatty acids in cancer. Curr. Opin. Clin. Nutr. Metab. Care. 2013, 16, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Kumlin, M.; Ingelman-Sundberg, M.; Wolk, A. Dietary long-chain n-3 fatty acids for the prevention of cancer: A review of potential mechanisms. Am. J. Clin. Nutr. 2004, 79, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Rationale and use of n-3 fatty acids in artificial nutrition. Proc. Nutr. Soc. 2010, 69, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Abbasoglu, O.; Hardy, G.; Manzanares, W.; Pontes-Arruda, A. Fish oil-containing lipid emulsions in adult parenteral nutrition: A review of the evidence. J. Parenter. Enteral. Nutr. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lambertsen, N. Fatty acid composition of fish fats. Comparisons based on eight fatty acids. Fisk. Dir. Skr. Ser. Ernæring 1978, 1, 105–116. [Google Scholar]
- Meguro, S.; Arai, Y.; Masukawa, Y.; Uie, K.; Tokimitsu, I. Relationship between covalently bound ceramides and transepidermal water loss (TEWL). Arch. Dermatol. Res. 2000, 292, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Ekanayake-Mudiyanselage, S.; Aschauer, H.; Schmook, F.P.; Jensen, J.M.; Meingassner, J.G.; Proksch, E. Expression of epidermal keratins and the cornified envelope protein involucrin is influenced by permeability barrier disruption. J. Investig. Dermatol. 1998, 111, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Ilievska, B.; Loftsson, T.; Hjalmarsdottir, M.A.; Asgrimsdottir, G.M. Topical formulation comprising fatty acid extract from cod liver oil: Development, evaluation and stability studies. Mar. Drugs 2016, 14, 105. [Google Scholar] [CrossRef] [PubMed]
- Chee, K.M.; Gong, J.X.; Rees, D.M.; Meydani, M.; Ausman, L.; Johnson, J.; Siguel, E.N.; Schaefer, E.J. Fatty acid content of marine oil capsules. Lipids 1990, 25, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Porojnicu, A.C.; Bruland, O.S.; Aksnes, L.; Grant, W.B.; Moan, J. Sun beds and cod liver oil as vitamin D sources. J. Photochem. Photobiol. B 2008, 91, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Loftsson, T.; Gudmundsdóttir, T.K.; Fridriksdóttir, H.; Sigurdardóttir, A.M.; Thorkelsson, J.; Gudmundsson, G.; Hjaltason, B. Fatty acids from cod-liver oil as skin penetration enhancers. Pharmazie 1995, 50, 188–190. [Google Scholar] [PubMed]
- Osman, H.; Suriah, A.R.; Law, E.C. Fatty acid composition and cholesterol content of selected marine fish in Malaysian waters. Food Chem. 2001, 73, 55–60. [Google Scholar] [CrossRef]
- Saify, Z.S.; Akhtar, S.; Hassan, S.; Arif, M.; Ahmed, F.; Siddiqui, S. A study on fatty acid composition of fish oil from two marine fish, Eusphyra blochii and Carcharhinus bleekeri. Pak. J. Pharm. Sci. 2000, 13, 5–12. [Google Scholar] [PubMed]
- Ruiz-López, N.; Sayanova, O.; Napier, J.A.; Haslam, R.P. Metabolic engineering of the omega-3 long chain polyunsaturated fatty acid biosynthetic pathway into transgenic plants. J. Exp. Bot. 2012, 63, 2397–2410. [Google Scholar] [CrossRef] [PubMed]
- Bibus, D.; Lands, B. Balancing proportions of competing omega-3 and omega-6 highly unsaturated fatty acids (HUFA) in tissue lipids. Prostaglandins Leukot. Essent. Fatty Acids 2015, 99, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.D.; Ali, R.; Khasira, M.A.; Odera, D.; West, A.L.; Koster, G.; Akomo, P.; Talbert, A.W.; Goss, V.M.; Ngari, M.; et al. Ready-to-use therapeutic food with elevated n-3 polyunsaturated fatty acid content, with or without fish oil, to treat severe acute malnutrition: A randomized controlled trial. BMC Med. 2015, 13, 93. [Google Scholar] [CrossRef] [PubMed]
- Horrobin, D.F. Essential fatty acid metabolism and its modification in atopic eczema. Am. J. Clin. Nutr. 2000, 71 (Suppl. 1), 367S–372S. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Yokomizo, T. The role of leukotrienes in allergic diseases. Allergol. Int. 2015, 64, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Breiden, B.; Sandhoff, K. The role of sphingolipid metabolism in cutaneous permeability barrier formation. Biochim. Biophys. Acta 2014, 1841, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, A.J.; Li, D. The significance of alpha-linolenic acid for humans. J. Oleo Sci. 2001, 50, 373–379. [Google Scholar] [CrossRef]
- Lam, S.M.; Wang, Z.; Li, J.; Huang, X.; Shui, G. Sequestration of polyunsaturated fatty acids in membrane phospholipids of Caenorhabditis elegans dauer larva attenuates eicosanoid biosynthesis for prolonged survival. Redox Biol. 2017, 12, 967–977. [Google Scholar] [CrossRef] [PubMed]
- McCusker, M.M.; Grant-Kels, J.M. Healing fats of the skin: The structural and immunologic roles of the omega-6 and omega-3 fatty acids. Clin. Dermatol. 2010, 28, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R. The role of epidermal lipids in cutaneous permeability barrier homeostasis. J. Lipid Res. 2007, 48, 2531–2546. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Hossain, S.; Al Mamun, A.; Matsuzaki, K.; Arai, H. Docosahexaenoic acid: One molecule diverse functions. Crit. Rev. Biotechnol. 2017, 37, 579–597. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.Y.; Ma, T.; Winterthun, S.; Kristiansen, K.; Frøyland, L.; Madsen, L. β-oxidation modulates metabolic competition between eicosapentaenoic acid and arachidonic acid regulating prostaglandin E2 synthesis in rat hepatocytes—Kupffer cells. Biochim. Biophys. Acta 2010, 1801, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Lorente-Cebrián, S.; Costa, A.G.; Navas-Carretero, S.; Zabala, M.; Laiglesia, L.M.; Martínez, J.A.; Moreno-Aliaga, M.J. An update on the role of omega-3 fatty acids on inflammatory and degenerative diseases. J. Physiol. Biochem. 2015, 71, 341–349. [Google Scholar] [CrossRef] [PubMed]
- de Gruijl, F.R. UV adaptation: Pigmentation and protection against overexposure. Exp. Dermatol. 2017, 26, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Wagener, F.A.; Carels, C.E.; Lundvig, D.M. Targeting the redox balance in inflammatory skin conditions. Int. J. Mol. Sci. 2013, 14, 9126–9167. [Google Scholar] [CrossRef] [PubMed]
- Rundhaug, J.E.; Fischer, S.M. Cyclo-oxygenase-2 plays a critical role in UV-induced skin carcinogenesis. Photochem. Photobiol. 2008, 84, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Mechanisms of action of (n-3) fatty acids. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef] [PubMed]
- Pilkington, S.M.; Watson, R.E.; Nicolaou, A.; Rhodes, L.E. Omega-3 polyunsaturated fatty acids: Photoprotective macronutrients. Exp. Dermatol. 2011, 20, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Pernet, I.; Sagot, V.; Schmitt, D.; Viac, J. UVA1 and UVB radiation but not PGE2 stimulate IL-8 release in normal human keratinocytes. Arch. Dermatol. Res. 1999, 291, 527–529. [Google Scholar] [CrossRef] [PubMed]
- Storey, A.; McArdle, F.; Friedmann, P.S.; Jackson, M.J.; Rhodes, L.E. Eicosapentaenoic acid and docosahexaenoic acid reduce UVB- and TNF-alpha-induced IL-8 secretion in keratinocytes and UVB-induced IL-8 in fibroblasts. J. Investig. Dermatol. 2005, 124, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Serini, S.; Donato, V.; Piccioni, E.; Trombino, S.; Monego, G.; Toesca, A.; Innocenti, I.; Missori, M.; De Spirito, M.; Celleno, L.; Fasano, E.; Ranelletti, F.O.; Calviello, G. Docosahexaenoic acid reverts resistance to UV-induced apoptosis in human keratinocytes: Involvement of COX-2 and HuR. J. Nutr. Biochem. 2011, 22, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Amano, S. Characterization and mechanisms of photoageing-related changes in skin. Damages of basement membrane and dermal structures. Exp. Dermatol. 2016, 25 (Suppl. 3), 14–19. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.H.; Shin, C.M.; Park, C.H.; Kim, K.H.; Cho, K.H.; Eun, H.C.; Chung, J.H. Eicosapentaenoic acid inhibits UV-induced MMP-1 expression in human dermal fibroblasts. J. Lipid Res. 2005, 46, 1712–1720. [Google Scholar] [CrossRef] [PubMed]
- Danno, K.; Ikai, K.; Imamura, S. Anti-inflammatory effects of eicosapentaenoic acid on experimental skin inflammation models. Arch. Dermatol. Res. 1993, 285, 432–435. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Kundu, J.K.; Shin, J.W.; Na, H.K.; Surh, Y.J. Docosahexaenoic acid inhibits UVB-induced activation of NF-κB and expression of COX-2 and NOX-4 in HR-1 hairless mouse skin by blocking MSK1 signaling. PLoS One 2011, 6, e28065. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Park, S.Y.; Hwang, E.; Park, B.; Seo, S.A.; Cho, J.G.; Zhang, M.; Yi, T.H. Dietary Foeniculum vulgare Mill extract attenuated UVB irradiation-induced skin photoaging by activating of Nrf2 and inhibiting MAPK pathways. Phytomedicine 2016, 23, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Yum, H.W.; Park, J.; Park, H.J.; Shin, J.W.; Cho, Y.Y.; Kim, S.J.; Kang, J.X.; Surh, Y.J. Endogenous ω-3 fatty acid production by fat-1 transgene and topically applied docosahexaenoic acid protect against UVB-induced mouse skin carcinogenesis. Sci. Rep. 2017, 7, 11658. [Google Scholar] [CrossRef] [PubMed]
- Orengo, I.F.; Black, H.S.; Wolf, J.E., Jr. Influence of fish oil supplementation on the minimal erythema dose in humans. Arch. Dermatol. Res. 1992, 284, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.E.; Durham, B.H.; Fraser, W.D.; Friedmann, P.S. Dietary fish oil reduces basal and ultraviolet B-generated PGE2 levels in skin and increases the threshold to provocation of polymorphic light eruption. J. Investig. Dermatol. 1995, 105, 532–535. [Google Scholar] [CrossRef] [PubMed]
- Puglia, C.; Tropea, S.; Rizza, L.; Santagati, N.A.; Bonina, F. In vitro percutaneous absorption studies and in vivo evaluation of anti-inflammatory activity of essential fatty acids (EFA) from fish oil extracts. Int. J. Pharm. 2005, 299, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Shahbakhti, H.; Watson, R.E.; Azurdia, R.M.; Ferreira, C.Z.; Garmyn, M.; Rhodes, L.E. Influence of eicosapentaenoic acid, an omega-3 fatty acid, on ultraviolet-B generation of prostaglandin-E2 and proinflammatory cytokines interleukin-1β, tumor necrosis factor-α, interleukin-6 and interleukin-8 in human skin in vivo. Photochem. Photobiol. 2004, 80, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Clausen, B.E.; Kel, J.M. Langerhans cells: Critical regulators of skin immunity? Immunol. Cell Biol. 2010, 88, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Pilkington, S.M.; Gibbs, N.K.; Costello, P.; Bennett, S.P.; Massey, K.A.; Friedmann, P.S.; Nicolaou, A.; Rhodes, L.E. Effect of oral eicosapentaenoic acid on epidermal Langerhans cell numbers and PGD2 production in UVR-exposed human skin: A randomised controlled study. Exp. Dermatol. 2016, 25, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.H.; Cho, S.; Lee, S.; Kim, K.H.; Cho, K.H.; Eun, H.C.; Chung, J.H. Photoprotective and anti-skin-aging effects of eicosapentaenoic acid in human skin in vivo. J. Lipid Res. 2006, 47, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Takemura, N.; Takahashi, K.; Tanaka, H.; Ihara, Y.; Ikemoto, A.; Fujii, Y.; Okuyama, H. Dietary, but not topical, alpha-linolenic acid suppresses UVB-induced skin injury in hairless mice when compared with linoleic acids. Photochem. Photobiol. 2002, 76, 657–663. [Google Scholar] [CrossRef]
- Tsuboyama-Kasaoka, N.; Takahashi, M.; Tanemura, K.; Kim, H.J.; Tange, T.; Okuyama, H.; Kasai, M.; Ikemoto, S.; Ezaki, O. Conjugated linoleic acid supplementation reduces adipose tissue by apoptosis and develops lipodystrophy in mice. Diabetes 2000, 49, 1534–1542. [Google Scholar] [CrossRef] [PubMed]
- Fuke, G.; Nornberg, J.L. Systematic evaluation on the effectiveness of conjugated linoleic acid in human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Storey, A.; Rogers, J.S.; McArdle, F.; Jackson, M.J.; Rhodes, L.E. Conjugated linoleic acids modulate UVR-induced IL-8 and PGE2 in human skin cells: Potential of CLA isomers in nutritional photoprotection. Carcinogenesis 2007, 28, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Kabri, T.H.; Arab-Tehrany, E.; Belhaj, N.; Linder, M. Physico-chemical characterization of nano-emulsions in cosmetic matrix enriched on omega-3. J. Nanobiotechnology 2011, 9, 41. [Google Scholar] [CrossRef] [PubMed]
- Molho-Pessach, V.; Lotem, M. Ultraviolet radiation and cutaneous carcinogenesis. Curr. Probl. Dermatol. 2007, 35, 14–27. [Google Scholar] [PubMed]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K. UV-induced immune suppression and photocarcinogenesis: Chemoprevention by dietary botanical agents. Cancer Lett. 2007, 255, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nikolakopoulou, Z.; Shaikh, M.H.; Dehlawi, H.; Michael-Titus, A.T.; Parkinson, E.K. The induction of apoptosis in pre-malignant keratinocytes by omega-3 polyunsaturated fatty acids docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) is inhibited by albumin. Toxicol. Lett. 2013, 218, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.C.M.; Tong, K.F.; Ooi, V.E.C. Cytostatic and cytotoxic effects of cyclooxygenase inhibitors and their synergy with docosahexaenoic acid on the growth of human skin melanoma A-375 cells. Biomed. Pharmacother. 2005, 59 (Suppl. 2), S293–S297. [Google Scholar] [CrossRef]
- Elmets, C.A.; Ledet, J.J.; Athar, M. Cyclooxygenases: Mediators of UV-induced skin cancer and potential targets for prevention. J. Investig. Dermatol. 2014, 134, 2497–2502. [Google Scholar] [CrossRef] [PubMed]
- Hanna, E.; Abadi, R.; Abbas, O. Imiquimod in dermatology: An overview. Int. J. Dermatol. 2016, 55, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Rehman, K.; Mohd Amin, M.C.; Yuen, N.P.; Zulfakar, M.H. Immunomodulatory effectiveness of fish oil and omega-3 fatty acids in human non-melanoma skin carcinoma cells. J. Oleo Sci. 2016, 65, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Rehman, K.; Zulfakar, M.H. Novel fish oil-based bigel system for controlled drug delivery and its influence on immunomodulatory activity of imiquimod against skin cancer. Pharm. Res. 2017, 34, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Lupi, F.R.; Gentile, L.; Gabriele, D.; Mazzulla, S.; Baldino, N.; de Cindio, B. Olive oil and hyperthermal water bigels for cosmetic uses. J. Colloid Interface Sci. 2015, 459, 70–78. [Google Scholar]
- Richards, H.; Thomas, C.P.; Bowen, J.L.; Heard, C.M. In-vitro transcutaneous delivery of ketoprofen and polyunsaturated fatty acids from a pluronic lecithin organogel vehicle containing fish oil. J. Pharm. Pharmacol. 2006, 58, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, G.; Das, U.N. Effect of evening primrose and fish oils on two stage skin carcinogenesis in mice. Prostaglandins Leukot. Essent. Fatty Acids 1998, 59, 155–161. [Google Scholar] [CrossRef]
- Lou, Y.R.; Peng, Q.Y.; Li, T.; Medvecky, C.M.; Lin, Y.; Shih, W.J.; Conney, A.H.; Shapses, S.; Wagner, G.C.; Lu, Y.P. Effects of high-fat diets rich in either omega-3 or omega-6 fatty acids on UVB-induced skin carcinogenesis in SKH-1 mice. Carcinogenesis 2011, 32, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Hakim, I.A.; Harris, R.B.; Ritenbaugh, C. Fat intake and risk of squamous cell carcinoma of the skin. Nutr. Cancer 2000, 36, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Fortes, C.; Mastroeni, S.; Melchi, F.; Pilla, M.A.; Antonelli, G.; Camaioni, D.; Alotto, M.; Pasquini, P. A protective effect of the Mediterranean diet for cutaneous melanoma. Int. J. Epidemiol. 2008, 37, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Homsi, J.; Bedikian, A.Y.; Kim, K.B.; Papadopoulos, N.E.; Hwu, W.J.; Mahoney, S.L.; Hwu, P. Phase 2 open-label study of weekly docosahexaenoic acid-paclitaxel in cutaneous and mucosal metastatic melanoma patients. Melanoma Res. 2009, 19, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Bradley, M.O.; Webb, N.L.; Anthony, F.H.; Devanesan, P.; Witman, P.A.; Hemamalini, S.; Chander, M.C.; Baker, S.D.; He, L.; Horwitz, S.B.; Swindell, C.S. Tumor targeting by covalent conjugation of a natural fatty acid to paclitaxel. Clin. Cancer Res. 2001, 7, 3229–3238. [Google Scholar] [PubMed]
- Bradley, M.O.; Swindell, C.S.; Anthony, F.H.; Witman, P.A.; Devanesan, P.; Webb, N.L.; Baker, S.D.; Wolff, A.C.; Donehower, R.C. Tumor targeting by conjugation of DHA to paclitaxel. J. Control. Release 2001, 74, 233–236. [Google Scholar] [CrossRef]
- Belury, M.A.; Kavanaugh, C.J.; Liu, K.L. Conjugated linoleic acid modulates phorbol ester–induced PPAR-δ and K-FABP mRNA expression in mouse skin. Nutr. Res. 2007, 27, 48–55. [Google Scholar] [CrossRef]
- Leung, D.Y.M.; Bieber, T. Atopic dermatitis. Lancet 2003, 361, 151–160. [Google Scholar] [CrossRef]
- Williams, H.C. Atopic dermatitis. N. Engl. J. Med. 2005, 352, 2314–2324. [Google Scholar] [CrossRef] [PubMed]
- Barcelos, R.C.; de Mello-Sampayo, C.; Antoniazzi, C.T.; Segat, H.J.; Silva, H.; Veit, J.C.; Piccolo, J.; Emanuelli, T.; Bürger, M.E.; Silva-Lima, B.; et al. Oral supplementation with fish oil reduces dryness and pruritus in the acetone-induced dry skin rat model. J. Dermatol. Sci. 2015, 79, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.P.; Yang, S.H.; Lee, C.H.; Aljuffali, I.A.; Kao, H.C.; Fang, J.Y. What is the discrepancy between drug permeation into/across intact and diseased skins? Atopic dermatitis as a model. Int. J. Pharm. 2016, 497, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Basketter, D.A.; Kimber, I. Phthalic anhydride: Illustrating a conundrum in chemical allergy. J. Immunotoxicol. 2016, 13, 767–769. [Google Scholar] [CrossRef] [PubMed]
- Abdel Latif, M.; Abdul-Hamid, M.; Galaly, S.R. Effect of diethylcarbamazine citrate and omega-3 fatty acids on trimellitic anhydride-induced rat skin allergy. Asian Pac. J. Allergy Immunol. 2015, 33, 33–41. [Google Scholar] [PubMed]
- Weise, C.; Heunemann, C.; Loddenkemper, C.; Herz, U.; van Tol, E.A.; Worm, M. Dietary docosahexaenoic acid in combination with arachidonic acid ameliorates allergen-induced dermatitis in mice. Pediatr. Allergy Immunol. 2011, 22, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Kong, S.; Seong, K.; Cho, Y. γ-Linolenic acid in borage oil reverses epidermal hyperproliferation in guinea pigs. J. Nutr. 2002, 132, 3090–3097. [Google Scholar] [CrossRef] [PubMed]
- Brosche, T.; Platt, D. Effect of borage oil consumption on fatty acid metabolism, transepidermal water loss and skin parameters in elderly people. Arch. Gerontol. Geriatr. 2000, 30, 139–150. [Google Scholar] [CrossRef]
- Kawamura, A.; Ooyama, K.; Kojima, K.; Kachi, H.; Abe, T.; Amano, K.; Aoyama, T. Dietary supplementation of gamma-linolenic acid improves skin parameters in subjects with dry skin and mild atopic dermatitis. J. Oleo Sci. 2011, 60, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.; Eng, P.A.; Borelli, S.; Kägi, R.; Zimmermann, C.; Zahner, C.; Drewe, J.; Hess, L.; Ferrari, G.; Lautenschlager, S.; et al. Gamma-linolenic acid levels correlate with clinical efficacy of evening primrose oil in patients with atopic dermatitis. Adv. Ther. 2014, 31, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, H.; Tateishi, N.; Shiraishi, A.; Teraoka, N.; Tanaka, T.; Tanaka, A.; Matsuda, H.; Kiso, Y. Oral administration of dihomo-gamma-linolenic acid prevents development of atopic dermatitis in NC/Nga mice. Lipids 2008, 43, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Amagai, Y.; Oida, K.; Matsuda, A.; Jung, K.; Kakutani, S.; Tanaka, T.; Matsuda, K.; Jang, H.; Ahn, G.; Xia, Y.; Kawashima, H.; Shibata, H.; Matsuda, H.; Tanaka, A. Dihomo-γ-linolenic acid prevents the development of atopic dermatitis through prostaglandin D1 production in NC/Tnd mice. J. Dermatol. Sci. 2015, 79, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Lordani, T.V.A.; de Lara, C.E.; Ferreira, F.B.P.; de Souza Terron Monich, M.; Mesquita da Silva, C.; Felicetti Lordani, C.R.; Giacomini Bueno, F.; Vieira Teixeira, J.J.; Lonardoni, M.V.C. Therapeutic effects of medicinal plants on cutaneous wound healing in humans: A systematic review. Mediators Inflamm. 2018, 2018, 7354250. [Google Scholar] [CrossRef] [PubMed]
- Baum, C.L.; Arpey, C.J. Normal cutaneous wound healing: Clinical correlation with cellular and molecular events. Dermatol. Surg. 2005, 31, 674–686. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Kiecolt-Glaser, J.K.; Glaser, R.; Christian, L.M. Omega-3 fatty acids and stress-induced immune dysregulation: Implications for wound healing. Mil. Med. 2014, 179 (Suppl. 11), 129–133. [Google Scholar] [CrossRef] [PubMed]
- Shingel, K.I.; Faure, M.P.; Azoulay, L.; Roberge, C.; Deckelbaum, R.J. Solid emulsion gel as a vehicle for delivery of polyunsaturated fatty acids: Implications for tissue repair, dermal angiogenesis and wound healing. J. Tissue Eng. Regen. Med. 2008, 2, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Yao, X.; Zeng, R.; Sun, R.; Tian, H.; Shi, C.; Li, L.; Tian, J.; Yang, K. Safety and efficacy of a new parenteral lipid emulsion (SMOF) for surgical patients: A systematic review and meta-analysis of randomized controlled trials. Nutr. Rev. 2013, 71, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.C.; Yang, F.L.; Subeq, Y.M.; Tien, C.C.; Chao, Y.C.; Lee, R.P. Lipid emulsion enriched in omega-3 PUFA accelerates wound healing: A placebo-controlled animal study. World J. Surg. 2018, 42, 1714–1720. [Google Scholar] [CrossRef] [PubMed]
- Arantes, E.L.; Dragano, N.; Ramalho, A.; Vitorino, D.; de-Souza, G.F.; Lima, M.H.; Velloso, L.A.; Araújo, E.P. Topical docosahexaenoic acid (DHA) accelerates skin wound healing in rats and activates GPR120. Biol. Res. Nurs. 2016, 18, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Tian, H.; Hong, S. Novel 14,21-dihydroxy-docosahexaenoic acids: Structures, formation pathways, and enhancement of wound healing. J. Lipid Res. 2010, 51, 923–932. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, J.C.; Belury, M.; Ahijevych, K.; Blakely, W. Omega-3 fatty acids effect on wound healing. Wound Repair Regen. 2008, 16, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, C.R.; Souza, M.A.; Ferro, E.A.; Favoreto, S., Jr.; Pena, J.D. Influence of topical administration of n-3 and n-6 essential and n-9 nonessential fatty acids on the healing of cutaneous wounds. Wound Repair Regen. 2004, 12, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.M.; Hatanaka, E.; Martins, E.F.; Oliveira, F.; Liberti, E.A.; Farsky, S.H.; Curi, R.; Pithon-Curi, T.C. Effect of oleic and linoleic acids on the inflammatory phase of wound healing in rats. Cell Biochem. Funct. 2008, 26, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, H.G.; Vinolo, M.A.; Sato, F.T.; Magdalon, J.; Kuhl, C.M.; Yamagata, A.S.; Pessoa, A.F.; Malheiros, G.; Dos Santos, M.F.; et al. Oral administration of linoleic acid induces new vessel formation and improves skin wound healing in diabetic rats. PLoS ONE 2016, 11, e0165115. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.C.; Goldman, M.P. A Topical anti-inflammatory healing regimen Utilizing conjugated linolenic acid for use post-ablative laser resurfacing of the face: A randomized, controlled trial. J. Clin. Aesthet. Dermatol. 2017, 10, 12–17. [Google Scholar] [PubMed]
- Kondo, T.; Hearing, V.J. Update on the regulation of mammalian melanocyte function and skin pigmentation. Expert Rev. Dermatol. 2011, 6, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Kondoh, H.; Ichihashi, M.; Hearing, V.J. Approaches to identify inhibitors of melanin biosynthesis via the quality control of tyrosinase. J. Investig. Dermatol. 2007, 127, 751–761. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed]
- Balcos, M.C.; Kim, S.Y.; Jeong, H.S.; Yun, H.Y.; Baek, K.J.; Kwon, N.S.; Park, K.C.; Kim, D.S. Docosahexaenoic acid inhibits melanin synthesis in murine melanoma cells in vitro through increasing tyrosinase degradation. Acta Pharmacol. Sin. 2014, 35, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, M. MITF: A stream flowing for pigment cells. Pigment Cell Res. 2000, 13, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Funasaka, Y.; Oka, M.; Ohashi, A.; Furumura, M.; Matsunaga, J.; Matsunaga, N.; Hearing, V.J.; Ichihashi, M. Possible involvement of proteolytic degradation of tyrosinase in the regulatory effect of fatty acids on melanogenesis. J. Lipid Res. 1999, 40, 1312–1316. [Google Scholar] [PubMed]
- Ando, H.; Ryu, A.; Hashimoto, A.; Oka, M.; Ichihashi, M. Linoleic acid and α-linolenic acid lightens ultraviolet-induced hyperpigmentation of the skin. Arch. Dermatol. Res. 1998, 290, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Shigeta, Y.; Imanaka, H.; Ando, H.; Ryu, A.; Oku, N.; Baba, N.; Makino, T. Skin whitening effect of linoleic acid is enhanced by liposomal formulations. Biol. Pharm. Bull. 2004, 27, 591–594. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Composition | Experimental Model | UV Type | Benefit | Reference |
|---|---|---|---|---|
| DHA and EPA | Keratinocytes and skin fibroblasts | UV 270–400 nm, 25–100 mJ/cm2 | Reduced IL-8 | Storey et al. [43] |
| DHA | Keratinocytes | UV 290–400 nm, 10–60 mJ/cm2 | Reverted cell resistance to UV-induced apoptosis | Serini et al. [44] |
| EPA | Skin fibroblasts | UV 275–380 nm, 25–75 mJ/cm2 | Reduced MMP-1, ERK, and JNK | Kim et al. [46] |
| EPA | BALB/c mouse with ear edema | UVB, 500 mJ/cm2 | Suppressed ear edema by oral EPA administration | Danno et al. [47] |
| DHA | Hairless mouse with skin inflammation | UVB 312 nm, 180 mJ/cm2 | Decreased COX-2 and NOX-4 by blocking MSK1 | Rahman et al. [48] |
| DHA | Hairless mouse with skin inflammation | UVB 312 nm, 180 mJ/cm2 | Elevated Nrf2 activation and upregulation of cytoprotective genes | Yum et al. [50] |
| Fish oil rich of DHA and EPA | Human | UVB with a filter to eliminate wavelengths <295 nm | Increased minimal erythema dose and decreased serum triglyceride | Orengo et al. [51] |
| Fish oil rich in DHA and EPA | Human | UV 270–400 nm | Increased minimal erythema dose and decreased PGE2 | Rhodes et al. [52] |
| Fish oils from mackerel, sardine, and horse mackerel | Human | Broadband UVB, doubled MED | Inhibited UVB-induced erythema | Puglia et al., [53] |
| Ethyl esters of EPA and oleic acid | Human | UV 270–400 nm | Increased minimal erythema dose and decreased PGE2 | Shahbakhti et al. [54] |
| EPA | Human | UV 270–400 nm, 4 × MED | No function on Langerhans cell migration and PGD2 expression | Pilkington et al. [56] |
| EPA | Human | UV 285–350 nm | Decreased epidermal thickness, procollagen, COX-2, and MMPs | Kim et al. [57] |
| LA and ALA | Hairless mouse | UVB at 312.5 nm, 3.6 × MED | Lowered erythema score and PGE2 in skin | Takemura et al. [58] |
| Conjugated LA | Keratinocytes | UV 270–400 nm, 25–100 mJ/cm2 | Reduced IL-8 and PGE2 | Storey et al. [61] |
| Composition | Experimental Model | Tumor-Induced Approach | Benefit | Reference |
|---|---|---|---|---|
| DHA and EPA | Keratinocytes HaCaT | Growth factors in 3% FBS | Induced pre-malignant keratinocyte apoptosis | Nikolakopoulou et al. [66] |
| DHA | Melanoma A-375 | Standard culture medium | Synergistic growth inhibition combined with NSAIDs | Chiu et al. [67] |
| Fish oil, DHA, and EPA | BCC TE 354 and SCC A431 | Standard culture medium | Synergistic growth inhibition combined with imiquimod | Rehman et al. [70] |
| Fish oil | Swiss albino mouse | DMBA-induced papilloma | Reduced tumor size and enhanced IL-10 | Rehman and Zulfakar [71] |
| Fish oil | Swiss albino mouse | Benzo(a)pyrene and croton oil | Reduced papilloma number per mouse | Ramesh and Das [74] |
| Fish oil | Hairless mouse | UV 280–320 nm, 30 mJ/cm2 | Reduced size of papilloma, keratoacanthoma, and carcinoma | Lou et al. [75] |
| Omega-3 and omega-6 fatty acids | Human | SCC patients | Lower SCC risk with the higher omega-3/omega-6 intake | Hakim et al. [76] |
| Fish oil rich in omega-3 PUFAs | Human | Melanoma patients | Lower melanoma risk with the fish oil intake | Fortes et al. [77] |
| DNA-paclitaxel conjugate | Human | Melanoma patients | Prolonged median survival period | Homsi et al. [78] |
| Conjugated LA | Mouse | DMBA- and TPA-induced tumor | Reduced PPAR-δ and K-FABP | Belury et al. [81] |
| Composition | Experimental Model | Dermatitis-Induced Approach | Benefit | Reference |
|---|---|---|---|---|
| Fish oil | Rat | Acetone-induced dry skin | Increased skin hydration and scratching | Barcelos et al. [84] |
| Omega-3 PUFAs | Rat | Trimellitic anhydride-induced allergy | Decreased ear thickness, cutaneous eosinophils, and mast cells | Abdel Latif et al. [87] |
| DHA and AA | Mouse | Ovalbumin-induced dermatitis | Reduced Ki67 and elevated IL-10 expression | Weise et al. [88] |
| GLA | Human | Mild atopic dermatitis | Reduced TEWL and improved stratum corneum index | Kawamura et al. [91] |
| Dihomo-GLA | NC/Nga mouse | Dermatitis-like skin lesion | Suppressed clinical severity score and scratching behavior | Kawashima et al. [93] |
| Dihomo-GLA | NC/Tnd mouse | Dermatitis-like skin lesion | Upregulated PGD1 and reduced scratching behavior | Amagai et al. [94] |
| Composition | Experimental Model | Wound-Induced Approach | Benefit | Reference |
|---|---|---|---|---|
| Fish oil | Pig | Full-thickness skin excision | Fast wound closure at day 2 | Shingel et al. [99] |
| Fish oil | Rat | Full-thickness skin excision | Accelerated healing process and increased IL-10 | Peng et al. [101] |
| DHA | Rat | Full-thickness skin excision | Accelerated healing process and increased GPR120 and TGF-β | Arantes et al. [102] |
| 14R,21-dihydroxy-DHA and 14S,21-dihydroxy-DHA | Mouse | Full-thickness skin excision | Increased granulation tissue region (>65%) and reduced epithelial gap | Lu et al. [103] |
| DHA and EPA | Human | Blisters in the forearms | Increased IL-1β expression in the wound sites | McDaniel et al. [104] |
| ALA, LA, and oleic acid | Mouse | Full-thickness skin excision | Faster wound closure by oleic acid than ALA and LA | Cardoso et al. [105] |
| LA and oleic acid | Rat | Full-thickness skin excision | Reduced necrotic cell layer thickness | Pereira et al. [106] |
| LA | Rat | Streptozotocin- induced diabetic wound | Increased leukocyte accumulation and angiogenesis | Rodrigues et al. [107] |
| Conjugated LA | Human | Fractional laser ablation | Reduced edema and itching | Wu and Goldman [108] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, T.-H.; Wang, P.-W.; Yang, S.-C.; Chou, W.-L.; Fang, J.-Y. Cosmetic and Therapeutic Applications of Fish Oil’s Fatty Acids on the Skin. Mar. Drugs 2018, 16, 256. https://doi.org/10.3390/md16080256
Huang T-H, Wang P-W, Yang S-C, Chou W-L, Fang J-Y. Cosmetic and Therapeutic Applications of Fish Oil’s Fatty Acids on the Skin. Marine Drugs. 2018; 16(8):256. https://doi.org/10.3390/md16080256
Chicago/Turabian StyleHuang, Tse-Hung, Pei-Wen Wang, Shih-Chun Yang, Wei-Ling Chou, and Jia-You Fang. 2018. "Cosmetic and Therapeutic Applications of Fish Oil’s Fatty Acids on the Skin" Marine Drugs 16, no. 8: 256. https://doi.org/10.3390/md16080256
APA StyleHuang, T.-H., Wang, P.-W., Yang, S.-C., Chou, W.-L., & Fang, J.-Y. (2018). Cosmetic and Therapeutic Applications of Fish Oil’s Fatty Acids on the Skin. Marine Drugs, 16(8), 256. https://doi.org/10.3390/md16080256