Functional Comparison of High and Low Molecular Weight Chitosan on Lipid Metabolism and Signals in High-Fat Diet-Fed Rats



Abstract
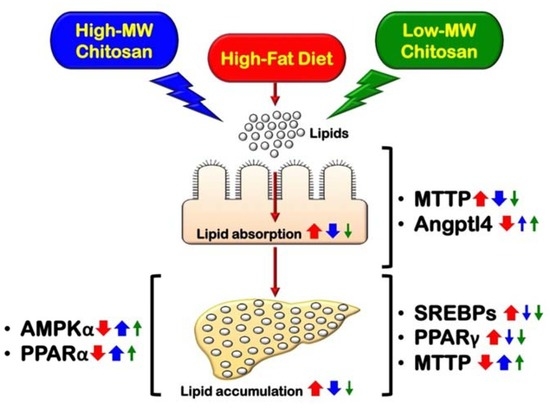
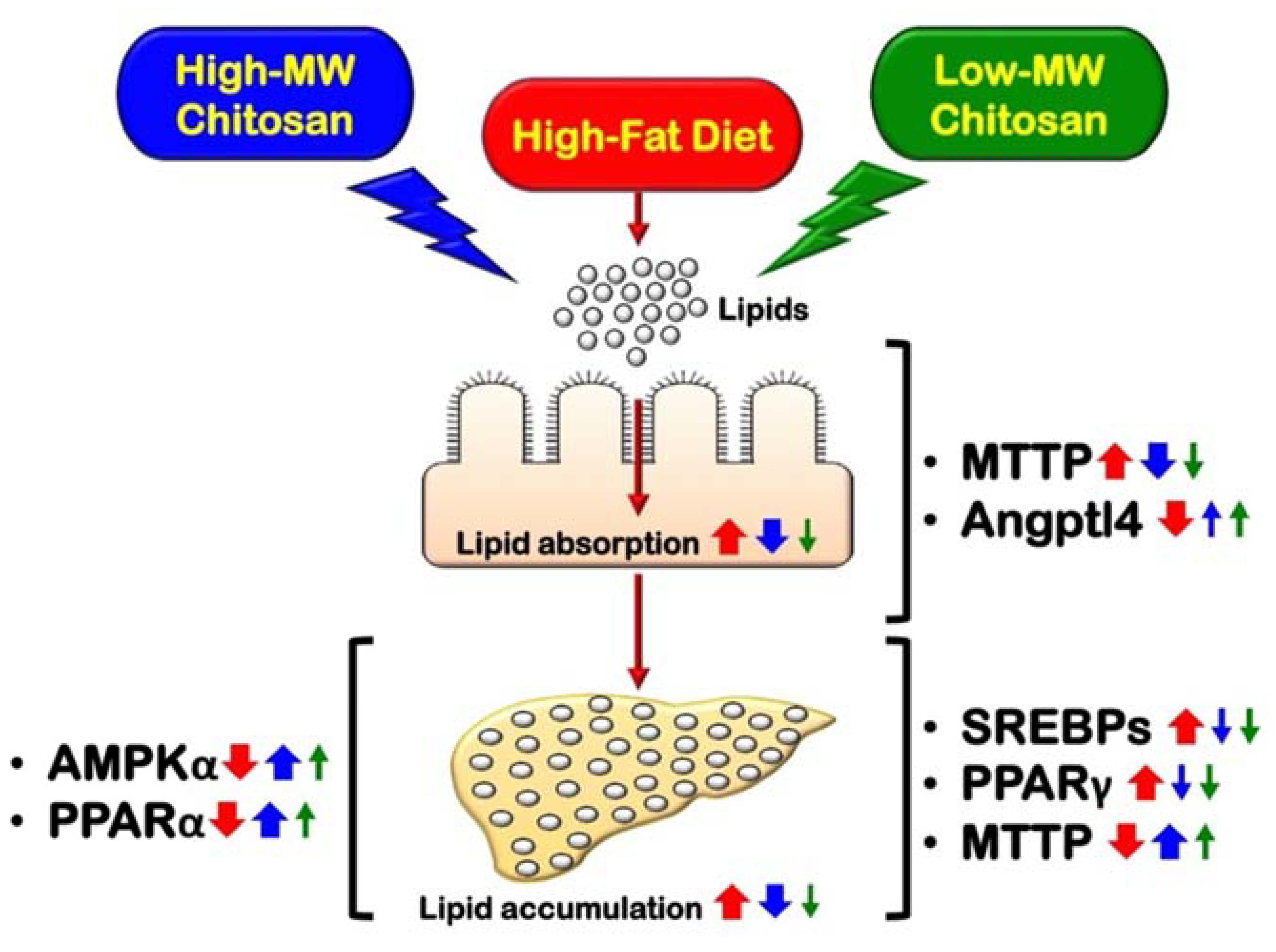
1. Introduction
2. Results
2.1. Effects of Low- and High-Mw Chitosan on Plasma Biochemical Indices, Organ Weight, and Body Weight in Hf Diet-Fed Rats
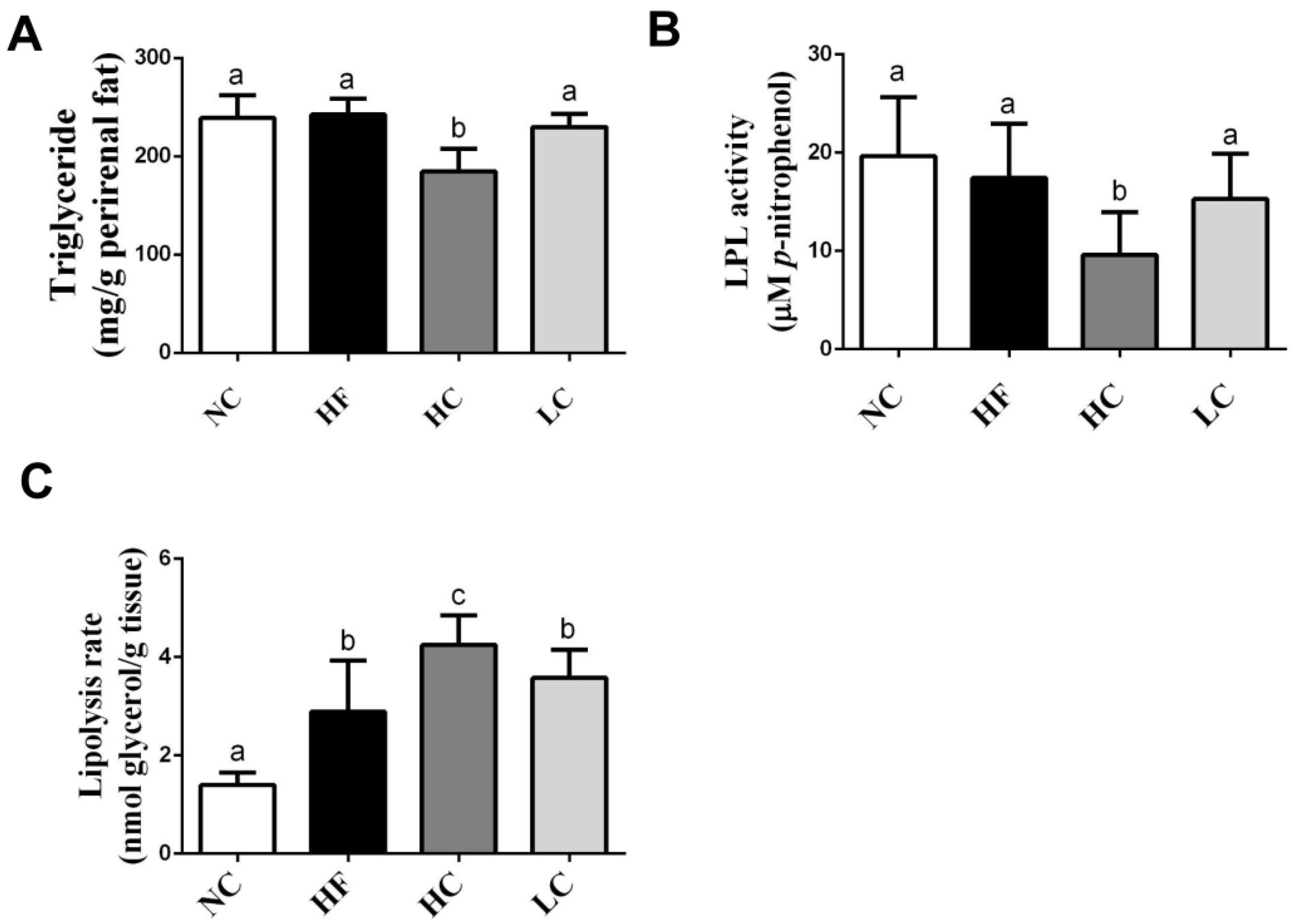
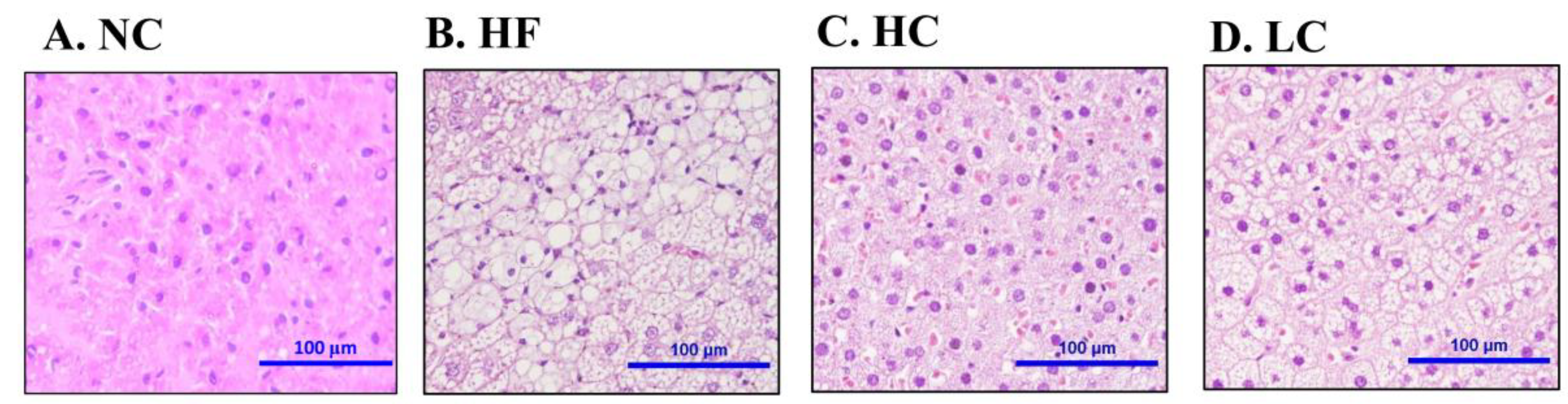
2.2. Effects of Low- and High-Mw Chitosan on The Lipometabolism in Adipose and Liver Tissues and Feces of Hf Diet-Fed Rats
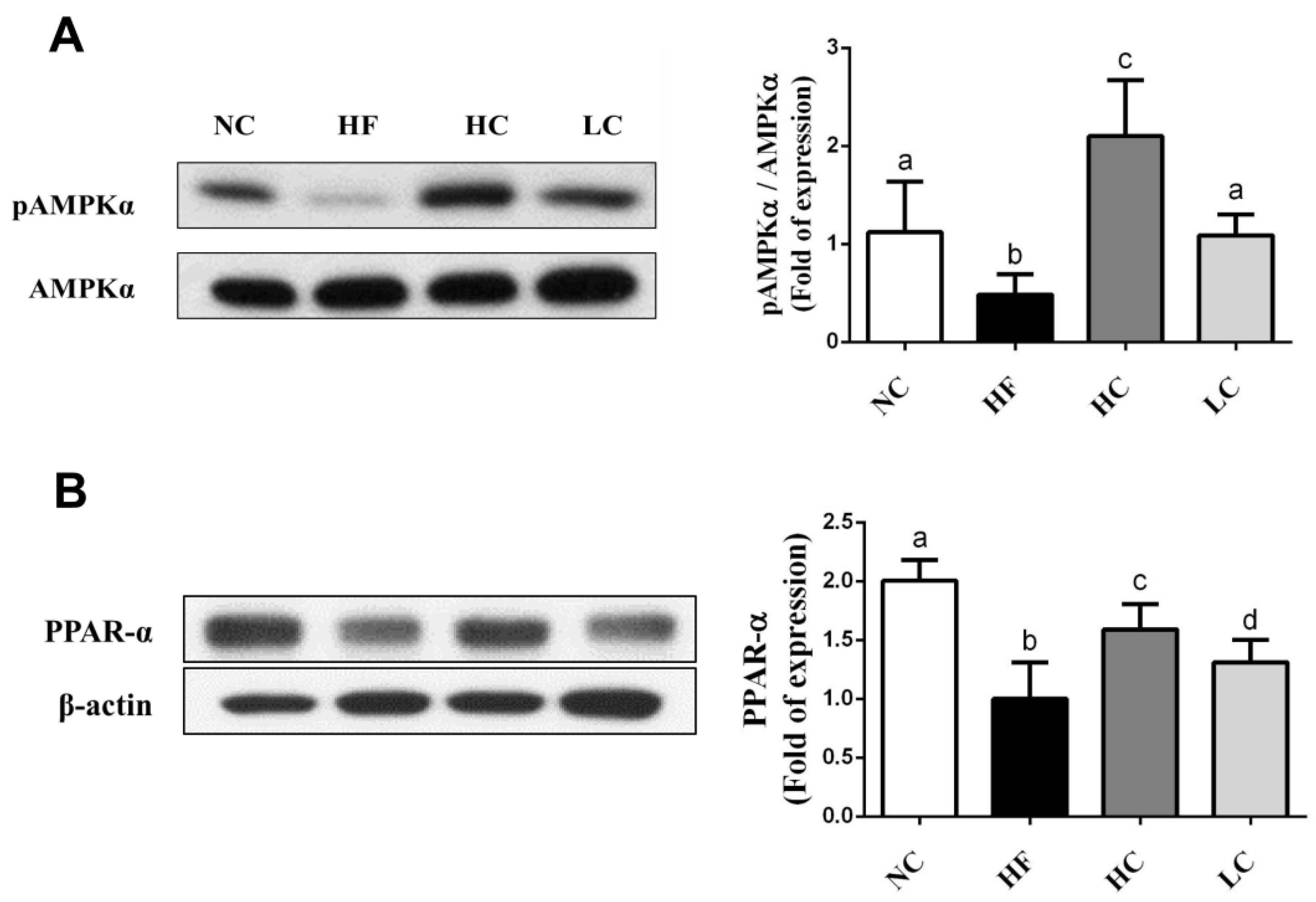
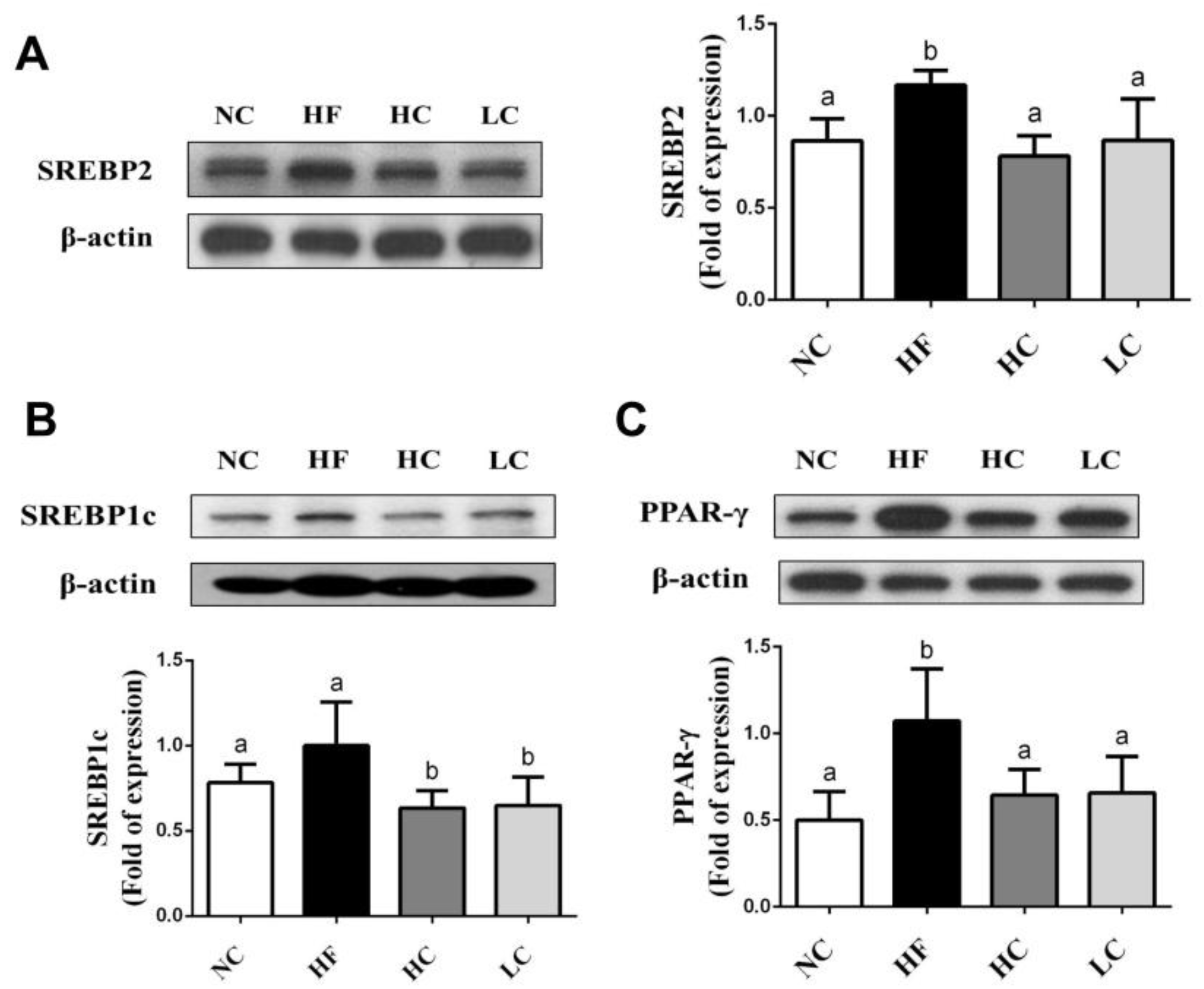
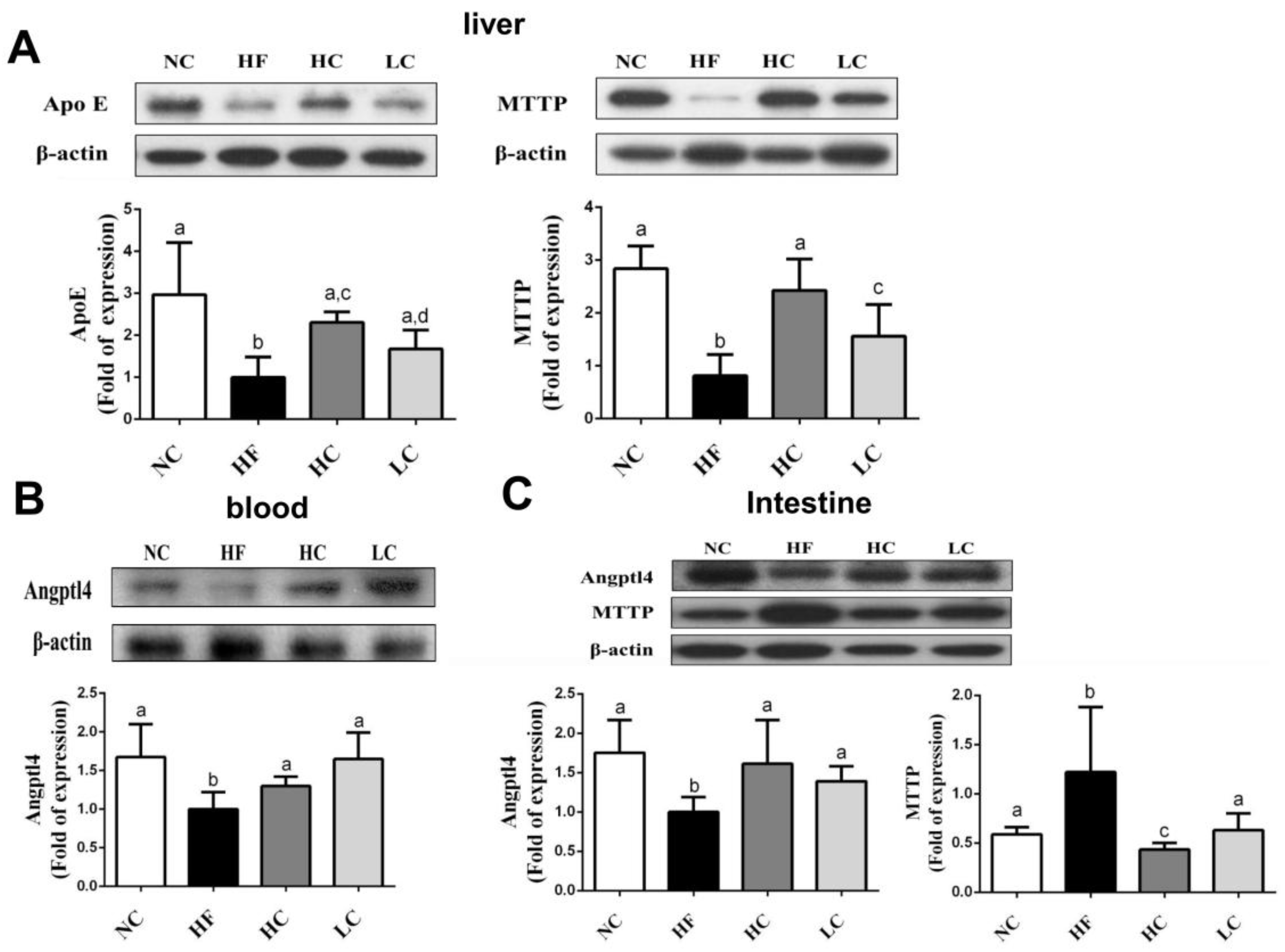
2.3. Effects of Low- and High-Mw Chitosan on Lipometabolic Signals in the Liver, Blood, and Intestine of Hf Diet-Fed Rats
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals and Diets
4.3. Measurement of Triglyceride (TG), Cholesterol (TC), Lipoproteins, and Activities of Aspartate Aminotransferase (AST) and Alanine Aminotransferase (ALT)
4.4. Measurement of Lipolysis Rate
4.5. Measurement of Lipoprotein Lipase (LPL) Activity
4.6. Histological Analysis of Liver
4.7. Protein Expression Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhu, S.; St-Onge, M.P.; Heshka, S.; Heymsfield, S.B. Lifestyle behaviors associated with lower risk of having the metabolic syndrome. Metabolism 2004, 53, 1503–1511. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Obesity and overweight. 2017. Available online: http://www.who.int/gho/ncd/risk_factors/overweight/en/ (accessed on 20 June 2018).
- Brunt, E.M. Nonalcoholic steatohepatitis: Definition and pathology. Semin. Liver Dis. 2001, 21, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Petrović, G.; Bjelaković, G.; Benedeto-Stojanov, D.; Nagorni, A.; Brzački, V.; Marković-Živković, B. Obesity and metabolic syndrome as risk factors for the development of non-alcoholic fatty liver disease as diagnosed by ultrasound. Vojnosanit. Pregl. 2016, 73, 910–920. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Chang, T.C.; Liu, S.H.; Chiang, M.T. The regulatory effects of fish oil and chitosan on hepatic lipogenic signals in high-fat diet-induced obese rats. J. Food Drug Anal. 2017, 25, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.L.; Yao, H.T.; Cheng, R.S.; Chiang, M.T. Chitosan reduces plasma adipocytokines and lipid accumulation in liver and adipose tissues and ameliorates insulin resistance in diabetic rats. J. Med. Food 2012, 15, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Chang, Y.H.; Chiang, M.T. Chitosan reduces gluconeogenesis and increases glucose uptake in skeletal muscle in streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2010, 58, 5795–5800. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Cai, F.Y.; Chiang, M.T. Long-term feeding of chitosan ameliorates glucose and lipid metabolism in a high-fructose-diet-impaired rat model of glucose tolerance. Mar. Drugs 2015, 13, 7302–7313. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Qin, C.; Wang, W.; Chi, W.; Li, W. Absorption and distribution of chitosan in mice after oral administration. Carbohydr. Polym. 2008, 71, 435–440. [Google Scholar] [CrossRef]
- Kondo, Y.; Nakatani, A.; Hayashi, K.; Ito, M. Low molecular weight chitosan prevents the progression of low dose streptozotocin-induced slowly progressive diabetes mellitus in mice. Biol. Pharm. Bull. 2000, 23, 1458–1464. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.T.; Huang, S.Y.; Chiang, M.T. A comparative study on hypoglycemic and hypocholesterolemic effects of high and low molecular weight chitosan in streptozotocin-induced diabetic rats. Food Chem. Toxicol. 2008, 46, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Lee, M.S.; Jo, K.; Lee, K.E.; Hwang, J.K. Therapeutic potential of panduratin A, LKB1-dependent AMP-activated protein kinase stimulator, with activation of PPARalpha/delta for the treatment of obesity. Diabetes Obes. Metab. 2011, 13, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Oike, Y.; Akao, M.; Kubota, Y.; Suda, T. Angiopoietin-like proteins: Potential new targets for metabolic syndrome therapy. Trends Mol. Med. 2005, 11, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Sacks, F.M. The crucial roles of apolipoproteins E and C-III in apoB lipoprotein metabolism in normolipidemia and hypertriglyceridemia. Curr. Opin. Lipidol. 2015, 26, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Shindo, N.; Fujisawa, T.; Sugimoto, K.; Nojima, K.; Oze-Fukai, A.; Yoshikawa, Y.; Wang, X.; Yasuda, O.; Ikegami, H.; Rakugi, H. Involvement of microsomal triglyceride transfer protein in nonalcoholic steatohepatitis in novel spontaneous mouse model. J. Hepatol. 2010, 52, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Sugano, M.; Fujikawa, Y.; Hiratsuji, K.; Nakashima, N.; Hasegawa, Y.A. novel use of chitosan as a hypocholesterolemic agent in rats. Am. J. Clin. Nutr. 1980, 33, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Gades, M.D.; Stern, J.S. Chitosan supplementation and fecal fat excretion in men. Obes. Res. 2003, 11, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Sugano, M.; Watanabe, S.; Kishi, A.; Izume, M.; Ohtakara, A. Hypocholesterolemic action of chitosans with different viscosity in rats. Lipids 1988, 23, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, W.; Mamadouba, B.; Xia, W. A comparative study on hypolipidemic activities of high and low molecular weight chitosan in rats. Int. J. Biol. Macromol. 2012, 51, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Yang, Q.; Huang, G.; Ding, C.; Cao, P.; Huang, L.; Xiao, T.; Guo, J.; Su, Z. Hypolipidemic effects of chitosan and its derivatives in hyperlipidemic rats induced by a high-fat diet. Food Nutr. Res. 2016, 60, 31137. [Google Scholar] [CrossRef] [PubMed]
- Gaidhu, M.P.; Anthony, N.M.; Patel, P.; Hawke, T.J.; Ceddia, R.B. Dysregulation of lipolysis and lipid metabolism in visceral and subcutaneous adipocytes by high-fat diet: Role of ATGL, HSL, and AMPK. J. Physiol. 2010, 298, C961–C971. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Dai, K.; Seimon, T.; Jungreis, R.; Oyadomari, M.; Kuriakose, G.; Ron, D.; Tabas, I.; Hussain, M.M. IRE1beta inhibits chylomicron production by selectively degrading MTP mRNA. Cell Metab. 2008, 7, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Mattijssen, F.; Alex, S.; Swarts, H.J.; Groen, A.K.; Schothorst, E.M.; Kersten, S. Angptl4 serves as an endogenous inhibitor of intestinal lipid digestion. Mol. Metab. 2013, 3, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, G.R.; Kemp, B.E. AMPK in Health and Disease. Physiol. Rev. 2009, 89, 1025–1078. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.K.; Marcinko, K.; Desjardins, E.M.; Lally, J.S.; Ford, R.J.; Steinberg, G.R. Treatment of nonalcoholic fatty liver disease: Role of AMPK. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E730–E740. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Bronner, M.; Hertz, R.; Bar-Tana, J. Kinase-independent transcriptional co-activation of peroxisome proliferator-activated receptor alpha by AMP-activated protein kinase. Biochem. J. 2004, 384, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Misra, P.; Reddy, J.K. Peroxisome proliferator-activated receptor-alpha activation and excess energy burning in hepatocarcinogenesis. Biochimie 2014, 98, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Jia, Y.; Yang, G.; Zhang, X.; Boddu, P.C.; Petersen, B.; Narsingam, S.; Zhu, Y.J. PPARα-deficient ob/ob obese mice become more obese and manifest severe hepatic steatosis due to decreased fatty acid oxidation. J. Pathol. 2015, 185, 1396–1408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, Z.X.; Choi, C.S.; Tian, L.; Kibbey, R.; Dong, J.; Cline, G.W.; Wood, P.A.; Shulman, G.I. Mitochondrial dysfunction due to long-chain Acyl-CoA dehydrogenase deficiency causes hepatic steatosis and hepatic insulin resistance. Proc. Natl. Acad. Sci. USA 2007, 104, 17075–17080. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, F.; Jong, M.C.; Lin, Y.; Eck, M.; Havinga, R.; Bloks, V.; Verkade, H.J.; Hofker, M.H.; Moshage, H.; Berkel, T.J.; Vonk, R.J.; Havekes, L.M. Impaired secretion of very low density lipoproteintriglycerides by apolipoprotein E-deficient mouse hepatocytes. J. Clin. Investig. 1997, 100, 2915–2922. [Google Scholar] [CrossRef] [PubMed]
- Maugeais, C.; Tietge, U.J.; Tsukamoto, K.; Glick, J.M.; Rader, D.J. Hepatic apolipoprotein E expression promotoes very low density lipoprotein-apolipoprotein B production in vivo in mice. J. Lipid Res. 2000, 41, 1673–1679. [Google Scholar] [PubMed]
- Tanoli, T.; Yue, P.; Yablonskiy, D.; Schonfeld, G. Fatty liver in familial hypobetalipoproteinemia: Roles of the APOB defects, intra-abdominal adipose tissue, and insulin sensitivity. J. Lipid Res. 2004, 45, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Newberry, E.P.; Norris, J.Y.; Xie, Y.; Luo, J.; Kennedy, S.M.; Davidson, N.O. ApoB100 is required for increased VLDL-triglyceride secretion by microsomal triglyceride transfer protein in ob/ob mice. J. Lipid Res. 2008, 49, 2013–2022. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.M.; Nijstad, N.; Franceschini, L. Regulation of microsomal triglyceride transfer protein. Clin. Lipidol. 2011, 6, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Koo, S.H. Nonalcoholic fatty liver disease: Molecular mechanisms for the hepatic steatosis. Clin. Mol. Hepatol. 2013, 19, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Guo, Z.; Johnson, C.M.; Hensrud, D.D.; Jensen, M.D. Splanchnic lipolysis in human obesity. J. Clin. Invest. 2004, 113, 1582–1588. [Google Scholar] [CrossRef] [PubMed]
- Fabbrini, E.; Mohammed, B.S.; Magkos, F.; Korenblat, K.M.; Patterson, B.W.; Klein, S. Alterations in adipose tissue and hepatic lipid kinetics in obese men and women with nonalcoholic fatty liver disease. Gastroenterology 2008, 134, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Biddinger, S.B.; Almind, K.; Miyazaki, M.; Kokkotou, E.; Ntambi, J.M.; Kahn, C.R. Effects of diet and genetic background on sterol regulatory element-binding protein-1c, stearoyl-CoA desaturase 1, and the development of the metabolic syndrome. Diabetes 2005, 54, 1314–1323. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Jou, W.; Gavrilova, O.; Hall, K.D. Persistent diet induced obesity in male C57BL/6 mice resulting from temporary obesigenic diets. PLoS ONE 2009, 4, e5370. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Chan, I.L.; Yang, T.H.; Liu, S.H.; Chiang, M.T. Supplementation of chitosan alleviates high-fat diet-enhanced lipogenesis in rats via adenosine monophosphate (AMP)-activated protein kinase activation and inhibition of lipogenesis-associated genes. J. Agric. Food Chem. 2015, 63, 2979–2988. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.P.; Yao, H.T.; Chiang, M.T. Effects of high and low molecular weight chitosan on plasma cholesterol, glucose and adipocytokines in diabetic rats induced by streptozotocin and nicotinamide. J. Food Drug Anal. 2012, 20, 661–667. [Google Scholar]
- National Research Council (NRC). Guide for the Care and Use of Laboratory Animals. 2011. Available online: https://grants.nih.gov/grants/olaw/guide-for-the-care-and-use-of-laboratory-animals.pdf (accessed on 20 June 2018).
- Chiu, C.Y.; Wang, L.P.; Liu, S.H.; Chiang, M.T. Fish oil supplementation alleviates the altered lipid homeostasis in blood, liver, and adipose tissues in high-fat diet-fed rats. J. Agric. Food Chem. 2018, 66, 4118–4128. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet | NC | HF | HC | LC |
|---|---|---|---|---|
| Food intake (g) | 27.9 ± 1.7 a | 25.3 ± 1.9 a | 26.7 ± 1.3 a | 25.9 ± 1.5 a |
| Caloric intake (kcal/kg/day) | 107.0± 6.4 a | 109.3 ± 8.3 a | 115.0 ± 5.8 a | 111.9 ± 6.5 a |
| Body weight (g) | 539.9 ± 22.5 a | 578 ± 47.3 b | 518.3 ± 34.1 a | 543.4 ± 18.9 a |
| Liver weight (g) | 14.5 ± 1.5 a | 33.3 ± 4.9 b | 20.0 ± 1.9 c | 23.0 ± 3.2 d |
| Relative liver weight (g/100 g BW) | 2.7 ± 0.2 a | 5.9 ± 0.7 b | 3.9 ± 0.4 a | 4.2 ± 0.5 a |
| Adipose tissue weight (g) | 28.7 ± 6.3 a,b | 23.1 ± 5.2 b | 18.0 ± 5.6 b,c | 20.7 ± 3.1 b,c |
| Relative adipose weight (g/100 g BW) | 5.2 ± 1.0 a | 4.1 ± 0.8 b | 3.5 ± 1.0 b | 3.8 ± 0.5 b |
| Perirenal adipose weight (g) | 16.7 ± 3.6 a | 14.0 ± 3.4 a | 10.7 ± 2.7 b | 12.1 ± 2.2 a |
| Epididymal adipose weight (g) | 12.0 ± 3.0 a | 9.1 ± 2.0 b | 7.3 ± 3.3 b | 8.6 ± 1.5 b |
| Small intestine length (cm) | 116.3 ± 2.6 a,b | 116.0 ± 5.2 a | 132.6 ± 11.4 b,c | 129.1 ± 10.3 b,c |
| Relative small intestine length (cm/100 g BW) | 21.4 ± 1.2 a | 20.7 ± 1.6 a | 25.6 ± 1.4 b | 23.5 ± 1.9 c |
| Diet | NC | HF | HC | LC |
|---|---|---|---|---|
| Total cholesterol (mg/dL) | 63.1 ± 9.3 a | 87.4 ± 11.7 b | 56.2 ± 7.9 a | 52.3 ± 17.0 a |
| HDL-C (mg/dL) | 44.7 ± 5.8 a | 9.1 ± 7.9 b | 26.7 ± 2.7 b | 25.5 ± 4.1 b |
| LDL-C + VLDL-C (mg/dL) | 18.4 ± 7.2 a | 58.3 ± 12.0 b | 29.5 ± 10.2 c | 26.7 ± 18.7 a,c |
| VLDL-C (mg/dL) | 14.9 ± 8.1 a | 28.8 ± 8.2 b | 16.0 ± 6.4 a | 14.0 ± 12.7 a |
| LDL-C (mg/dL) | 3.5 ± 1.6 a | 29.5 ± 11.4 b | 13.6 ± 7.7 c | 12.7 ± 10.5 c |
| TC/HDL-C (mg/dL) | 1.4 ± 0.2 a | 3.2 ± 0.9 b | 2.1 ± 0.5 c | 2.1 ± 0.9 a,c |
| HDL-C/LDL-C + VLDL-C ratio | 2.8 ± 1.2 a | 0.5 ± 0.2 b | 1.0 ± 0.4 c | 2.2 ± 2.7 a,b,c |
| Triglyceride (mg/dL) | 96.2 ± 43.3 a | 34.5 ± 5.1 b | 44.1 ± 8.8 c | 45.8 ± 13.0 c |
| TNF-α (pg/dL) | 10.9 ± 2.3 a | 36.8 ± 13.7 b | 17.1 ± 3.6 c | 22.8 ± 7.3 c |
| ALT (U/L) | 15.7 ± 3.5 a | 72.2 ± 32.1 b | 25.5 ± 15.9 a,c | 35.1 ± 25.0 c |
| AST (U/L) | 42.1 ± 16.6 a | 79.1 ± 42.8 b | 39.4 ± 22.2 a | 59.7 ± 31.2 a,b |
| Diet | NC | HF | HC | LC |
|---|---|---|---|---|
| Total cholesterol | - | - | - | - |
| (mg/g liver) | 2.7 ± 1.0 a | 152.6 ± 13.7 b | 52.6 ± 28.4 c | 65.4 ± 33.2 c |
| (g/liver) | 0.04 ± 0.02 a | 5.1 ± 0.8 b | 1.1 ± 0.6 c | 1.6 ± 0.9 c |
| Triglyceride | - | - | - | - |
| (mg/g liver) | 12.1 ± 5.0 a | 96.0 ± 22.2 b | 49.2 ± 20.1 c | 58.7 ± 29.0 c |
| (g/liver) | 0.2 ± 0.1 a | 3.1 ± 0.5 b | 1.0 ± 0.4 c | 1.4 ± 0.8 c |
| Diet | NC | HF | HC | LC |
|---|---|---|---|---|
| Feces wet weight (g/day) | 2.9 ± 0.2 a | 2.8 ± 0.2 a | 3.1 ± 0.5 b | 3.6 ± 0.5 b |
| Feces dry weight (g/day) | 2.2 ± 0.2 a,b | 2.0 ± 0.2 b | 2.4 ± 0.5 a | 2.9 ± 0.5 a,c |
| Total cholesterol | - | - | - | - |
| (mg/g feces) | 2.4 ± 0.4 a | 12.2 ± 1.5 b | 24.2 ± 4.8 c | 23.2 ± 4.7 c |
| (mg/day) | 5.4 ± 1.1 a | 24.3 ± 3.4 b | 59.4 ± 20 c | 66.5 ± 17.3 c |
| Triglyceride | - | - | - | - |
| (mg/g feces) | 0.4 ± 0.2 a | 0.7 ± 0.2 b | 1.8 ± 0.4 c | 2.0 ± 0.3 c |
| (mg/day) | 0.9 ± 0.4 a | 1.4 ± 0.5 b | 4.5 ± 1.5 c | 5.7 ± 1.4 c |
| Ingredient (%) | NC | HF | HC | LC |
|---|---|---|---|---|
| Casein | 20 | 20 | 20 | 20 |
| Lard | 3 | 13 | 13 | 13 |
| Soybean oil | 2 | 2 | 2 | 2 |
| Vitamin mixture 1 | 1 | 1 | 1 | 1 |
| Minerals 2 | 4 | 4 | 4 | 4 |
| Cholesterol | - | 0.5 | 0.5 | 0.5 |
| Choline chloride | 0.2 | 0.2 | 0.2 | 0.2 |
| Cholic acid | - | 0.2 | 0.2 | 0.2 |
| Corn starch | 64.8 | 54.1 | 54.1 | 54.1 |
| Cellulose | 5 | 5 | - | - |
| High molecular weight chitosan 3 | - | - | 5 | - |
| Low molecular weight chitosan 4 | - | - | - | 5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.-H.; Chiu, C.-Y.; Shi, C.-M.; Chiang, M.-T. Functional Comparison of High and Low Molecular Weight Chitosan on Lipid Metabolism and Signals in High-Fat Diet-Fed Rats. Mar. Drugs 2018, 16, 251. https://doi.org/10.3390/md16080251
Liu S-H, Chiu C-Y, Shi C-M, Chiang M-T. Functional Comparison of High and Low Molecular Weight Chitosan on Lipid Metabolism and Signals in High-Fat Diet-Fed Rats. Marine Drugs. 2018; 16(8):251. https://doi.org/10.3390/md16080251
Chicago/Turabian StyleLiu, Shing-Hwa, Chen-Yuan Chiu, Ching-Ming Shi, and Meng-Tsan Chiang. 2018. "Functional Comparison of High and Low Molecular Weight Chitosan on Lipid Metabolism and Signals in High-Fat Diet-Fed Rats" Marine Drugs 16, no. 8: 251. https://doi.org/10.3390/md16080251
APA StyleLiu, S.-H., Chiu, C.-Y., Shi, C.-M., & Chiang, M.-T. (2018). Functional Comparison of High and Low Molecular Weight Chitosan on Lipid Metabolism and Signals in High-Fat Diet-Fed Rats. Marine Drugs, 16(8), 251. https://doi.org/10.3390/md16080251