Multiple Isoforms of Anti-Lipopolysaccharide Factors and Their Antimicrobial Functions in the Ridgetail Prawn Exopalaemon carinicauda

Abstract
:1. Introduction
2. Results
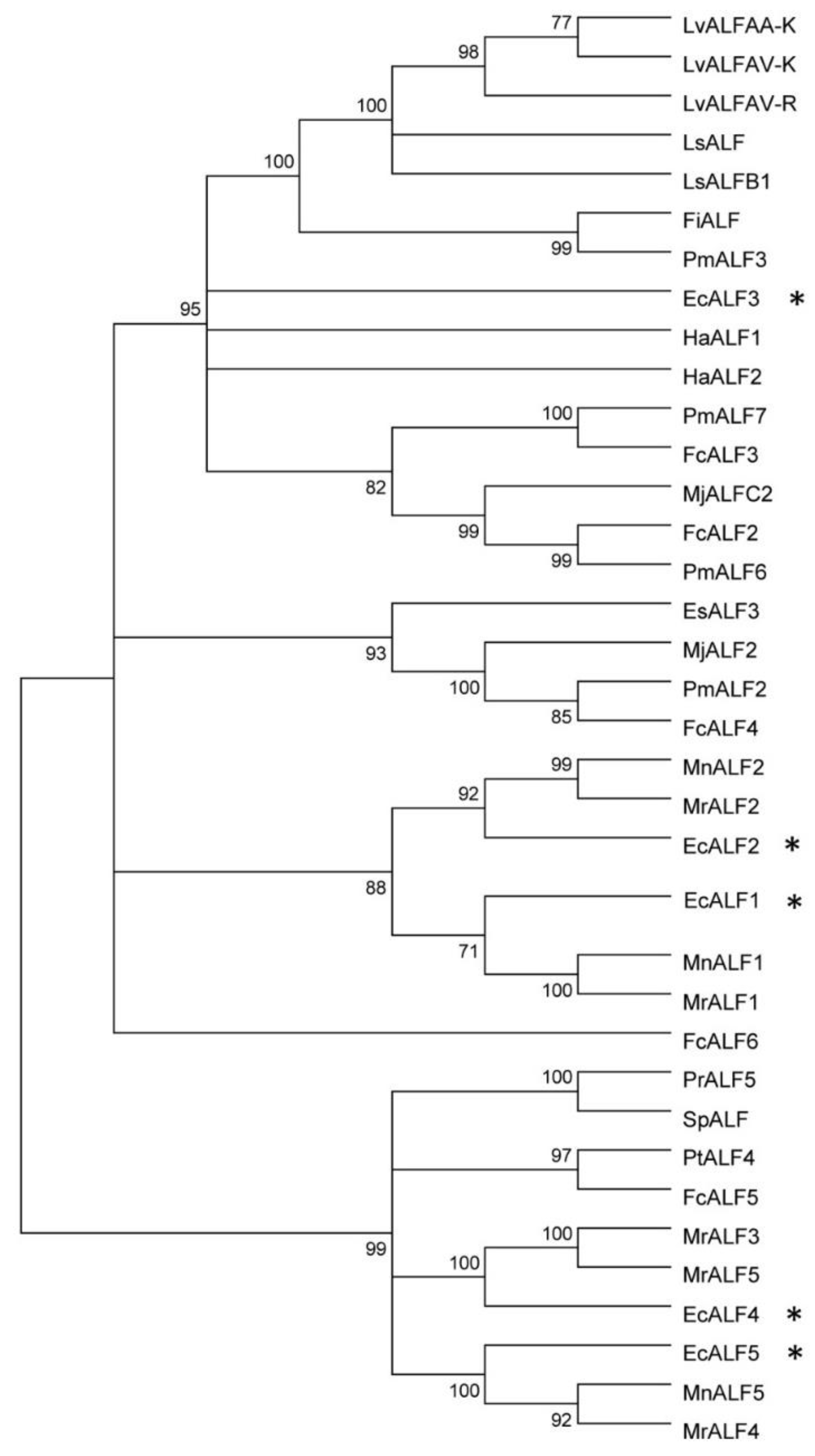
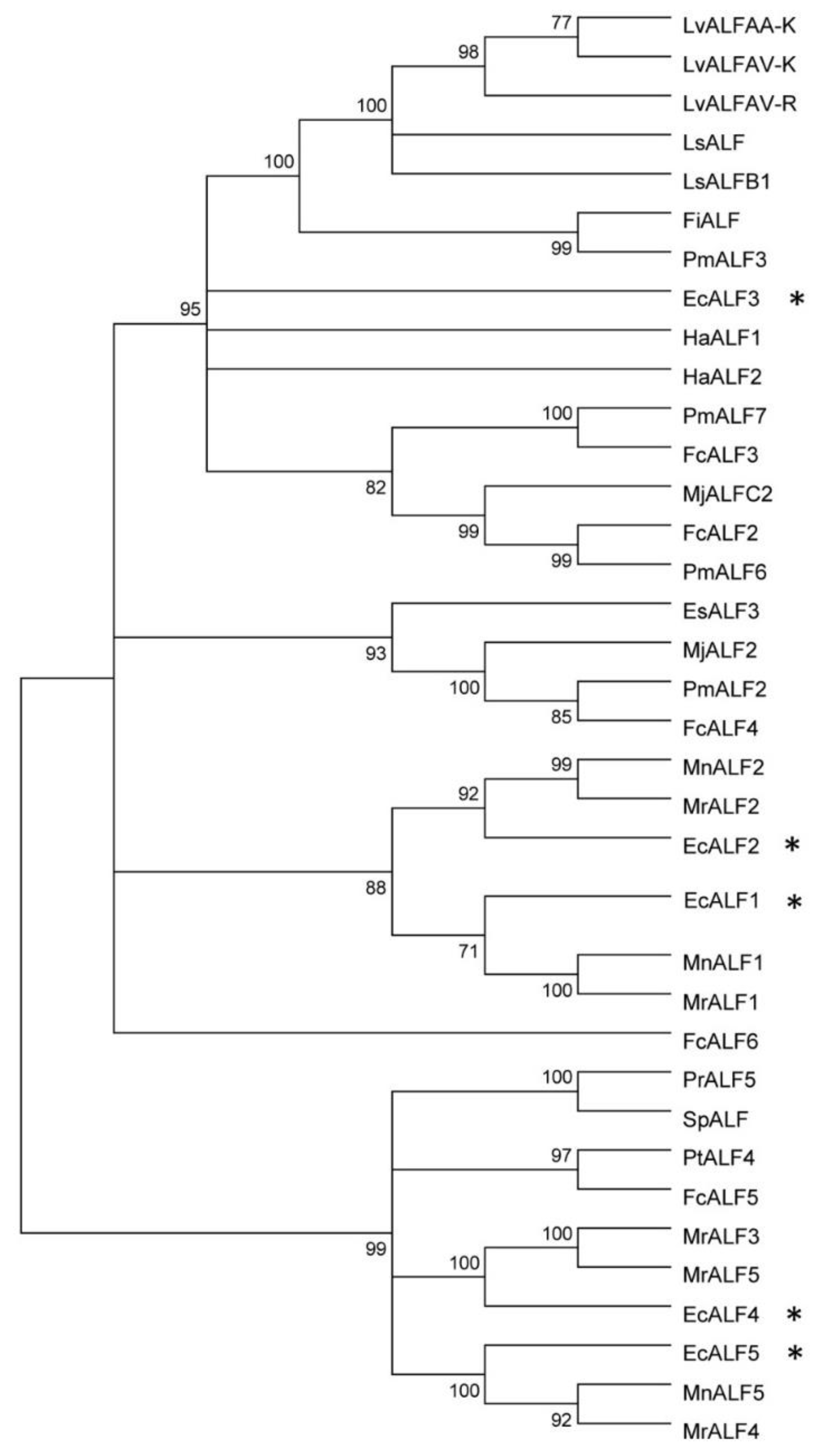
2.1. Sequences Characters and Phylogenic Analysis of EcALFs
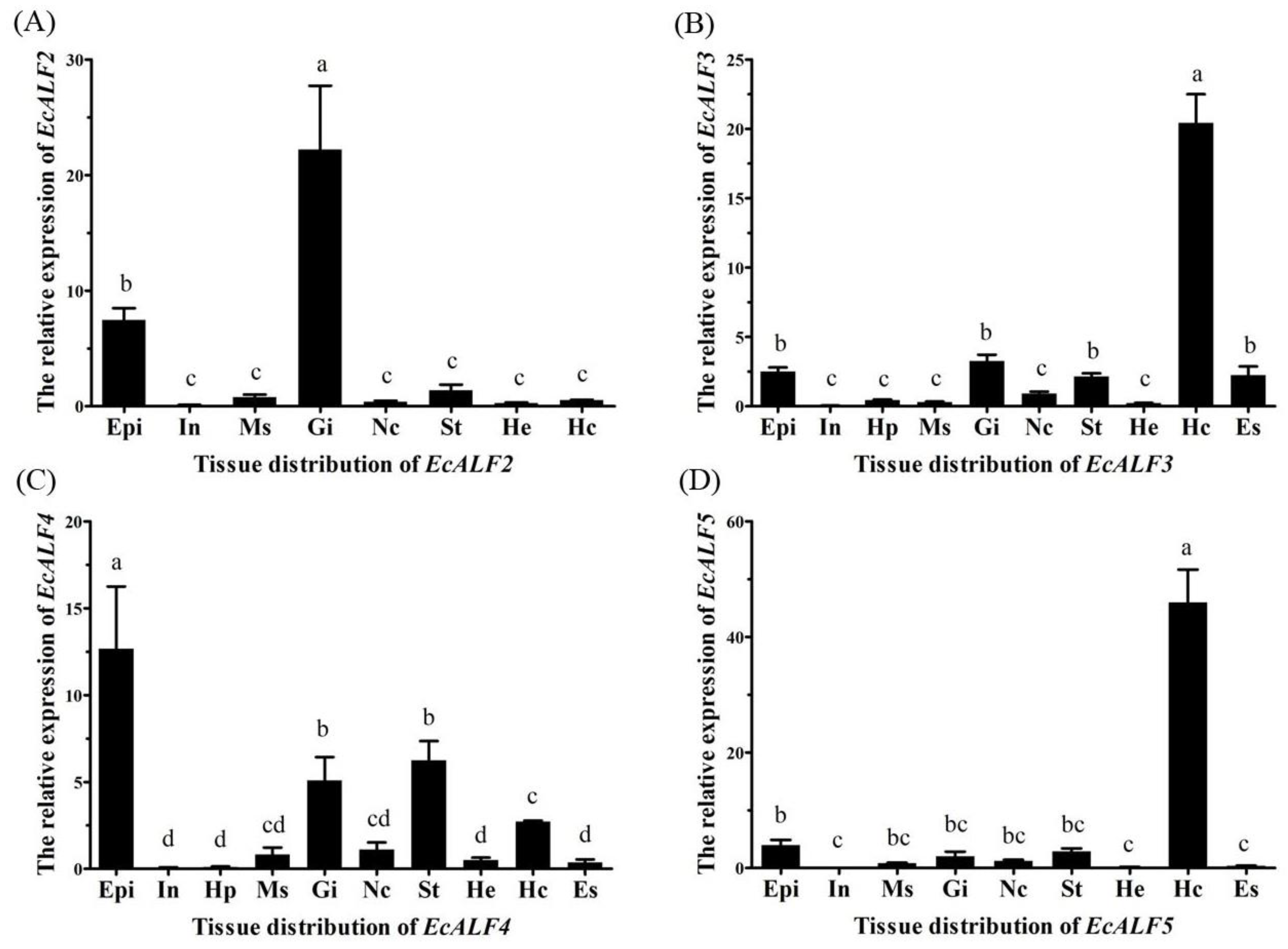
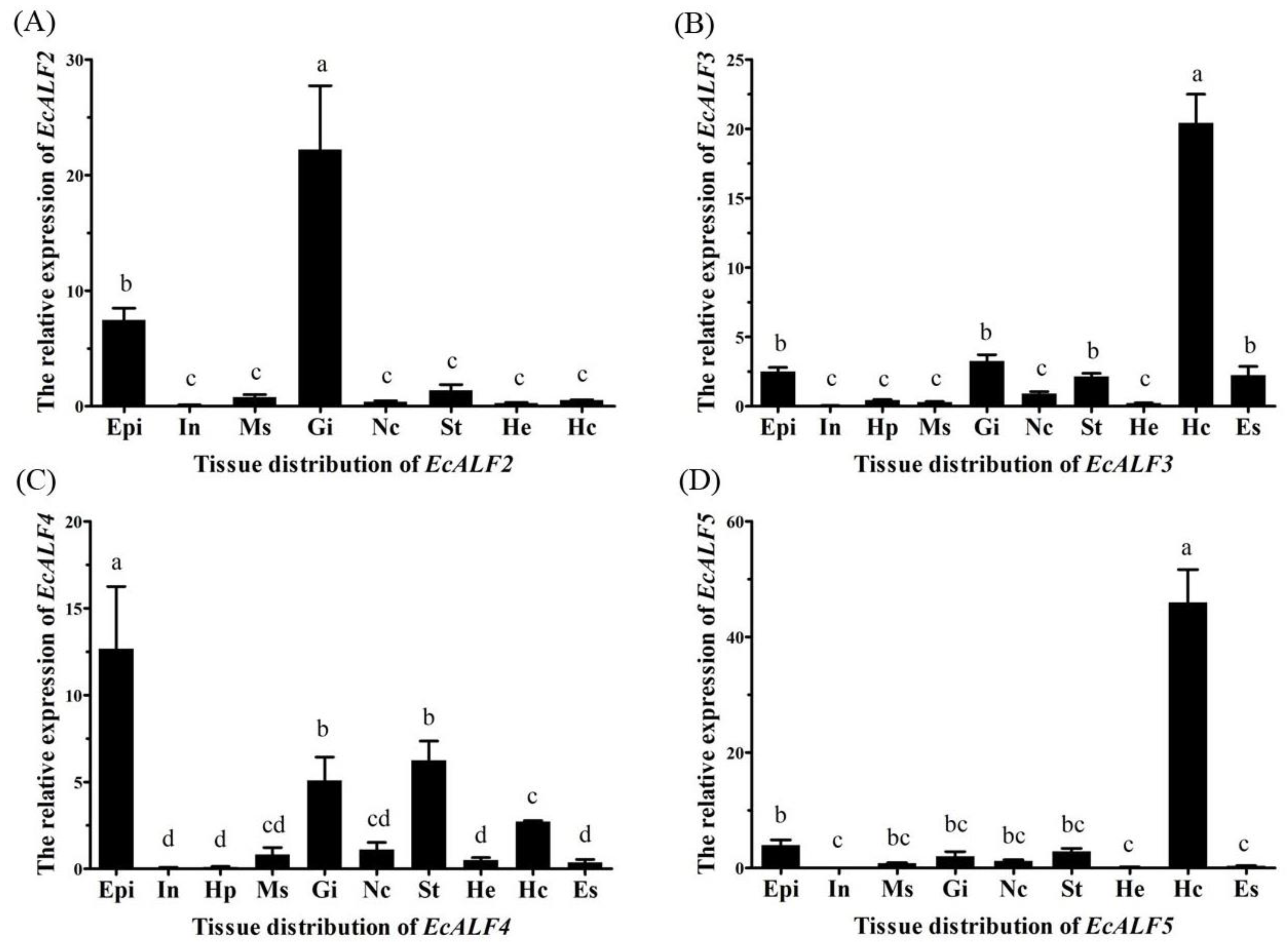
2.2. Tissue Distribution of Different EcALFs Transcripts
2.3. Minimal Inhibitory Concentrations (MIC) of the Synthetic EcLBD Peptides
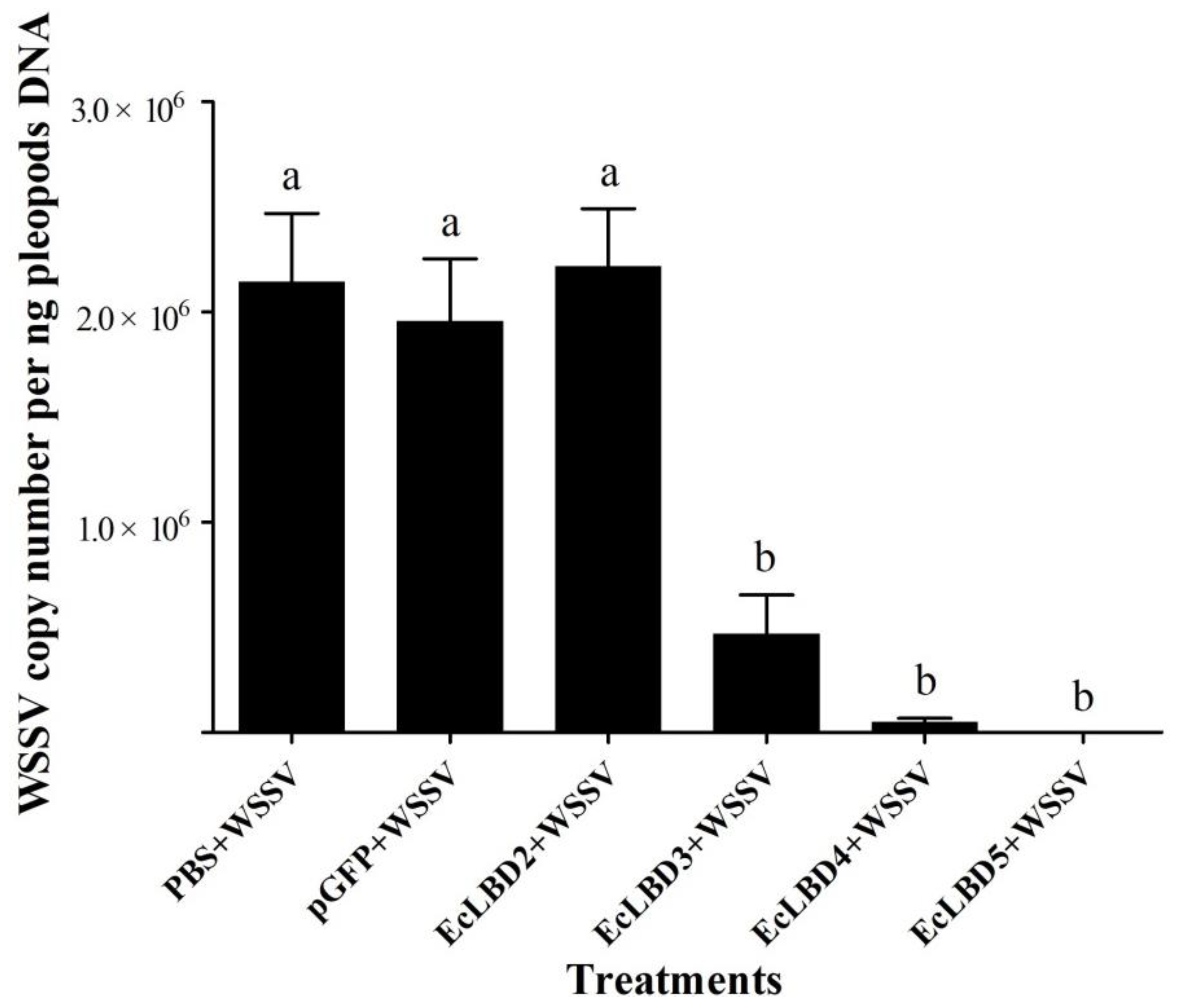
2.4. Antiviral Activity of EcLBD Peptides
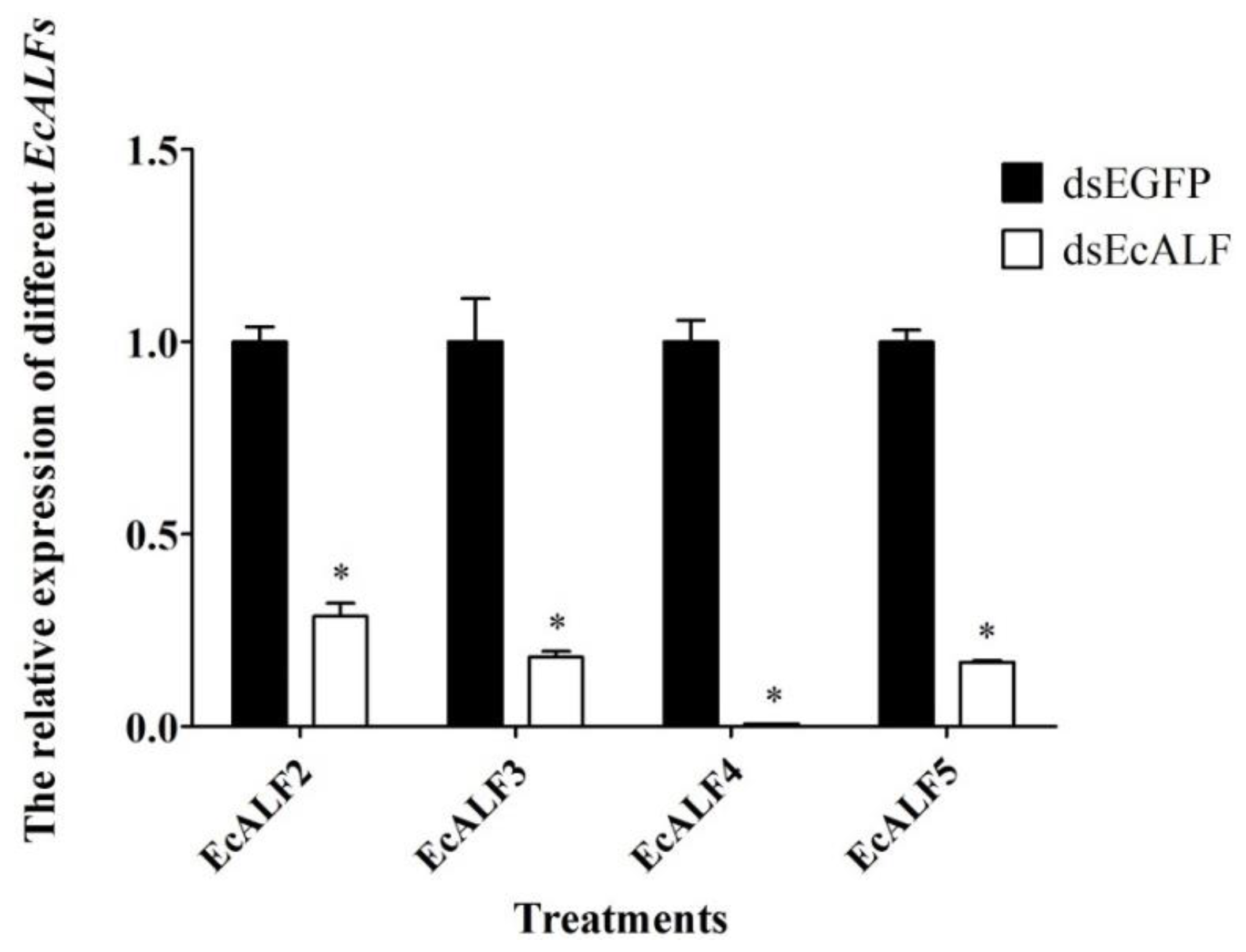
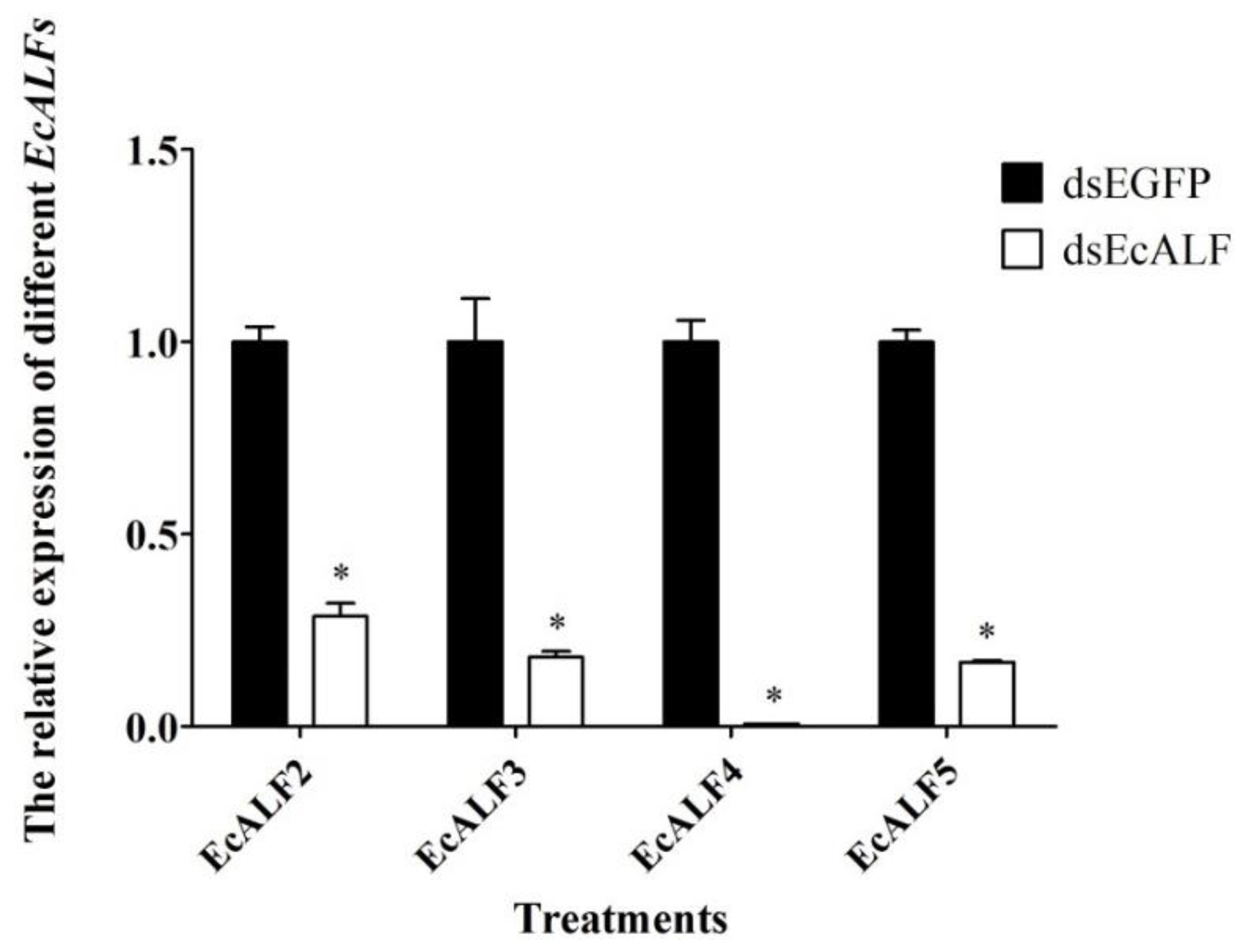
2.5. Optimization of dsRNA Dosage for Different EcALFs in E. carinicauda
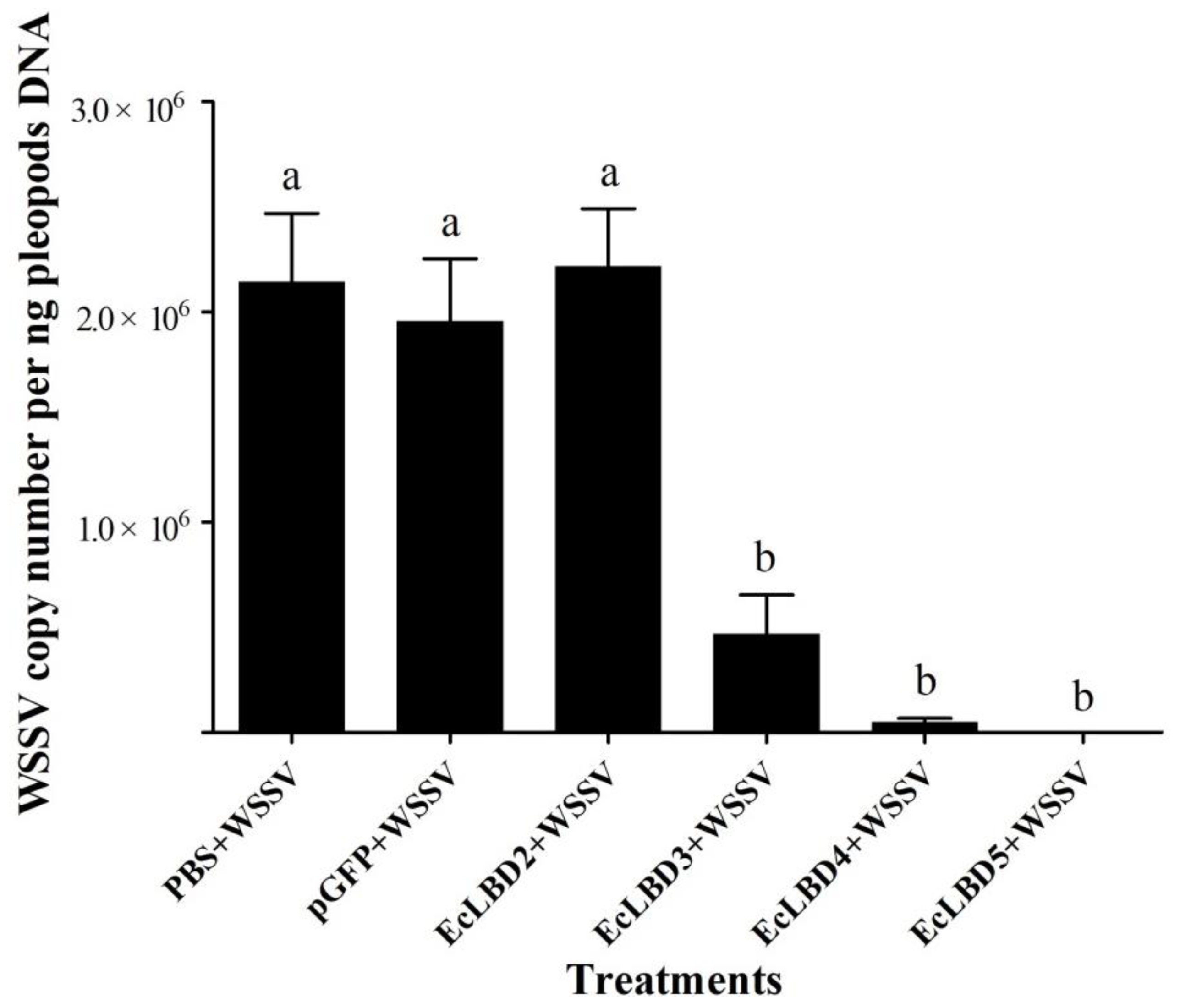
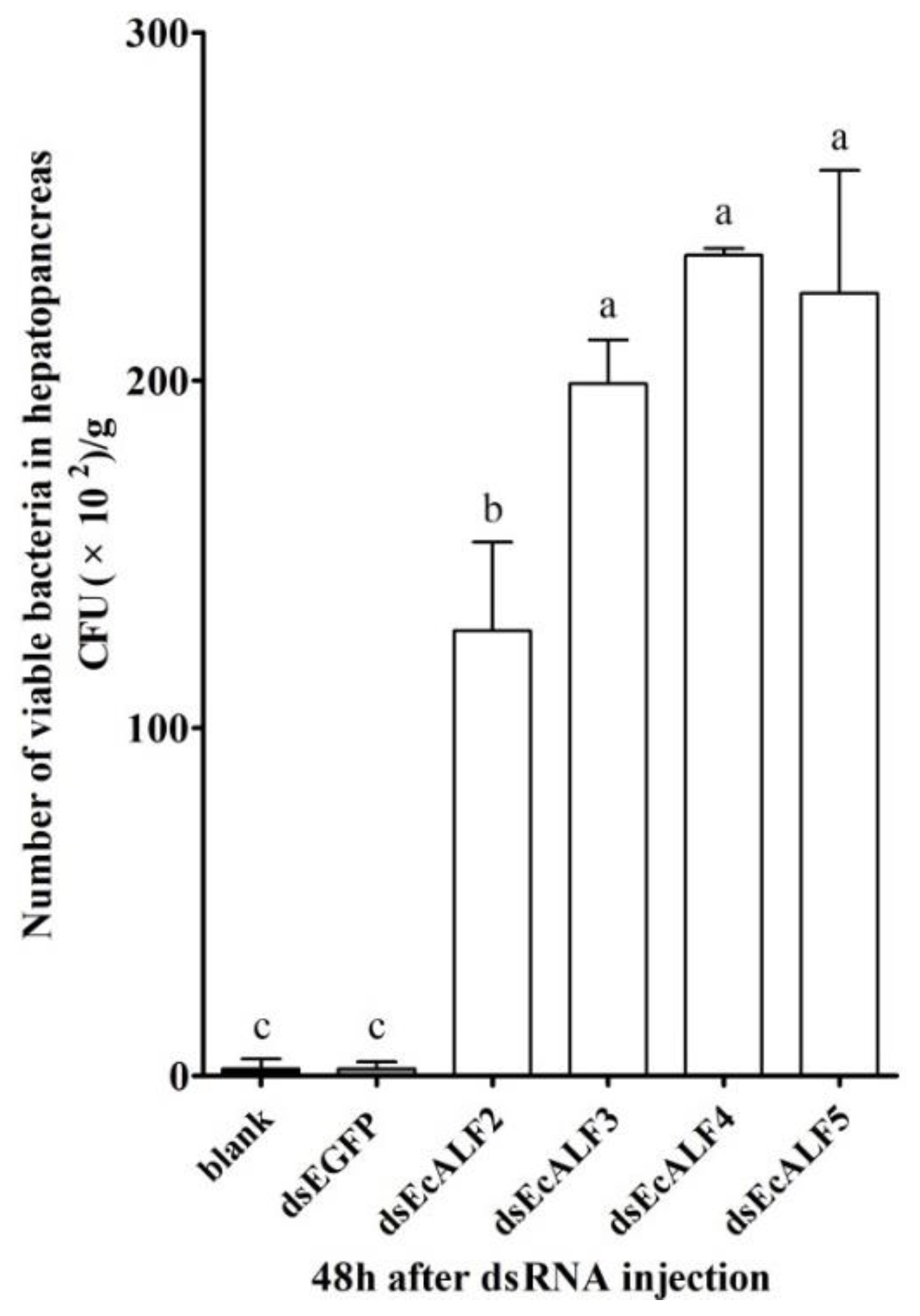
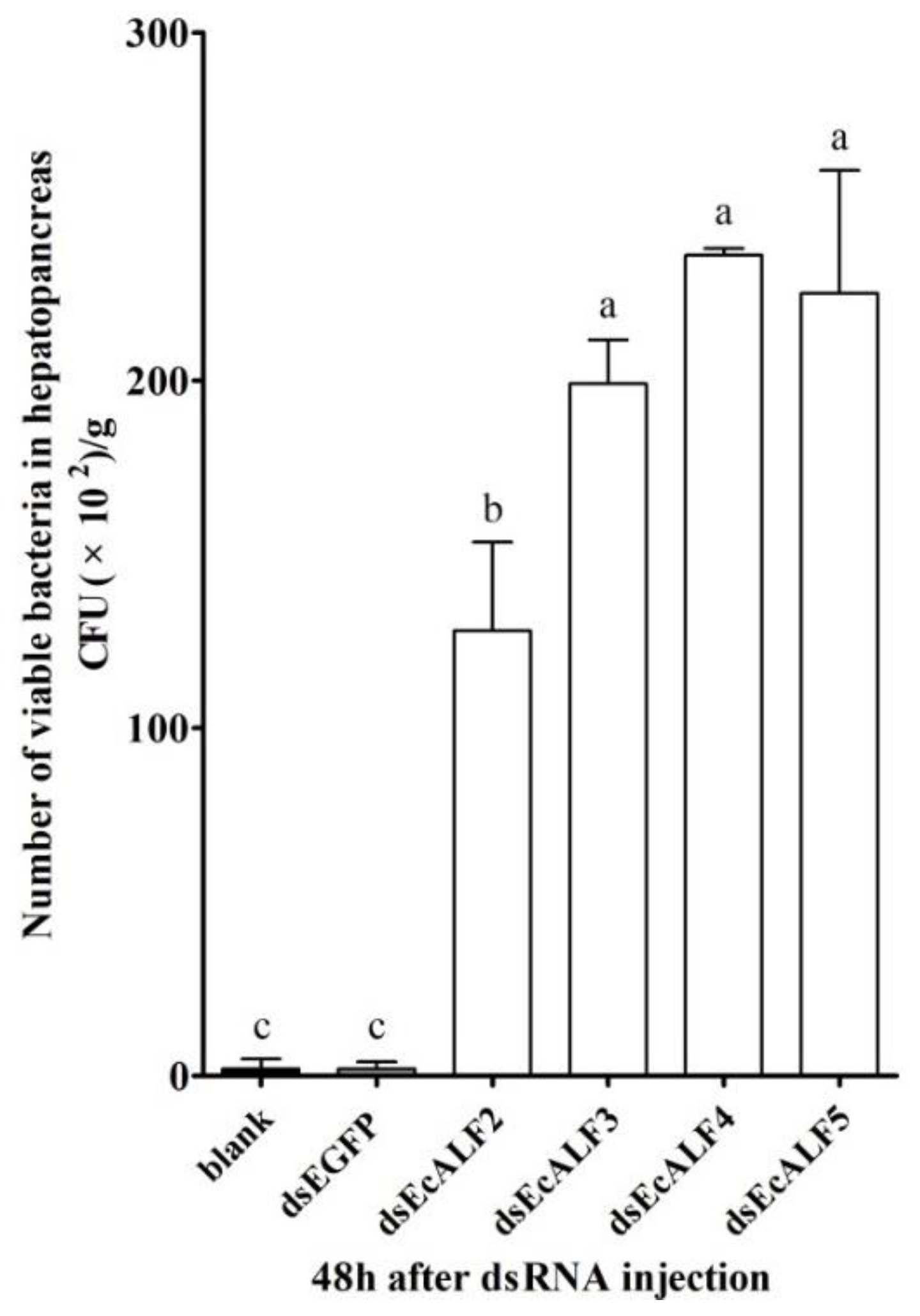
2.6. Effects of EcALFs Silencing on Bacterial Proliferation in the Hepatopancreas of the Prawn
2.7. Hemolytic Property of EcLBD Peptides
3. Discussions
4. Materials and Methods
4.1. Animals and Tissue Sampling
4.2. Sequence Analysis of EcALFs
4.3. Total RNA Extraction and RT-qPCR Analysis
4.4. Synthesis of EcLBD Peptides
4.5. Minimal Inhibitory Concentration (MIC) Assay
4.6. Antiviral Activity of EcLBDs
4.7. DsRNA Synthesis and Gene Silencing of EcALFs
4.8. Hemolytic Activity of EcLBDs
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Steiner, H.; Hultmark, D.; Engstrom, A.; Bennich, H.; Boman, H.G. Sequence and specificity of 2 anti-bacterial proteins involved in insect immunity. Nature 1981, 292, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; van Meegern, A.; Doemming, S.; Schuerholz, T. Antimicrobial peptides in human sepsis. Front. Immunol. 2015, 6, 404. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial peptides from plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.D.; Yu, X.Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.D.; Barracco, M.A. Antimicrobial peptides in crustaceans. Invert. Surviv. J. 2010, 7, 262–284. [Google Scholar]
- Tassanakajon, A.; Amparyup, P.; Somboonwiwat, K.; Supungul, P. Cationic antimicrobial peptides in penaeid shrimp. Mar. Biotechnol. 2011, 13, 639–657. [Google Scholar] [CrossRef] [PubMed]
- Hoess, A.; Watson, S.; Siber, G.R.; Liddington, R. Crystal structure of an endotoxin-neutralizing protein from the horseshoe crab, Limulus anti-LPS factor, at 1.5 A resolution. EMBO J. 1993, 12, 3351–3356. [Google Scholar] [PubMed]
- Hancock, G.E.; Speelman, D.J.; Heers, K.; Bortell, E.; Smith, J.; Cosco, C. Generation of atypical pulmonary inflammatory responses in BALB/c mice after immunization with the native attachment (G) glycoprotein of respiratory syncytial virus. J. Virol. 1996, 70, 7783–7791. [Google Scholar] [PubMed]
- Andra, J.; Lamata, M.; Martinez de Tejada, G.; Bartels, R.; Koch, M.H.; Brandenburg, K. Cyclic antimicrobial peptides based on Limulus anti-lipopolysaccharide factor for neutralization of lipopolysaccharide. Biochem. Pharmacol. 2004, 68, 1297–1307. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Nakamura, T.; Morita, T.; Iwanaga, S. Limulus anti-LPS factor: An anticoagulant which inhibits the endotoxin-mediated activation of Limulus coagulation system. Biochem. Biophys. Res. Commun. 1982, 105, 717–723. [Google Scholar] [CrossRef]
- Morita, T.; Ohtsubo, S.; Nakamura, T.; Tanaka, S.; Iwanaga, S.; Ohashi, K.; Niwa, M. Isolation and biological activities of Limulus anticoagulant (anti-LPS factor) which interacts with lipopolysaccharide (LPS). J. Biochem. 1985, 97, 1611–1620. [Google Scholar] [CrossRef] [PubMed]
- Muta, T.; Miyata, T.; Tokunaga, F.; Nakamura, T.; Iwanaga, S. Primary structure of antilipopolysaccharide factor from American horseshoe crab, Limulus polyphemus. J. Biochem. 1987, 101, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Beale, K.M.; Towle, D.W.; Jayasundara, N.; Smith, C.M.; Shields, J.D.; Small, H.J.; Greenwood, S.J. Anti-lipopolysaccharide factors in the American lobster Homarus americanus: Molecular characterization and transcriptional response to Vibrio fluvialis challenge. Comp. Biochem. Phys. D 2008, 3, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Imjongjirak, C.; Amparyup, P.; Tassanakajon, A.; Sittipraneed, S. Antilipopolysaccharide factor (ALF) of mud crab Scylla paramamosain: Molecular cloning, genomic organization and the antimicrobial activity of its synthetic LPS binding domain. Mol. Immunol. 2007, 44, 3195–3203. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, H.; Inagawa, H.; Morii, K.; Harada, H.; Kohchi, C.; Nishizawa, T.; Taniguchi, Y.; Uenobe, M.; Honda, T.; Kondoh, M.; et al. Cloning and characterization of a LPS-regulatory gene having an LPS binding domain in kuruma prawn Marsupenaeus japonicus. Mol. Immunol. 2006, 43, 2061–2069. [Google Scholar] [CrossRef] [PubMed]
- Somboonwiwat, K.; Marcos, M.; Tassanakajon, A.; Klinbunga, S.; Aumelas, A.; Romestand, B.; Gueguen, Y.; Boze, H.; Moulin, G.; Bachere, E. Recombinant expression and anti-microbial activity of anti-lipopolysaccharide factor (ALF) from the black tiger shrimp Penaeus monodon. Dev. Comp. Immunol. 2005, 29, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Prapavorarat, A.; Pongsomboon, S.; Tassanakajon, A. Identification of genes expressed in response to yellow head virus infection in the black tiger shrimp, Penaeus monodon, by suppression subtractive hybridization. Dev. Comp. Immunol. 2010, 34, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Soonthornchai, W.; Rungrassamee, W.; Karoonuthaisiri, N.; Jarayabhand, P.; Klinbunga, S.; Soderhall, K.; Jiravanichpaisal, P. Expression of immune-related genes in the digestive organ of shrimp, Penaeus monodon, after an oral infection by Vibrio harveyi. Dev. Comp. Immunol. 2010, 34, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Tharntada, S.; Ponprateep, S.; Somboonwiwat, K.; Liu, H.; Soderhall, I.; Soderhall, K.; Tassanakajon, A. Role of anti-lipopolysaccharide factor from the black tiger shrimp, Penaeus monodon, in protection from white spot syndrome virus infection. J. Gen. Virol. 2009, 90, 1491–1498. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Guo, S.; Li, F.; Xiang, J. Characterization and function analysis of an anti-lipopolysaccharide factor (ALF) from the Chinese shrimp Fenneropenaeus chinensis. Dev. Comp. Immunol. 2014, 46, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Ponprateep, S.; Somboonwiwat, K.; Tassanakajon, A. Recombinant anti-lipopolysaccharide factor isoform 3 and the prevention of vibriosis in the black tiger shrimp, Penaeus monodon. Aquaculture 2009, 289, 219–224. [Google Scholar] [CrossRef]
- Ponprateep, S.; Tharntada, S.; Somboonwiwat, K.; Tassanakajon, A. Gene silencing reveals a crucial role for anti-lipopolysaccharide factors from Penaeus monodon in the protection against microbial infections. Fish Shellfish Immunol. 2012, 32, 26–34. [Google Scholar] [CrossRef] [PubMed]
- De la Vega, E.; O’Leary, N.A.; Shockey, J.E.; Robalino, J.; Payne, C.; Browdy, C.L.; Warr, G.W.; Gross, P.S. Anti-lipopolysaccharide factor in Litopenaeus vannamei (LvALF): A broad spectrum antimicrobial peptide essential for shrimp immunity against bacterial and fungal infection. Mol. Immunol. 2008, 45, 1916–1925. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.S.; Zhang, Q.; Zhao, Y.R.; Jia, W.M.; Zhao, X.F.; Wang, J.X. A new group of anti-lipopolysaccharide factors from Marsupenaeus japonicus functions in antibacterial response. Dev. Comp. Immunol. 2015, 48, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Guo, S.; Li, F.; Xiang, J. Functional diversity of anti-lipopolysaccharide factor isoforms in shrimp and their characters related to antiviral activity. Mar. Drugs 2015, 13, 2602–2616. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Mishra, B.; Lau, K.; Lushnikova, T.; Golla, R.; Wang, X. Antimicrobial peptides in 2014. Pharmaceuticals 2015, 8, 123–150. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Li, S.; Liu, F.; Li, F.; Xiang, J. Identification and function analysis of an anti-lipopolysaccharide factor from the ridgetail prawn Exopalaemon carinicauda. Dev. Comp. Immunol. 2017, 70, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.W.; Keyser, P.; Sritunyalucksana, K.; Soderhall, K. Crustacean haemocytes and haematopoiesis. Aquaculture 2000, 191, 45–52. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, T.; Gu, J.; Li, X.; Yang, X.; Gao, X.; Liu, F.; Wang, J. Identification of five anti-lipopolysaccharide factors in oriental river prawn, Macrobrachium nipponense. Fish Shellfish Immunol. 2015, 46, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Imjongjirak, C.; Amparyup, P.; Tassanakajon, A. Molecular cloning, genomic organization and antibacterial activity of a second isoform of antilipopolysaccharide factor (ALF) from the mud crab, Scylla paramamosain. Fish Shellfish Immunol. 2011, 30, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Haldar, S. Vibrio related diseases in aquaculture and development of rapid and accurate identification methods. J. Mar. Sci. Res. Dev. 2012, S1, 002. [Google Scholar]
- Lightner, D.V.; Redman, R.M.; Pantoja, C.R.; Noble, B.L.; Tran, L. Early mortality syndrome affects shrimp in Asia. Glob. Aquac. Adv. 2012, 15, 40. [Google Scholar]
- Rosa, R.D.; Vergnes, A.; de Lorgeril, J.; Goncalves, P.; Perazzolo, L.M.; Sauné, L.; Romestand, B.; Fievet, J.; Gueguen, Y.; Bachère, E.; et al. Functional divergence in shrimp anti-lipopolysaccharide factors (ALFs): From recognition of cell wall components to antimicrobial activity. PLoS ONE 2013, 8, e67937. [Google Scholar] [CrossRef] [PubMed]
- Boto, A.; de la Lastra, J.M.P.; González, C.C. The road from host-defense peptides to a new generation of antimicrobial drugs. Molecules 2018, 23, 311. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, S.; Li, F.; Xiang, J. Structure and bioactivity of a modified peptide derived from the LPS-binding domain of an anti-lipopolysaccharide factor (ALF) of shrimp. Mar. Drugs 2016, 14, 96. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.M.; Li, F.H.; Xiang, J.H. Analysis on the dynamic changes of the amount of WSSV in Chinese shrimp Fenneropenaeus chinensis during infection. Aquaculture 2013, 376, 124–132. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EcLBD2 | Ac-V(CSYQVKPTIRKFELYFKGTFWC)P-NH2 |
| EcLBD3 | Ac-T(CNYRVDPKIKRFQLYFKGRMWC)P-NH2 |
| EcLBD4 | Ac-Y(CVYKRIGYFYKWELNYKAEVRC)P-NH2 |
| EcLBD5 | Ac-A(CQYKRIPYIKRLELHYRAEVRC)L-NH2 |
| pGFP | Ac-TTGKLPVPWPTLVTTFSYGVQCFS-NH2 |
| Bacteria | MIC (μM) | |||
|---|---|---|---|---|
| EcLBD2 | EcLBD3 | EcLBD4 | EcLBD5 | |
| Gram-positive bacteria | ||||
| K. rhizophila ATCC 9341 | >64 | 32–64 | >64 | >64 |
| S. epidermidis ATCC 12228 | >64 | >64 | 32–64 | >64 |
| B. licheniformis ATCC 11946 | >64 | >64 | >64 | >64 |
| Gram-negative bacteria | ||||
| V. alginolyticus ATCC 17749 | >64 | >64 | 16–32 | 16–32 |
| V. harveyi ATCC 33842 | 32–64 | 8–16 | >64 | >64 |
| P. damselae ATCC 33539 | >64 | 32–64 | >64 | >64 |
| E. coli ATCC 25922 | >64 | >64 | >64 | >64 |
| V. parahaemolyticus ATCC 17802 | >64 | >64 | >64 | >64 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, X.; Li, S.; Zhang, C.; Xiang, J.; Li, F. Multiple Isoforms of Anti-Lipopolysaccharide Factors and Their Antimicrobial Functions in the Ridgetail Prawn Exopalaemon carinicauda. Mar. Drugs 2018, 16, 145. https://doi.org/10.3390/md16050145
Lv X, Li S, Zhang C, Xiang J, Li F. Multiple Isoforms of Anti-Lipopolysaccharide Factors and Their Antimicrobial Functions in the Ridgetail Prawn Exopalaemon carinicauda. Marine Drugs. 2018; 16(5):145. https://doi.org/10.3390/md16050145
Chicago/Turabian StyleLv, Xinjia, Shihao Li, Chengsong Zhang, Jianhai Xiang, and Fuhua Li. 2018. "Multiple Isoforms of Anti-Lipopolysaccharide Factors and Their Antimicrobial Functions in the Ridgetail Prawn Exopalaemon carinicauda" Marine Drugs 16, no. 5: 145. https://doi.org/10.3390/md16050145
APA StyleLv, X., Li, S., Zhang, C., Xiang, J., & Li, F. (2018). Multiple Isoforms of Anti-Lipopolysaccharide Factors and Their Antimicrobial Functions in the Ridgetail Prawn Exopalaemon carinicauda. Marine Drugs, 16(5), 145. https://doi.org/10.3390/md16050145