Eicosanoid Diversity of Stony Corals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
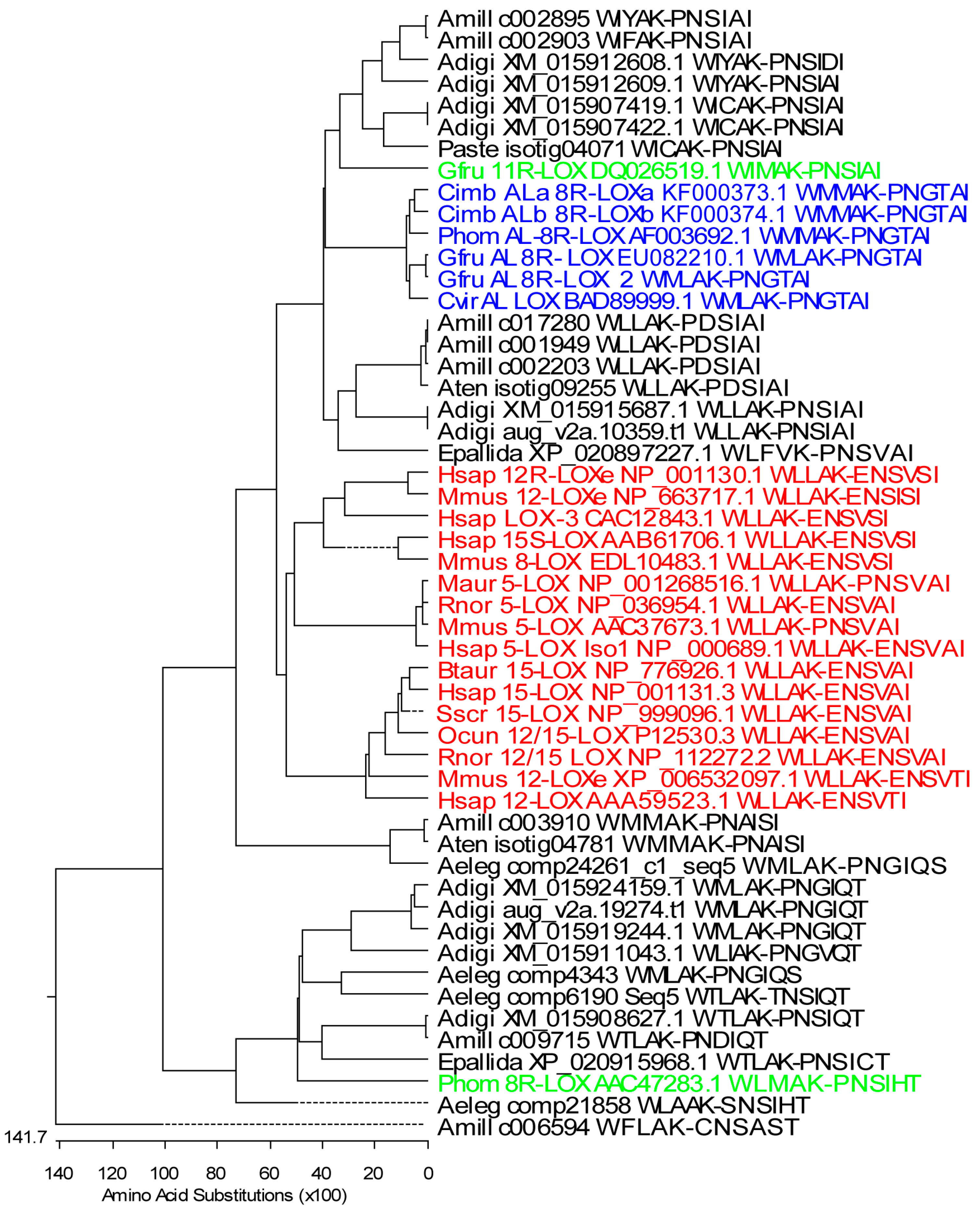
2.1. Stony Coral Dioxygenases
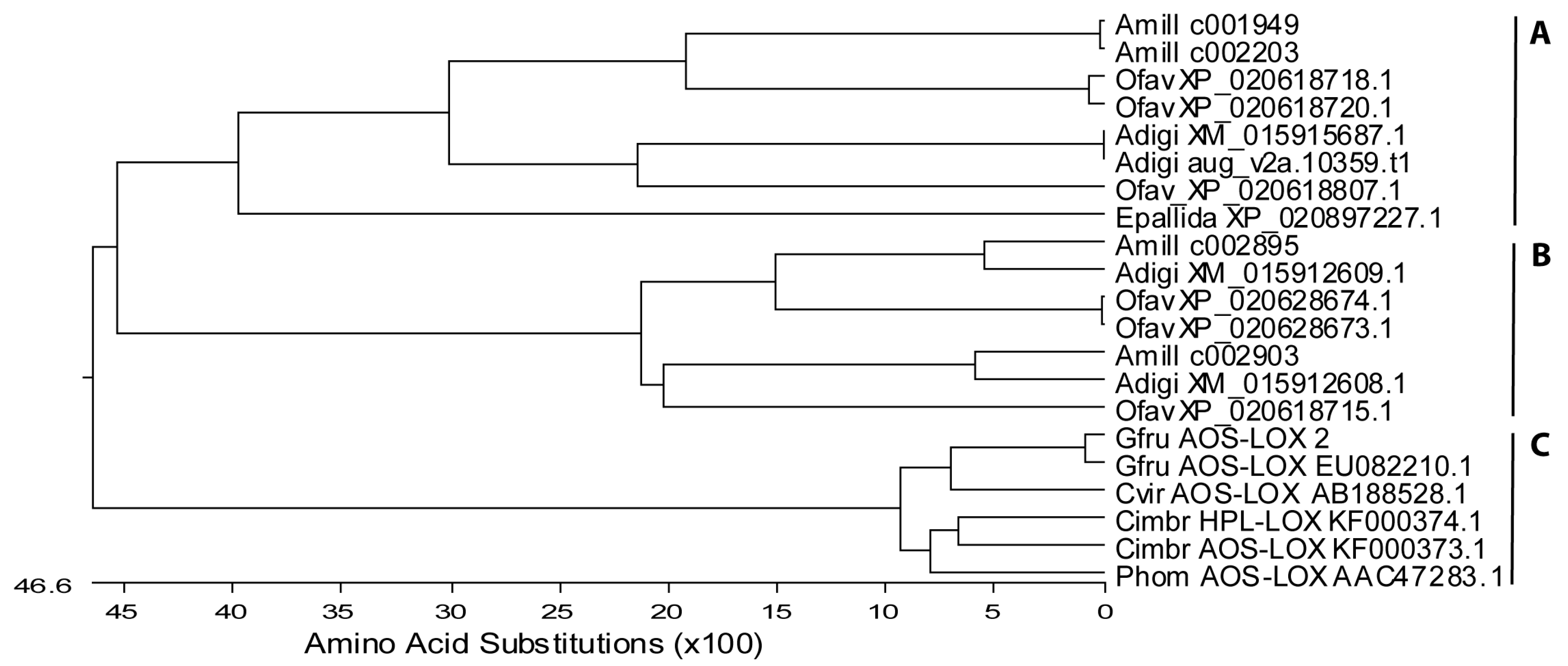
2.1.1. Lipoxygenases
2.1.2. Fusion Proteins
2.1.3. Cyclooxygenases
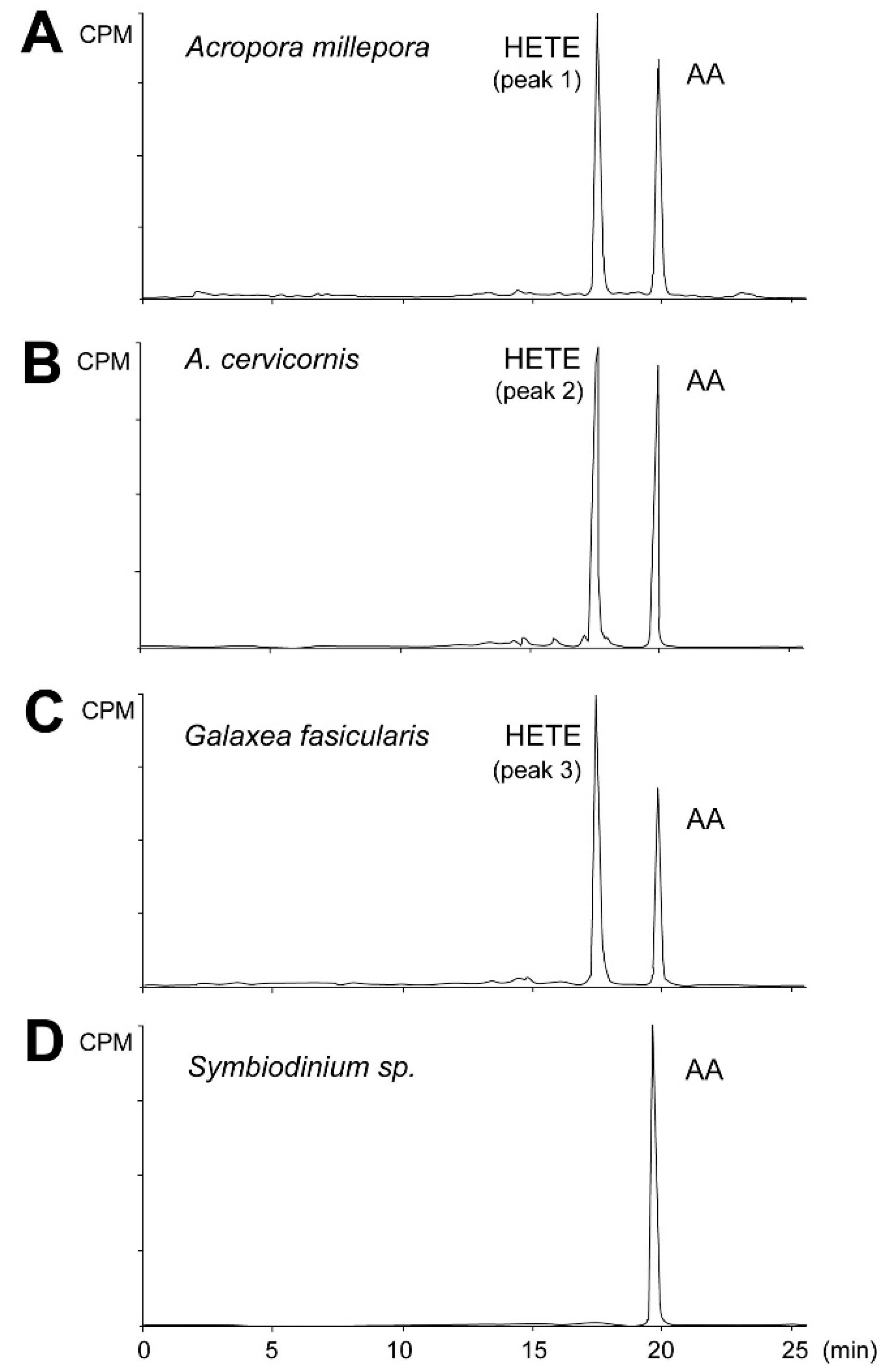
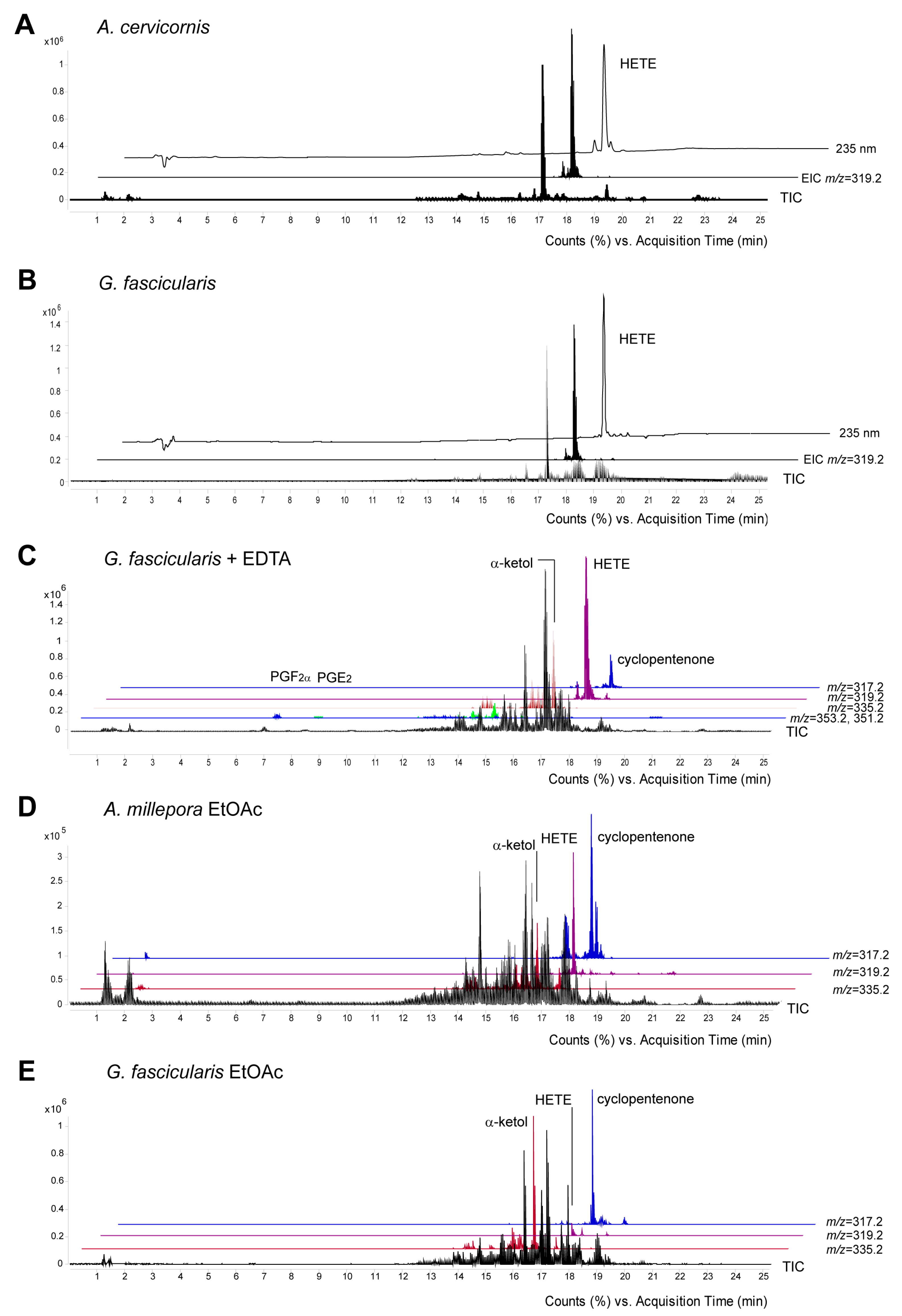
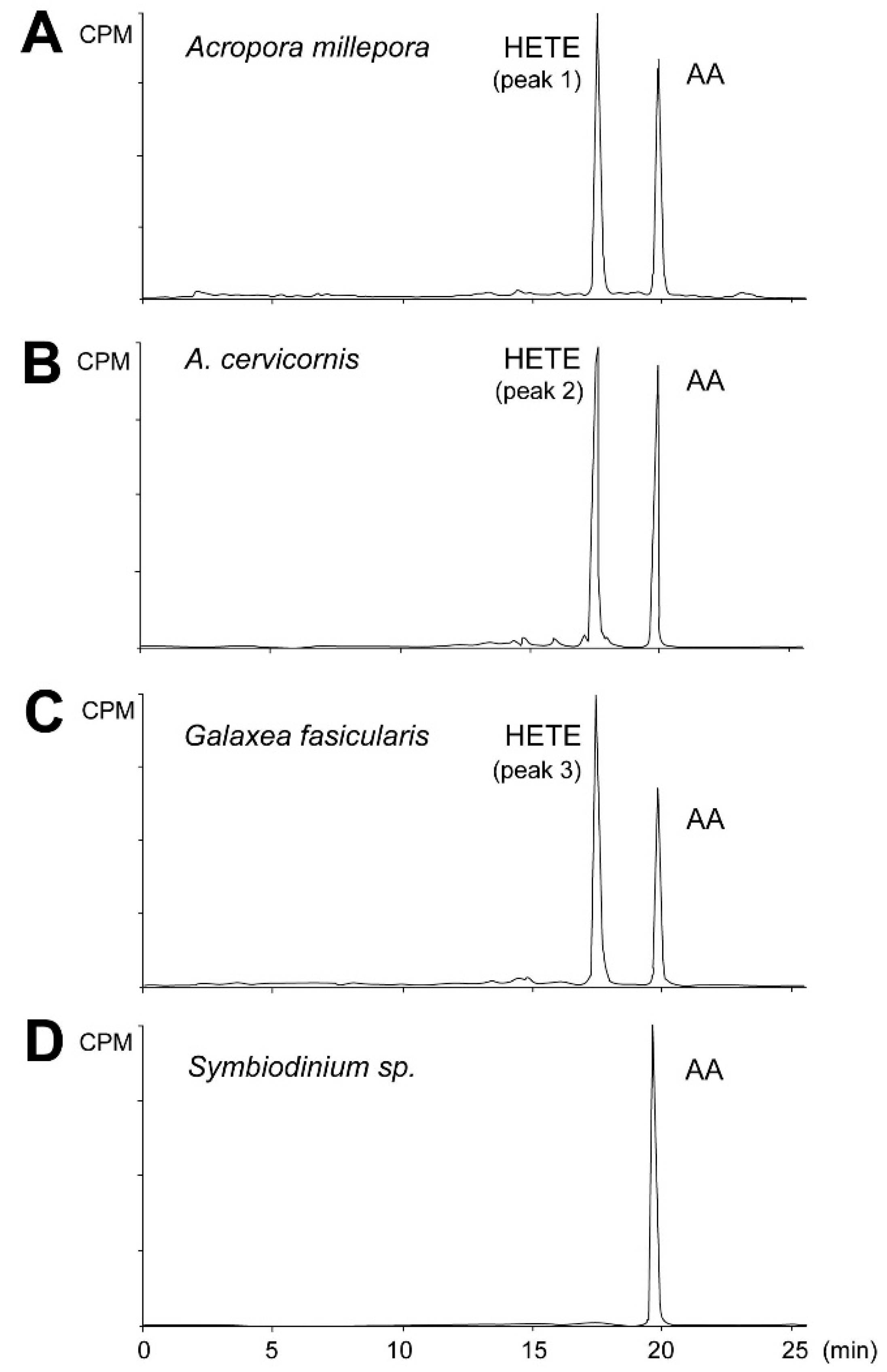
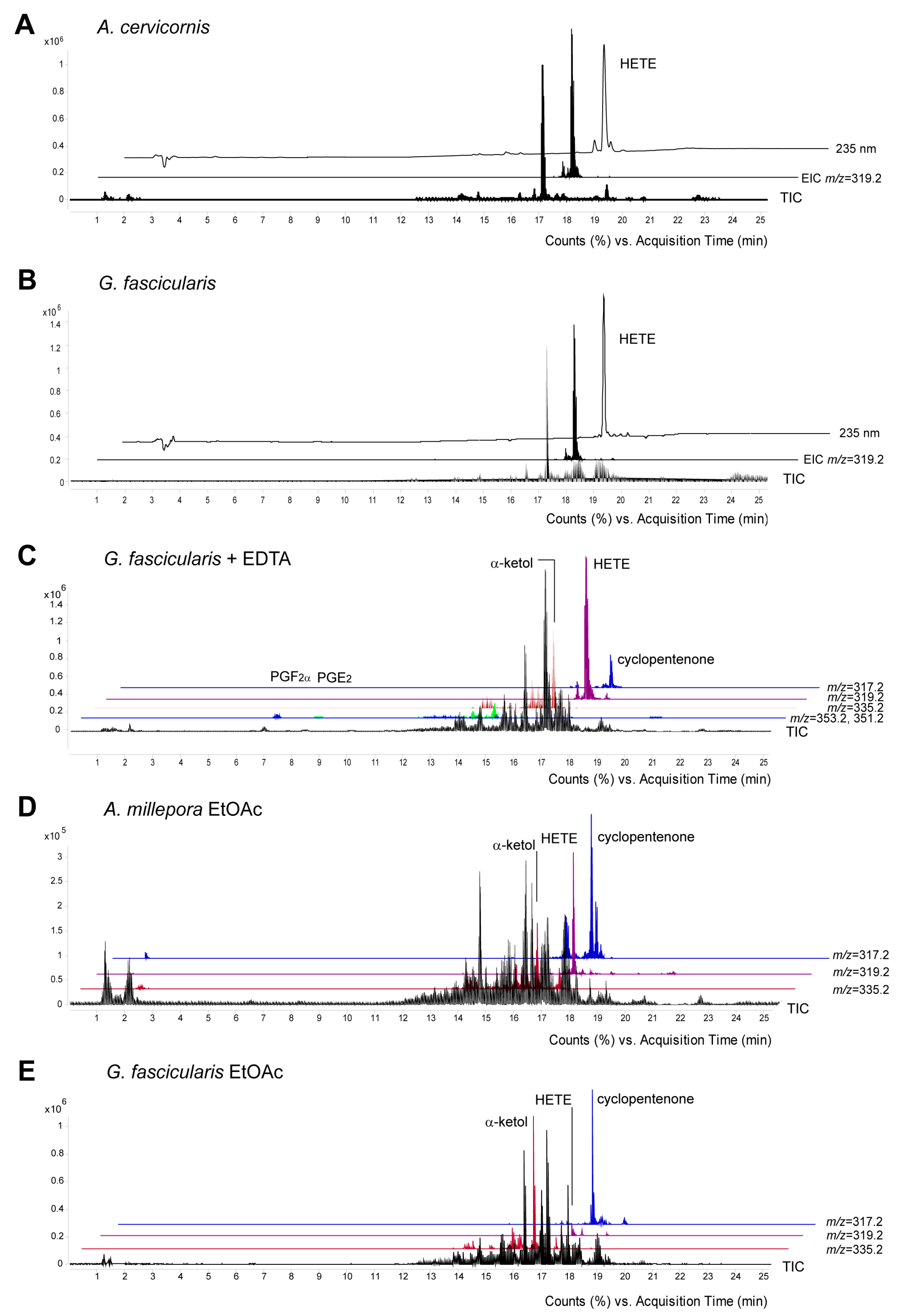
2.2. Eicosanoid Profiling
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Corals
4.3. Sequence Analysis
4.4. Preparation of Samples
4.5. Isolation of Zooxanthellae Cells
4.6. RP-HPLC
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.L.; Urade, Y.; Jakobsson, P.-J. Enzymes of the cyclooxygenase pathways of prostanoid biosynthesis. Chem. Rev. 2011, 111, 5821–5865. [Google Scholar] [CrossRef] [PubMed]
- Brash, A.R. Lipoxygenases: Occurrence, functions, catalysis, and acquisition of substrate. J. Biol. Chem. 1999, 274, 23679–23682. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Buckley, C.D.; Gilroy, D.W.; Serhan, C.N. Proresolving lipid mediators and mechanisms in the resolution of acute inflammation. Immunity 2014, 40, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Z.; DuBois, R.N. Eicosanoids and cancer. Nat. Rev. Cancer 2010, 10, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Kunio, Y.; George, I.; Hiroyuki, N.; Naoki, O.; Yukiko, N. Eicosanoids derived from arachidonic acid and their family prostaglandins and cyclooxygenase in psychiatric disorders. Curr. Neuropharmacol. 2015, 13, 776–785. [Google Scholar]
- Birnie, M.; Morrison, R.; Camara, R.; Strauss, K.I. Temporal changes of cytochrome P450 (Cyp) and eicosanoid-related gene expression in the rat brain after traumatic brain injury. BMC Genomics 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Mashima, R.; Okuyama, T. The role of lipoxygenases in pathophysiology; new insights and future perspectives. Redox Biology 2015, 6, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Al-Shabrawey, M.; Wang, M.-H. Cyclooxygenase- and cytochrome P450-derived eicosanoids in stroke. Prostaglandins Other Lipid Med. 2016, 122, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Schauberger, E.; Peinhaupt, M.; Cazares, T.; Lindsley, A.W. Lipid mediators of allergic disease: Pathways, treatments, and emerging therapeutic targets. Curr. Allergy Asthma Rep. 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Weinheimer, A.J.; Spraggins, R.L. The occurrence of two new prostaglandin derivatives (15-epi-PGA2 and its acetate, methyl ester) in the gorgonian Plexaura homomalla chemistry of coelenterates. XV. Tetrahedron Lett. 1969, 10, 5185–5188. [Google Scholar] [CrossRef]
- Corey, E.J.; Washburn, W.N.; Chen, J.C. Studies on the prostaglandin A2 synthetase complex from Plexaura homomalla. J. Am. Chem. Soc. 1973, 95, 2054–2055. [Google Scholar] [CrossRef] [PubMed]
- Varvas, K.; Järving, I.; Koljak, R.; Vahemets, A.; Pehk, T.; Müürisepp, A.-M.; Lille, Ü.; Samel, N. Invitro niosynthesis of prostaglandins in the White Sea soft coral Gersemia fruticosa: Formation of optically active Pgd2, Pge2α, Pgf2-Alpha and 15-Keto-Pgf2α from arachidonicacid. Tetrahedron Lett. 1993, 34, 3643–3646. [Google Scholar] [CrossRef]
- Imbs, A.B.; Demina, O.A.; Demidkova, D.A. Lipid class and fatty acid composition of the boreal soft coral Gersemia rubiformis. Lipids 2006, 41, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Imbs, A.B.; Yakovleva, I.M. Dynamics of lipid and fatty acid composition of shallow-water corals under thermal stress: An experimental approach. Coral Reefs 2011, 31, 41–53. [Google Scholar] [CrossRef]
- Varvas, K.; Järving, I.; Koljak, R.; Valmsen, K.; Brash, A.R.; Samel, N. Evidence of a cyclooxygenase-related prostaglandin synthesis in coral. The allene oxide pathway is not involved in prostaglandin biosynthesis. J. Biol. Chem. 1999, 274, 9923–9929. [Google Scholar] [CrossRef] [PubMed]
- Koljak, R.; Järving, I.; Kurg, R.; Boeglin, W.E.; Varvas, K.; Valmsen, K.; Ustav, M.; Brash, A.R.; Samel, N. The basis of prostaglandin synthesis in coral: Molecular cloning and expression of a cyclooxygenase from the Arctic soft coral Gersemia fruticosa. J. Biol. Chem. 2001, 276, 7033–7040. [Google Scholar] [CrossRef] [PubMed]
- Valmsen, K.; Järving, I.; Boeglin, W.E.; Varvas, K.; Koljak, R.; Pehk, T.; Brash, A.R.; Samel, N. The origin of 15R-prostaglandins in the Caribbean coral Plexaura homomalla: Molecular cloning and expression of a novel cyclooxygenase. Proc. Natl. Acad. Sci. USA 2001, 98, 7700–7705. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, M.; Järving, R.; Brash, A.R.; Samel, N.; Järving, I. Identification and characterization of an arachidonate 11R-lipoxygenase. Arch. Biochem. Biophys. 2006, 445, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Koljak, R.; Boutaud, O.; Shieh, B.-H.; Samel, N.; Brash, A.R. Identification of a naturally occurring peroxidase-lipoxygenase fusion protein. Science 1997, 277, 1994–1996. [Google Scholar] [CrossRef] [PubMed]
- Lõhelaid, H.; Järving, R.; Valmsen, K.; Varas, K.; Kreen, M.; Järving, I.; Samel, N. Identification of a functional allene oxide synthase-lipoxygenase fusion protein in the soft coral Gersemia fruticosa suggests the generality of this pathway in octocorals. Biochim. Biophys. Acta 2008, 1780, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Teder, T.; Lõhelaid, H.; Boeglin, W.E.; Calcutt, W.M.; Brash, A.R.; Samel, N. A Catalase-related hemoprotein in coral is specialized for synthesis of short-chain aldehydes. J. Biol. Chem. 2015, 290, 19823–19832. [Google Scholar] [CrossRef] [PubMed]
- Teder, T.; Lõhelaid, H.; Samel, N. Structural and functional insights into the reaction specificity of catalase-related hydroperoxide lyase: A shift from lyase activity to allene oxide synthase by site-directed mutagenesis. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Lõhelaid, H.; Teder, T.; Samel, N. Lipoxygenase-allene oxide synthase pathway in octocoral thermal stress response. Coral Reefs 2014, 34, 143–154. [Google Scholar] [CrossRef]
- Lõhelaid, H.; Teder, T.; Tõldsepp, K.; Ekins, M.; Samel, N. Up-regulated expression of AOS-LOXa and increased eicosanoid synthesis in response to coral wounding. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Bellantuono, A.J.; Hoegh-Guldberg, O.; Rodriguez-Lanetty, M. Resistance to thermal stress in corals without changes in symbiont composition. Proc. R. Soc. B Biol. Sci. 2011, 279, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Siboni, N.; Abrego, D.; Seneca, F.; Motti, C.A.; Andreakis, N.; Tebben, J.; Blackall, L.L.; Harder, T. Using bacterial extract along with differential gene expression in acropora millepora larvae to decouple the processes of attachment and metamorphosis. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Moya, A.; Huisman, L.; Ball, E.E.; Hayward, D.C.; Grasso, L.C.; Chua, C.M.; Woo, H.N.; Gattuso, J.-P.; Forêt, S.; Miller, D.J. Whole transcriptome analysis of the coral acropora millepora reveals complex responses to CO2-driven acidification during the Initiation of calcification. Mol. Ecol. 2012, 21, 2440–2454. [Google Scholar] [CrossRef] [PubMed]
- Polato, N.R.; Vera, J.C.; Baums, I.B. Gene discovery in the threatened elkhorn coral: 454 sequencing of the Acropora palmata transcriptome. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Meyer, E.; Aglyamova, G.V.; Matz, M.V. Profiling gene expression responses of coral larvae (Acropora millepora) to elevated temperature and settlement inducers using a novel RNA-Seq procedure. Mol. Ecol. 2011, 20, 3599–3616. [Google Scholar] [CrossRef] [PubMed]
- Voolstra, C.R.; Sunagawa, S.; Matz, M.V.; Bayer, T.; Aranda, M.; Buschiazzo, E.; Desalvo, M.K.; Lindquist, E.; Szmant, A.M.; Coffroth, M.A.; et al. Rapid evolution of coral proteins responsible for interaction with the environment. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.M.; Aglyamova, G.V.; Meyer, E.; Matz, M.V. Gene expression associated with white syndromes in a reef building coral, Acropora hyacinthus. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Barshis, D.J.; Ladner, J.T.; Oliver, T.A.; Seneca, F.O.; Traylor-Knowles, N.; Palumbi, S.R. Genomic basis for coral resilience to climate change. Proc. Natl. Acad. Sci. USA 2013, 110, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Souter, P.; Bay, L.K.; Andreakis, N.; Császár, N.; Seneca, F.O.; Van Oppen, M.J.H. A multilocus, temperature stress-related gene expression profile assay in Acropora millepora, a dominant reef-building coral. Mol. Ecol. Resour. 2011, 11, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Portune, K.J.; Voolstra, C.R.; Medina, M.; Szmant, A.M. Development and heat stress-induced transcriptomic changes during embryogenesis of the scleractinian coral Acropora palmata. Mar. Genom. 2010, 3, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Dupiol, J.; Zoccola, D.; Tambutté, E.; Grunau, C.; Cosseau, C.; Smith, K.M.; Freitag, M.; Dheilly, N.M.; Allemand, D.; Tambutté, S. Genes related to ion-transport and energy production are upregulated in response to CO2-driven pH decrease in corals: New insights from transcriptome analysis. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Libro, S.; Kaluziak, S.T.; Vollmer, S.V. RNA-seq profiles of immune related genes in the staghorn coral acropora cervicornis infected with white band disease. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Latyshev, N.A.; Naumenko, N.V.; Svetashev, V.I.; Latypov, Y.Y. Fatty-acids of reef-building corals. Mar. Ecol. Prog. Ser. 1991, 76, 295–301. [Google Scholar] [CrossRef]
- Figueiredo, J.; Baird, A.H.; Cohen, M.F.; Flot, J.-F.; Kamiki, T.; Meziane, T.; Tsuchiya, M.; Yamasaki, H. Ontogenetic change in the lipid and fatty acid composition of scleractinian coral larvae. Coral Reefs 2012, 31, 613–619. [Google Scholar] [CrossRef]
- Dunn, S.R.; Thomas, M.C.; Nette, G.W.; Dove, S.G. A lipidomic approach to understanding free fatty acid lipogenesis derived from dissolved inorganic carbon within cnidarian-dinoflagellate symbiosis. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Varvas, K.; Kasvandik, S.; Hansen, K.; Järving, I.; Morell, I.; Samel, N. Structural and catalytic insights into the algal prostaglandin H synthase reveal atypical features of the first non-animal cyclooxygenase. (BBA)-Mol. Cell Biol. Lipids 2013, 1831, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Varvas, K.; Kurg, R.; Hansen, K.; Järving, R.; Järving, I.; Valmsen, K.; Lõhelaid, H.; Samel, N. Direct evidence of the cyclooxygenase pathway of prostaglandin synthesis in arthropods: Genetic and biochemical characterization of two crustacean cyclooxygenases. Insect Biochem. Mol. Biol. 2009, 39, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Selinsky, B.S. Bacterial and algal orthologs of prostaglandin H2 synthase: Novel insights into the evolution of an integral membrane protein. Biochim. Biophys. Acta (BBA) Biomembr. 2015, 1848, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Havird, J.C.; Kocot, K.M.; Brannock, P.M.; Cannon, J.T.; Waits, D.S.; Weese, D.A.; Santos, S.R.; Halanych, K.M. Reconstruction of cyclooxygenase evolution in animals suggests variable, lineage-specific duplications, and homologs with low sequence identity. J. Mol. Evol. 2015, 80, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Nioche, P.; Hamberg, M.; Raman, C.S. Structural insights into the evolutionary paths of oxylipin biosynthetic enzymes. Nature 2008, 455, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Coffa, G.; Schneider, C.; Brash, A.R. A comprehensive model of positional and stereo control in lipoxygenases. Biochem. Biophys. Res. Commun. 2005, 338, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Corey, E.J.; Matsuda, S.P.T.; Nagata, R.; Cleaver, M.B. Biosynthesis of 8-R-Hpete and preclavulone-a from arachidonate in several species of caribbean coral. A Widespread Route to Marine Prostanoids. Tetrahedron Lett. 1988, 29, 2555–2558. [Google Scholar] [CrossRef]
- Murphy, R.C.; Gijón, M.A. Biosynthesis and metabolism of leukotrienes. Biochem. J. 2007, 405, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.V.; Traylor-Knowles, N.G.; Willis, B.L.; Bythell, J.C. Corals use similar immune cells and wound-healing processes as those of higher organisms. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Shinzato, C.; Shoguchi, E.; Kawashima, T.; Hamada, M.; Hisata, K.; Tanaka, M.; Fujie, M.; Fujiwara, M.; Koyanagi, R.; Ikuta, T.; et al. Using the Acropora digitifera genome to understand coral responses to environmental change. Nature 2011, 476, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Kenkel, C.D.; Meyer, E.; Matz, M.V. Gene expression under chronic heat stress in populations of the mustard hill coral (Porites astreoides) from different thermal environments. Mol. Ecol. 2013, 22, 4322–4334. [Google Scholar] [CrossRef] [PubMed]
- matzLABDATA. Available online: http://www.bio.utexas.edu/research/matz_lab/matzlab/Data.html (accessed on 15 January 2017).
- Annotated Reference Traqnscriptome Assemblies. Available online: http://people.oregonstate.edu/~meyere/data.html (accessed on 15 January 2017).
- Translate. Available online: http://web.expasy.org/translate/ (accessed on 16 January 2017).
- Rowan, R.; Powers, D.A. Molecular Genetic Identification of Symbiotic Dinoflagellates (Zooxanthellae). Mar. Ecol. Prog. Ser. 1991, 71, 65–73. [Google Scholar] [CrossRef]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lõhelaid, H.; Samel, N. Eicosanoid Diversity of Stony Corals. Mar. Drugs 2018, 16, 10. https://doi.org/10.3390/md16010010
Lõhelaid H, Samel N. Eicosanoid Diversity of Stony Corals. Marine Drugs. 2018; 16(1):10. https://doi.org/10.3390/md16010010
Chicago/Turabian StyleLõhelaid, Helike, and Nigulas Samel. 2018. "Eicosanoid Diversity of Stony Corals" Marine Drugs 16, no. 1: 10. https://doi.org/10.3390/md16010010
APA StyleLõhelaid, H., & Samel, N. (2018). Eicosanoid Diversity of Stony Corals. Marine Drugs, 16(1), 10. https://doi.org/10.3390/md16010010