
Alleviation of Hepatic Steatosis by Alpha-Defensin Is Associated with Enhanced Lipolysis
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Core Tip:
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Plasma Analysis
2.3. Quantification of Total Hepatic Lipid
2.4. Liver Fat Fraction Quantification Using EchoMRI
2.5. Tissue Processing and Histological Analyses
2.6. Cell Isolation, Staining, and Flow Cytometric Analysis
2.7. Metabolic Caging Analysis
2.8. Statistical Analysis
3. Results
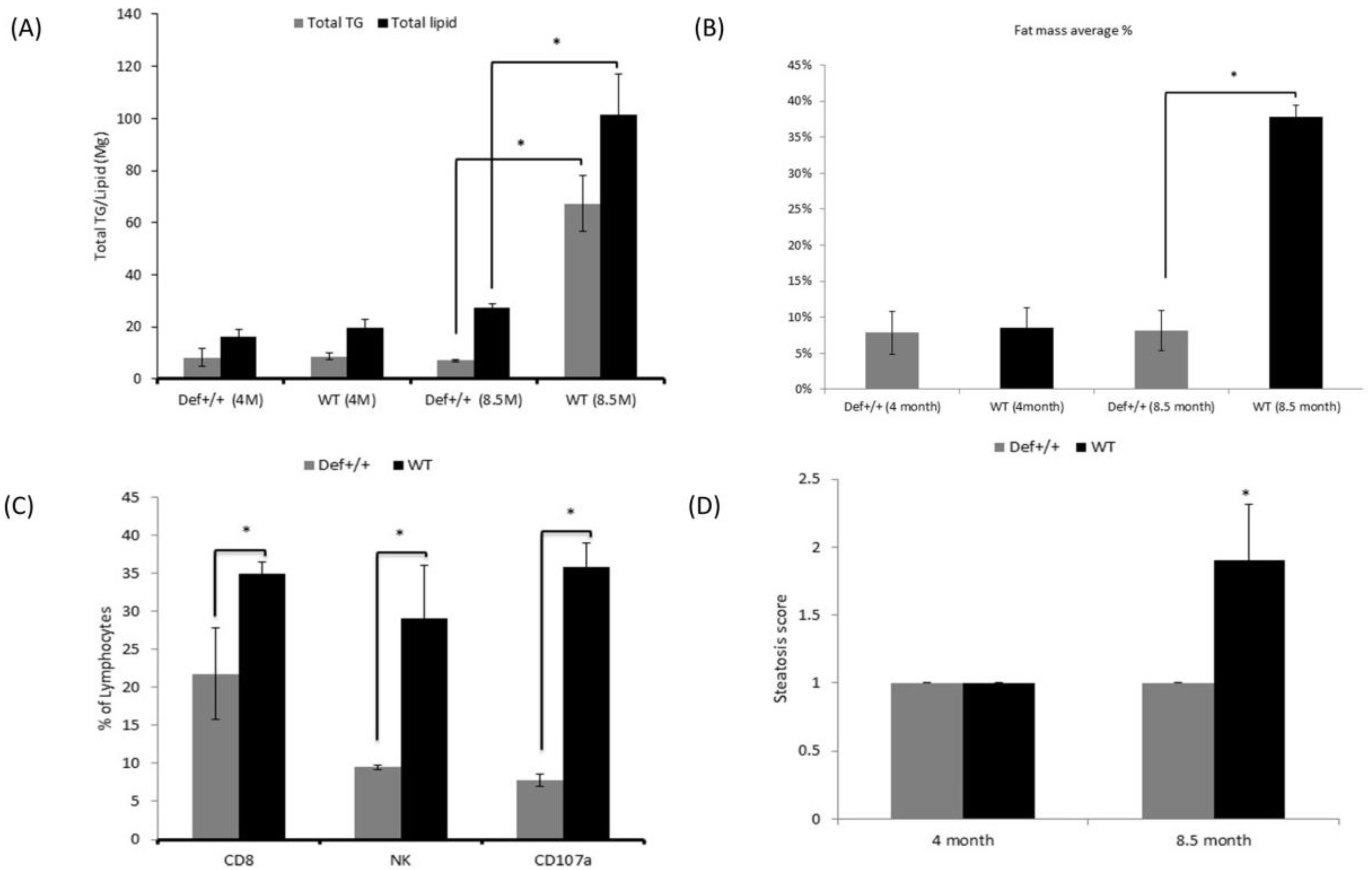
3.1. Plasma Biochemical Markers
3.2. Quantification of Total Hepatic Lipid
3.3. Liver Fat Fraction Quantification using EchoMRI
3.4. Lymphocyte Isolation
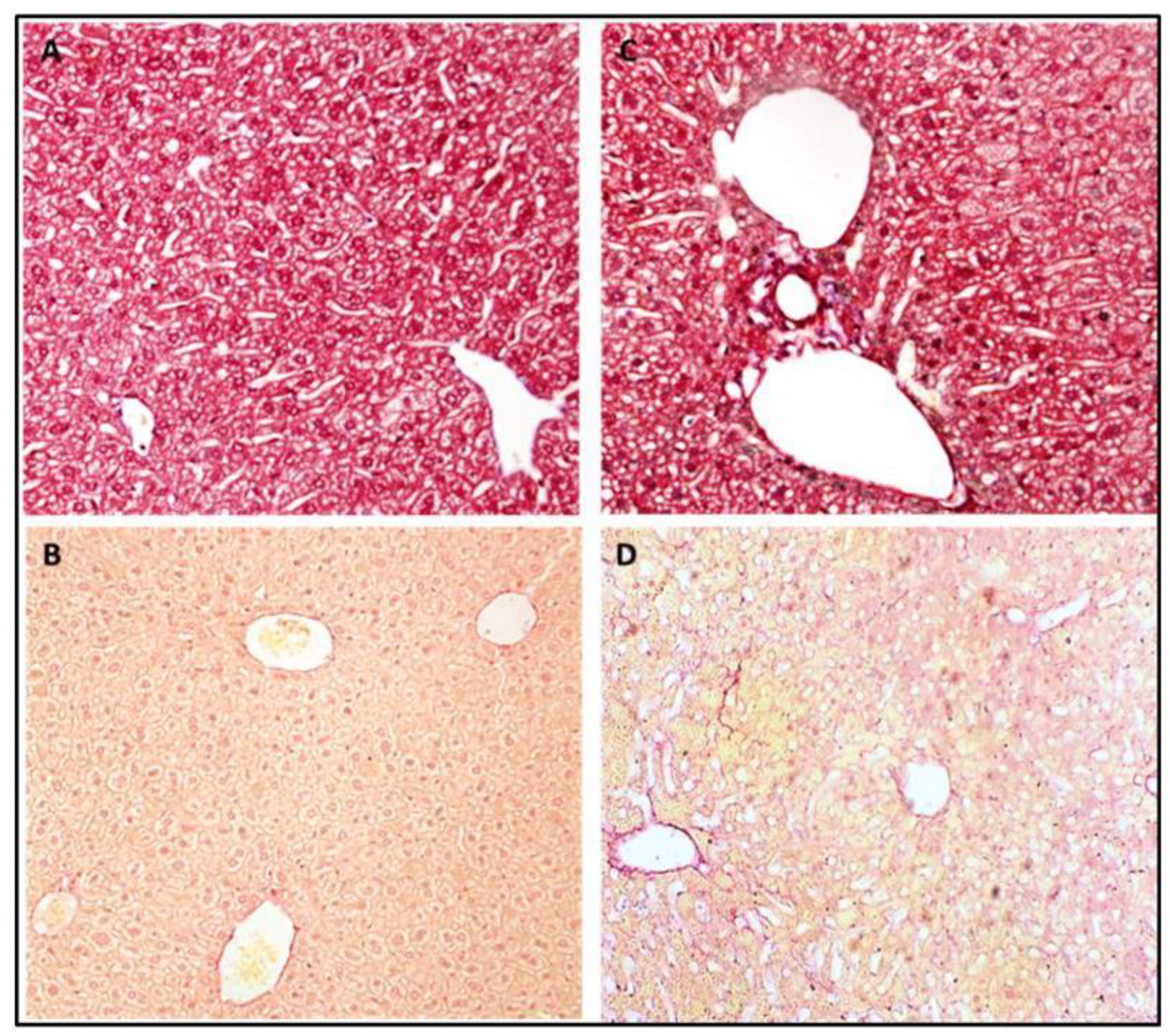
3.5. Tissue Processing and Histological Profile
3.6. Metabolic Caging Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martin, K.; Hatab, A.; Athwal, V.S.; Jokl, E.; Piper Hanley, K. Genetic Contribution to Non-alcoholic Fatty Liver Disease and Prognostic Implications. Curr. Diabetes Rep. 2021, 21, 8. [Google Scholar] [CrossRef]
- Salari, N.; Darvishi, N.; Mansouri, K.; Ghasemi, H.; Hosseinian-Far, M.; Darvishi, F.; Mohammadi, M. Association between PNPLA3 rs738409 polymorphism and nonalcoholic fatty liver disease: A systematic review and meta-analysis. BMC Endocr. Disord. 2021, 21, 125. [Google Scholar] [CrossRef]
- Rojano-Toimil, A.; Rivera-Esteban, J.; Manzano-Nuñez, R.; Bañares, J.; Selva, D.M.; Gabriel-Medina, P.; Ferrer, R.; Pericàs, J.M.; Ciudin, A. When Sugar Reaches the Liver: Phenotypes of Patients with Diabetes and NAFLD. J. Clin. Med. 2022, 11, 3286. [Google Scholar] [CrossRef] [PubMed]
- Atamni, H.J.; Botzman, M.; Mott, R.; Gat-Viks, I.; Iraqi, F.A. Mapping liver fat female-dependent quantitative trait loci in collaborative cross mice. Mamm. Genome 2016, 27, 565–573. [Google Scholar] [CrossRef]
- Huang, D.Q.; El-Serag, H.B.; Loomba, R. Global epidemiology of NAFLD-related HCC: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 223–238. [Google Scholar] [CrossRef]
- Juanola, O.; Martínez-López, S.; Francés, R.; Gómez-Hurtado, I. Non-Alcoholic Fatty Liver Disease: Metabolic, Genetic, Epigenetic and Environmental Risk Factors. Int. J. Environ. Res. Public Health 2021, 18, 5227. [Google Scholar] [CrossRef] [PubMed]
- Lindenmeyer, C.C.; McCullough, A.J. The Natural History of Nonalcoholic Fatty Liver Disease-An Evolving View. Clin. Liver Dis. 2018, 22, 11–21. [Google Scholar] [CrossRef]
- Targher, G. Long-term outcomes in patients with non-alcoholic fatty liver disease: Further evidence that a multidisciplinary and patient-centred approach to treatment is needed. Hepatobiliary Surg. Nutr. 2022, 11, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Jitrukthai, S.; Kositamongkol, C.; Boonchai, P.; Mepramoon, E.; Ariyakunaphan, P.; Nimitpunya, P.; Srivanichakorn, W.; Chaisathaphol, T.; Washirasaksiri, C.; Auesomwang, C.; et al. Long-Term Outcomes Associated with NAFLD, ASCVD, and All-Cause Mortality of Patients with Metabolic Syndrome. J. Clin. Med. 2022, 11, 4627. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Van Natta, M.L.; Clark, J.; Neuschwander-Tetri, B.A.; Diehl, A.; Dasarathy, S.; Loomba, R.; Chalasani, N.; Kowdley, K.; Hameed, B.; et al. Prospective Study of Outcomes in Adults with Nonalcoholic Fatty Liver Disease. N. Engl. J. Med. 2021, 385, 1559–1569. [Google Scholar] [CrossRef]
- Wasilewska, N.; Lebensztejn, D.M. Non-alcoholic fatty liver disease and lipotoxicity. Clin. Exp. Hepatol. 2021, 7, 1–6. [Google Scholar] [CrossRef]
- Abu-Toamih Atamni, H.J.; Ziner, Y.; Mott, R.; Wolf, L.; Iraqi, F.A. Glucose tolerance female-specific QTL mapped in collaborative cross mice. Mamm. Genome 2017, 28, 20–30. [Google Scholar] [CrossRef]
- Rensen, S.S.; Slaats, Y.; Nijhuis, J.; Jans, A.; Bieghs, V.; Driessen, A.; Malle, E.; Greve, J.W.; Buurman, W.A. Increased hepatic myeloperoxidase activity in obese subjects with nonalcoholic steatohepatitis. Am. J. Pathol. 2009, 175, 1473–1482. [Google Scholar] [CrossRef]
- Ikura, Y.; Ohsawa, M.; Suekane, T.; Fukushima, H.; Itabe, H.; Jomura, H.; Nishiguchi, S.; Inoue, T.; Naruko, T.; Ehara, S.; et al. Localization of oxidized phosphatidylcholine in nonalcoholic fatty liver disease: Impact on disease progression. Hepatology 2006, 43, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Rensen, S.S.; Bieghs, V.; Xanthoulea, S.; Arfianti, E.; Bakker, J.A.; Shiri-Sverdlov, R.; Hofker, M.H.; Greve, J.W.; Buurman, W.A. Neutrophil-derived myeloperoxidase aggravates non-alcoholic steatohepatitis in low-density lipoprotein receptor-deficient mice. PLoS ONE. 2012, 7, e52411. [Google Scholar] [CrossRef] [PubMed]
- Chairatana, P.; Nolan, E.M. Human α-Defensin 6: A Small Peptide That Self-Assembles and Protects the Host by Entangling Microbes. Acc. Chem. Res. 2017, 50, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Lu, W. Defensins: A Double-Edged Sword in Host Immunity. Front. Immunol. 2020, 11, 764. [Google Scholar] [CrossRef]
- Jin, G.; Weinberg, A. Human antimicrobial peptides and cancer. Semin. Cell Dev. Biol. 2019, 88, 156–162. [Google Scholar] [CrossRef]
- Ibusuki, R.; Uto, H.; Arima, S.; Mawatari, S.; Setoguchi, Y.; Iwashita, Y.; Hashimoto, S.; Maeda, T.; Tanoue, S.; Kanmura, S.; et al. Transgenic expression of human neutrophil peptide-1 enhances hepatic fibrosis in mice fed a choline-deficient, L-amino acid–defined diet. Liver Intl. 2013, 33, 1549–1556. [Google Scholar] [CrossRef]
- Yang, D.; Biragyn, A.; Hoover, D.M.; Lubkowski, J.; Oppenheim, J.J. Multiple roles of antimicrobial defensins, cathelicidins, and eosinophil-derived neurotoxin in host defense. Annu. Rev. Immunol. 2004, 22, 181–215. [Google Scholar] [CrossRef]
- Bdeir, K.; Higazi, A.A.R.; Kulikovskaya, I.; Christofidou-Solomidou, M.; Vinogradov, S.A.; Allen, T.C.; Idell, S.; Linzmeier, R.; Ganz, T.; Cines, D.B. Neutrophil α-defensins cause lung injury by disrupting the capillary-epithelialbarrier. Am. J. Respir. Crit. Care Med. 2010, 181, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Abu-Fanne, R.; Maraga, E.; Abd-Elrahman, I.; Hankin, A.; Blum, G.; Abdeen, S.; Hijazi, N.; Cines, D.B.; Higazi, A.A.R. α-Defensins Induce a Post-translational Modification of Low Density Lipoprotein (LDL) That Promotes Atherosclerosis at Normal Levels of Plasma Cholesterol. J. Biol. Chem. 2016, 291, 2777–2786. [Google Scholar] [CrossRef] [PubMed]
- Glenthøj, A.; Cowland, J.B.; Heegaard, N.H.; Larsen, M.T.; Borregaard, N. Serglycin participates in retention of α-defensin in granules during myelopoiesis. Blood 2011, 118, 4440–4448. [Google Scholar] [CrossRef]
- Mopuri, R.; Kalyesubula, M.; Rosov, A.; Edery, N.; Moallem, U.; Dvir, H. Improved Folch method for liver-fat quantification. Front. Vet. Sci. 2021, 7, 594853. [Google Scholar] [CrossRef]
- Cataldo, I.; Sarcognato, S.; Sacchi, D.; Cacciatore, M.; Baciorri, F.; Mangia, A.; Cazzagon, N.; Guido, M. Pathology of non-alcoholic fatty liver disease. Pathologica 2021, 113, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Kasper, P.; Martin, A.; Lang, S.; Kütting, F.; Goeser, T.; Demir, M.; Steffen, H.-M. NAFLD and cardiovascular diseases: A clinical review. Clin. Res. Cardiol. 2021, 110, 921–937. [Google Scholar] [CrossRef]
- Soehnlein, O.; Kai-Larsen, Y.; Frithiof, R.; Sorensen, O.E.; Kenne, E.; Scharffetter-Kochanek, K.; Eriksson, E.E.; Herwald, H.; Agerberth, B.; Lindbom, L. Neutrophil primary granule proteins HBP and HNP1–3 boost bacterial phagocytosis by human and murine macrophages. J. Clin. Investig. 2008, 118, 3491–3502. [Google Scholar] [CrossRef]
- Quinn, K.L.; Henriques, M.; Tabuchi, A.; Han, B.; Yang, H.; Cheng, W.E.; Tole, S.; Yu, H.; Luo, A.; Charbonney, E.; et al. Human neutrophil peptides mediate endothelial-monocyte interaction, foam cell formation, and platelet activation. Arter. Thromb. Vasc. Biol. 2011, 31, 2070–2079. [Google Scholar] [CrossRef]
- Crespo, J.; Fern, P.; Hern, M.; Mayorga, M.; Pons-Romero, F. Gene expression of tumor necrosis factor alpha and TNF receptors, p55 and p75, in nonalcoholic steatohepatitis patients. Hepatology 2001, 34, 1158–1163. [Google Scholar] [CrossRef]
- Potoupni, V.; Georgiadou, M.; Chatzigriva, E.; Polychronidou, G.; Markou, E.; Zapantis Gakis, C.; Filimidou, I.; Karagianni, M.; Anastasilakis, D.; Evripidou, K.; et al. Circulating tumor necrosis factor-α levels in non-alcoholic fatty liver disease: A systematic review and a meta-analysis. J. Gastroenterol. Hepatol. 2021, 36, 3002–3014. [Google Scholar] [CrossRef]
- Zhao, S.; Jiang, J.; Jing, Y.; Liu, W.; Yang, X.; Hou, X.; Gao, L.; Wei, L. The concentration of tumor necrosis factor-α determines its protective or damaging effect on liver injury by regulating Yap activity. Cell Death Dis. 2020, 11, 70. [Google Scholar] [CrossRef] [PubMed]
- Golabi, P.; Paik, J.; Fukui, N.; Locklear, C.T.; de Avilla, L.; Younossi, Z.M. Patients With Lean Nonalcoholic Fatty Liver Disease Are Metabolically Abnormal and Have a Higher Risk for Mortality. Clin. Diabetes 2019, 37, 65–72. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maraga, E.; Safadi, R.; Amer, J.; Higazi, A.A.-r.; Fanne, R.A. Alleviation of Hepatic Steatosis by Alpha-Defensin Is Associated with Enhanced Lipolysis. Medicina 2023, 59, 983. https://doi.org/10.3390/medicina59050983
Maraga E, Safadi R, Amer J, Higazi AA-r, Fanne RA. Alleviation of Hepatic Steatosis by Alpha-Defensin Is Associated with Enhanced Lipolysis. Medicina. 2023; 59(5):983. https://doi.org/10.3390/medicina59050983
Chicago/Turabian StyleMaraga, Emad, Rifaat Safadi, Johnny Amer, Abd Al-roof Higazi, and Rami Abu Fanne. 2023. "Alleviation of Hepatic Steatosis by Alpha-Defensin Is Associated with Enhanced Lipolysis" Medicina 59, no. 5: 983. https://doi.org/10.3390/medicina59050983
APA StyleMaraga, E., Safadi, R., Amer, J., Higazi, A. A.-r., & Fanne, R. A. (2023). Alleviation of Hepatic Steatosis by Alpha-Defensin Is Associated with Enhanced Lipolysis. Medicina, 59(5), 983. https://doi.org/10.3390/medicina59050983