
Suppression of UVB-Induced MMP-1 Expression in Human Skin Fibroblasts Using Lysate of Lactobacillus iners Derived from Korean Women’s Skin in Their Twenties

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
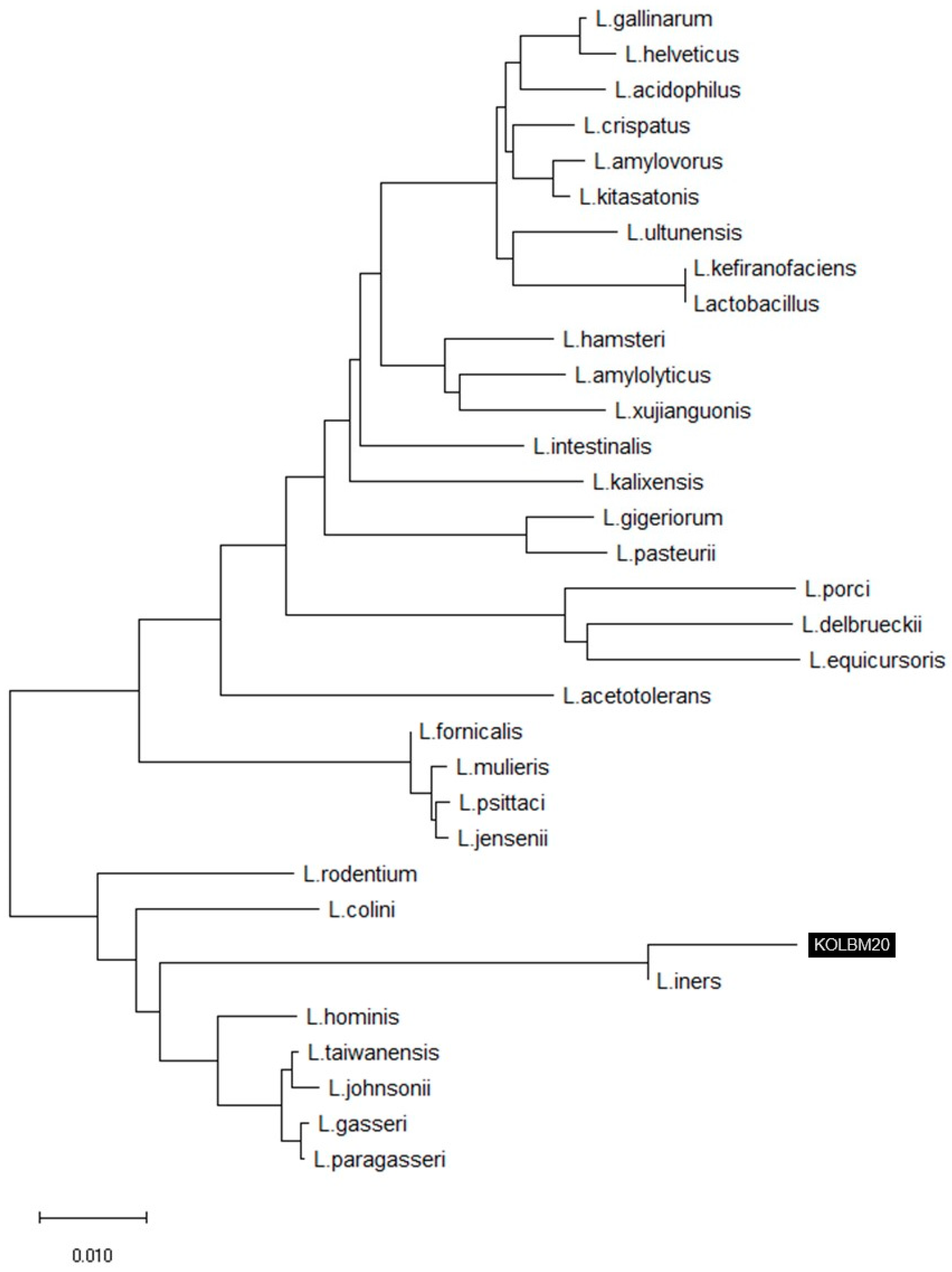
2.2. 16S rRNA Gene Sequence and Phylogenetic Analysis
2.3. Preparation of Lactobacillus Iners KOLBM20 Lysate
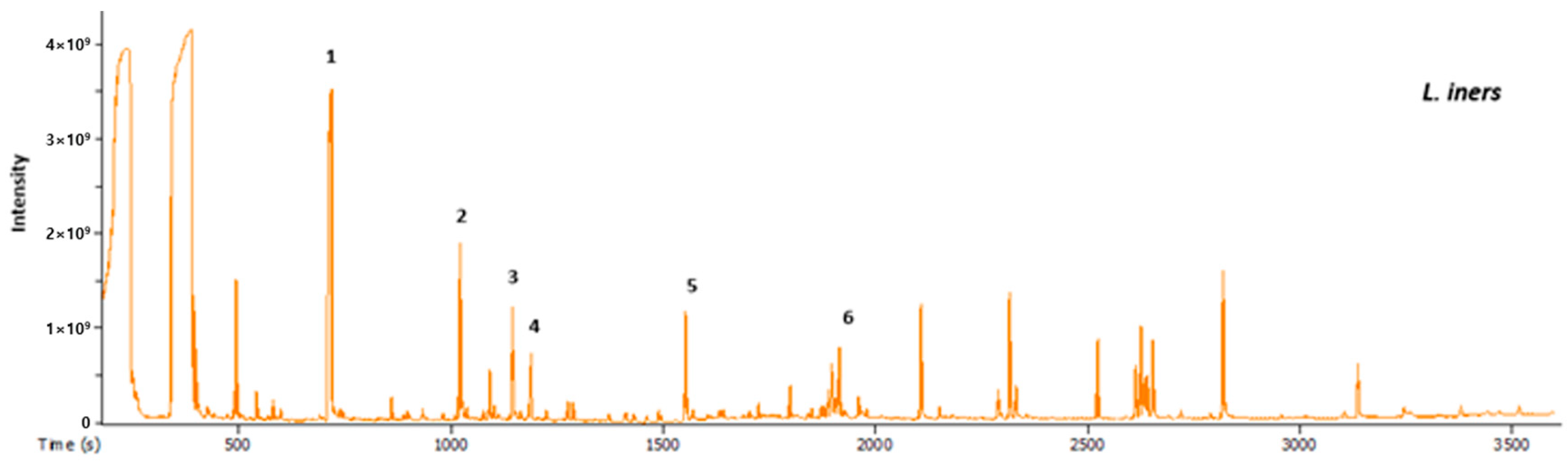
2.4. GC-MS Analysis of Extracellular Metabolites
2.5. Cell Cultures and Viability Assay
2.6. Ultraviolet Irradiation
2.7. Cytotoxicity
2.8. Enzyme-Linked Immunosorbent Assay (ELISA) for MMP-1 and Type I Collagen Expression
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
3.1. 16S rRNA Gene Sequence and Phylogenetic Analysis
3.2. Metabolomics Analysis by GC-MS
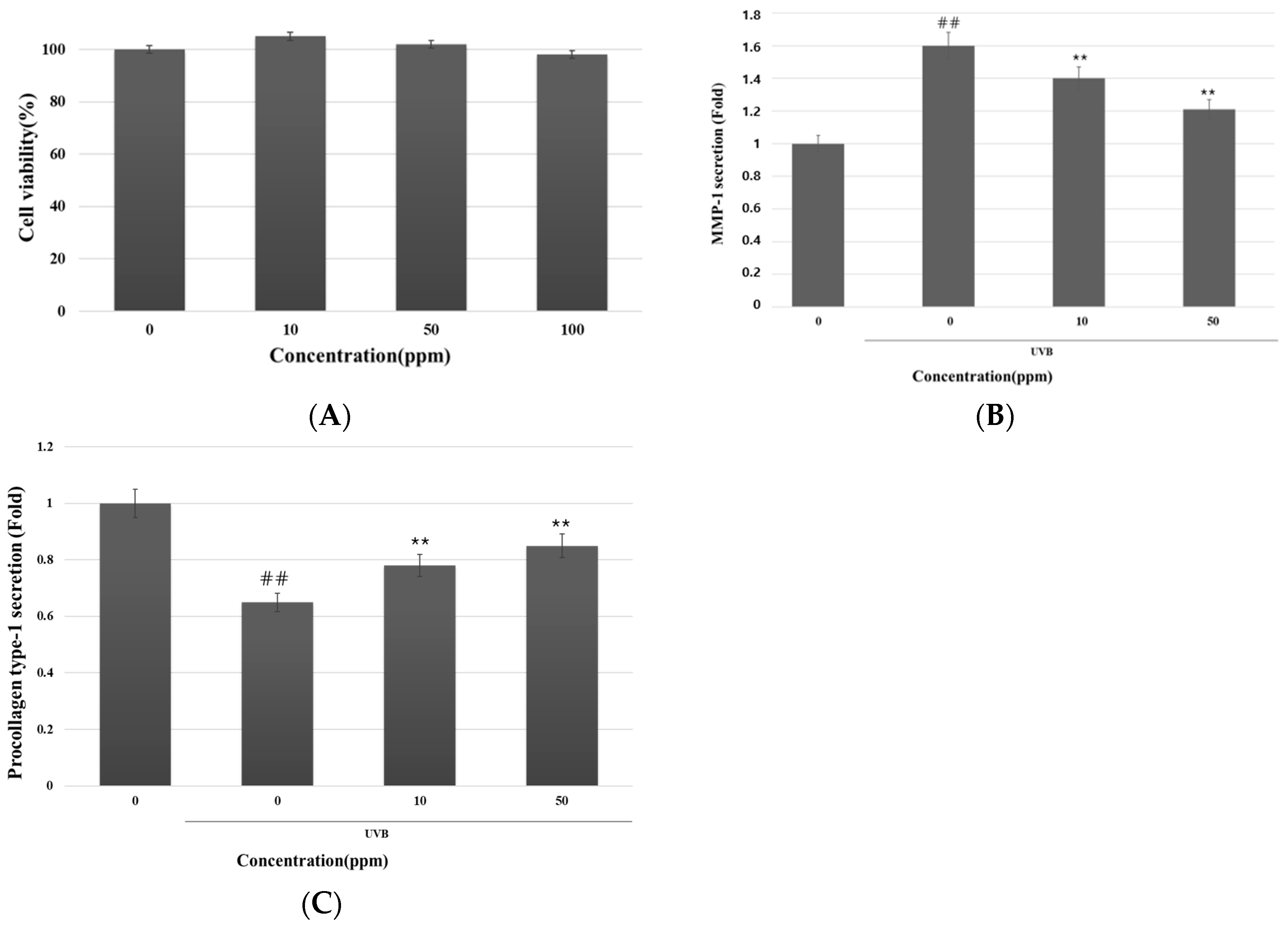
3.3. Inhibitory Effect of Strain KOLBM20 Lysate on MMP-1 Secretion and Type I Procollagen Degradation in UVB-Stimulated Dermal Fibroblasts
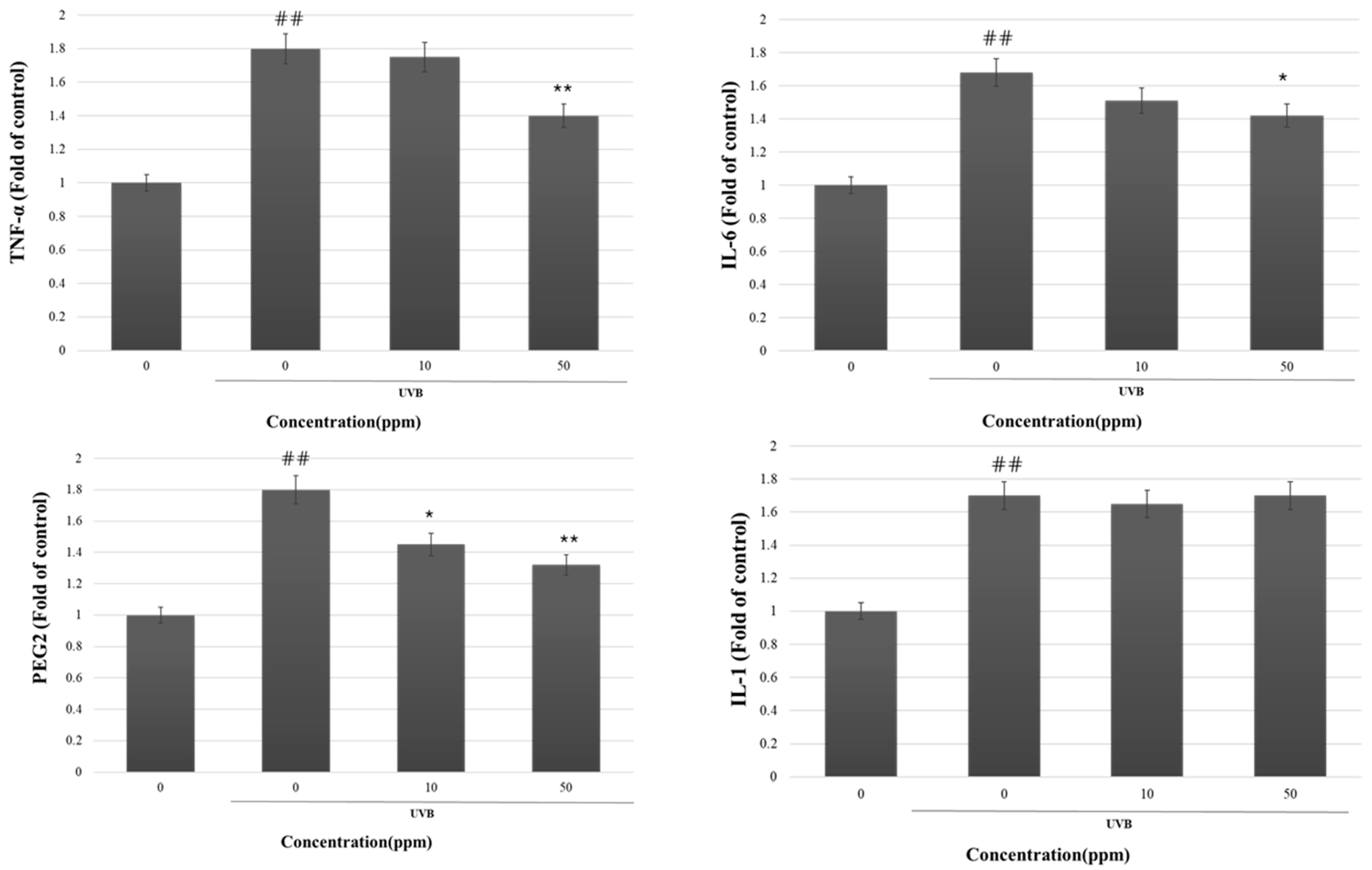
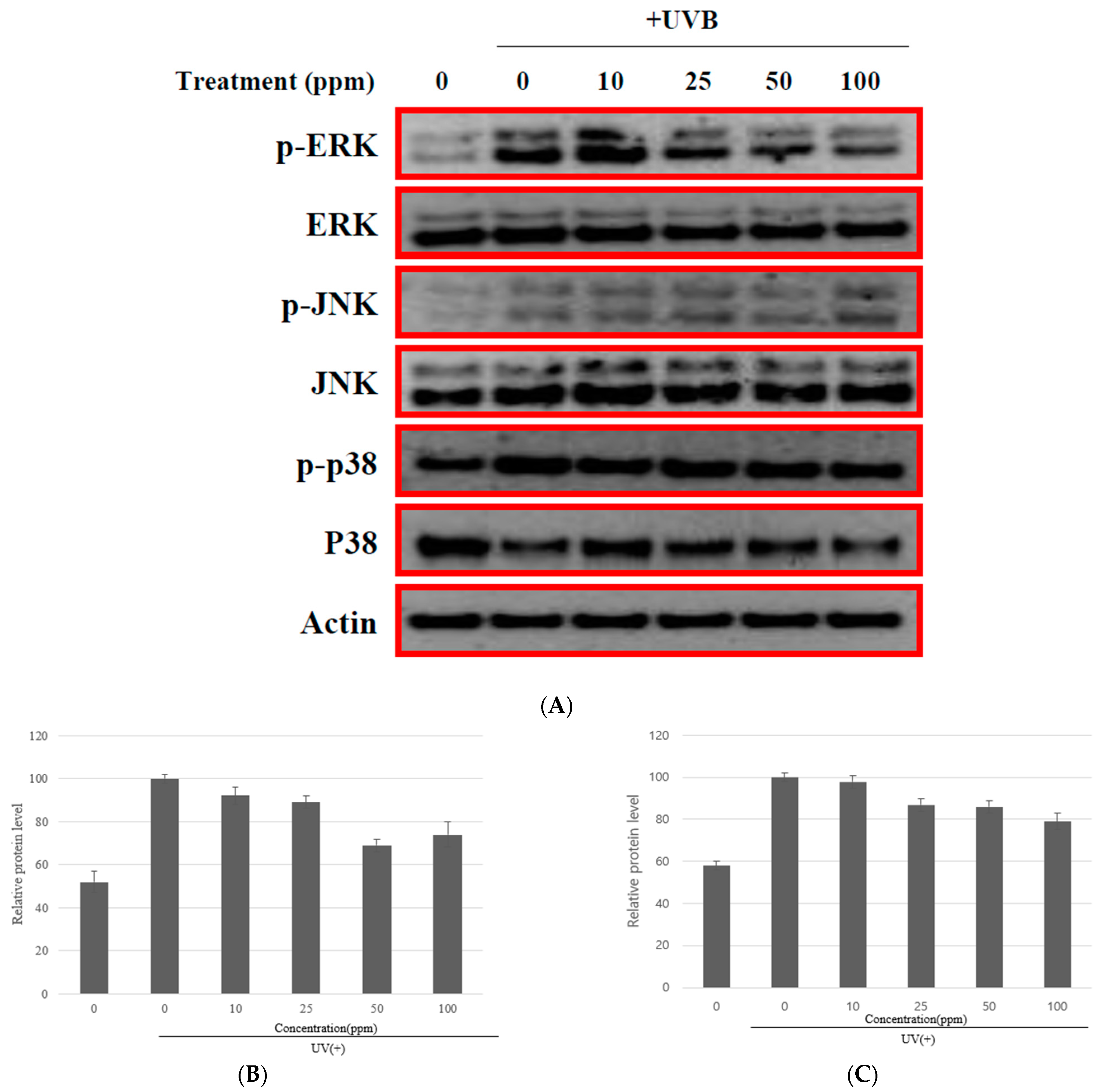
3.4. The MMP-1 Inhibition by Strain KOLBM20 Lysate through the Suppression of Signalling Pathway
4. Discussion
5. Conclusions
- Strain KOLBM20 was identified as a member of the Lactobacillus genus and found to have a close genetic affinity with L. iners DSM13335(T), sharing a 99.20% similarity in 16S rRNA gene sequences.
- We also confirmed the anti-aging properties of strain KOLBM20 lysate, specifically its potential to inhibit MMP-1 expression.
- While previous research has shown that Lactobacillus species can stimulate dermal fibroblast proliferation and enhance extracellular matrix deposition in vitro, the specific impact of Lactobacillus iners strain KOLBM20 on MMP-1 expression had not been previously explored.
- Our research revealed that strain KOLBM20 lysate effectively reduced the expression of MMP-1 induced by UVB irradiation in in vitro experiments. This inhibitory effect persisted over time, possibly due to the presence of uracil in KOLBM20, a compound known to inhibit MMP-1 expression.
- We identified the involvement of the ERK and JNK signaling pathways in UVB-induced MMP-1 expression. Strain KOLBM20 lysate was found to suppress these pathways, suggesting a potential mechanism for its inhibition of MMP-1 expression.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- El-Domyati, M.; Attia, S.; Saleh, F.; Brown, D.; Birk, D.E.; Gasparro, F.; Ahmad, H.; Uitto, J. Intrinsic aging vs. photoaging: A comparative histopathological, immunohistochemical, and ultrastructural study of skin. Exp. Dermatol. 2002, 11, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Cavinato, M.; Jansen-Durr, P. Molecular mechanisms of UVB-induced senescence of dermal fibroblasts and its relevance for photoaging of the human skin. Exp. Gerontol. 2017, 94, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Gilchrest, B.A. Photoaging. J. Inv. Dermatol. 2013, 133, E2–E6. [Google Scholar] [CrossRef] [PubMed]
- Scharffetter-Kochanek, K.; Brenneisen, P.; Wenk, J.; Herrmann, G.; Ma, W.; Kuhr, L.; Meewes, C.; Wlaschek, M. Photoaging of the skin from phenotype to mechanisms. Exp. Gerontol. 2000, 35, 307–316. [Google Scholar] [CrossRef]
- Kang, S.; Fisher, G.J.; Voorhees, J.J. Photoaging and topical tretinoin: Therapy, pathogenesis, and prevention. Arch. Dermatol. 1997, 133, 1280–1284. [Google Scholar] [CrossRef] [PubMed]
- Kligman, A.M. Early destructive effect of sunlight on human skin. JAMA 1969, 210, 2377–2380. [Google Scholar] [CrossRef]
- Vierkotter, A.; Huls, A.; Yamamoto, A.; Stolz, S.; Kramer, U.; Matsui, M.S.; Morita, A.; Wang, S.; Li, Z.; Jin, L.; et al. Extrinsic skin ageing in German, Chinese and Japanese women manifests differently in all three groups depending on ethnic background, age and anatomical site. J. Dermatol. Sci. 2016, 83, 219–225. [Google Scholar] [CrossRef]
- Shah, H.; Rawal Mahajan, S. Photoaging: New insights into its stimulators, complications, biochemical changes and therapeutic interventions. Biomed. Aging Pathol. 2013, 3, 161–169. [Google Scholar] [CrossRef]
- Goukassian, D.A.; Gilchrest, B.A. The interdependence of skin aging, skin cancer, and DNA repair capacity: A novel perspective with therapeutic implications. Rejuvenation Res. 2004, 7, 175–185. [Google Scholar] [CrossRef]
- Berardesca, E.; Maibach, H. Racial differences in skin pathophysiology. J. Am. Acad. Dermatol. 1996, 34, 667–672. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chiang, H.I.; Jiang, S.B.; Nagarajan, H.; Zengler, K.; Gallo, R.L. The microbiome extends to subepidermal compartments of normal skin. Nat. Commun. 2013, 4, 1431. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.; Garges, S.; Giovanni, M.; McInnes, P.; Wang, L.; Schloss, J.A.; Bonazzi, V.; McEwen, J.E.; Wetterstrand, K.A.; Deal, C.; et al. The NIH Human Microbiome Project. Genome Res. 2009, 19, 2317–2323. [Google Scholar] [CrossRef] [PubMed]
- Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Rijkers, G.T.; de Vos, W.M.; Brummer, R.J.; Morelli, L.; Corthier, G.; Marteau, P. Health benefits and health claims of probiotics: Bridging science and marketing. Br. J. Nutr. 2011, 106, 1291–1296. [Google Scholar] [CrossRef] [PubMed]
- Reid, G.; Jass, J.; Sebulsky, M.T.; McCormick, J.K. Potential uses of probiotics in clinical practice. Clin. Microbiol. Rev. 2003, 16, 658–672. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, E.; Park, Y.M.; Hong, S.J. Microbiome in the Gut-Skin Axis in Atopic Dermatitis. Allergy Asthma Immunol. Res. 2018, 10, 354–362. [Google Scholar] [CrossRef]
- Lim, H.Y.; Jeong, D.; Park, S.H.; Shin, K.K.; Hong, Y.H.; Kim, E.; Yu, Y.G.; Kim, T.R.; Kim, H.; Lee, J.; et al. Antiwrinkle and Antimelanogenesis Effects of Tyndallized Lactobacillus acidophilus KCCM12625P. Int. J. Mol. Sci. 2020, 21, 1620. [Google Scholar] [CrossRef]
- Teng, Y.; Huang, Y.; Danfeng, X.; Tao, X.; Fan, Y. The Role of Probiotics in Skin Photoaging and Related Mechanisms: A Review. Clin. Cosmet. Investig. Dermatol. 2022, 15, 2455–2464. [Google Scholar] [CrossRef]
- You, G.E.; Jung, B.J.; Kim, H.R.; Kim, H.G.; Kim, T.R.; Chung, D.K. Lactobacillus sakei lipoteichoic acid inhibits MMP-1 induced by UVA in normal dermal fibroblasts of human. J. Microbiol. Biotechnol. 2013, 23, 1357–1364. [Google Scholar] [CrossRef]
- Bowe, W.P.; Logan, A.C. Acne vulgaris, probiotics and the gut-brain-skin axis—back to the future? Gut Pathog. 2011, 3, 1. [Google Scholar] [CrossRef]
- Romagnani, S. Coming back to a missing immune deviation as the main explanatory mechanism for the hygiene hypothesis. J. Allergy Clin. Immunol. 2007, 119, 1511–1513. [Google Scholar] [CrossRef] [PubMed]
- Fredricks, D.N. Microbial ecology of human skin in health and disease. J. Investig. Dermatol. Symp. Proc. 2001, 6, 167–169. [Google Scholar] [CrossRef]
- Im, A.R.; Lee, B.; Kang, D.J.; Chae, S. Protective effects of tyndallized Lactobacillus acidophilus IDCC 3302 against UVB-induced photodamage to epidermal keratinocytes cells. Int. J. Mol. Med. 2019, 43, 2499–2506. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Lee, Y.; Huang, Y.H.; Lim, H.W.; Jang, K.; Kim, D.D.; Lim, C.J. Probiotic fermentation augments the skin anti-photoaging properties of Agastache rugosa through up-regulating antioxidant components in UV-B-irradiated HaCaT keratinocytes. BMC Complement. Altern. Med. 2018, 18, 196. [Google Scholar] [CrossRef] [PubMed]
- Khmaladze, I.; Butler, E.; Fabre, S.; Gillbro, J.M. Lactobacillus reuteri DSM 17938-A comparative study on the effect of probiotics and lysates on human skin. Exp. Dermatol. 2019, 28, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.B.; Jeong, M.; Han, K.S.; Hwan Kim, J.; Park, Y.; Suh, H.J. Photoprotective effects of galacto-oligosaccharide and/or Bifidobacterium longum supplementation against skin damage induced by ultraviolet irradiation in hairless mice. Int. J. Food Sci. Nutr. 2015, 66, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Habeebuddin, M.; Karnati, R.K.; Shiroorkar, P.N.; Nagaraja, S.; Asdaq, S.M.B.; Khalid Anwer, M.; Fattepur, S. Topical Probiotics: More Than a Skin Deep. Pharmaceutics 2022, 14, 557. [Google Scholar] [CrossRef]
- Sharma, D.; Kober, M.M.; Bowe, W.P. Anti-Aging Effects of Probiotics. J. Drugs Dermatol. 2016, 15, 9–12. [Google Scholar]
- Kang, B.S.; Seo, J.G.; Lee, G.S.; Kim, J.H.; Kim, S.Y.; Han, Y.W.; Kang, H.; Kim, H.O.; Rhee, J.H.; Chung, M.J.; et al. Antimicrobial activity of enterocins from Enterococcus faecalis SL-5 against Propionibacterium acnes, the causative agent in acne vulgaris, and its therapeutic effect. J. Microbiol. 2009, 47, 101–109. [Google Scholar] [CrossRef]
- Notay, M.; Saric-Bosanac, S.; Vaughn, A.R.; Dhaliwal, S.; Trivedi, M.; Reiter, P.N.; Rybak, I.; Li, C.C.; Weiss, L.B.; Ambrogio, L.; et al. The use of topical Nitrosomonas eutropha for cosmetic improvement of facial wrinkles. J. Cosmet. Dermatol. 2020, 19, 689–693. [Google Scholar] [CrossRef]
- Howard, B.; Bascom, C.C.; Hu, P.; Binder, R.L.; Fadayel, G.; Huggins, T.G.; Jarrold, B.B.; Osborne, R.; Rocchetta, H.L.; Swift, D.; et al. Aging-Associated Changes in the Adult Human Skin Microbiome and the Host Factors that Affect Skin Microbiome Composition. J. Inv. Dermatol. 2022, 142, 1934–1946.e1921. [Google Scholar] [CrossRef] [PubMed]
- Jo, C.S.; Myung, C.H.; Yoon, Y.C.; Ahn, B.H.; Min, J.W.; Seo, W.S.; Lee, D.H.; Kang, H.C.; Heo, Y.H.; Choi, H.; et al. The Effect of Lactobacillus plantarum Extracellular Vesicles from Korean Women in Their 20s on Skin Aging. Curr. Issues Mol. Biol. 2022, 44, 526–540. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.C.; Kim, K.M.; Lee, S.H.; Kim, Y.H.; Lee, S.Y.; Choi, D.H.; Choi, J.W.; Lee, S.H. Inhibitory Effect of Novel Lactobacillus paracasei SKB1192 on Skin Photoaging. J. K. Soc. Cosm. 2019, 25, 591–599. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Oxford University Press: Oxford, UK, 1999; pp. 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. The Neutral Theory of Molecular Evolution; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tamura, K.; Jakobsen, I.B.; Nei, M. MEGA2: Molecular evolutionary genetics analysis software. Bioinformatics 2001, 17, 1244–1245. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Park, B.; Hwang, H.; Chang, J.Y.; Hong, S.W.; Lee, S.H.; Jung, M.Y.; Sohn, S.O.; Park, H.W.; Lee, J.H. Identification of 2-hydroxyisocaproic acid production in lactic acid bacteria and evaluation of microbial dynamics during kimchi ripening. Sci. Rep. 2017, 7, 10904. [Google Scholar] [CrossRef]
- Kim, S.R.; Jung, Y.R.; An, H.J.; Kim, D.H.; Jang, E.J.; Choi, Y.J.; Moon, K.M.; Park, M.H.; Park, C.H.; Chung, K.W.; et al. Anti-wrinkle and anti-inflammatory effects of active garlic components and the inhibition of MMPs via NF-κB signaling. PLoS ONE 2013, 8, e73877. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-J.; Dai, R.-Y.; Leu, Y.-L.; Tsai, T.-Y. Effects of the melanogenic inhibitor, uracil, derived from Lactobacillus plantarum TWK10-fermented soy milk on anti-melanogenesis in B16F0 mouse melanoma cells. J. Funct. Foods 2015, 17, 314–327. [Google Scholar] [CrossRef]
- Yun, H.J.; Yoo, W.H.; Han, M.K.; Lee, Y.R.; Kim, J.S.; Lee, S.I. Epigallocatechin-3-gallate suppresses TNF-alpha -induced production of MMP-1 and -3 in rheumatoid arthritis synovial fibroblasts. Rheumatol. Int. 2008, 29, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Fushimi, K.; Nakashima, S.; You, F.; Takigawa, M.; Shimizu, K. Prostaglandin E2 downregulates TNF-alpha-induced production of matrix metalloproteinase-1 in HCS-2/8 chondrocytes by inhibiting Raf-1/MEK/ERK cascade through EP4 prostanoid receptor activation. J. Cell Biochem. 2007, 100, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M.; Lee, I.T.; Lin, C.C.; Wang, C.H.; Cherng, W.J.; Hsiao, L.D. c-Src-dependent MAPKs/AP-1 activation is involved in TNF-α-induced matrix metalloproteinase-9 expression in rat heart-derived H9c2 cells. Biochem. Pharmacol. 2013, 85, 1115–1123. [Google Scholar] [CrossRef] [PubMed]
- Ziv, E.; Rotem, C.; Miodovnik, M.; Ravid, A.; Koren, R. Two modes of ERK activation by TNF in keratinocytes: Different cellular outcomes and bi-directional modulation by vitamin D. J. Cell Biochem. 2008, 104, 606–619. [Google Scholar] [CrossRef]
- Brenneisen, P.; Sies, H.; Scharffetter-Kochanek, K. Ultraviolet-B irradiation and matrix metalloproteinases: From induction via signaling to initial events. Ann. N. Y. Acad. Sci. 2002, 973, 31–43. [Google Scholar] [CrossRef]
- Thornfeldt, C.R. Chronic inflammation is etiology of extrinsic aging. J. Cosmet. Dermatol. 2008, 7, 78–82. [Google Scholar] [CrossRef]
- Yoshizumi, M.; Nakamura, T.; Kato, M.; Ishioka, T.; Kozawa, K.; Wakamatsu, K.; Kimura, H. Release of cytokines/chemokines and cell death in UVB-irradiated human keratinocytes, HaCaT. Cell Biol. Int. 2008, 32, 1405–1411. [Google Scholar] [CrossRef]
- Miller, C.C.; Hale, P.; Pentland, A.P. Ultraviolet B injury increases prostaglandin synthesis through a tyrosine kinase-dependent pathway. Evidence for UVB-induced epidermal growth factor receptor activation. J. Biol. Chem. 1994, 269, 3529–3533. [Google Scholar] [CrossRef]
- Pająk, J.; Nowicka, D.; Szepietowski, J.C. Inflammaging and Immunosenescence as Part of Skin Aging-A Narrative Review. Int. J. Mol. Sci. 2023, 24, 7784. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-S.; Min, J.-W.; Gye, S.-B.; Kim, Y.-W.; Kang, H.-C.; Choi, Y.-S.; Seo, W.-S.; Lee, B.-Y. Suppression of UVB-Induced MMP-1 Expression in Human Skin Fibroblasts Using Lysate of Lactobacillus iners Derived from Korean Women’s Skin in Their Twenties. Curr. Issues Mol. Biol. 2024, 46, 513-526. https://doi.org/10.3390/cimb46010033
Lee J-S, Min J-W, Gye S-B, Kim Y-W, Kang H-C, Choi Y-S, Seo W-S, Lee B-Y. Suppression of UVB-Induced MMP-1 Expression in Human Skin Fibroblasts Using Lysate of Lactobacillus iners Derived from Korean Women’s Skin in Their Twenties. Current Issues in Molecular Biology. 2024; 46(1):513-526. https://doi.org/10.3390/cimb46010033
Chicago/Turabian StyleLee, Jin-Sung, Jin-Woo Min, Seong-Bong Gye, Yong-Woo Kim, Hee-Cheol Kang, Yoon-Seo Choi, Won-Sang Seo, and Bun-Yeoul Lee. 2024. "Suppression of UVB-Induced MMP-1 Expression in Human Skin Fibroblasts Using Lysate of Lactobacillus iners Derived from Korean Women’s Skin in Their Twenties" Current Issues in Molecular Biology 46, no. 1: 513-526. https://doi.org/10.3390/cimb46010033
APA StyleLee, J.-S., Min, J.-W., Gye, S.-B., Kim, Y.-W., Kang, H.-C., Choi, Y.-S., Seo, W.-S., & Lee, B.-Y. (2024). Suppression of UVB-Induced MMP-1 Expression in Human Skin Fibroblasts Using Lysate of Lactobacillus iners Derived from Korean Women’s Skin in Their Twenties. Current Issues in Molecular Biology, 46(1), 513-526. https://doi.org/10.3390/cimb46010033