
TRPV4 Activation during Guinea Pig Airway Smooth Muscle Contraction Promotes Ca2+ and Na+ Influx
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Intracellular Ca2+ Increase Mediated by TRPV4 Activation with GSK101 Depends Exclusively on External Ca2+ Influx
2.2. Pharmacological TRPV4 Activation with GSK101 Mediates Extracellular Na+ Influx
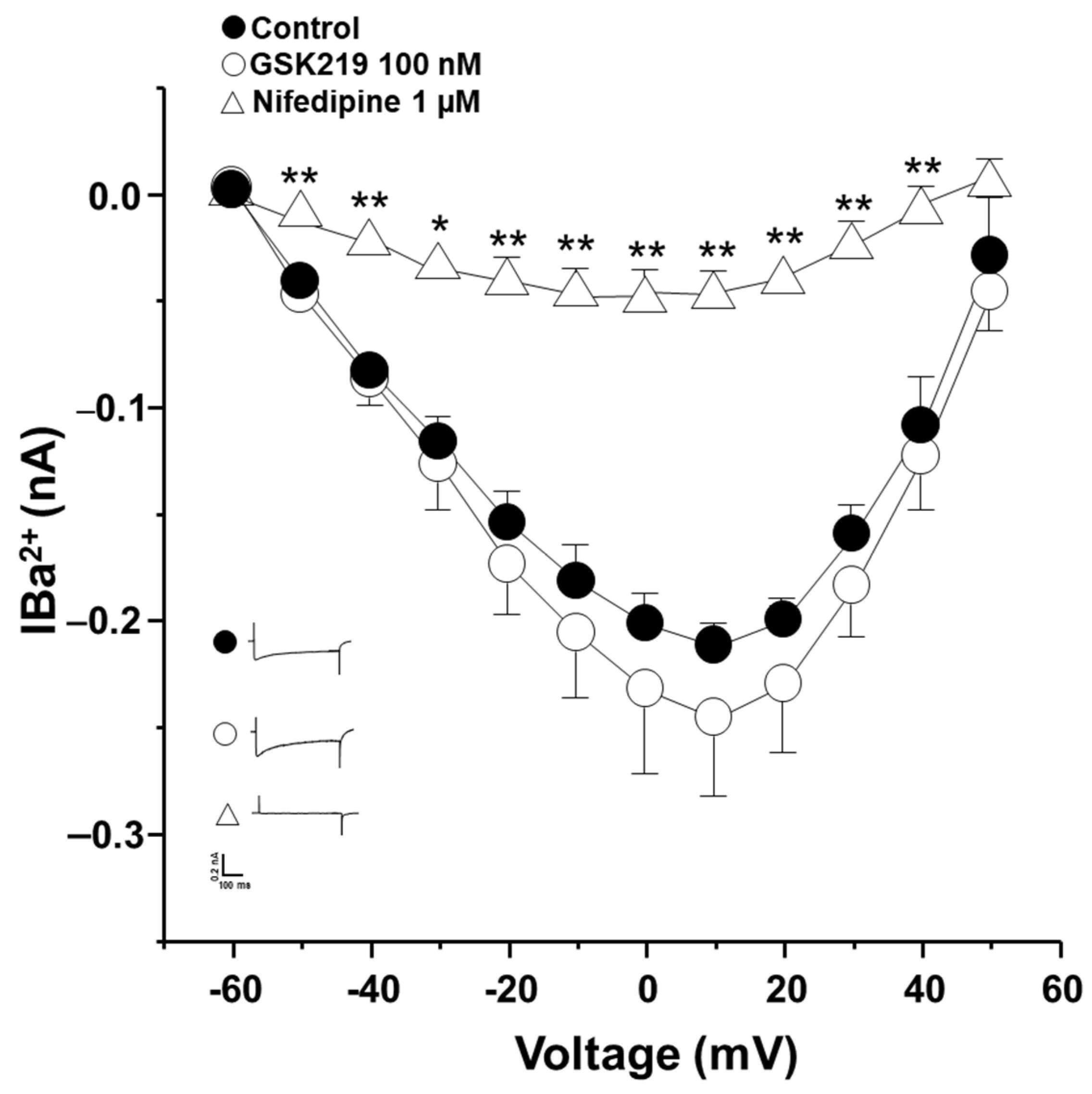
2.3. L-VDCCs Are Involved in Airway Smooth Muscle Contraction Mediated by TRPV4 Stimulation
2.4. TRPV4 Blockade Decreases Airway Smooth Muscle Reactivity to Carbachol and Histamine
2.5. TRPV4 Is Involved in Intracellular Ca2+ Increases Induced by Carbachol and Histamine
2.6. TRPV4 Blockade Also Lowers Smooth Muscle Reactivity to KCl
2.7. Role of TRPV4 in Sarcoplasmic Reticulum Ca2+ Refilling
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Intracellular Ca2+ Measurements in Tracheal Myocytes
4.3. Organ Baths
4.4. Patch-Clamp Recordings
4.5. Intracellular Na+ Measurements in Tracheal Myocytes
4.6. Western Blotting for TRPV4
4.7. Drugs and Chemicals
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lam, M.; Lamanna, E.; Bourke, J.E. Regulation of airway smooth muscle contraction in health and disease. Adv. Exp. Med. Biol. 2019, 1124, 381–422. [Google Scholar] [CrossRef]
- Abohalaka, R. Bronchial epithelial and airway smooth muscle cell interactions in health and disease. Heliyon 2023, 9, e19976. [Google Scholar] [CrossRef]
- Shetty, S.M.; Archana, S. Anatomy and Physiology of the Airway Applied Aspects; Ubaradka, R.S., Gupta, N., Bidkar, P.U., Tripathy, D.K., Gupta, A., Eds.; The Airway Manual; Springer: Singapore, 2023; pp. 21–43. [Google Scholar] [CrossRef]
- Montaño, L.M.; Flores-Soto, E.; Reyes-García, J.; Solis-Chagoyán, H.; Sommer, B. Airway smooth muscle functioning in basal, agonists stimulated conditions and novel androgen asthma therapy. Adv. Med. Biol. 2020, 157, 1–64. [Google Scholar]
- Xiong, D.J.P.; Martin, J.G.; Lauzon, A.M. Airway smooth muscle function in asthma. Front. Physiol. 2022, 13, 993406. [Google Scholar] [CrossRef]
- Yang, Q.; Miao, Q.; Chen, H.; Li, D.; Luo, Y.; Chiu, J.; Wang, H.J.; Chuvanjyan, M.; Parmacek, M.S.; Shi, W. Myocd regulates airway smooth muscle cell remodeling in response to chronic asthmatic injury. J. Pathol. 2023, 259, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Hongping, H.; Chufang, L.; Cuomu, M.; Jintao, L.; Kaiyin, C.; Lvyi, C.; Weiwu, C.; Zuguang, Y.; Nanshan, Z. Effects of Shiwei Longdanhua formula on LPS induced airway mucus hypersecretion, cough hypersensitivity, oxidative stress and pulmonary inflammation. Biomed. Pharmacother. 2023, 163, 114793. [Google Scholar] [CrossRef] [PubMed]
- Yeganeh, B.; Xia, C.; Movassagh, H.; Koziol-White, C.; Chang, Y.; Al-Alwan, L.; Bourke, J.E.; Oliver, B.G. Emerging mediators of airway smooth muscle dysfunction in asthma. Pulm. Pharmacol. Ther. 2013, 26, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Miki, H.; Kiosses, W.B.; Manresa, M.C.; Gupta, R.K.; Sethi, G.S.; Herro, R.; Da Silva Antunes, R.; Dutta, P.; Miller, M.; Fung, K.; et al. Lymphotoxin beta receptor signaling directly controls airway smooth muscle deregulation and asthmatic lung dysfunction. J. Allergy Clin. Immunol. 2023, 151, 976–990.e5. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Liu, S.; Ye, D.; Bai, G.; Guo, M.; Sun, R.; Ran, P. PM2.5 induces lung inflammation and fibrosis via airway smooth muscle cell expression of the Wnt5a/JNK pathway. J. Thorac. Dis. 2023, 15, 6094–6105. [Google Scholar] [CrossRef] [PubMed]
- Kume, H.; Yamada, R.; Sato, Y.; Togawa, R. Airway smooth muscle regulated by oxidative stress in COPD. Antioxidants 2023, 12, 142. [Google Scholar] [CrossRef]
- Wang, R.; Li, W.; Dai, H.; Zhu, M.; Li, L.; Si, G.; Bai, Y.; Wu, H.; Hu, X.; Xing, Y. PKD1 deficiency induces Bronchiectasis in a porcine ADPKD model. Respir. Res. 2022, 23, 292. [Google Scholar] [CrossRef]
- Gebski, E.B.; Anaspure, O.; Panettieri, R.A.; Koziol-White, C.J. Airway smooth muscle and airway hyperresponsiveness in asthma: Mechanisms of airway smooth muscle dysfunction. Minerva Med. 2022, 113, 4–16. [Google Scholar] [CrossRef]
- An, S.S.; Mitzner, W.; Tang, W.Y.; Ahn, K.; Yoon, A.R.; Huang, J.; Kilic, O.; Yong, H.M.; Fahey, J.W.; Kumar, S.; et al. An inflammation-independent contraction mechanophenotype of airway smooth muscle in asthma. J. Allergy Clin. Immunol. 2016, 138, 294–297.e4. [Google Scholar] [CrossRef]
- Yan, F.; Gao, H.; Zhao, H.; Bhatia, M.; Zeng, Y. Roles of airway smooth muscle dysfunction in chronic obstructive pulmonary disease. J. Transl. Med. 2018, 16, 262. [Google Scholar] [CrossRef] [PubMed]
- Alhamdan, F.; Marsh, L.M.; Pedersen, F.; Alhamwe, B.A.; Tholken, C.; Pfefferle, P.I.; Bahmer, T.; Greulich, T.; Potaczek, D.P.; Garn, H. Differential Regulation of Interferon Signaling Pathways in CD4+ T Cells of the Low Type-2 Obesity-Associated Asthma Phenotype. Int. J. Mol. Sci. 2021, 22, 10144. [Google Scholar] [CrossRef] [PubMed]
- Mazerik, J.; Gondas, E.; Smiesko, L.; Franova, S.; Sutovska, M. Effects of TRPV4 channel blocker on airway inflammation and airway defense reflexes in experimentally induced model of allergic asthma. Respir. Physiol. Neurobiol. 2023, 316, 104123. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, X.; Yang, X.; Liu, X.; Zeng, Y.; Li, J. Melatonin antagonizes ozone-exacerbated asthma by inhibiting the TRPV1 channel and stabilizing the Nrf2 pathway. Environ. Sci. Pollut. Res. Int. 2021, 28, 59858–59867. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Cheng, M.; Li, M.; Wang, T.; Wen, F.; Sanderson, M.J.; Sneyd, J.; Shen, Y.; Chen, J. Inherent differences of small airway contraction and Ca2+ oscillations in airway smooth muscle cells between BALB/c and C57BL/6 mouse strains. Front. Cell Dev. Biol. 2023, 11, 1202573. [Google Scholar] [CrossRef] [PubMed]
- Gerthoffer, W.T. Regulation of the contractile element of airway smooth muscle. Am. J. Physiol. 1991, 261, L15–L28. [Google Scholar] [CrossRef]
- Somlyo, A.P.; Somlyo, A.V. Signal transduction and regulation in smooth muscle. Nature 1994, 372, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Kitazawa, T.; Matsui, T.; Katsuki, S.; Goto, A.; Akagi, K.; Hatano, N.; Tokumitsu, H.; Takeya, K.; Eto, M. A temporal Ca2+ desensitization of myosin light chain kinase in phasic smooth muscles induced by CaMKKβ/PP2A pathways. Am. J. Physiol. Cell Physiol. 2021, 321, C549–C558. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Santos, M.D.; Alvarez-Gonzalez, M.; Estrada-Soto, S.; Bazan-Perkins, B. Regulation of myosin light-chain phosphatase activity to generate airway smooth muscle hypercontractility. Front. Physiol. 2020, 11, 701. [Google Scholar] [CrossRef] [PubMed]
- Gazzola, M.; Henry, C.; Lortie, K.; Khadangi, F.; Park, C.Y.; Fredberg, J.J.; Bosse, Y. Airway smooth muscle tone increases actin filamentogenesis and contractile capacity. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 318, L442–L451. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Wang, L.; Seow, C.Y.; Ikebe, M. p116(Rip) promotes myosin phosphatase activity in airway smooth muscle cells. J. Cell Physiol. 2020, 235, 114–127. [Google Scholar] [CrossRef] [PubMed]
- Reyes-García, J.; Flores-Soto, E.; Carbajal-García, A.; Sommer, B.; Montaño, L.M. Maintenance of intracellular Ca2+ basal concentration in airway smooth muscle (Review). Int. J. Mol. Med. 2018, 42, 2998–3008. [Google Scholar] [CrossRef] [PubMed]
- Bourreau, J.P.; Abela, A.P.; Kwan, C.Y.; Daniel, E.E. Acetylcholine Ca2+ stores refilling directly involves a dihydropyridine-sensitive channel in dog trachea. Am. J. Physiol. 1991, 261, C497–C505. [Google Scholar] [CrossRef]
- Campos-Bedolla, P.; Vargas, M.H.; Segura, P.; Carbajal, V.; Calixto, E.; Figueroa, A.; Flores-Soto, E.; Barajas-López, C.; Mendoza-Patino, N.; Montaño, L.M. Airway smooth muscle relaxation induced by 5-HT2A receptors: Role of Na+/K+-ATPase pump and Ca2+-activated K+ channels. Life Sci. 2008, 83, 438–446. [Google Scholar] [CrossRef]
- Carbajal, V.; Vargas, M.H.; Flores-Soto, E.; Martínez-Cordero, E.; Bazán-Perkins, B.; Montaño, L.M. LTD4 induces hyperresponsiveness to histamine in bovine airway smooth muscle: Role of SR-ATPase Ca2+ pump and tyrosine kinase. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 288, L84–L92. [Google Scholar] [CrossRef]
- Montaño, L.M.; Flores-Soto, E.; Reyes-García, J.; Díaz-Hernández, V.; Carbajal-García, A.; Campuzano-González, E.; Ramirez-Salinas, G.L.; Velasco-Velazquez, M.A.; Sommer, B. Testosterone induces hyporesponsiveness by interfering with IP3 receptors in guinea pig airway smooth muscle. Mol. Cell Endocrinol. 2018, 473, 17–30. [Google Scholar] [CrossRef]
- Baron, C.B.; Cunningham, M.; Strauss, J.F., 3rd; Coburn, R.F. Pharmacomechanical coupling in smooth muscle may involve phosphatidylinositol metabolism. Proc. Natl. Acad. Sci. USA 1984, 81, 6899–6903. [Google Scholar] [CrossRef]
- Berridge, M.J. Inositol trisphosphate and calcium signalling. Nature 1993, 361, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Trice, J.; Shinde, P.; Willis, R.E.; Pressley, T.A.; Perez-Zoghbi, J.F. Ca2+ oscillations, Ca2+ sensitization, and contraction activated by protein kinase C in small airway smooth muscle. J. Gen. Physiol. 2013, 141, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Perusquía, M.; Flores-Soto, E.; Sommer, B.; Campuzano-González, E.; Martinez-Villa, I.; Martinez-Banderas, A.I.; Montaño, L.M. Testosterone-induced relaxation involves L-type and store-operated Ca2+ channels blockade, and PGE2 in guinea pig airway smooth muscle. Pflugers Arch. 2015, 467, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Flores-Soto, E.; Reyes-García, J.; Sommer, B.; Montaño, L.M. Sarcoplasmic reticulum Ca2+ refilling is determined by L-type Ca2+ and store operated Ca2+ channels in guinea pig airway smooth muscle. Eur. J. Pharmacol. 2013, 721, 21–28. [Google Scholar] [CrossRef]
- Pelaia, G.; Renda, T.; Gallelli, L.; Vatrella, A.; Busceti, M.T.; Agati, S.; Caputi, M.; Cazzola, M.; Maselli, R.; Marsico, S.A. Molecular mechanisms underlying airway smooth muscle contraction and proliferation: Implications for asthma. Respir. Med. 2008, 102, 1173–1181. [Google Scholar] [CrossRef]
- Dwivedi, R.; Drumm, B.T.; Griffin, C.S.; Dudem, S.; Bradley, E.; Alkawadri, T.; Martin, S.L.; Sergeant, G.P.; Hollywood, M.A.; Thornbury, K.D. Excitatory cholinergic responses in mouse primary bronchial smooth muscle require both Ca2+ entry via L-type Ca2+ channels and store operated Ca2+ entry via Orai channels. Cell Calcium 2023, 112, 102721. [Google Scholar] [CrossRef]
- Hirota, S.; Helli, P.; Janssen, L.J. Ionic mechanisms and Ca2+ handling in airway smooth muscle. Eur. Respir. J. 2007, 30, 114–133. [Google Scholar] [CrossRef]
- Liu, Q.H.; Zheng, Y.M.; Korde, A.S.; Yadav, V.R.; Rathore, R.; Wess, J.; Wang, Y.X. Membrane depolarization causes a direct activation of G protein-coupled receptors leading to local Ca2+ release in smooth muscle. Proc. Natl. Acad. Sci. USA 2009, 106, 11418–11423. [Google Scholar] [CrossRef]
- Flores-Soto, E.; Carbajal, V.; Reyes-García, J.; Garcia-Hernandez, L.M.; Figueroa, A.; Checa, M.; Barajas-López, C.; Montaño, L.M. In airways ATP refills sarcoplasmic reticulum via P2X smooth muscle receptors and induces contraction through P2Y epithelial receptors. Pflugers Arch. 2011, 461, 261–275. [Google Scholar] [CrossRef]
- Tan, H.; Lei, J.; Xue, L.; Cai, C.; Liu, Q.H.; Shen, J. Relaxing effect of TSU-68, an antiangiogenic agent, on mouse airway smooth muscle. Cell Physiol. Biochem. 2017, 41, 2350–2362. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xue, L.; Zhao, Q.; Cai, C.; Liu, Q.H.; Shen, J. Nelumbo nucifera leaves extracts inhibit mouse airway smooth muscle contraction. BMC Complement. Altern. Med. 2017, 17, 159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Luo, X.J.; Sai, W.B.; Yu, M.F.; Li, W.E.; Ma, Y.F.; Chen, W.; Zhai, K.; Qin, G.; Guo, D.; et al. Non-selective cation channels mediate chloroquine-induced relaxation in precontracted mouse airway smooth muscle. PLoS ONE 2014, 9, e101578. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 218. [Google Scholar] [CrossRef]
- Zhang, M.; Ma, Y.; Ye, X.; Zhang, N.; Pan, L.; Wang, B. TRP (transient receptor potential) ion channel family: Structures, biological functions and therapeutic interventions for diseases. Signal Transduct. Target. Ther. 2023, 8, 261. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, Y.; Qin, L.; Chen, H. Functional cooperation between KCa3.1 and TRPV4 channels in bronchial smooth muscle cell proliferation associated with chronic asthma. Front. Pharmacol. 2017, 8, 559. [Google Scholar] [CrossRef]
- Bonvini, S.J.; Birrell, M.A.; Dubuis, E.; Adcock, J.J.; Wortley, M.A.; Flajolet, P.; Bradding, P.; Belvisi, M.G. Novel airway smooth muscle-mast cell interactions and a role for the TRPV4-ATP axis in non-atopic asthma. Eur. Respir. J. 2020, 56, 1901458. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Borkar, N.A.; Yao, Y.; Ye, X.; Vogel, E.R.; Pabelick, C.M.; Prakash, Y.S. Mechanosensitive channels in lung disease. Front. Physiol. 2023, 14, 1302631. [Google Scholar] [CrossRef] [PubMed]
- Reyes-García, J.; Carbajal-García, A.; Montaño, L.M. Transient receptor potential cation channel subfamily V (TRPV) and its importance in asthma. Eur. J. Pharmacol. 2022, 915, 174692. [Google Scholar] [CrossRef]
- Migulina, N.; Kelley, B.; Zhang, E.Y.; Pabelick, C.M.; Prakash, Y.S.; Vogel, E.R. Mechanosensitive channels in lung health and disease. Compr. Physiol. 2023, 13, 5157–5178. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, X.; Varty, L.; Rizzo, C.A.; Yang, R.; Correll, C.C.; Phelps, P.T.; Egan, R.W.; Hey, J.A. Functional TRPV4 channels are expressed in human airway smooth muscle cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2004, 287, L272–L278. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, Y.; Bai, S.; Ding, S.; Gao, H.; Yin, S.; Chen, S.; Lu, J.; Wang, H.; Shen, Y.; et al. TRPV4 complexes with the Na+/Ca2+ exchanger and IP3 receptor 1 to regulate local intracellular calcium and tracheal tension in mice. Front. Physiol. 2019, 10, 1471. [Google Scholar] [CrossRef] [PubMed]
- McAlexander, M.A.; Luttmann, M.A.; Hunsberger, G.E.; Undem, B.J. Transient receptor potential vanilloid 4 activation constricts the human bronchus via the release of cysteinyl leukotrienes. J. Pharmacol. Exp. Ther. 2014, 349, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Naumov, D.E.; Kolosov, V.P.; Perelman, J.M.; Prikhodko, A.G. Influence of TRPV4 gene polymorphisms on the development of osmotic airway hyperresponsiveness in patients with bronchial asthma. Dokl. Biochem. Biophys. 2016, 469, 260–263. [Google Scholar] [CrossRef]
- Zhu, G.; Investigators, I.; Gulsvik, A.; Bakke, P.; Ghatta, S.; Anderson, W.; Lomas, D.A.; Silverman, E.K.; Pillai, S.G. Association of TRPV4 gene polymorphisms with chronic obstructive pulmonary disease. Hum. Mol. Genet. 2009, 18, 2053–2062. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Sullivan, M.N.; Chase, M.; Gonzales, A.L.; Earley, S. Calcineurin/nuclear factor of activated T cells-coupled vanilliod transient receptor potential channel 4 Ca2+ sparklets stimulate airway smooth muscle cell proliferation. Am. J. Respir. Cell Mol. Biol. 2014, 50, 1064–1075. [Google Scholar] [CrossRef] [PubMed]
- Rajan, S.; Schremmer, C.; Weber, J.; Alt, P.; Geiger, F.; Dietrich, A. Ca2+ signaling by TRPV4 channels in respiratory function and disease. Cells 2021, 10, 822. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Zaman, M.K.; Das, S.; Goyary, D.; Pathak, M.P.; Chattopadhyay, P. Transient receptor potential vanilloid (TRPV4) channel inhibition: A novel promising approach for the treatment of lung diseases. Biomed. Pharmacother. 2023, 163, 114861. [Google Scholar] [CrossRef]
- Zaccor, N.W.; Sumner, C.J.; Snyder, S.H. The nonselective cation channel TRPV4 inhibits angiotensin II receptors. J. Biol. Chem. 2020, 295, 9986–9997. [Google Scholar] [CrossRef]
- Flores-Soto, E.; Martinez-Villa, I.; Solis-Chagoyan, H.; Sommer, B.; Lemini, C.; Montaño, L.M. 17β-Aminoestrogens induce guinea pig airway smooth muscle hyperresponsiveness through L-type Ca2+ channels activation. Steroids 2015, 101, 64–70. [Google Scholar] [CrossRef]
- Liu, B.B.; Peng, Y.B.; Zhang, W.J.; Zhao, X.X.; Chen, L.P.; Liu, M.S.; Wang, G.G.; Liu, Y.J.; Shen, J.; Zhao, P.; et al. NS8593 inhibits Ca2+ permeant channels reversing mouse airway smooth muscle contraction. Life Sci. 2019, 238, 116953. [Google Scholar] [CrossRef]
- Bazán-Perkins, B.; Carbajal, V.; Sommer, B.; Macias-Silva, M.; Gonzalez-Martinez, M.; Valenzuela, F.; Daniel, E.E.; Montaño, L.M. Involvement of different Ca2+ pools during the canine bronchial sustained contraction in Ca2+-free medium: Lack of effect of PKC inhibition. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1998, 358, 567–573. [Google Scholar] [CrossRef]
- Bazán-Perkins, B.; Flores-Soto, E.; Barajas-López, C.; Montaño, L.M. Role of sarcoplasmic reticulum Ca2+ content in Ca2+ entry of bovine airway smooth muscle cells. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2003, 368, 277–283. [Google Scholar] [CrossRef]
- Wang, Y.X.; Fleischmann, B.K.; Kotlikoff, M.I. M2 receptor activation of nonselective cation channels in smooth muscle cells: Calcium and Gi/Go requirements. Am. J. Physiol. 1997, 273, C500–C508. [Google Scholar] [CrossRef]
- Fleischmann, B.K.; Wang, Y.X.; Kotlikoff, M.I. Muscarinic activation and calcium permeation of nonselective cation currents in airway myocytes. Am. J. Physiol. 1997, 272, C341–C349. [Google Scholar] [CrossRef] [PubMed]
- Wen, N.; Xue, L.; Yang, Y.; Shi, S.; Liu, Q.H.; Cai, C.; Shen, J. Coptisine, a protoberberine alkaloid, relaxes mouse airway smooth muscle via blockade of VDLCCs and NSCCs. Biosci. Rep. 2020, 40, BSR20190534. [Google Scholar] [CrossRef]
- Zhao, P.; Liu, J.; Ming, Q.; Tian, D.; He, J.; Yang, Z.; Shen, J.; Liu, Q.H.; Yang, X. Dandelion extract relaxes mouse airway smooth muscle by blocking VDLCC and NSCC channels. Cell Biosci. 2020, 10, 125. [Google Scholar] [CrossRef] [PubMed]
- Jo, T.; Nagata, T.; Iida, H.; Imuta, H.; Iwasawa, K.; Ma, J.; Hara, K.; Omata, M.; Nagai, R.; Takizawa, H.; et al. Voltage-gated sodium channel expressed in cultured human smooth muscle cells: Involvement of SCN9A. FEBS Lett. 2004, 567, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, T.; Jo, T.; Meguro, K.; Oonuma, H.; Ma, J.; Kubota, N.; Imuta, H.; Takano, H.; Iida, H.; Nagase, T.; et al. Effect of dexamethasone on voltage-gated Na+ channel in cultured human bronchial smooth muscle cells. Life Sci. 2008, 82, 1210–1215. [Google Scholar] [CrossRef]
- Bradley, E.; Webb, T.I.; Hollywood, M.A.; Sergeant, G.P.; McHale, N.G.; Thornbury, K.D. The cardiac sodium current NaV1.5 is functionally expressed in rabbit bronchial smooth muscle cells. Am. J. Physiol. Cell Physiol. 2013, 305, C427–C435. [Google Scholar] [CrossRef][Green Version]
- Matthews, R.M.; Bradley, E.; Griffin, C.S.; Lim, X.R.; Mullins, N.D.; Hollywood, M.A.; Lundy, F.T.; McGarvey, L.P.; Sergeant, G.P.; Thornbury, K.D. Functional expression of NaV1.7 channels in freshly dispersed mouse bronchial smooth muscle cells. Am. J. Physiol. Cell Physiol. 2022, 323, C749–C762. [Google Scholar] [CrossRef]
- Snetkov, V.A.; Hirst, S.J.; Ward, J.P. Ion channels in freshly isolated and cultured human bronchial smooth muscle cells. Exp. Physiol. 1996, 81, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Scheraga, R.G.; Southern, B.D.; Grove, L.M.; Olman, M.A. The role of transient receptor potential vanilloid 4 in pulmonary inflammatory diseases. Front. Immunol. 2017, 8, 503. [Google Scholar] [CrossRef]
- Yin, J.; Michalick, L.; Tang, C.; Tabuchi, A.; Goldenberg, N.; Dan, Q.; Awwad, K.; Wang, L.; Erfinanda, L.; Nouailles, G.; et al. Role of transient receptor potential vanilloid 4 in neutrophil activation and acute lung injury. Am. J. Respir. Cell Mol. Biol. 2016, 54, 370–383. [Google Scholar] [CrossRef]
- Balakrishna, S.; Song, W.; Achanta, S.; Doran, S.F.; Liu, B.; Kaelberer, M.M.; Yu, Z.; Sui, A.; Cheung, M.; Leishman, E.; et al. TRPV4 inhibition counteracts edema and inflammation and improves pulmonary function and oxygen saturation in chemically induced acute lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 307, L158–L172. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kanju, P.; Patterson, M.; Chew, W.L.; Cho, S.H.; Gilmour, I.; Oliver, T.; Yasuda, R.; Ghio, A.; Simon, S.A.; et al. TRPV4-mediated calcium influx into human bronchial epithelia upon exposure to diesel exhaust particles. Environ. Health Perspect. 2011, 119, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Pairet, N.; Mang, S.; Fois, G.; Keck, M.; Kuhnbach, M.; Gindele, J.; Frick, M.; Dietl, P.; Lamb, D.J. TRPV4 inhibition attenuates stretch-induced inflammatory cellular responses and lung barrier dysfunction during mechanical ventilation. PLoS ONE 2018, 13, e0196055. [Google Scholar] [CrossRef]
- Wiesner, D.L.; Merkhofer, R.M.; Ober, C.; Kujoth, G.C.; Niu, M.; Keller, N.P.; Gern, J.E.; Brockman-Schneider, R.A.; Evans, M.D.; Jackson, D.J.; et al. Club cell TRPV4 serves as a damage sensor driving lung allergic inflammation. Cell Host Microbe 2020, 27, 614–628.e6. [Google Scholar] [CrossRef]
- Duan, J.; Xie, J.; Deng, T.; Xie, X.; Liu, H.; Li, B.; Chen, M. Exposure to both formaldehyde and high relative humidity exacerbates allergic asthma by activating the TRPV4-p38 MAPK pathway in Balb/c mice. Environ. Pollut. 2020, 256, 113375. [Google Scholar] [CrossRef]
- Niu, L.; Lu, Y.J.; Zu, X.W.; Yang, W.; Shen, F.K.; Xu, Y.Y.; Jiang, M.; Xie, Y.; Li, S.Y.; Gao, J.; et al. Magnolol alleviates pulmonary fibrosis inchronic obstructive pulmonary disease by targeting transient receptor potential vanilloid 4-ankyrin repeat domain. Phytother. Res. 2023, 37, 4282–4297. [Google Scholar] [CrossRef]
- Fernandez-Fernandez, J.M.; Nobles, M.; Currid, A.; Vazquez, E.; Valverde, M.A. Maxi K+ channel mediates regulatory volume decrease response in a human bronchial epithelial cell line. Am. J. Physiol. Cell Physiol. 2002, 283, C1705–C1714. [Google Scholar] [CrossRef]
- Woodrow, J.S.; Sheats, M.K.; Cooper, B.; Bayless, R. Asthma: The use of animal models and their translational utility. Cells 2023, 12, 1091. [Google Scholar] [CrossRef] [PubMed]
- Brocklehurst, W.E. The release of histamine and formation of a slow-reacting substance (SRS-A) during anaphylactic shock. J. Physiol. 1960, 151, 416–435. [Google Scholar] [CrossRef] [PubMed]
- Adams, G.K., 3rd; Lichtenstein, L.M. Antagonism of antigen-induced contraction of guinea pig and human airways. Nature 1977, 270, 255–257. [Google Scholar] [CrossRef] [PubMed]
- Lamm, W.J.; Lai, Y.L.; Hildebrandt, J. Histamine and leukotrienes mediate pulmonary hypersensitivity to antigen in guinea pigs. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1984, 56, 1032–1038. [Google Scholar] [CrossRef] [PubMed]
- Bjorck, T.; Dahlen, S.E. Leukotrienes and histamine mediate IgE-dependent contractions of human bronchi: Pharmacological evidence obtained with tissues from asthmatic and non-asthmatic subjects. Pulm. Pharmacol. 1993, 6, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Roquet, A.; Dahlen, B.; Kumlin, M.; Ihre, E.; Anstren, G.; Binks, S.; Dahlen, S.E. Combined antagonism of leukotrienes and histamine produces predominant inhibition of allergen-induced early and late phase airway obstruction in asthmatics. Am. J. Respir. Crit. Care Med. 1997, 155, 1856–1863. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Liu, Q.; Canning, B.J. Evidence for autocrine and paracrine regulation of allergen-induced mast cell mediator release in the guinea pig airways. Eur. J. Pharmacol. 2018, 822, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Canning, B.J.; Chou, Y. Using guinea pigs in studies relevant to asthma and COPD. Pulm. Pharmacol. Ther. 2008, 21, 702–720. [Google Scholar] [CrossRef]
- Albuquerque, A.A.; Carvalho, M.T.; Evora, P.M.; de Nadai, T.R.; Celotto, A.C.; Evora, P.R. In vitro reactivity (“organ chamber”) of guinea pig tracheal rings-methodology considerations. Ann. Transl. Med. 2016, 4, 216. [Google Scholar] [CrossRef]
- Sommer, B.; Flores-Soto, E.; Reyes-García, J.; Díaz-Hernández, V.; Carbajal, V.; Montaño, L.M. Na+ permeates through L-type Ca2+ channel in bovine airway smooth muscle. Eur. J. Pharmacol. 2016, 782, 77–88. [Google Scholar] [CrossRef]
- Flores-Soto, E.; Reyes-García, J.; Carbajal-García, A.; Campuzano-González, E.; Perusquía, M.; Sommer, B.; Montaño, L.M. Sex steroids effects on guinea pig airway smooth muscle tone and intracellular Ca2+ basal levels. Mol. Cell Endocrinol. 2017, 439, 444–456. [Google Scholar] [CrossRef] [PubMed]
- Imam, F.; Rehman, N.U.; Ansari, M.N.; Qamar, W.; Afzal, M.; Alharbi, K.S. Effect of Roflumilast in airways disorders via dual inhibition of phosphodiesterase and Ca2+-channel. Saudi Pharm. J. 2020, 28, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, S. Heating-induced contraction in airways smooth muscle: A possible causative factor of exercise-induced bronchoconstriction. Respir. Physiol. Neurobiol. 2019, 259, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [CrossRef]
- Kajita, J.; Yamaguchi, H. Calcium mobilization by muscarinic cholinergic stimulation in bovine single airway smooth muscle. Am. J. Physiol. 1993, 264, L496–L503. [Google Scholar] [CrossRef]
- Thorneloe, K.S.; Sulpizio, A.C.; Lin, Z.; Figueroa, D.J.; Clouse, A.K.; McCafferty, G.P.; Chendrimada, T.P.; Lashinger, E.S.; Gordon, E.; Evans, L.; et al. N-((1S)-1-[4-((2S)-2-[(2,4-dichlorophenyl)sulfonyl]amino-3-hydroxypropanoyl)-1-piperazinyl]carbonyl-3-methylbutyl)-1-benzothiophene-2-carboxamide (GSK1016790A), a novel and potent transient receptor potential vanilloid 4 channel agonist induces urinary bladder contraction and hyperactivity: Part I. J. Pharmacol. Exp. Ther. 2008, 326, 432–442. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montaño, L.M.; Carbajal-García, A.; Casas-Hernández, M.F.; Arredondo-Zamarripa, D.; Reyes-García, J. TRPV4 Activation during Guinea Pig Airway Smooth Muscle Contraction Promotes Ca2+ and Na+ Influx. Pharmaceuticals 2024, 17, 293. https://doi.org/10.3390/ph17030293
Montaño LM, Carbajal-García A, Casas-Hernández MF, Arredondo-Zamarripa D, Reyes-García J. TRPV4 Activation during Guinea Pig Airway Smooth Muscle Contraction Promotes Ca2+ and Na+ Influx. Pharmaceuticals. 2024; 17(3):293. https://doi.org/10.3390/ph17030293
Chicago/Turabian StyleMontaño, Luis M., Abril Carbajal-García, María F. Casas-Hernández, David Arredondo-Zamarripa, and Jorge Reyes-García. 2024. "TRPV4 Activation during Guinea Pig Airway Smooth Muscle Contraction Promotes Ca2+ and Na+ Influx" Pharmaceuticals 17, no. 3: 293. https://doi.org/10.3390/ph17030293
APA StyleMontaño, L. M., Carbajal-García, A., Casas-Hernández, M. F., Arredondo-Zamarripa, D., & Reyes-García, J. (2024). TRPV4 Activation during Guinea Pig Airway Smooth Muscle Contraction Promotes Ca2+ and Na+ Influx. Pharmaceuticals, 17(3), 293. https://doi.org/10.3390/ph17030293