

Antibacterial Activity and Mechanisms of Plant Flavonoids against Gram-Negative Bacteria Based on the Antibacterial Statistical Model

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Structure, Antibacterial Activity, and Physicochemical Parameters
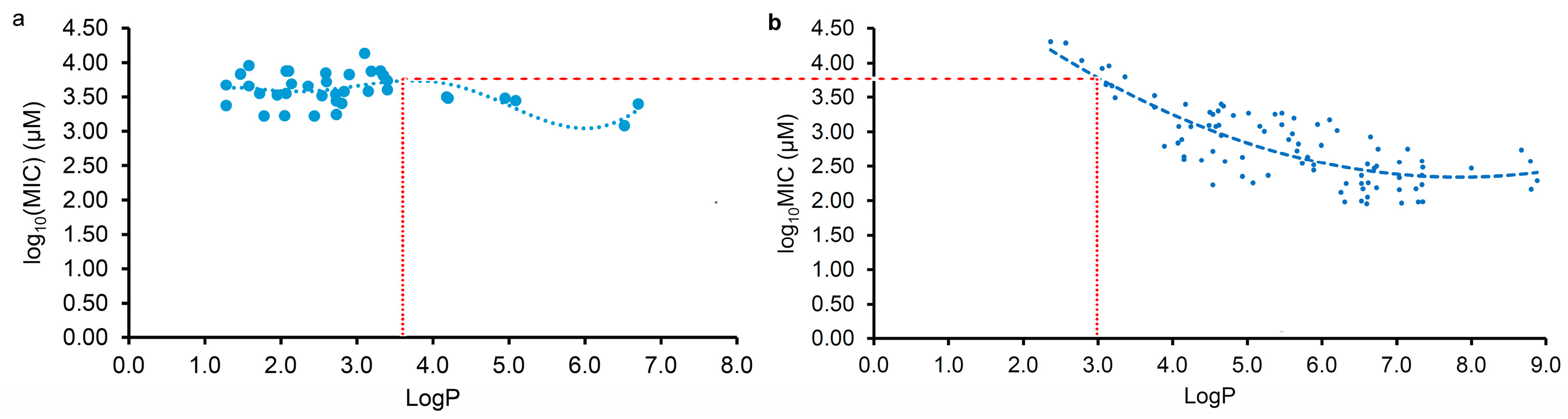
2.2. Correlation and Regression Analyses of the Antibacterial Activity and the Physicochemical Parameter
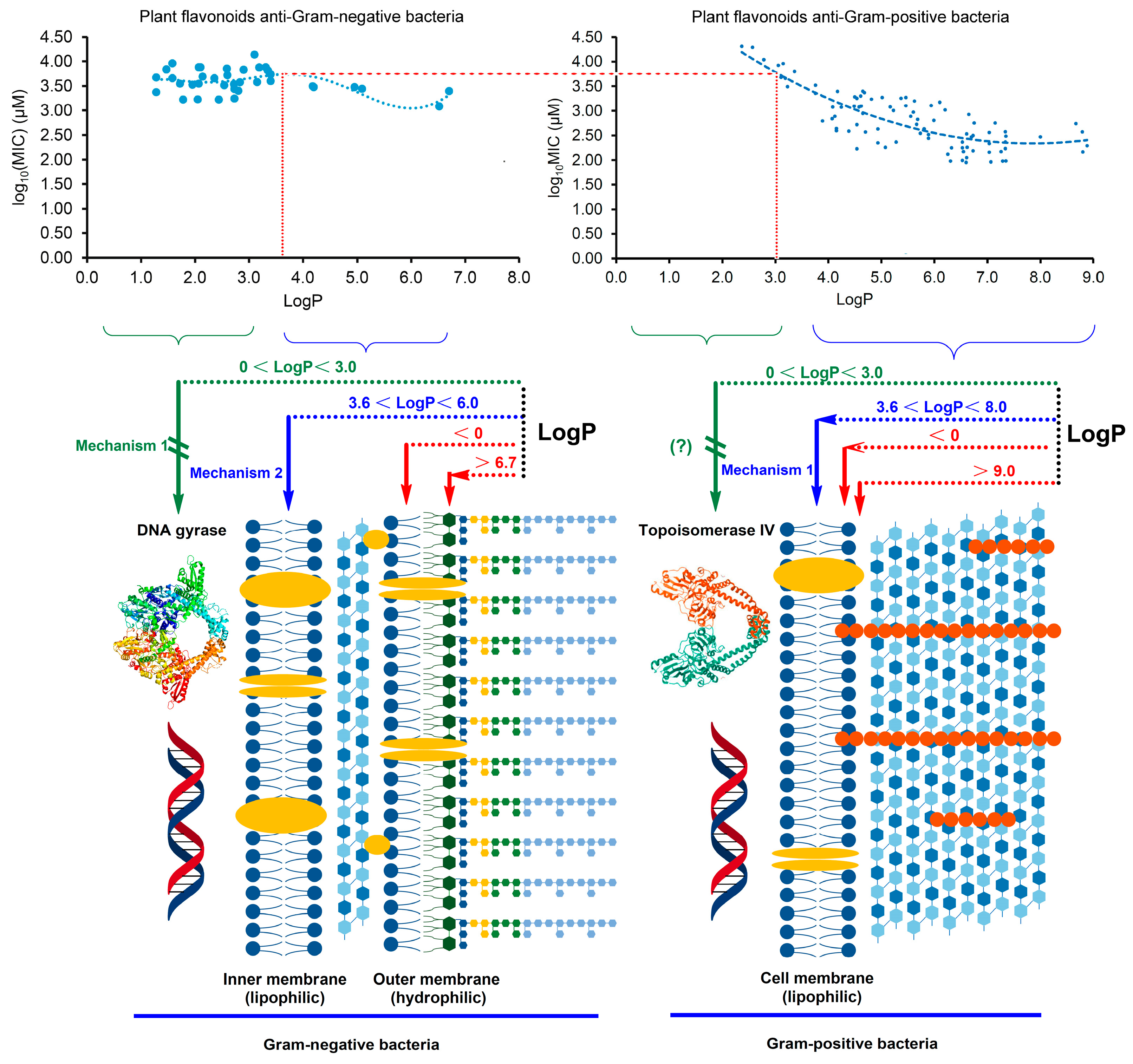
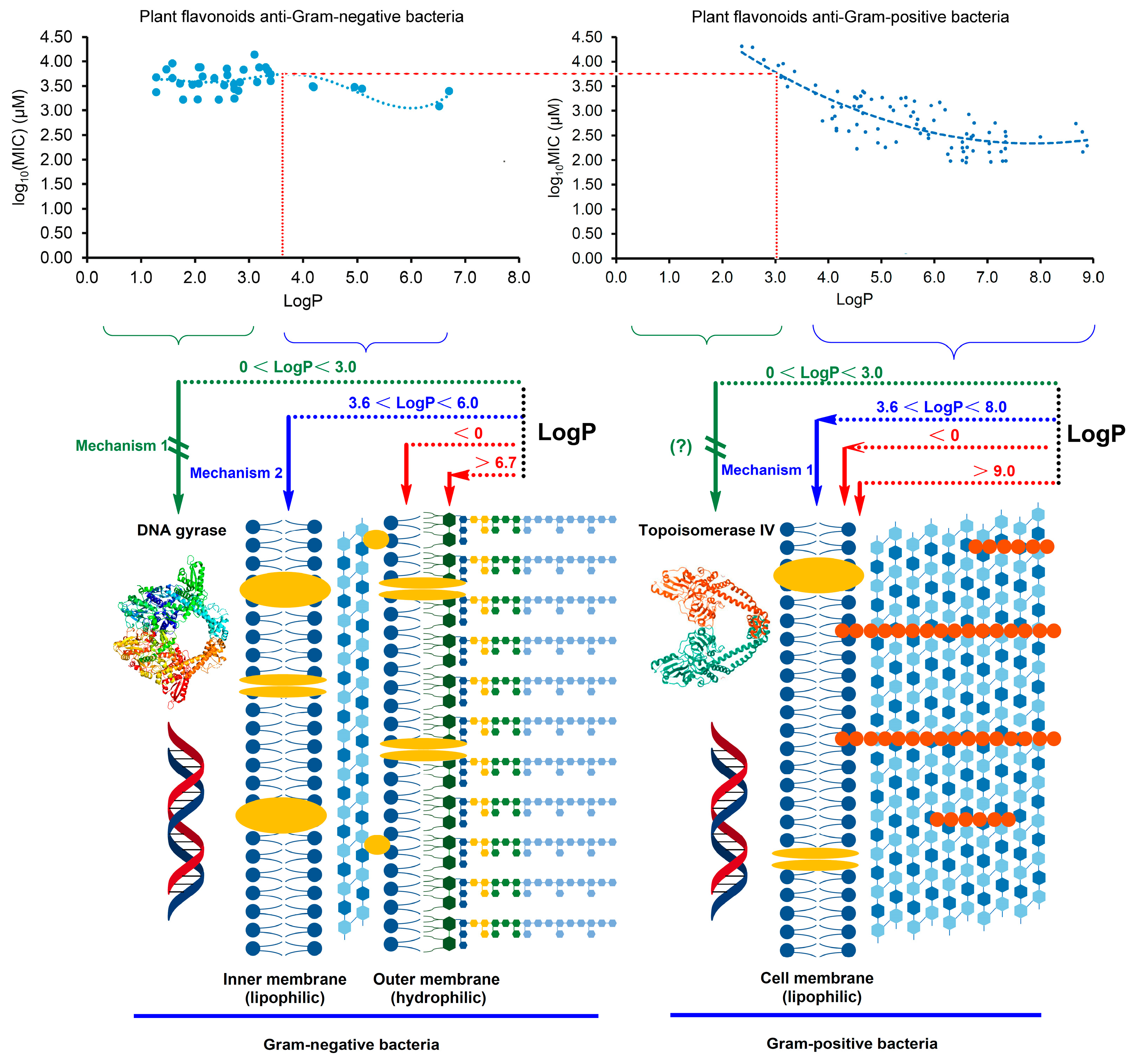
2.3. Different Antibacterial Effects and Mechanisms of Plant Flavonoids against Gram-Negative and Gram-Positive Bacteria
3. Discussion
4. Materials and Methods
4.1. Materials, Chemicals and Reagents
4.2. Bacterial Strains and Growth Conditions
4.3. Antimicrobial Susceptibility Assay and Prediction
4.4. Structures and MICs of Reported Plant Flavonoids
4.5. Correlation and Regression Analyses
4.6. Comparison of the Characteristics of Regression Curves
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Sridhar, D.; Blaser, M.; Wang, M.; Woolhouse, M. Achieving global targets for antimicrobial resistance. Science 2016, 353, 874–875. [Google Scholar] [CrossRef]
- Voulgaris, G.L.; Voulgari, M.L.; Falagas, M.E. Developments on antibiotics for multidrug resistant bacterial Gram-negative infections. Expert Rev. Anti. Infect. Ther. 2019, 17, 387–401. [Google Scholar] [CrossRef]
- Jean, S.S.; Gould, I.M.; Lee, W.S.; Hsueh, P.R.; International Society of Antimicrobial Chemotherapy (ISAC). New drugs for multidrug-resistant Gram-negative organisms: Time for stewardship. Drugs 2019, 79, 705–714. [Google Scholar] [CrossRef]
- Kaur, R.; Rani, P.; Atanasov, A.G.; Alzahrani, Q.; Gupta, R.; Kapoor, B.; Gulati, M.; Chawla, P. Discovery and development of antibacterial agents: Fortuitous and designed. Mini. Rev. Med. Chem. 2022, 22, 984–1029. [Google Scholar] [CrossRef]
- Lim, J.S.; Chai, Y.Y.; Ser, W.X.; Haeren, A.V.; Lim, Y.H.; Raja, T.; Foo, J.B.; Hamzah, S.; Sellappans, R.; Yow, H.Y. Novel drug candidates against antibiotic-resistant microorganisms: A review. Iran J. Basic. Med. Sci. 2024, 27, 134–150. [Google Scholar] [CrossRef]
- Xu, X.; Xu, L.; Yuan, G.; Wang, Y.; Qu, Y.; Zhou, M. Synergistic combination of two antimicrobial agents closing each other’s mutant selection windows to prevent antimicrobial resistance. Sci. Rep. 2018, 8, 7237. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Ge, X.; Xua, H.; Ma, K.; Zhang, W.; Zan, Y.; Efferth, T.; Xue, Z.; Hua, X. Phytochemicals with activity against methicillin-resistant Staphylococcus aureus. Phytomedicine 2022, 100, 154073. [Google Scholar] [CrossRef]
- Abdallah, E.M.; Alhatlani, B.Y.; de Paula Menezes, R.; Martins, C.H.G. Back to nature: Medicinal plants as promising sources for antibacterial drugs in the post-antibiotic era. Plants 2023, 12, 3077. [Google Scholar] [CrossRef] [PubMed]
- Thebti, A.; Meddeb, A.; Ben Salem, I.; Bakary, C.; Ayari, S.; Rezgui, F.; Essafi-Benkhadir, K.; Boudabous, A.; Ouzari, H.-I. Antimicrobial activities and mode of flavonoid actions. Antibiotics 2023, 12, 225. [Google Scholar] [CrossRef] [PubMed]
- Dey, J.; Mahapatra, S.R.; Raj, T.K.; Misra, N.; Suar, M. Identification of potential flavonoid compounds as antibacterial therapeutics against Klebsiella pneumoniae infection using structure-based virtual screening and molecular dynamics simulation. Mol. Divers. 2023. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Yang, W.; Tang, F.; Chen, X.; Ren, L. Antibacterial activities of flavonoids: Structure-activity relationship and mechanism. Curr. Med. Chem. 2015, 22, 132–149. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Deng, J.; Ye, Q.; Zhang, Z. The antibacterial activity of natural-derived flavonoids. Curr. Top. Med. Chem. 2022, 22, 1009–1019. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Hu, X.; Ren, X.; Liu, J.; Liu, X. Antibacterial modes of herbal flavonoids combat resistant bacteria. Front. Pharmacol. 2022, 13, 873374. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Yang, S.; Li, S.M. Naturally occurring prenylated chalcones from plants: Structural diversity, distribution, activities and biosynthesis. Nat. Prod. Rep. 2021, 38, 2236–2260. [Google Scholar] [CrossRef] [PubMed]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure-activity relationship: An update review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef]
- Panda, L.; Duarte-Sierra, A. Recent advancements in enhancing antimicrobial activity of plant-derived polyphenols by biochemical means. Horticulturae 2022, 8, 401. [Google Scholar] [CrossRef]
- Shamsudin, N.F.; Ahmed, Q.U.; Mahmood, S.; Ali Shah, S.A.; Khatib, A.; Mukhtar, S.; Alsharif, M.A.; Parveen, H.; Zakaria, Z.A. Antibacterial effects of flavonoids and their structure-activity relationship study: A comparative interpretation. Molecules 2022, 27, 1149. [Google Scholar] [CrossRef]
- Yuan, G.; Xia, X.; Guan, Y.; Yi, H.; Lai, S.; Sun, Y.; Cao, S. Antimicrobial quantitative relationship and mechanism of plant flavonoids to gram-positive bacteria. Pharmaceuticals 2022, 15, 1190. [Google Scholar] [CrossRef]
- Yuan, G.; Guan, Y.; Yi, H.; Lai, S.; Sun, Y.; Cao, S. Antibacterial activity and mechanism of plant flavonoids to gram-positive bacteria predicted from their lipophilicities. Sci. Rep. 2021, 11, 10471. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yan, Y.; Zhu, J.; Xia, X.; Yuan, G.; Li, S.; Deng, B.; Luo, X. Quinone pool, a key target of plant flavonoids inhibiting gram-positive bacteria. Molecules 2023, 28, 4972. [Google Scholar] [CrossRef]
- Mbaveng, A.T.; Sandjo, L.P.; Tankeo, S.B.; Ndifor, A.R.; Pantaleon, A.; Nagdjui, B.T.; Kuete, V. Antibacterial activity of nineteen selected natural products against multi-drug resistant Gram-negative phenotypes. SpringerPlus 2015, 4, 823. [Google Scholar] [CrossRef]
- Chukwujekwu, J.C.; Van Heerden, F.R.; Van Staden, J. Antibacterial activity of flavonoids from the stem bark of Erythrina caffra Thunb. Phytother. Res. 2011, 25, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Bennett, R.N.; Bisignano, G.; Trombetta, D.; Saija, A.; Faulds, C.B.; Gasson, M.J.; Narbad, A. Antimicrobial activity of flavonoids extracted from bergamot (Citrus bergamia Risso) peel, a byproduct of the essential oil industry. J. Appl. Microbiol. 2007, 103, 2056–2064. [Google Scholar] [CrossRef] [PubMed]
- Martini, N.D.; Katerere, D.R.; Eloff, J.N. Biological activity of five antibacterial flavonoids from Combretum erythrophyllum (Combretaceae). J. Ethnopharmacol. 2004, 93, 207–212. [Google Scholar] [CrossRef]
- Bagla, V.P.; McGaw, L.J.; Elgorashi, E.E.; Eloff, J.N. Antimicrobial activity, toxicity and selectivity index of two biflavonoids and a flavone isolated from Podocarpus henkelii (Podocarpaceae) leaves. BMC Complement. Altern. Med. 2014, 14, 383. [Google Scholar] [CrossRef]
- Wu, T.; Zang, X.; He, M.; Pan, S.; Xu, X. Structure-activity relationship of flavonoids on their anti-Escherichia coli activity and inhibition of DNA gyrase. J. Agric. Food. Chem. 2013, 61, 8185–8190. [Google Scholar] [CrossRef]
- Edziri, H.; Mastouri, M.; Mahjoub, M.A.; Mighri, Z.; Mahjoub, A.; Verschaeve, L. Antibacterial, antifungal and cytotoxic activities of two flavonoids from Retama raetam flowers. Molecules 2012, 17, 7284–7293. [Google Scholar] [CrossRef]
- Mohamed, M.S.; Abdelkader, K.; Gomaa, H.A.M.; Batubara, A.S.; Gamal, M.; Sayed, A.M. Mechanistic study of the antibacterial potential of the prenylated flavonoid auriculasin against Escherichia coli. Arch. Pharm. (Weinheim) 2022, 355, e2200360. [Google Scholar] [CrossRef]
- Eumkeb, G.; Siriwong, S.; Thumanu, K. Synergistic activity of luteolin and amoxicillin combination against amoxicillin-resistant Escherichia coli and mode of action. J. Photochem. Photobiol. B. 2012, 117, 247–253. [Google Scholar] [CrossRef]
- Jaisinghani, R.N. Antibacterial properties of quercetin. Microbiol. Res. 2017, 8, 6877. [Google Scholar] [CrossRef]
- Dzoyem, J.P.; Melong, R.; Tsamo, A.T.; Tchinda, A.T.; Kapche, D.G.; Ngadjui, B.T.; McGaw, L.J.; Eloff, J.N. Cytotoxicity, antimicrobial and antioxidant activity of eight compounds isolated from Entada abyssinica (Fabaceae). BMC Res. Notes 2017, 10, 118. [Google Scholar] [CrossRef]
- Eumkeb, G.; Siriwong, S.; Phitaktim, S.; Rojtinnakorn, N.; Sakdarat, S. Synergistic activity and mode of action of flavonoids isolated from smaller galangal and amoxicillin combinations against amoxicillin-resistant Escherichia coli. J. Appl. Microbiol. 2012, 112, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Ozçelik, B.; Orhan, I.; Toker, G. Antiviral and antimicrobial assessment of some selected flavonoids. Z. Naturforsch. C J. Biosci. 2006, 61, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Simo, I.K.; Ngameni, B.; Bigoga, J.D.; Watchueng, J.; Kapguep, R.N.; Etoa, F.X.; Tchaleu, B.N.; Beng, V.P. Antimicrobial activity of the methanolic extract, fractions and four flavonoids from the twigs of Dorstenia angusticornis Engl. (Moraceae). J. Ethnopharmacol. 2007, 112, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.d.l.A.; Zarelli, V.E.; Pappano, N.B.; Debattista, N.B. Bacteriostatic action of synthetic polyhydroxylated chalcones against Escherichia coli. Biocell. 2004, 28, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Liu, Y.; Li, T.; Liu, X.; Hao, Z.; Ding, S.; Panichayupakaranant, P.; Zhu, K.; Shen, J. Plant natural flavonoids against multidrug resistant pathogens. Adv. Sci. 2021, 8, e2100749. [Google Scholar] [CrossRef]
- Ohemeng, K.A.; Schwender, C.F.; Fu, K.P.; Barrett, J.F. DNA gyrase inhibitory and antibacterial activity of some flavones (l). Bioorg. Med. Chem. Lett. 1993, 3, 225–230. [Google Scholar] [CrossRef]
- Fang, Y.; Lu, Y.; Zang, X.; Wu, T.; Qi, X.; Pan, S.; Xu, X. 3D-QSAR and docking studies of flavonoids as potent Escherichia coli inhibitors. Sci. Rep. 2016, 6, 23634. [Google Scholar] [CrossRef] [PubMed]
- Verghese, J.; Nguyen, T.; Oppegard, L.M.; Seivert, L.M.; Hiasa, H.; Ellis, K.C. Flavone-based analogues inspired by the natural product simocyclinone D8 as DNA gyrase inhibitors. Bioorg. Med. Chem. Lett. 2013, 23, 5874–5877. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Hu, J.; Mei, F.; Zhang, Y.; Ma, Y.; Chen, Q.; Wang, C.; Fu, J.; Yang, M.; Wen, Z.; et al. Anti-microbial efficacy, mechanisms and druggability evaluation of the natural flavonoids. J. Appl. Microbiol. 2022, 133, 1975–1988. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory and Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 10th ed.; Approved Standards, CLSI Document M07-A10; Clinical and Laboratory and Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds (No.) a | Structure Types | LogP b | LogD7.40 b | E. coli ATCC 25922 | S. aureus ATCC 25923 | ||
|---|---|---|---|---|---|---|---|
| MIC (μM) c | Log10(MIC) | MIC (μM) c | Predicted MIC (μM) d | ||||
| Sophoraflavanone G (1) | Dihydroflavones | 6.52 | 6.33 | 1206.15 | 3.0814 | 9.42 | 16.37 |
| Neohesperidin (2) | Dihydroflavones | 2.44 | 1.99 | 1677.12 | 3.2246 | >1677.12 | 1816.01 |
| Naringin (3) | Dihydroflavones | 2.73 | 2.30 | 1763.88 | 3.2465 | >1763.88 | 1151.90 |
| Hesperidin (4) | Dihydroflavones | 1.78 | 1.33 | 1677.12 | 3.2246 | >1677.12 | 4440.63 |
| Methyl hesperidin (5) | Dihydroflavones | 2.54 | 2.10 | 3278.95 | 3.5157 | >3278.95 | 8410.66 |
| Eriodictyol (6) | Dihydroflavones | 2.59 | 2.34 | 7104.70 | 3.8515 | >7104.70 | 1442.76 |
| Eriocitrin (7) | Dihydroflavones | 1.47 | 1.03 | >3433.13 | >3.5357 | >3433.13 | 6397.02 |
| Hesperitin (8) | Dihydroflavones | 2.90 | 2.65 | 6775.18 | 3.8309 | >6775.18 | 863.94 |
| Naringenin (9) | Dihydroflavones | 3.19 | 2.96 | 7527.47 | 3.8766 | 3763.74 | 509.64 |
| didymin (10) | Dihydroflavones | 2.72 | 2.29 | 3444.51 | 3.5371 | >3444.51 | 1170.98 |
| Narirutin (11) | Dihydroflavones | 2.07 | 1.65 | 3527.75 | 3.5475 | >3527.75 | 3063.61 |
| Baicalein (12) | Flavones | 3.31 | 2.60 | >3789.22 | >3.5785 | >3789.22 | 404.47 |
| Licoflavone C (13) | Flavones | 4.20 | 3.77 | 3026.36 | 3.4809 | 3026.36 | 85.1 |
| Tangeritin (14) | Flavones | 2.73 | 2.73 | 2749.95 | 3.4393 | >2749.95 | 1151.9 |
| Nobiletin (15) | Flavones | 2.8 | 2.80 | 2544.73 | 3.4056 | >2544.74 | 1025.21 |
| Vitexin (16) | Flavones | 1.28 | 0.45 | 2368.29 | 3.3744 | 2368.29 | 7888.23 |
| Isovitexin (17) | Flavones | 1.28 | 0.15 | >2368.29 | >3.3744 | 2368.29 | 7888.23 |
| Diosmin (18) | Flavones | 2.05 | 1.23 | 1682.69 | 3.2260 | >1682.69 | 3146.25 |
| Rhoifolin (19) | Flavones | 1.72 | 0.91 | 3540.07 | 3.5490 | >3540.07 | 4777.21 |
| Apigenin (20) | Flavones | 2.10 | 1.57 | 7578.45 | 3.8796 | >7578.45 | 2942.82 |
| Diosmetin (21) | Flavones | 3.10 | 2.55 | >6820.53 | >3.8338 | >6820.53 | 603.30 |
| 5-Demethylnobiletin (22) | Flavones | 2.60 | 2.28 | 5273.32 | 3.7221 | >5273.32 | 1420.23 |
| 4’,5,7-Trimethoxyflavone (23) | Flavones | 3.35 | 3.35 | 6557.38 | 3.8167 | >6557.38 | 373.96 |
| Sinensetin (24) | Flavones | 3.40 | 3.40 | 5499.91 | 3.7404 | >5499.91 | 338.78 |
| Orientin (25) | Flavones | 1.58 | 0.72 | >4567.55 | >3.6597 | >4567.55 | 5639.27 |
| Isoorientin (26) | Flavones | 1.58 | 0.41 | 4567.55 | 3.6597 | >4567.55 | 5639.27 |
| Quercetin (27) | Flavonols | 2.07 | 1.40 | >3388.04 | >3.5299 | 13552.14 | 3063.61 |
| Galangin (28) | Flavonols | 2.83 | 2.16 | 3789.22 | 3.5785 | >3789.22 | 974.47 |
| Icaritin (29) | Flavonols | 5.09 | 4.54 | 2779.66 | 3.4430 | 2779.66 | 75.22 |
| Rutin (30) | Flavonols | 1.95 | 1.22 | >1677.26 | >3.2246 | 1677.26 | 3585.77 |
| Quercitrin (31) | Flavonols | 2.36 | 1.63 | 4567.55 | 3.6597 | >4567.55 | 2043.95 |
| Isoliquiritigenin (32) | Chalcones | 3.40 | 3.26 | 3995.94 | 3.6016 | 3995.94 | 338.78 |
| Licochalcone A (33) | Chalcones | 4.95 | 4.85 | 3026.00 | 3.4809 | 11.82 | 74.44 |
| Formononetin (34) | Isoflavones | 3.15 | 2.91 | 3817.05 | 3.5817 | >3817.04 | 549.61 |
| Puerarin (35) | Isoflavones | 2.14 | 1.59 | 4918.58 | 3.6918 | 614.82 | 2787.56 |
| Glabridin (36) | Isoflavanes | 4.18 | 4.18 | 3156.79 | 3.4992 | 49.32 | 86.81 |
| α-Mangostin (37) | Xanthones | 6.70 | 6.10 | 2494.70 | 3.3970 | 4.87 | 8.17 |
| Compounds (No.) a | Structure Types | LogP b | LogD7.40 b | MIC (μM) c | Log10(MIC) c | References |
|---|---|---|---|---|---|---|
| Candidone (38) | Dihydroflavones | 5.64 | 5.64 | 11.35 | 1.0540 | [23] |
| Atalantoflavone (39) | Flavones | 4.31 | 3.58 | 761.13 | 2.8815 | [23] |
| 2’-hydroxyatalantoflavone (40) | Flavones | 3.87 | 3.10 | 726.57 | 2.8613 | [23] |
| Neocyclomorusin (41) | Flavones | 4.30 | 2.87 | 18.33 | 1.2632 | [23] |
| Neobavaisoflavone (42) | Isoflavones | 4.68 | 4.69 | 23.64 | 1.3736 | [23] |
| Daidzein (43) | Isoflavones | 3.14 | 2.61 | 236.83 | 2.3744 | [23] |
| Isowighteone (44) | Isoflavones | 4.78 | 4.23 | 380.57 | 2.5804 | [23] |
| Isoneorautenol (45) | Dihydroisoflavones | 4.16 | 4.16 | 794.14 | 2.8999 | [23] |
| Abyssione-V 4′-O-methyl ether (46) | Dihydroflavones | 8.03 | 7.79 | 9.23 | 0.9652 | [24] |
| 6,8-diprenylgenistein (47) | Isoflavones | 7.33 | 7.16 | 19.19 | 1.2831 | [24] |
| Alpinumisoflavone (48) | Isoflavones | 5.80 | 4.93 | 11.60 | 1.0645 | [24] |
| Eriodictyol (49) | Dihydroflavones | 2.59 | 2.34 | 867.27 | 2.9382 | [25] |
| Hesperetin (50) | Dihydroflavones | 2.90 | 2.65 | 3308.19 | 3.5196 | [25] |
| Neohesperidin (51) | Dihydroflavones | 2.44 | 1.99 | >1637.81 | >3.2143 | [25] |
| Neoeriocitrin (52) | Dihydroflavones | 2.13 | 1.68 | 1341.07 | 3.1275 | [25] |
| Naringin (53) | Dihydroflavones | 2.73 | 2.30 | >1722.53 | >3.2362 | [25] |
| Naringenin (54) | Dihydroflavones | 3.19 | 2.96 | 2938.37 | 3.4681 | [25] |
| 5-hydroxy-7,4’-dimethoxyflavone (55) | Flavones | 3.40 | 2.78 | 167.62 | 2.2243 | [26] |
| Genkwanin (56) | Flavones | 2.36 | 1.75 | 351.78 | 2.5463 | [26] |
| Quercetin-5,3’-dimethylether (57) | Flavonols | 2.30 | 1.74 | 151.38 | 2.1801 | [26] |
| Rhamnazin (58) | Flavonols | 2.51 | 1.73 | 302.76 | 2.4811 | [26] |
| Rhamnocitrin (59) | Flavonols | 2.56 | 1.82 | 166.52 | 2.2215 | [26] |
| 7,4’, 7’’, 4’’’-tetramethoxy amentoflavone (TMA) (60) | Bioflavones | 5.80 | 4.10 | 420.47 | 2.6237 | [27] |
| Isoginkgetin (IGG) (61) | Bioflavones | 5.68 | 4.32 | 242.29 | 2.3843 | [27] |
| Podocarpusflavone A (PFA) (62) | Bioflavones | 4.15 | 2.53 | 452.5 | 2.6556 | [27] |
| Nobiletin (63) | Flavones | 2.80 | 2.80 | 439.86 | 2.6433 | [28] |
| Kaempferol (64) | Flavonols | 2.05 | 1.40 | 87.34 | 1.9412 | [28] |
| Licoflavone C (65) | Flavones | 4.20 | 3.77 | 23.08 | 1.3632 | [29] |
| Derrone (66) | Isoflavones | 5.55 | 4.82 | 23.22 | 1.3659 | [29] |
| Epimedokoreanin B (67) | Flavones | 6.59 | 6.14 | >151.49 | >2.1804 | [30] |
| Auriculasin (68) | Isoflavones | 7.70 | 6.89 | 9.51 | 0.9782 | [30] |
| Pomiferin (69) | Isoflavones | 7.44 | 7.06 | 19.03 | 1.2794 | [30] |
| Gancaonin L (70) | Isoflavones | 5.03 | 4.58 | 22.58 | 1.3537 | [30] |
| Mopanin (71) | Flavonols | 1.94 | 1.04 | >214.59 | >2.3316 | [30] |
| Luteolin (72) | Flavones | 2.40 | 1.85 | 349.36 | 2.5433 | [31] |
| Quercetin (73) | Flavonols | 2.07 | 1.40 | 1323.45 | 3.1217 | [32] |
| Quercetin 3-O-β-D-glucosyl (1→4)-α-L-rhamnoside (74) | Flavonols | 2.92 | 2.20 | 40.02 | 1.6023 | [33] |
| Quercetin-3-O-α-L-rhamnoside or quercitrin (75) | Flavonols | 2.51 | 1.78 | 108.13 | 2.0339 | [33] |
| Entadanin (76) | Flavonols | 2.00 | 0.086 | 36.31 | 1.5600 | [33] |
| Kaempferide (77) | Flavonols | 3.00 | 2.33 | 333.03 | 2.5225 | [34] |
| Kaempferide-3-O-β-D-glucoside (78) | Flavonols | 2.31 | 1.60 | 432.52 | 2.6360 | [34] |
| Galangin (79) | Flavonols | 2.83 | 2.16 | 370.04 | 2.5682 | [34] |
| Tiliroside (80) | Flavonols | 4.02 | 3.32 | 3.36 | 0.5263 | [35] |
| Quercetin-3,7-O-α-L-dirhamnoside (81) | Flavonols | 1.53 | 0.53 | 3.36 | 0.5263 | [35] |
| Kaempferol-3,7-O-α-L-dirhamnoside (82) | Flavonols | 1.73 | 0.76 | 3.46 | 0.5391 | [35] |
| Scandenone (83) | Isoflavones | 7.90 | 7.12 | 4.94 | 0.6937 | [35] |
| Angusticornin B (84) | Chalcones | 4.31 | 4.07 | 2.87 | 0.4579 | [36] |
| Bartericin A (85) | Chalcones | 5.46 | 5.77 | 1.49 | 0.1732 | [36] |
| 2’,4’,2-(OH)3-chalcone (86) | Chalcones | 3.95 | 3.66 | 476.08 | 2.6777 | [37] |
| 2’,4’,3-(OH)3-chalcone (87) | Chalcones | 3.35 | 3.05 | 179.51 | 2.2541 | [37] |
| Isobavachalcone (88) | Chalcones | 5.49 | 5.44 | >394.6 | >2.5962 | [38] |
| α-Mangostin (89) | Xanthones | 6.70 | 6.10 | >311.84 | >2.4939 | [38] |
| Equation Number | Sample Numbers (n) | Parameters a (x) | Regression Equation (r b) | Coefficient of Determination (R2) |
|---|---|---|---|---|
| (1) | 37 | LogP | y = 154.74x4 – 2328.1x3 + 11,774x2 – 23,630x + 20,574 (0.3736) c | 0.1396 |
| (2) | y = −0.0022x6 + 0.0574x5 – 0.5624x4 + 2.6776x3–6.4916x2 + 7.5985x + 0.2396 (0.4714) d | 0.2222 | ||
| (3) | 37 | LogD7.40 | y = 51.533x4 – 648.46x3 + 2399.6x2 – 2834x + 5407.4 (0.3412) c | 0.1164 |
| (4) | 46 | LogP | y = 3.4251x6 – 99.079x5 + 1128.5x4 – 6389x3 + 18,606x2 – 25,841x + 13,552 (0.4108) c | 0.1688 |
| (5) | y = −0.0358x4 + 0.7264x3–5.2356x2 + 15.438x–13.244 (0.6670) d | 0.4449 | ||
| (6) | y = 0.0621x3–0.9417x2 + 4.0824x – 2.9354 (0.5875) d | 0.3452 | ||
| (7) | 46 | LogD7.40 | / e | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Y.; Xia, X.; Fatima, A.; Zhang, L.; Yuan, G.; Lian, F.; Wang, Y. Antibacterial Activity and Mechanisms of Plant Flavonoids against Gram-Negative Bacteria Based on the Antibacterial Statistical Model. Pharmaceuticals 2024, 17, 292. https://doi.org/10.3390/ph17030292
Yan Y, Xia X, Fatima A, Zhang L, Yuan G, Lian F, Wang Y. Antibacterial Activity and Mechanisms of Plant Flavonoids against Gram-Negative Bacteria Based on the Antibacterial Statistical Model. Pharmaceuticals. 2024; 17(3):292. https://doi.org/10.3390/ph17030292
Chicago/Turabian StyleYan, Yu, Xuexue Xia, Aiman Fatima, Li Zhang, Ganjun Yuan, Fengxian Lian, and Yu Wang. 2024. "Antibacterial Activity and Mechanisms of Plant Flavonoids against Gram-Negative Bacteria Based on the Antibacterial Statistical Model" Pharmaceuticals 17, no. 3: 292. https://doi.org/10.3390/ph17030292
APA StyleYan, Y., Xia, X., Fatima, A., Zhang, L., Yuan, G., Lian, F., & Wang, Y. (2024). Antibacterial Activity and Mechanisms of Plant Flavonoids against Gram-Negative Bacteria Based on the Antibacterial Statistical Model. Pharmaceuticals, 17(3), 292. https://doi.org/10.3390/ph17030292