
The Potential Protective Role of Naringenin against Dasatinib-Induced Hepatotoxicity

 , , , , , , ,
, , , , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Kits
2.2. Animals
2.3. Experimantal Design
2.3.1. Determine DASA Tolerability and Survival Rates
2.3.2. Determine the Efficacy of Naringenin on DASA-Induced Liver Toxicity
2.4. Samples Collection
2.5. Tissue Analysis
2.6. Histological Evaluation
2.7. Gene Expression by Real-Time PCR
2.8. Western Blotting for NF-kB
2.9. Statistical Analyses
3. Results
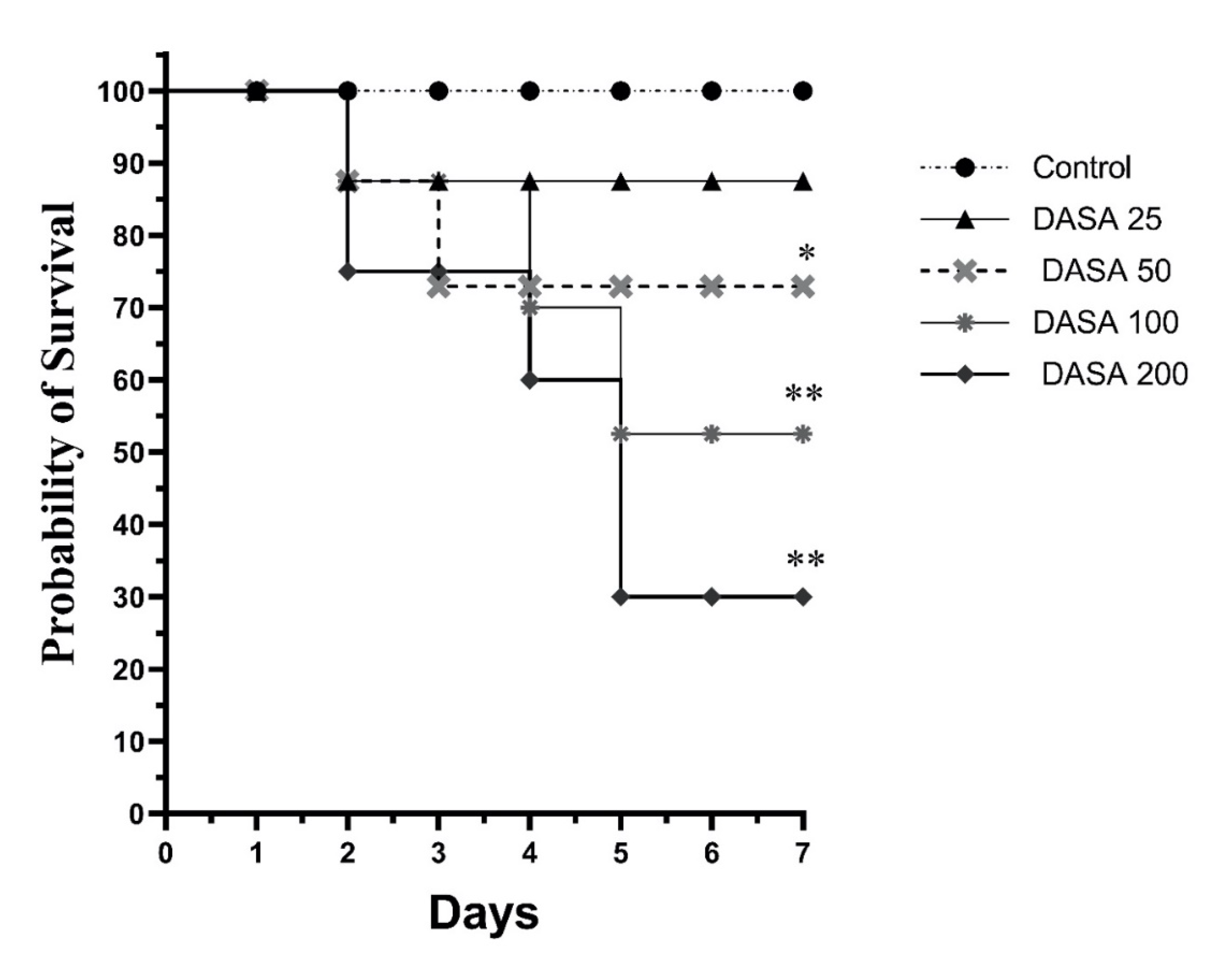
3.1. Effect of DASA Doses on Survival Rate
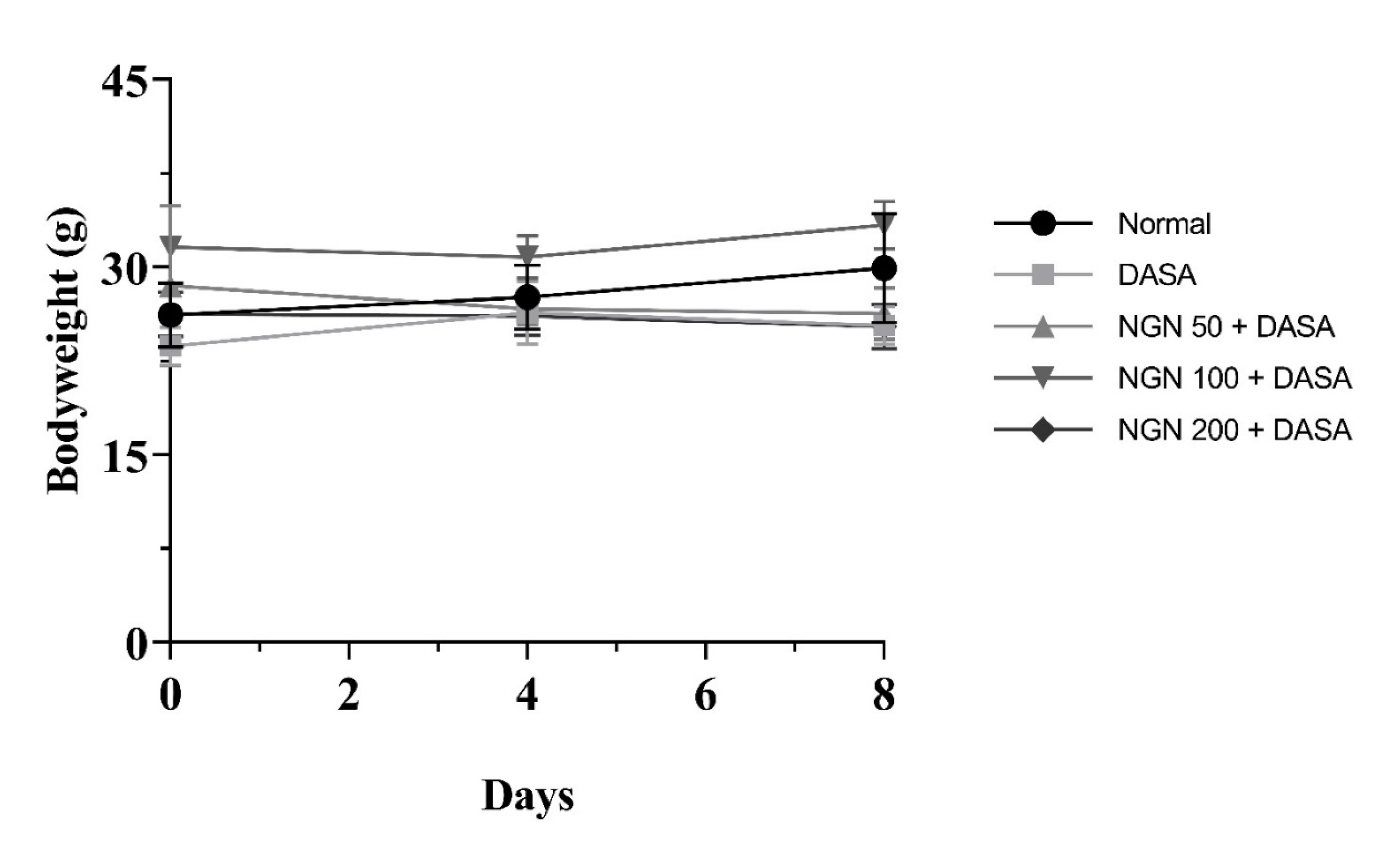
3.2. Effect of NGN ± DASA on Mice’s Bodyweights
3.3. Effect of NGN ± DASA on Liver Enzymes
3.4. Effect of NGN ± DASA on Liver Histopathology
3.5. Effect of NGN ± DASA on Gene Expression of TNF-α and IL-10
3.6. Effect of NGN ± DASA on Antioxidant Genes (CAT, SOD2, GST, and GPX)
3.7. Effect of NGN ± DASA on Tissue Concentrations of TNFα, IL-6, CAT, GSH, and MDA Levels
3.8. Effect of NGN ± DASA on the Expression of NF-κB Protein
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rix, U.; Hantschel, O.; Dürnberger, G.; Remsing Rix, L.L.; Planyavsky, M.; Fernbach, N.V.; Kaupe, I.; Bennett, K.L.; Valent, P.; Colinge, J. Chemical Proteomic Profiles of the BCR-ABL Inhibitors Imatinib, Nilotinib, and Dasatinib Reveal Novel Kinase and Nonkinase Targets. Blood 2007, 110, 4055–4063. [Google Scholar] [CrossRef] [PubMed]
- Ottmann, O.; Dombret, H.; Martinelli, G.; Simonsson, B.; Guilhot, F.; Larson, R.A.; Rege-Cambrin, G.; Radich, J.; Hochhaus, A.; Apanovitch, A.M. Dasatinib Induces Rapid Hematologic and Cytogenetic Responses in Adult Patients with Philadelphia Chromosome–Positive Acute Lymphoblastic Leukemia with Resistance or Intolerance to Imatinib: Interim Results of a Phase 2 Study. Blood J. Am. Soc. Hematol. 2007, 110, 2309–2315. [Google Scholar] [CrossRef] [PubMed]
- Schlenk, R.F.; Kayser, S. Small Molecules in Hematology; Springer: Berlin/Heidelberg, Germany, 2018; Volume 212. [Google Scholar] [CrossRef]
- Yang, X.; Wang, J.; Dai, J.; Shao, J.; Ma, J.; Chen, C.; Ma, S.; He, Q.; Luo, P.; Yang, B. Autophagy Protects against Dasatinib-Induced Hepatotoxicity via P38 Signaling. Oncotarget 2015, 6, 6203. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Luo, P.; Zhu, H.; Zhao, Y.; Wu, H.; Gai, R.; Wu, Y.; Yang, B.; Yang, X.; He, Q. Oxidative Stress Is Involved in Dasatinib-Induced Apoptosis in Rat Primary Hepatocytes. Toxicol. Appl. Pharm. 2012, 261, 280–291. [Google Scholar] [CrossRef]
- Suh, K.J.; Lee, J.Y.; Shin, D.Y.; Koh, Y.; Bang, S.M.; Yoon, S.S.; Park, S.; Kim, I.; Lee, J.O. Analysis of Adverse Events Associated with Dasatinib and Nilotinib Treatments in Chronic-Phase Chronic Myeloid Leukemia Patients Outside Clinical Trials. Int. J. Hematol. 2017, 106, 229–239. [Google Scholar] [CrossRef]
- Wei, G.; Rafiyath, S.; Liu, D. First-Line Treatment for Chronic Myeloid Leukemia: Dasatinib, Nilotinib, or Imatinib. J. Hematol. Oncol. 2010, 3, 45. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The Therapeutic Potential of Naringenin: A Review of Clinical Trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef]
- Hernández-Aquino, E.; Muriel, P. Beneficial Effects of Naringenin in Liver Diseases: Molecular Mechanisms. World J. Gastroenterol. 2018, 24, 1679. [Google Scholar] [CrossRef]
- Alam, M.A.; Subhan, N.; Rahman, M.M.; Uddin, S.J.; Reza, H.M.; Sarker, S.D. Effect of Citrus Flavonoids, Naringin and Naringenin, on Metabolic Syndrome and Their Mechanisms of Action. Adv. Nutr. 2014, 5, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Ozkaya, A.; Sahin, Z.; Dag, U.; Ozkaraca, M. Effects of Naringenin on Oxidative Stress and Histopathological Changes in the Liver of Lead Acetate Administered Rats. J. Biochem. Mol. Toxicol. 2016, 30, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-J.; Choi, Y.-K.; Im, H.-S.; Yarimaga, O.; Yoon, E.; Kim, H.-S. Aspartate Aminotransferase (AST/GOT) and Alanine Aminotransferase (ALT/GPT) Detection Techniques. Sensors 2006, 6, 756–782. [Google Scholar] [CrossRef]
- Brown, R.A.M.; Epis, M.R.; Horsham, J.L.; Kabir, T.D.; Richardson, K.L.; Leedman, P.J. Total RNA Extraction from Tissues for MicroRNA and Target Gene Expression Analysis: Not All Kits Are Created Equal. BMC Biotechnol. 2018, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mori, R.; Wang, Q.; Danenberg, K.D.; Pinski, J.K.; Danenberg, P.V. Both Β-actin and GAPDH Are Useful Reference Genes for Normalization of Quantitative RT-PCR in Human FFPE Tissue Samples of Prostate Cancer. Prostate 2008, 68, 1555–1560. [Google Scholar] [CrossRef]
- Dong, Y.; Shi, O.; Zeng, Q.; Lu, X.; Wang, W.; Li, Y.; Wang, Q. Leukemia Incidence Trends at the Global, Regional, and National Level between 1990 and 2017. Exp. Hematol. Oncol. 2020, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Illnesses, S. Leukemia. In Tumors in Adolescents and Young Adults; Karger Publishers: Basel, Switzerland, 2016; Volume 43, pp. 87–100. [Google Scholar] [CrossRef]
- Eşkazan, A.E. Tyrosine Kinase Inhibitors (TKIs) Used in the Management of Chronic Myeloid Leukaemia Are Associated with Haematologic Toxicities—Which TKI Is the Safest? Br. J. Clin. Pharm. 2019, 85, 2241. [Google Scholar] [CrossRef]
- Mathisen, M.S.; O’Brien, S.; Thomas, D.; Cortes, J.; Kantarjian, H.; Ravandi, F. Role of Tyrosine Kinase Inhibitors in the Management of Philadelphia Chromosome–Positive Acute Lymphoblastic Leukemia. Curr. Hematol. Malig. Rep. 2011, 6, 187. [Google Scholar] [CrossRef]
- Karim, N.; Jia, Z.; Zheng, X.; Cui, S.; Chen, W. A Recent Review of Citrus Flavanone Naringenin on Metabolic Diseases and Its Potential Sources for High Yield-Production. In Trends in Food Science and Technology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 35–54. [Google Scholar] [CrossRef]
- Pinho-Ribeiro, F.A.; Zarpelon, A.C.; Fattori, V.; Manchope, M.F.; Mizokami, S.S.; Casagrande, R.; Verri, W.A. Naringenin Reduces Inflammatory Pain in Mice. Neuropharmacology 2016, 105, 508–519. [Google Scholar] [CrossRef]
- Yin, J.; Liang, Y.; Wang, D.; Yan, Z.; Yin, H.; Wu, D.; Su, Q. Naringenin Induces Laxative Effects by Upregulating the Expression Levels of C-Kit and SCF, as Well as Those of Aquaporin 3 in Mice with Loperamide-Induced Constipation. Int. J. Mol. Med. 2018, 41, 649–658. [Google Scholar] [CrossRef]
- Morton, J.P.; Karim, S.A.; Graham, K.; Timpson, P.; Jamieson, N.; Athineos, D.; Doyle, B.; McKay, C.; Heung, M.; Oien, K.A. Dasatinib Inhibits the Development of Metastases in a Mouse Model of Pancreatic Ductal Adenocarcinoma. Gastroenterology 2010, 139, 292–303. [Google Scholar] [CrossRef]
- Cortes, J.E.; Abruzzese, E.; Chelysheva, E.; Guha, M.; Wallis, N.; Apperley, J.F. The Impact of Dasatinib on Pregnancy Outcomes. Am. J. Hematol. 2015, 90, 1111–1115. [Google Scholar] [CrossRef] [PubMed]
- Pari, L.; Gnanasoundari, M. Influence of Naringenin on Oxytetracycline Mediated Oxidative Damage in Rat Liver. Basic Clin. Pharm. Toxicol. 2006, 98, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Renugadevi, J.; Prabu, S.M. Cadmium-Induced Hepatotoxicity in Rats and the Protective Effect of Naringenin. Exp. Toxicol. Pathol. 2010, 62, 171–181. [Google Scholar] [CrossRef]
- Lv, Y.; Zhang, B.; Xing, G.; Wang, F.; Hu, Z. Protective Effect of Naringenin against Acetaminophen-Induced Acute Liver Injury in Metallothionein (MT)-Null Mice. Food Funct. 2013, 4, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Wali, A.F.; Rashid, S.; Rashid, S.M.; Ansari, M.A.; Khan, M.R.; Haq, N.; Alhareth, D.Y.; Ahmad, A.; Rehman, M.U. Naringenin Regulates Doxorubicin-Induced Liver Dysfunction: Impact on Oxidative Stress and Inflammation. Plants 2020, 9, 550. [Google Scholar] [CrossRef]
- Jayaraman, J.; Jesudoss, V.A.S.; Menon, V.P.; Namasivayam, N. Anti-Inflammatory Role of Naringenin in Rats with Ethanol Induced Liver Injury. Toxicol. Mech. Methods 2012, 22, 568–576. [Google Scholar] [CrossRef]
- Hernández-Aquino, E.; Quezada-Ramírez, M.A.; Silva-Olivares, A.; Casas-Grajales, S.; Ramos-Tovar, E.; Flores-Beltrán, R.E.; Segovia, J.; Shibayama, M.; Muriel, P. Naringenin Attenuates the Progression of Liver Fibrosis via Inactivation of Hepatic Stellate Cells and Profibrogenic Pathways. Eur. J. Pharm. 2019, 865, 172730. [Google Scholar] [CrossRef]
- Wang, Q.; Ou, Y.; Hu, G.; Wen, C.; Yue, S.; Chen, C.; Xu, L.; Xie, J.; Dai, H.; Xiao, H.; et al. Naringenin Attenuates Non-Alcoholic Fatty Liver Disease by down-Regulating the NLRP3/NF-ΚB Pathway in Mice. Br. J. Pharm. 2020, 177, 1806–1821. [Google Scholar] [CrossRef]
- Alfrey, E.J.; Most, D.; Wang, X.; Lee, L.K.; Holm, B.; Krieger, N.R.; Sibley, R.K.; Huie, P.; Dafoe, D.C. Interferon-Gamma and Interleukin-10 Messenger RNA Are up-Regulated after Orthotopic Liver Transplantation in Tolerant Rats: Evidence for Cytokine-Mediated Immune Dysregulation. Surgery 1995, 118, 399–404; discussion 404–405. [Google Scholar] [CrossRef]
- Knolle, P.; Schlaak, J.; Uhrig, A.; Kempf, P.; Meyer zum Büschenfelde, K.H.; Gerken, G. Human Kupffer Cells Secrete IL-10 in Response to Lipopolysaccharide (LPS) Challenge. J. Hepatol. 1995, 22, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Arai, T.; Hiromatsu, K.; Kobayashi, N.; Takano, M.; Ishida, H.; Nimura, Y.; Yoshikai, Y. IL-10 Is Involved in the Protective Effect of Dibutyryl Cyclic Adenosine Monophosphate on Endotoxin-Induced Inflammatory Liver Injury. J. Immunol. 1995, 155, 5743–5749. [Google Scholar] [CrossRef]
- Gazzinelli, R.T.; Wysocka, M.; Hieny, S.; Scharton-Kersten, T.; Cheever, A.; Kühn, R.; Müller, W.; Trinchieri, G.; Sher, A. In the Absence of Endogenous IL-10, Mice Acutely Infected with Toxoplasma Gondii Succumb to a Lethal Immune Response Dependent on CD4+ T Cells and Accompanied by Overproduction of IL-12, IFN-Gamma and TNF-Alpha. J. Immunol. 1996, 157, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Louis, H.; Le Moine, O.; Peny, M.O.; Gulbis, B.; Nisol, F.; Goldman, M.; Devière, J. Hepatoprotective Role of Interleukin 10 in Galactosamine/Lipopolysaccharide Mouse Liver Injury. Gastroenterology 1997, 112, 935–942. [Google Scholar] [CrossRef]
- Louis, H.; Le Moine, O.; Peny, M.O.; Quertinmont, E.; Fokan, D.; Goldman, M.; Devière, J. Production and Role of Interleukin-10 in Concanavalin A-Induced Hepatitis in Mice. Hepatology 1997, 25, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Louis, H.; Le Moine, O.; Goldman, M.; Devière, J. Modulation of Liver Injury by Interleukin-10. Acta Gastroenterol. Belg. 2003, 66, 7–14. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | n | Treatment Plan |
|---|---|---|
| Control | 8 | Received the vehicle (0.5% carboxymethyl cellulose) by oral gavage daily for 8 days. |
| DASA | 8 | Received the vehicle (0.5% carboxymethyl cellulose) daily for 7 days and subsequently treated with DASA (25 mg/kg) on the 8th day. |
| NGN50 + DASA | 8 | Received NGN (50 mg/kg, solubilized in 0.5% carboxymethyl cellulose) daily for 7 days and were subsequently treated with DASA (25 mg/kg) on the 8th day. |
| NGN100 + DASA | 8 | Received NGN (100 mg/kg, solubilized in 0.5% carboxymethyl cellulose) daily for 7 days and were subsequently treated with DASA (25 mg/kg) on the 8th day. |
| NGN200 + DASA | 8 | Received NGN (200 mg/kg, solubilized in 0.5% carboxymethyl cellulose) daily for 7 days and were subsequently treated with DASA (25 mg/kg) on the 8th day. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alanazi, A.Z.; Alhazzani, K.; Alrewily, S.Q.; Aljerian, K.; Algahtani, M.M.; Alqahtani, Q.H.; Haspula, D.; Alhamed, A.S.; Alqinyah, M.; Raish, M. The Potential Protective Role of Naringenin against Dasatinib-Induced Hepatotoxicity. Pharmaceuticals 2023, 16, 921. https://doi.org/10.3390/ph16070921
Alanazi AZ, Alhazzani K, Alrewily SQ, Aljerian K, Algahtani MM, Alqahtani QH, Haspula D, Alhamed AS, Alqinyah M, Raish M. The Potential Protective Role of Naringenin against Dasatinib-Induced Hepatotoxicity. Pharmaceuticals. 2023; 16(7):921. https://doi.org/10.3390/ph16070921
Chicago/Turabian StyleAlanazi, Ahmed Z., Khalid Alhazzani, Salah Q. Alrewily, Khaldoon Aljerian, Mohammad M. Algahtani, Qamraa H. Alqahtani, Dhanush Haspula, Abdullah S. Alhamed, Mohammed Alqinyah, and Mohammad Raish. 2023. "The Potential Protective Role of Naringenin against Dasatinib-Induced Hepatotoxicity" Pharmaceuticals 16, no. 7: 921. https://doi.org/10.3390/ph16070921
APA StyleAlanazi, A. Z., Alhazzani, K., Alrewily, S. Q., Aljerian, K., Algahtani, M. M., Alqahtani, Q. H., Haspula, D., Alhamed, A. S., Alqinyah, M., & Raish, M. (2023). The Potential Protective Role of Naringenin against Dasatinib-Induced Hepatotoxicity. Pharmaceuticals, 16(7), 921. https://doi.org/10.3390/ph16070921