
Recently Approved Drugs for Lowering and Controlling Intraocular Pressure to Reduce Vision Loss in Ocular Hypertensive and Glaucoma Patients

Abstract
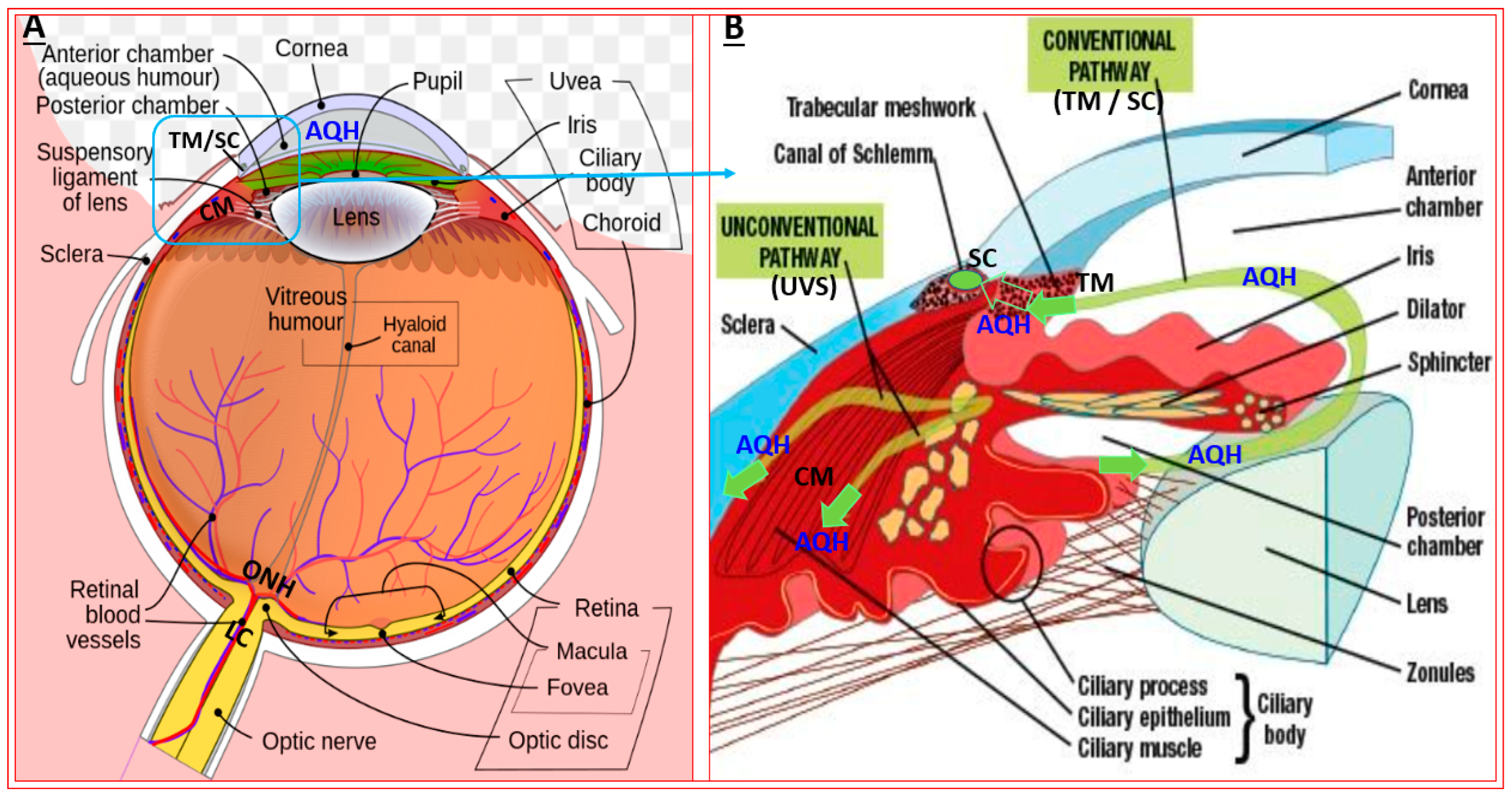
1. Introduction
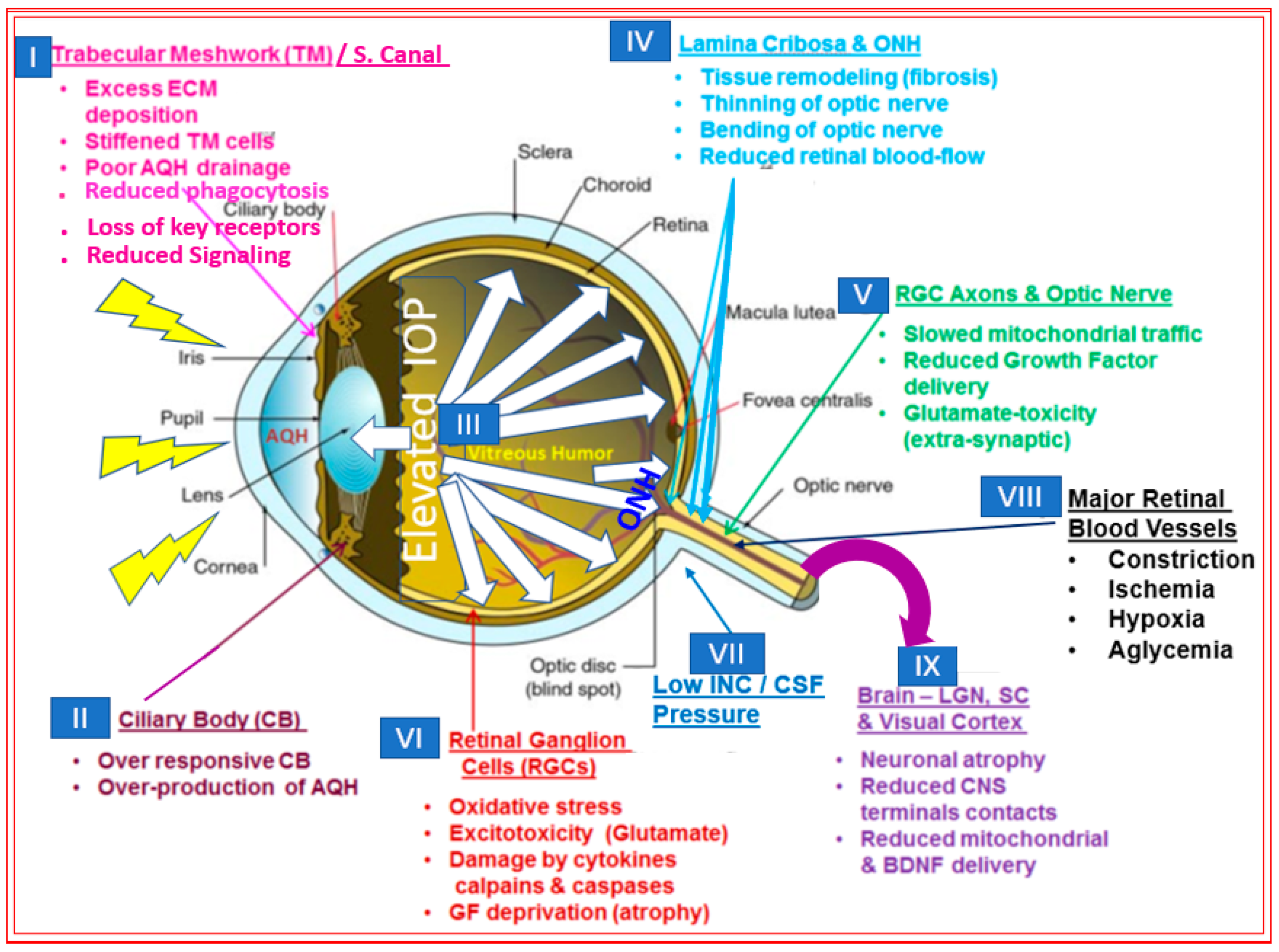
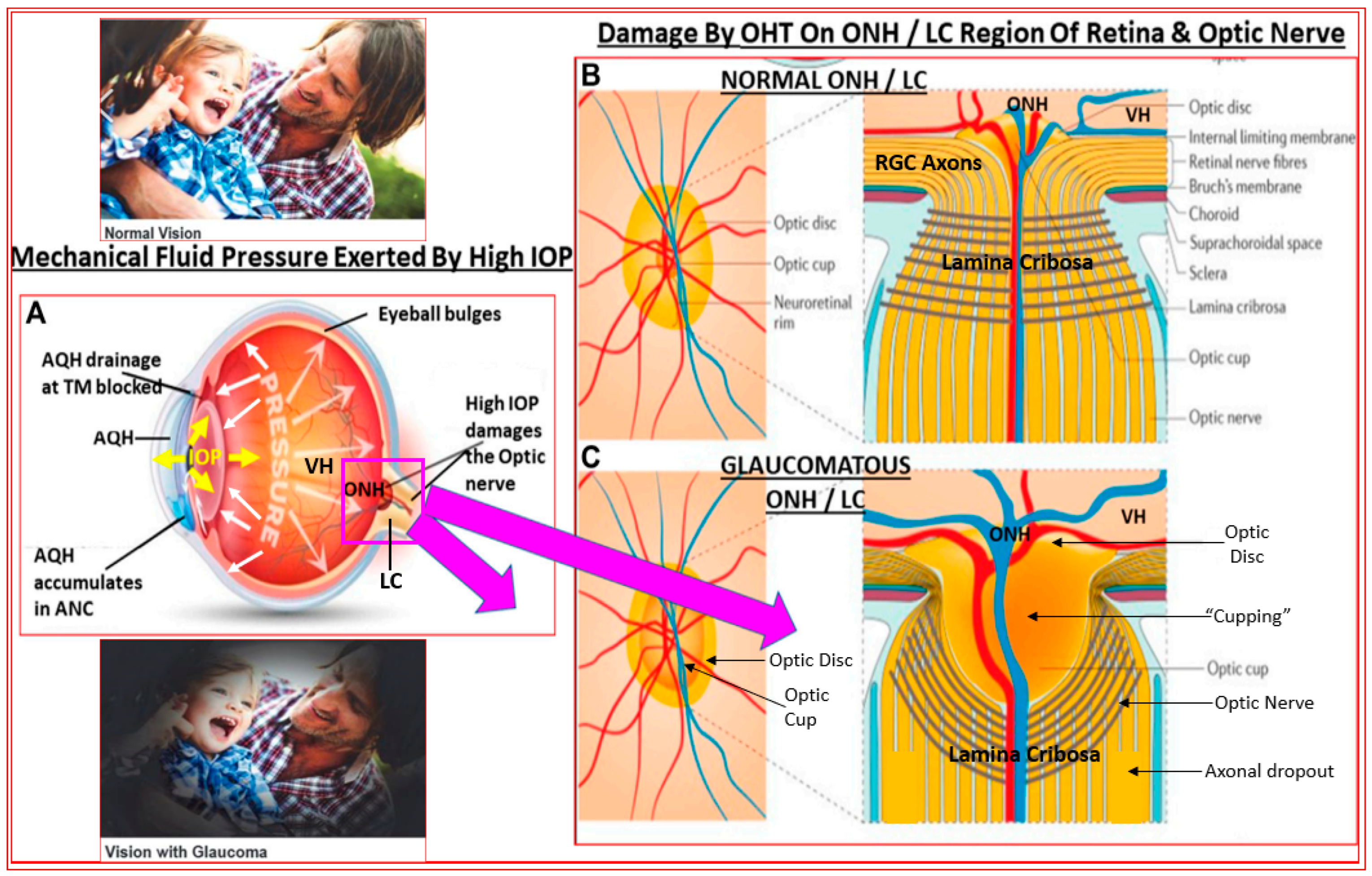
2. Open-Angle Glaucoma
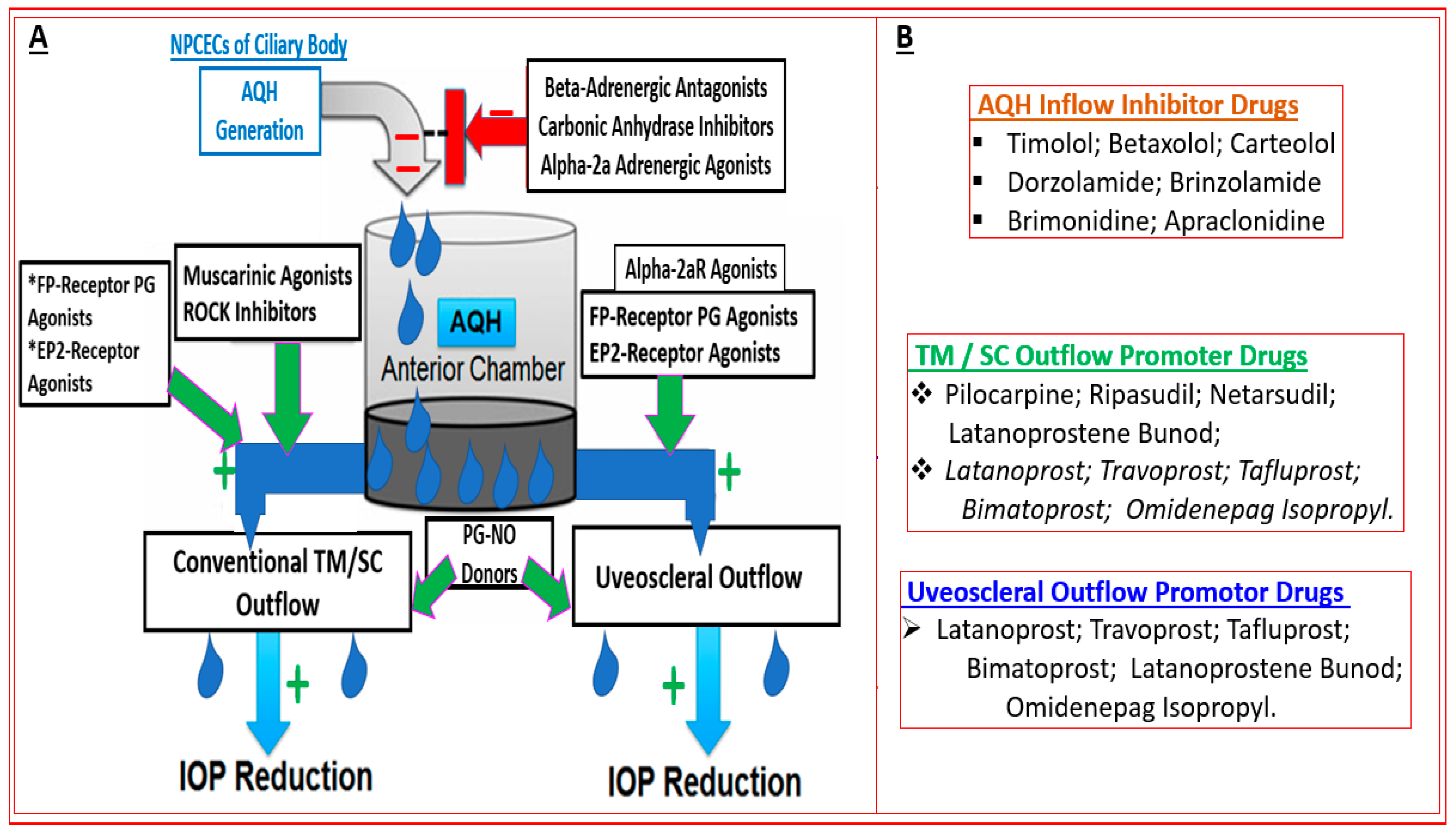
3. Major Methods to Lower and Control IOP/OHT
4. Recently Approved Drug-Based Therapeutics for OAG/OHT Treatment
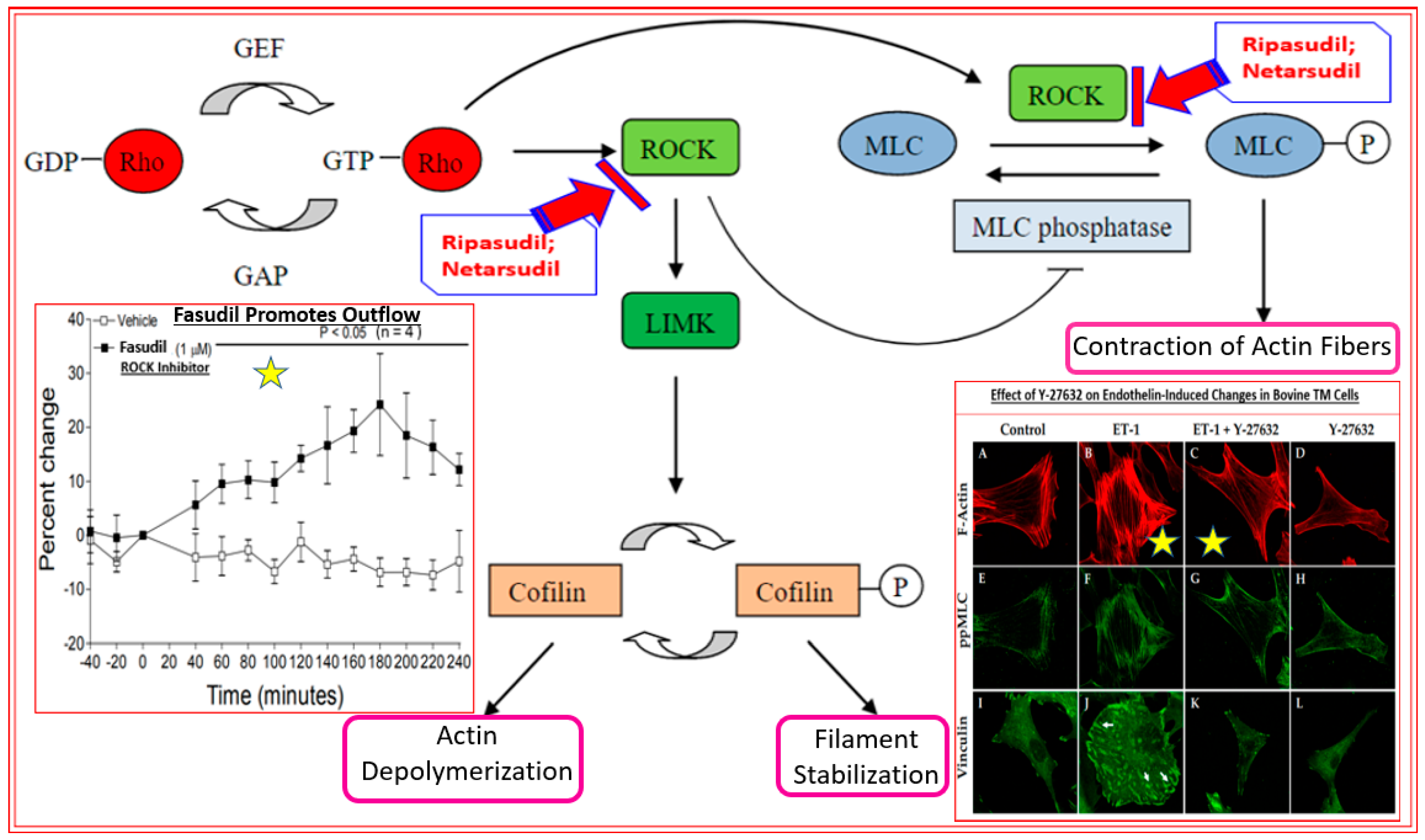
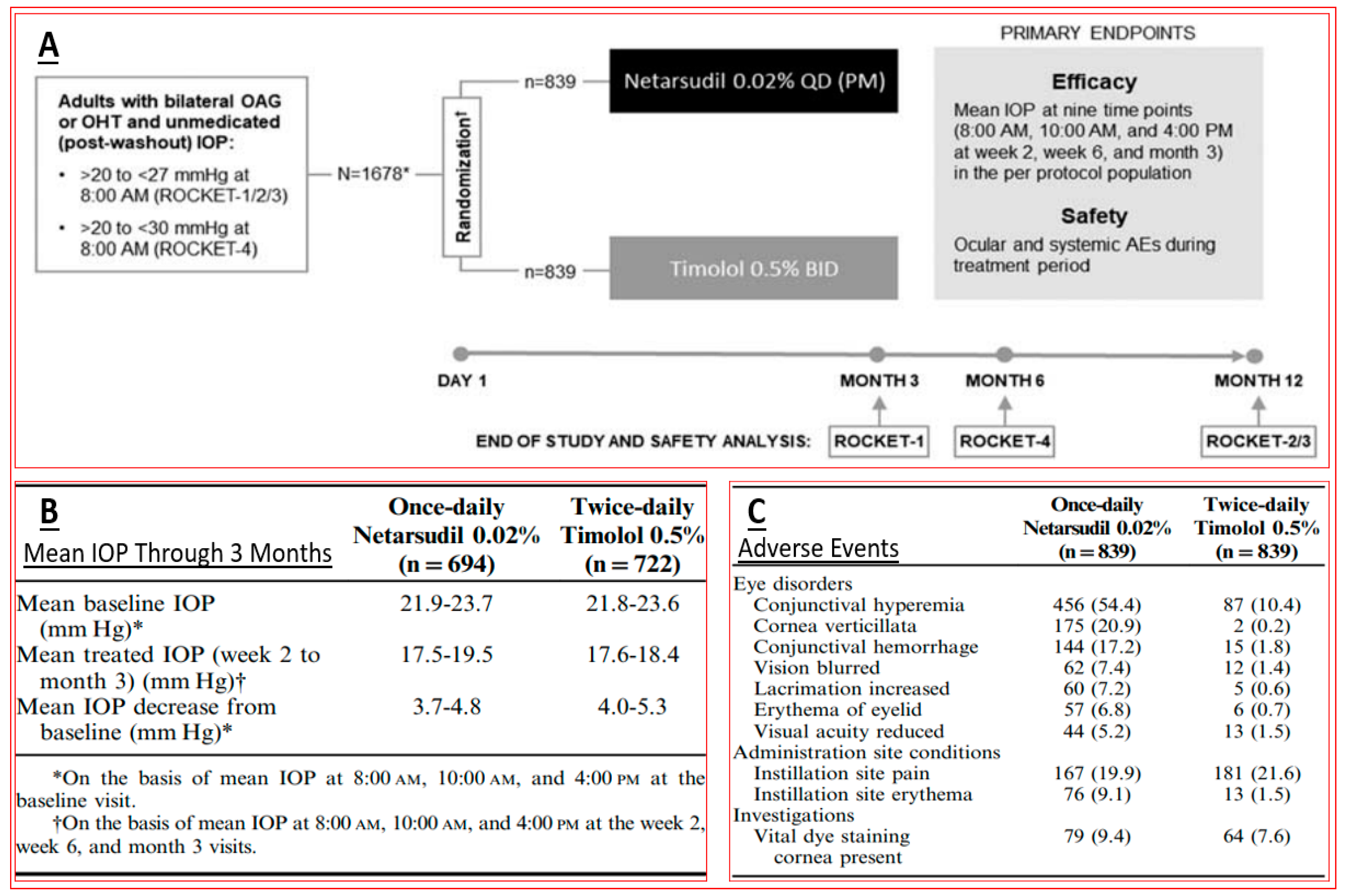
4.1. Rho Kinase Inhibitors (Ripasudil and Netarsudil)
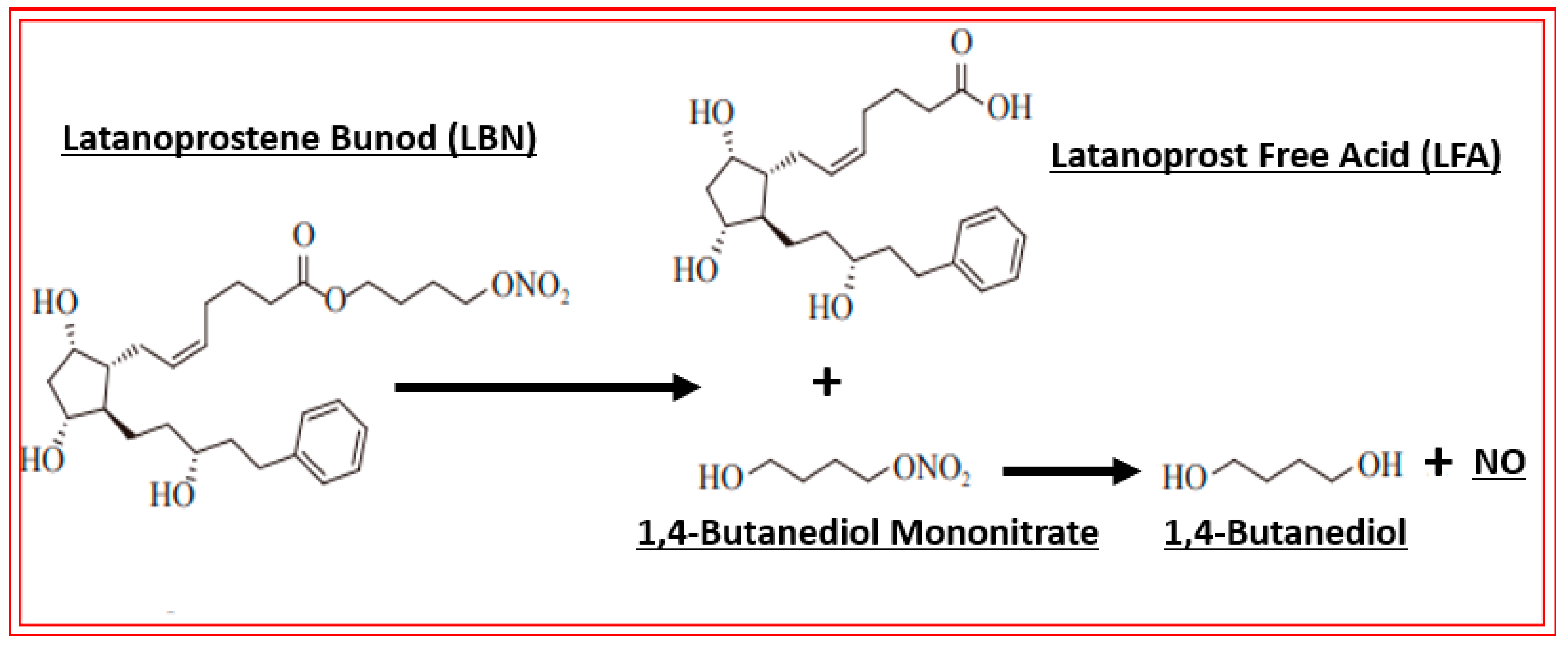
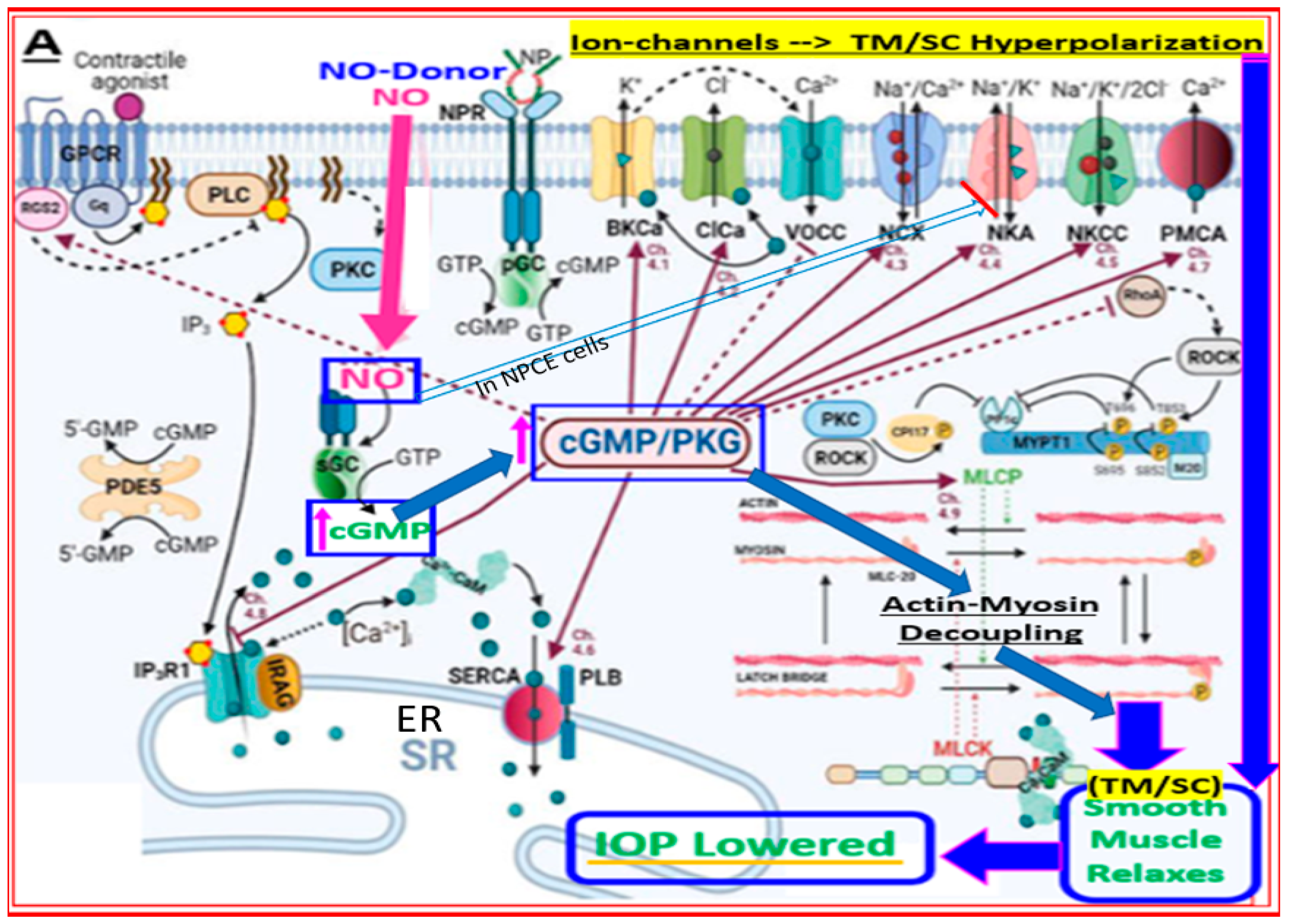
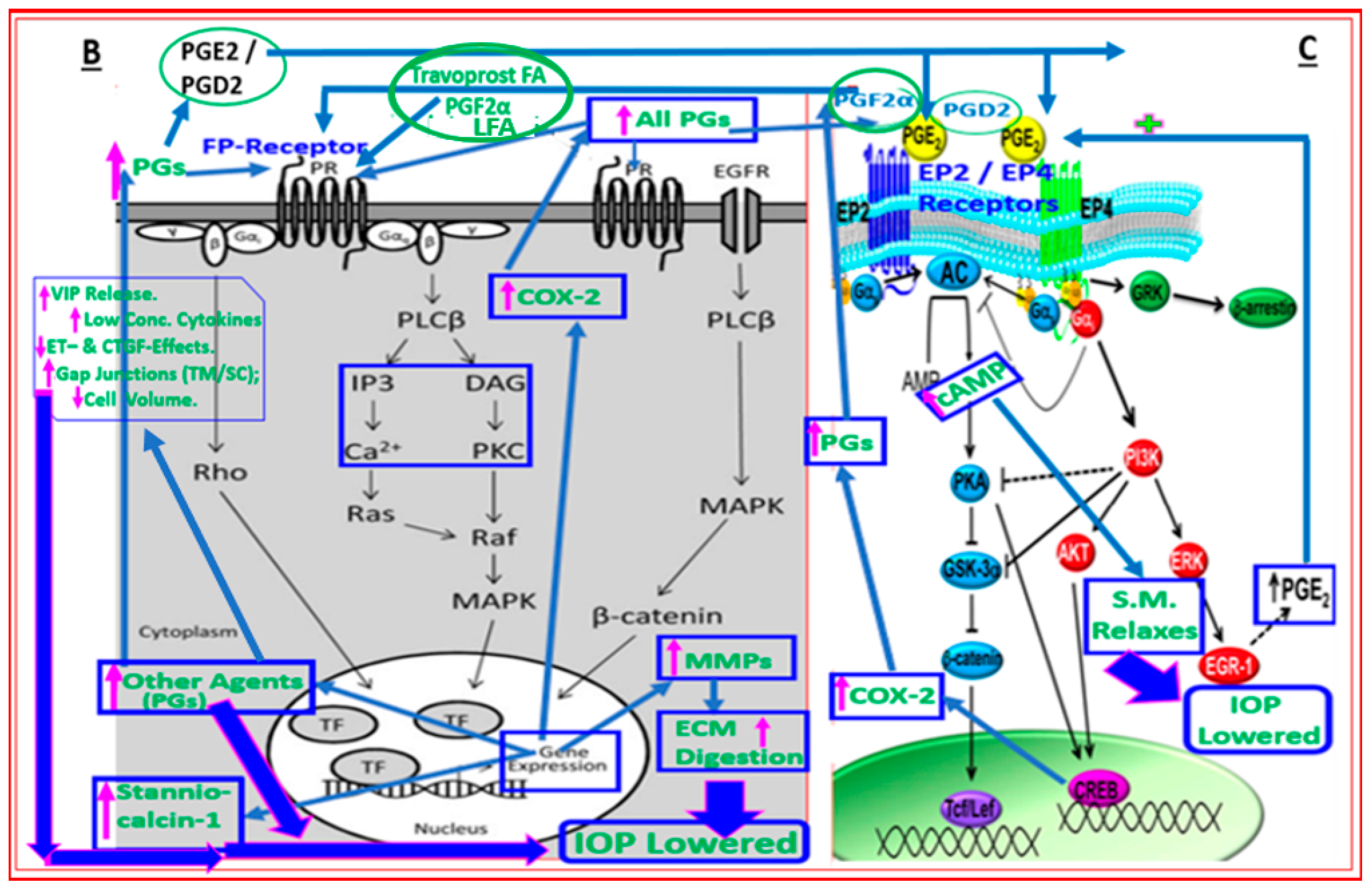
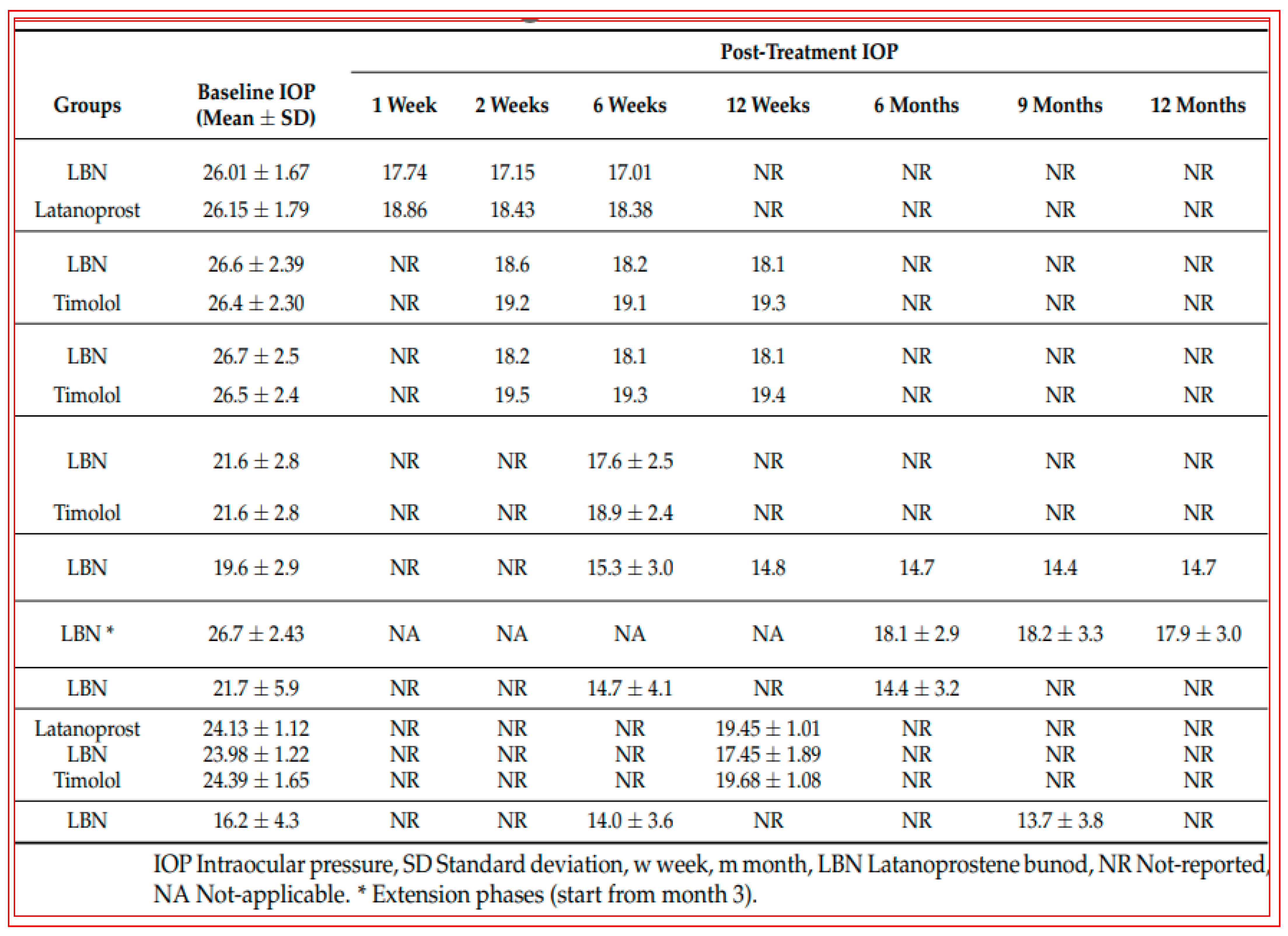
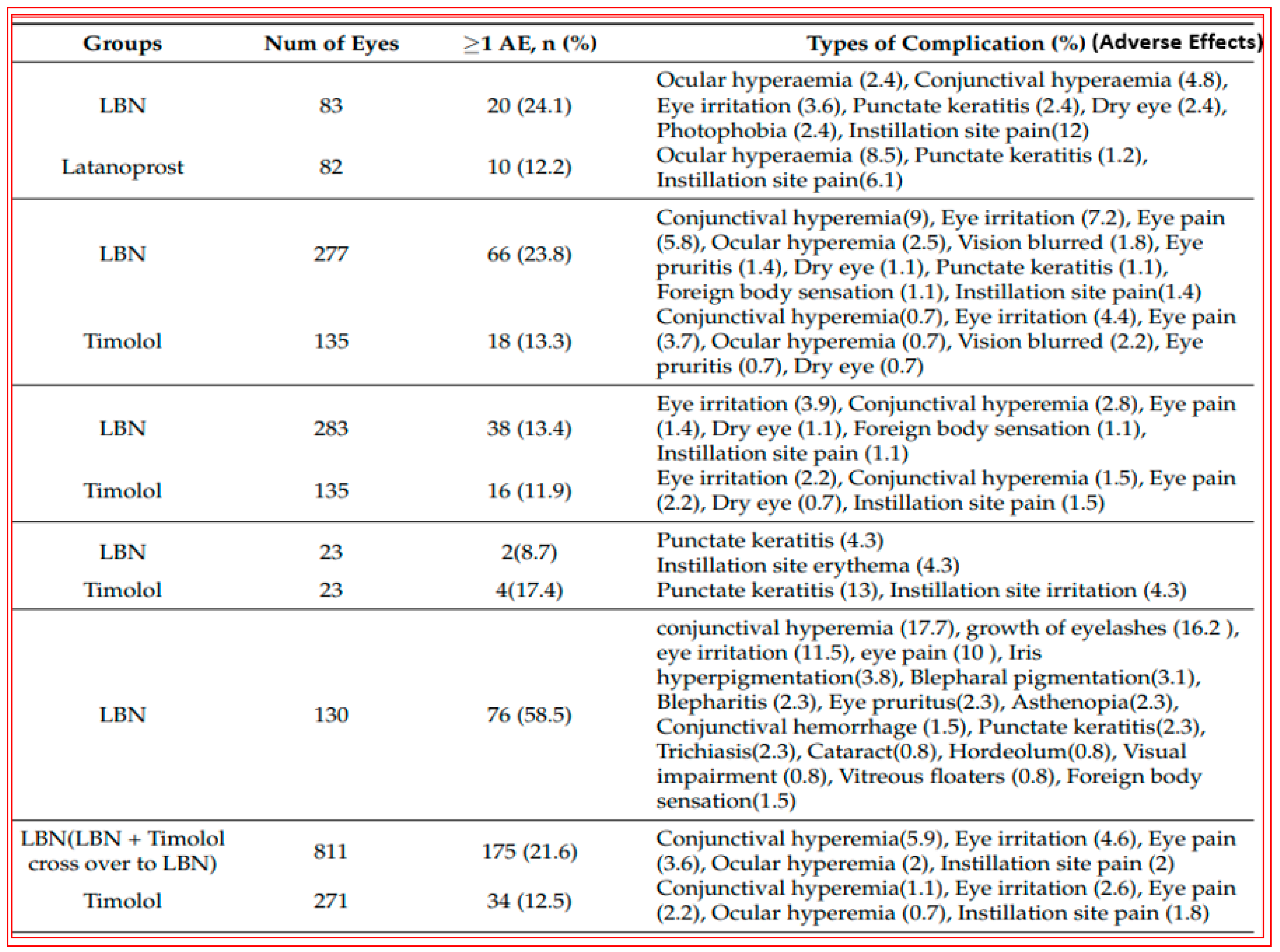
4.2. Conjugate of Latanoprost and NO-Donor (Latanoprostene Bunod)
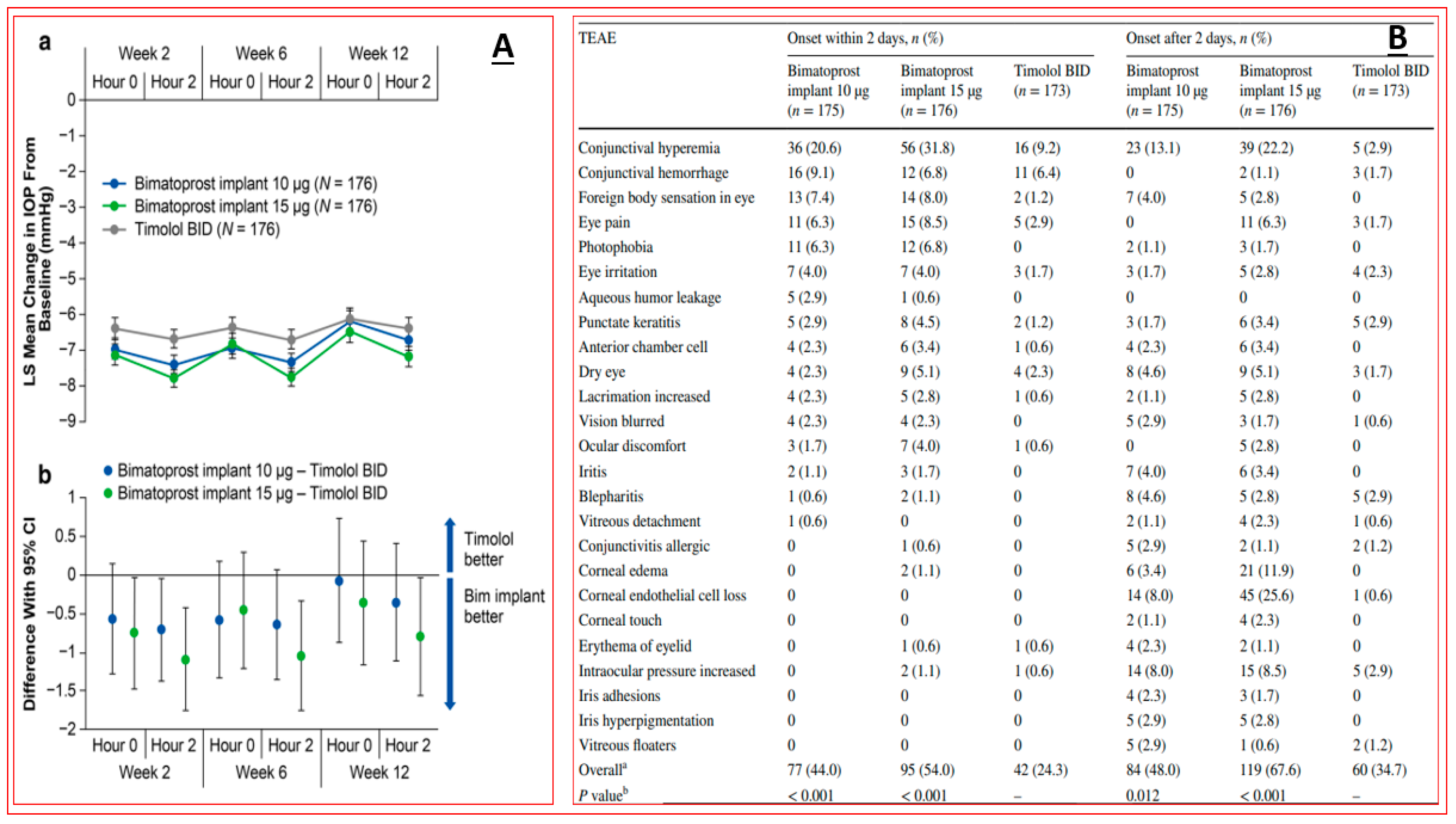
4.3. Sustained Delivery Bimatoprost Implant
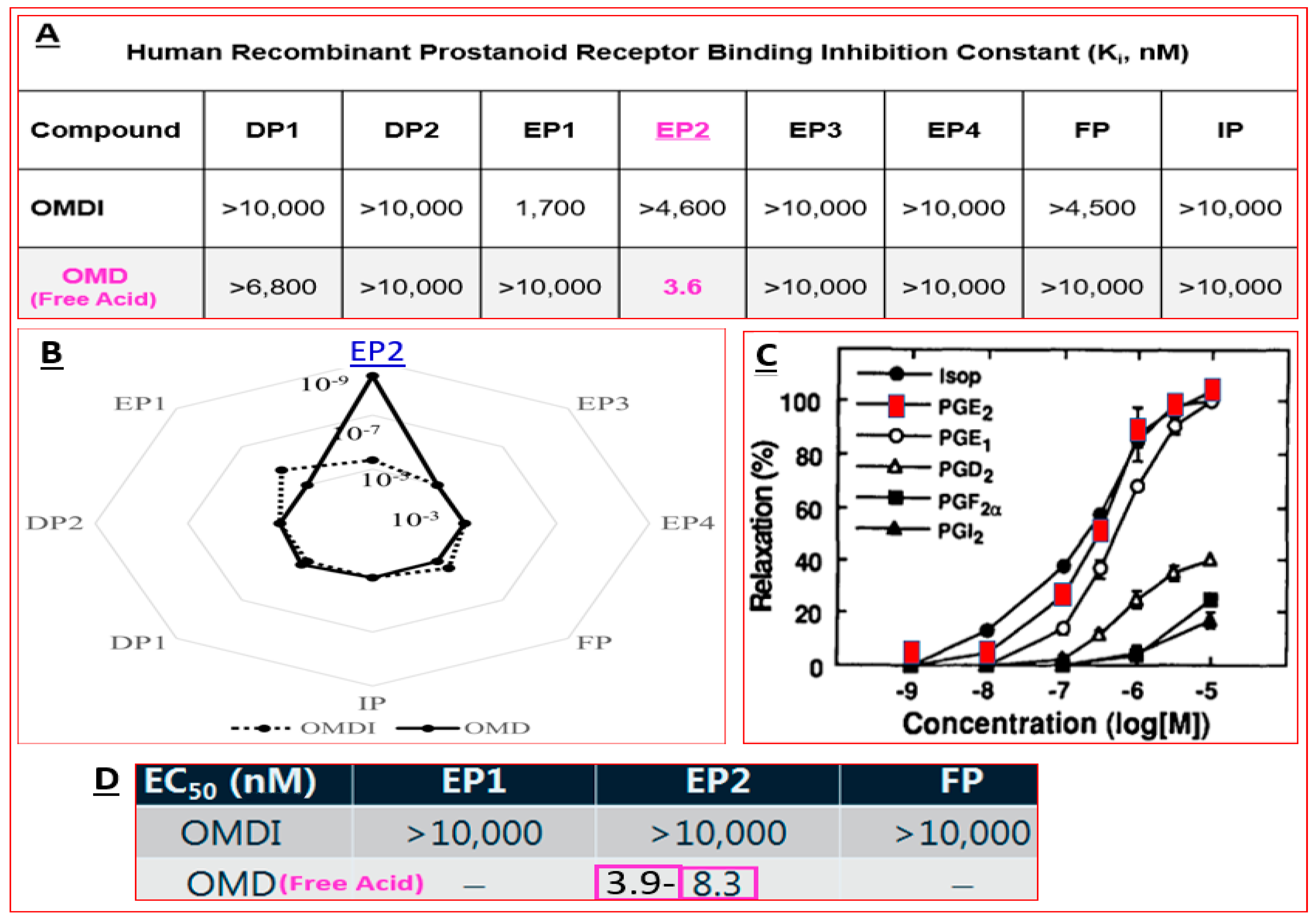
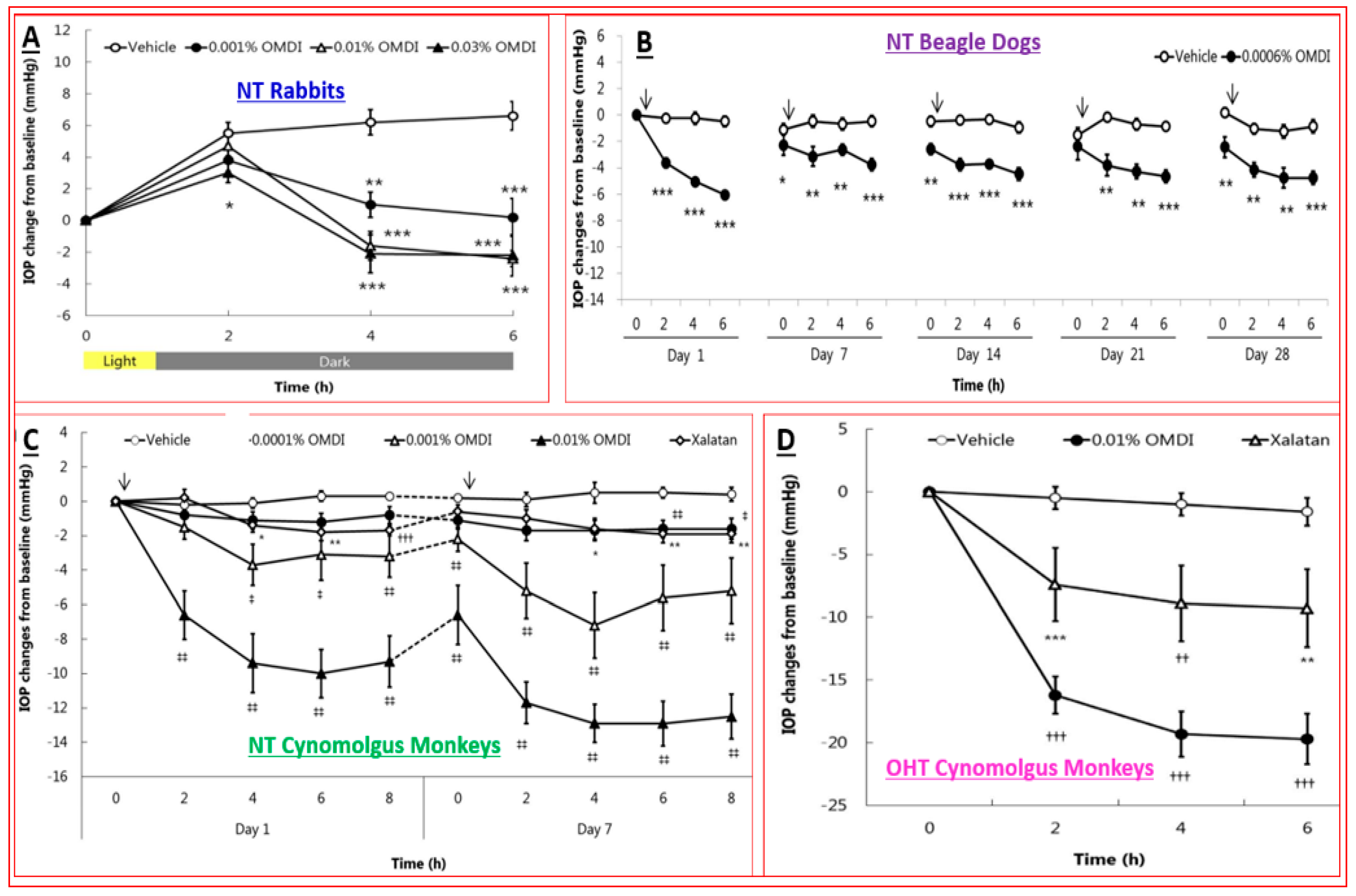
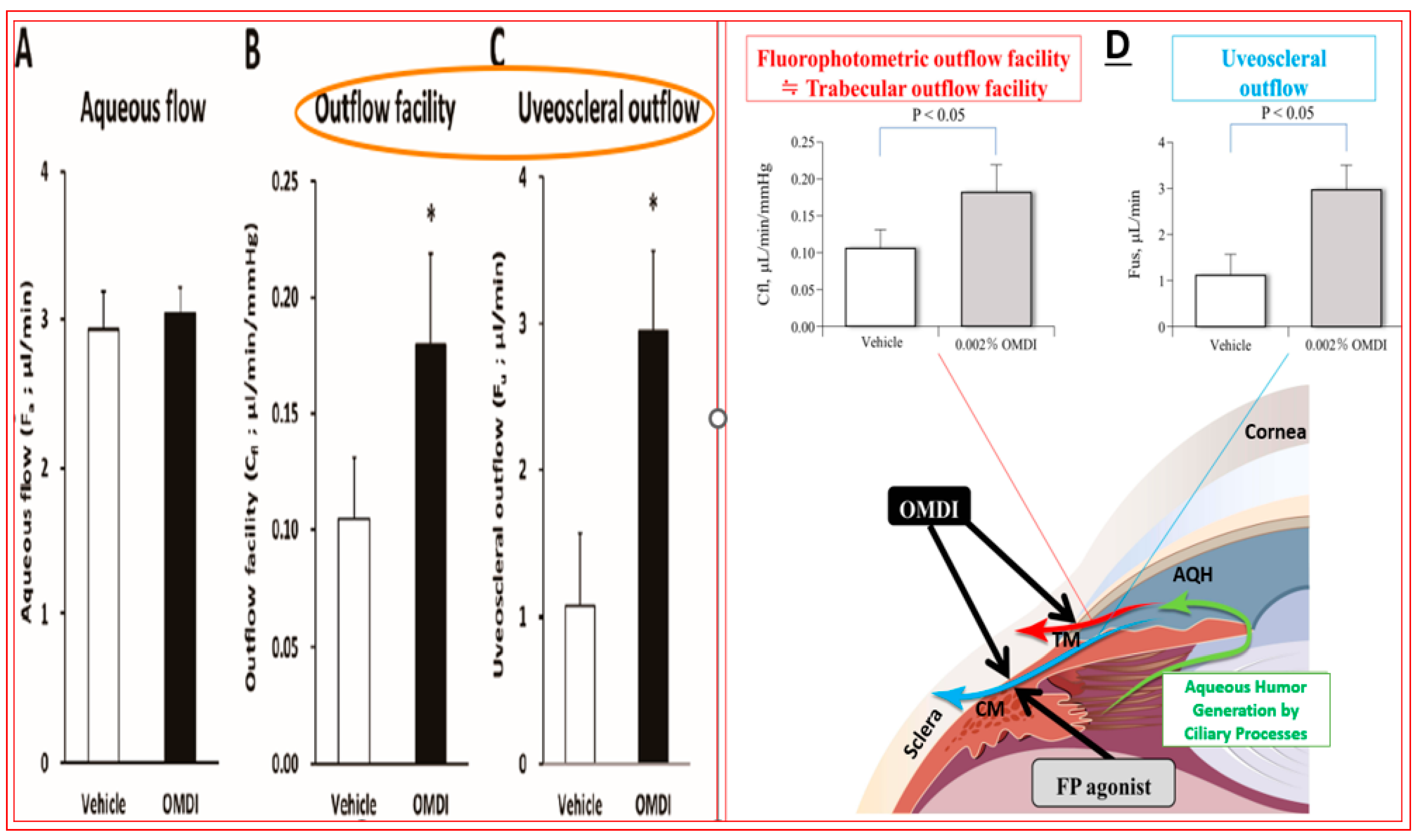
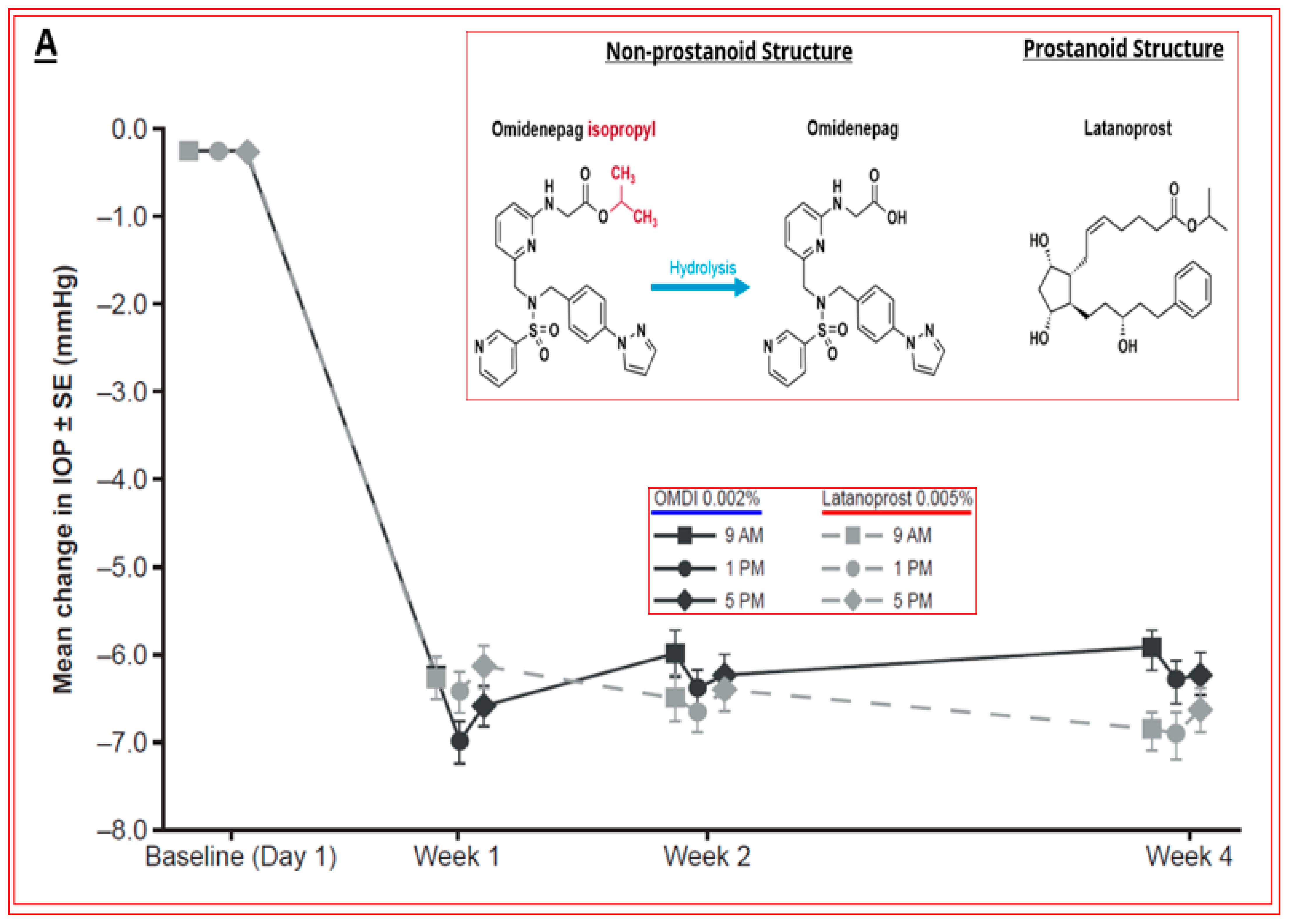
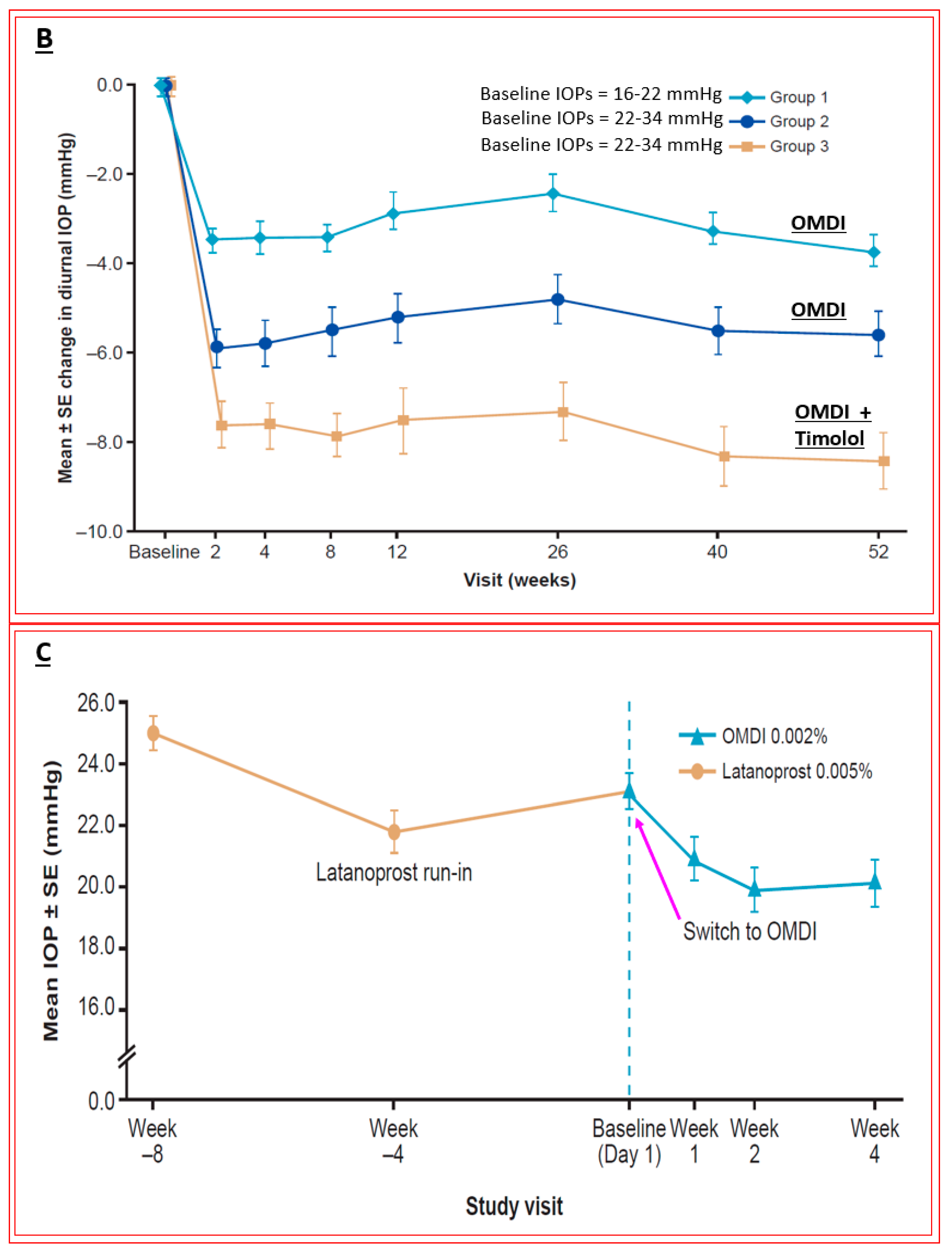
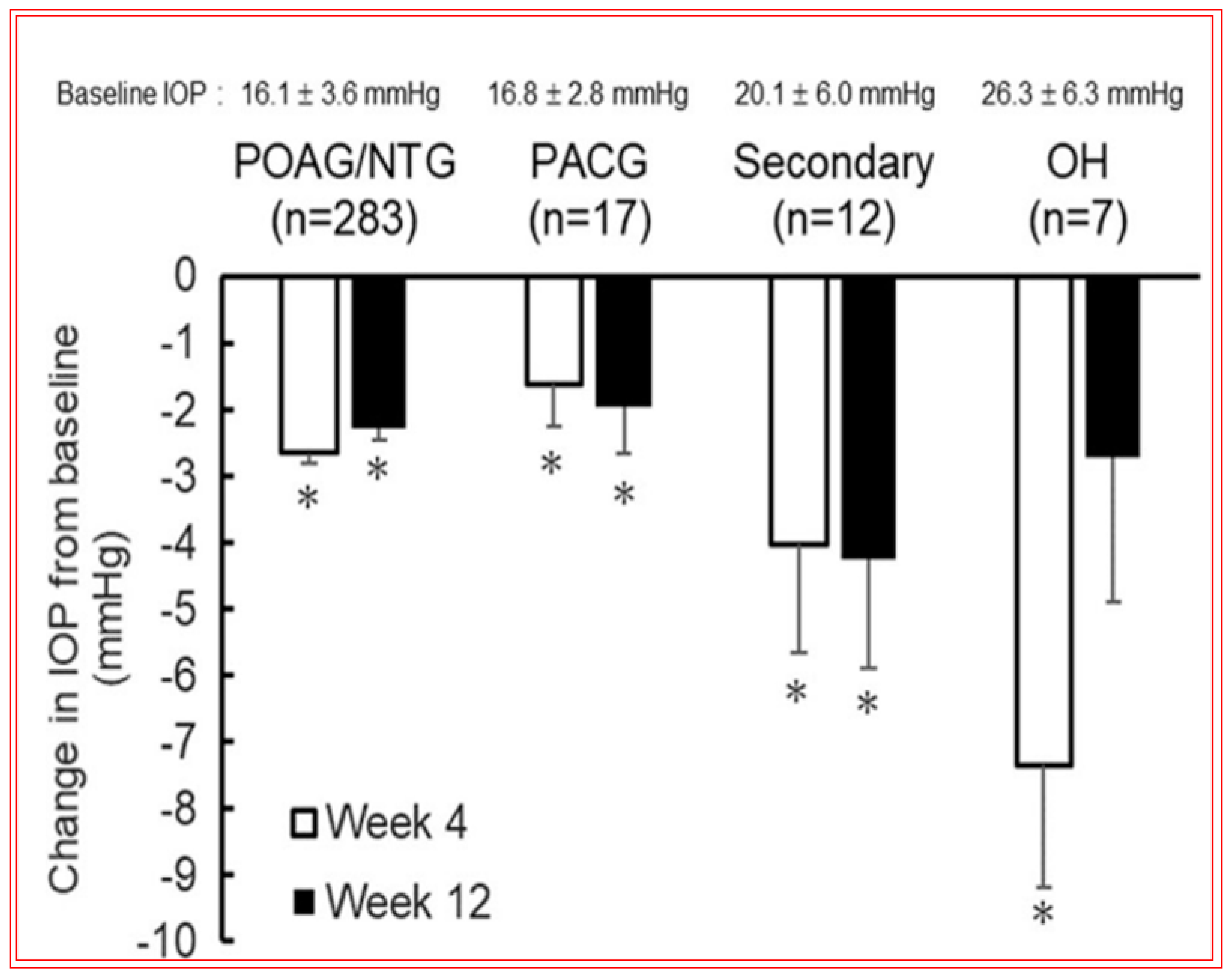
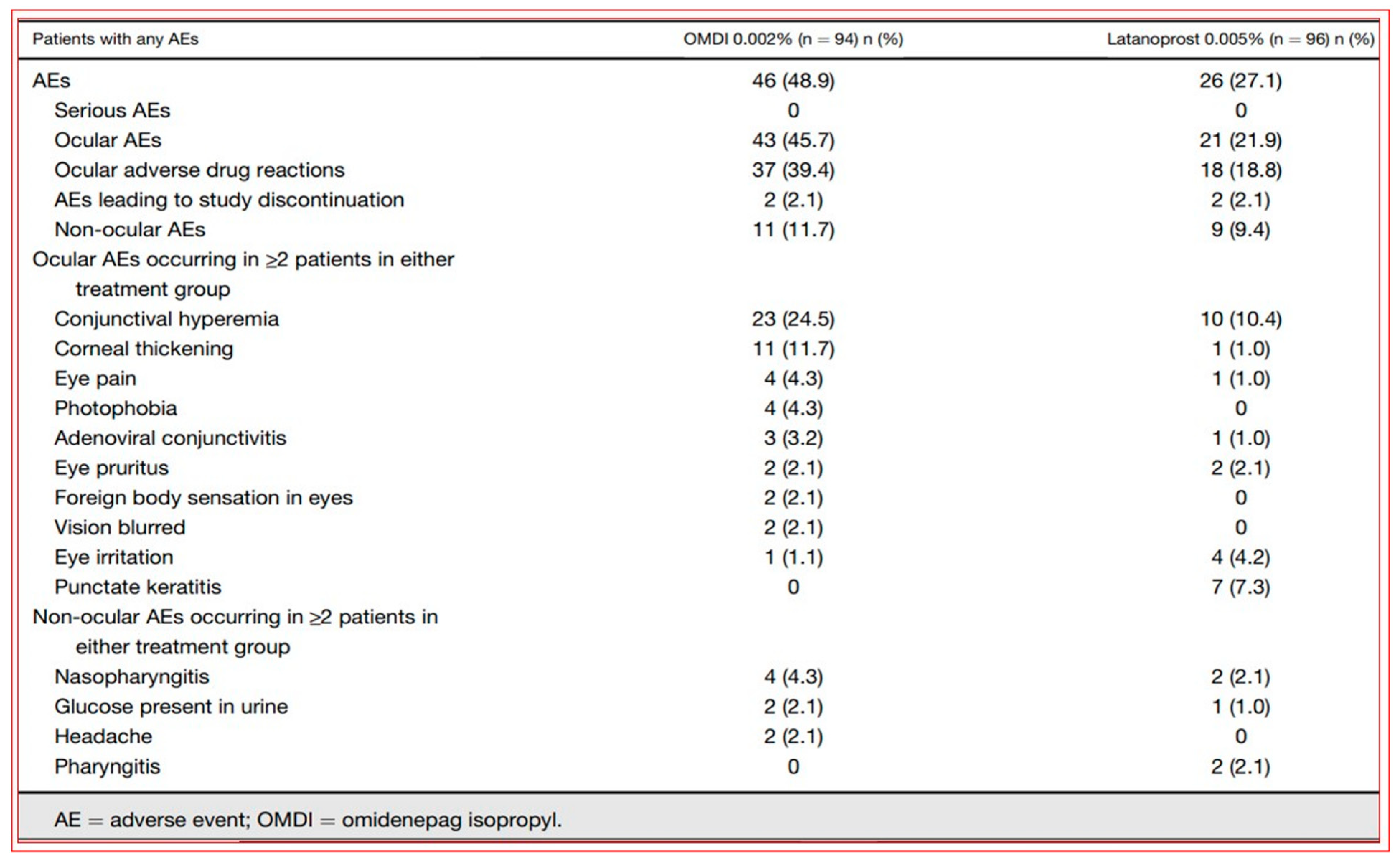
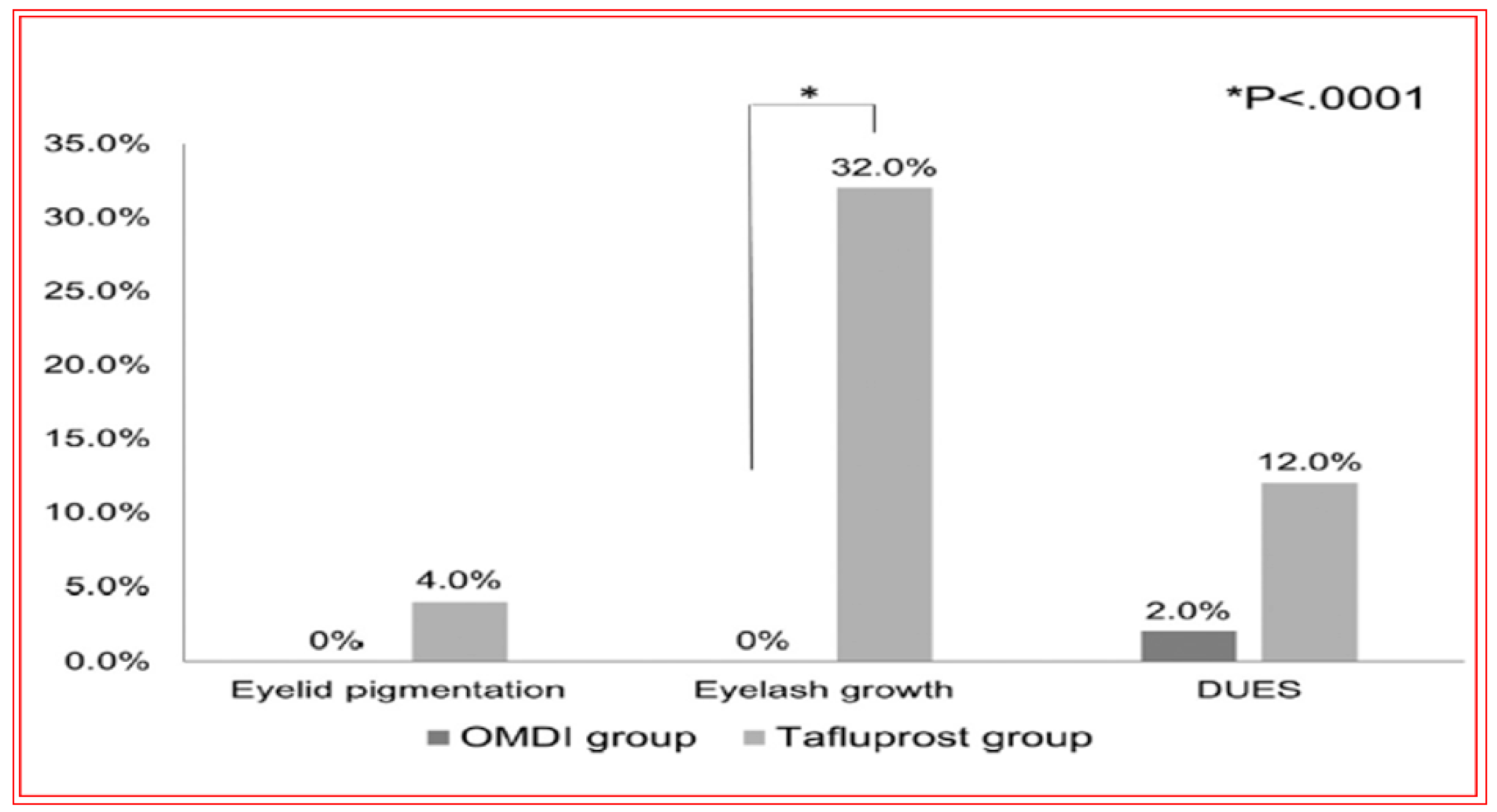
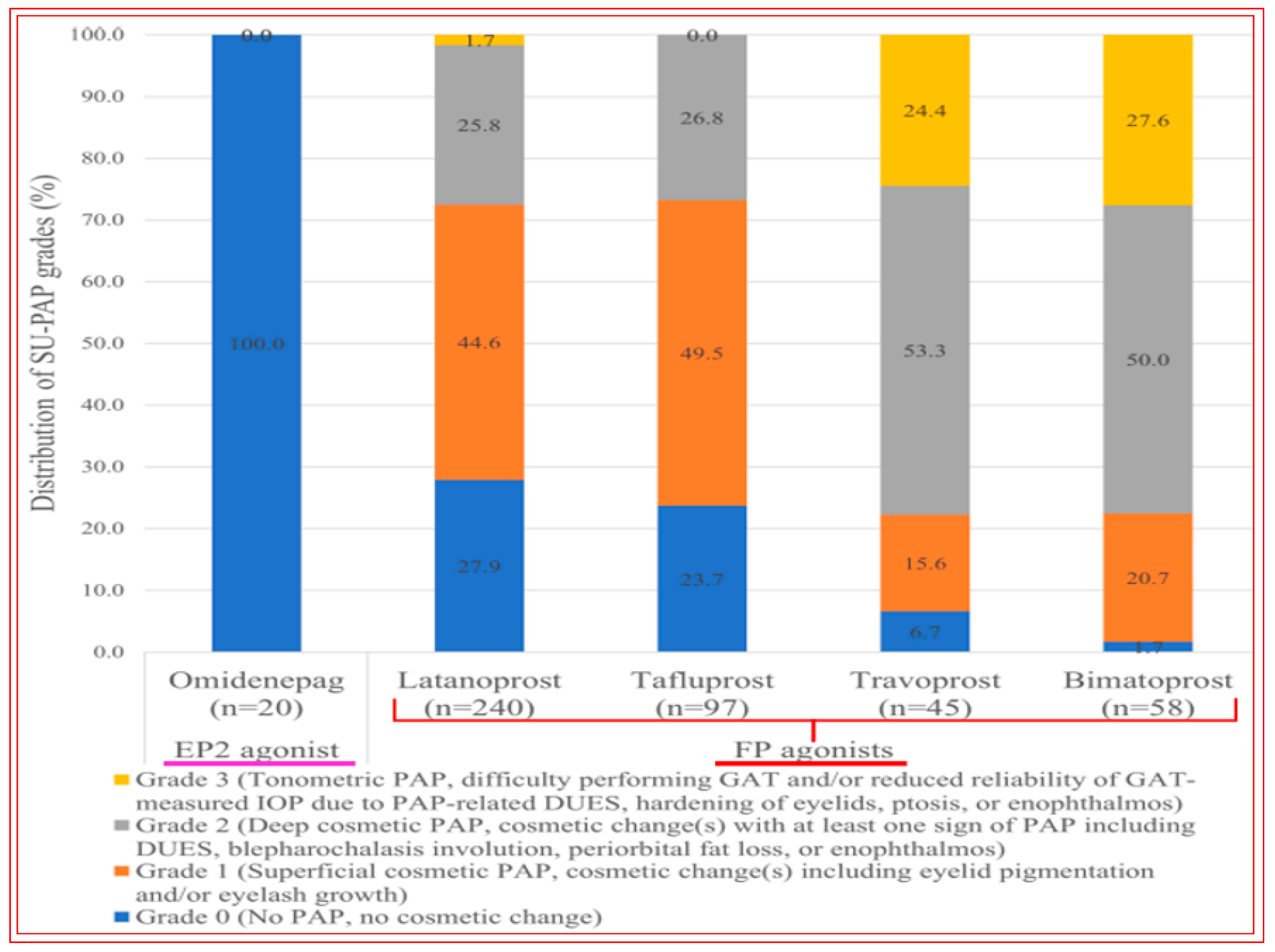
4.4. Novel Non-Prostaglandin EP2-Prostanoid Receptor Agonist (Omidenepag Isopropyl)
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Blindness and Vision Impairment. Fact Sheets. WHO Priority Eye Diseases. Available online: https://www.who.int/blindness/causes/priority/en/ (accessed on 2 December 2022).
- Burton, M.J.; Ramke, J.; Marques, A.P.; Bourne, R.R.A.; Congdon, N.; Jones, I.; Ah Tong, B.A.M.; Arunga, S.; Bachani, D.; Bascaran, C.; et al. The Lancet global health commission on global eye health: Vision beyond. Lancet Glob. Health 2020, 9, e489–e551. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Chen, A.; Zou, M.; Zhang, Y.; Jin, L.; Li, Y.; Zheng, D.; Jin, G.; Congdon, N. Time trends, associations and prevalence of blindness and vision loss due to glaucoma: An analysis of observational data from the Global Burden of Disease Study 2017. BMJ Open 2022, 12, e053805. [Google Scholar] [CrossRef] [PubMed]
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The pathophysiology and treatment of glaucoma: A review. JAMA 2014, 311, 1901–1911. [Google Scholar] [CrossRef]
- Jonas, J.B.; Aung, T.; Bourne, R.R.; Bron, A.M.; Ritch, R.; Panda-Jonas, S. Glaucoma. Lancet 2017, 390, 2183–2193. [Google Scholar] [CrossRef]
- Sharif, N.A. Therapeutic Drugs and Devices for Tackling Ocular Hypertension and Glaucoma, and Need for Neuroprotection and Cytoprotective Therapies. Front. Pharmacol. 2021, 12, 729249. [Google Scholar] [CrossRef]
- European Glaucoma Society terminology and guidelines for glaucoma. Br. J. Ophthalmol. 2021, 105 (Suppl. S1), 1–169. [CrossRef]
- Chan, P.P.; Pang, J.C.; Tham, C.C. Acute primary angle closure-treatment strategies, evidence and economic considerations. Eye 2019, 33, 110–119. [Google Scholar] [CrossRef]
- Mallick, J.; Devi, L.; Malik, P.K.; Mallick, J. Update on normal tension glaucoma. J. Ophthalmic Vis. Res. 2016, 11, 204–208. [Google Scholar] [CrossRef]
- Collaborative Normal-Tension Glaucoma Study Group. Comparison of glaucomatous progression between untreated patients with normal-tension glaucoma and patients with therapeutically reduced intraocular pressures. Am. J. Ophthalmol. 1998, 126, 487–497. [Google Scholar] [CrossRef]
- Collaborative Normal-Tension Glaucoma Study Group. The effectiveness of intraocular pressure reduction in the treatment of normal-tension glaucoma. Am. J. Ophthalmol. 1998, 126, 498–505. [Google Scholar] [CrossRef] [PubMed]
- AGIS Investigators The Advanced Glaucoma Intervention Study (AGIS): The relationship between control of intraocular pressure and visual field deterioration. Am. J. Ophthalmol. 2000, 130, 429–440. [CrossRef] [PubMed]
- Kass, M.A.; Heuer, D.K.; Higginbotham, E.J.; Johnson, C.A.; Keltner, J.L.; Miller, J.P.; Parrish, R.K., 2nd; Wilson, M.R.; Gordon, M.O. The Ocular Hypertension Treatment Study: A randomized trial determines that topical ocular hypotensive medication delays or prevents the onset of primary open-angle glaucoma. Arch. Ophthalmol. 2002, 120, 701–713, discussion 829–830. [Google Scholar] [CrossRef] [PubMed]
- Tezel, G.; Siegmund, K.D.; Trinkaus, K.; Wax, M.B.; Kass, M.A.; Kolker, A.E. Clinical factors associated with progression of glaucomatous optic disc damage in treated patients. Arch. Ophthalmol. 2001, 119, 813–818. [Google Scholar] [CrossRef]
- Gordon, M.O.; Beiser, J.A.; Brandt, J.D.; Heuer, D.K.; Higginbotham, E.J.; Johnson, C.A.; Keltner, J.L.; Miller, J.P.; Parrish, R.K., 2nd; Wilson, M.R.; et al. The Ocular Hypertension Treatment Study: Baseline factors that predict the onset of primary open-angle glaucoma. Arch. Ophthalmol. 2002, 120, 714–720, discussion 829–830. [Google Scholar] [CrossRef]
- Heijl, A.; Leske, M.C.; Bengtsson, B.; Hyman, L.; Bengtsson, B.; Hussein, M.; Early Manifest Glaucoma Trial Group. Reduction of intraocular pressure and glaucoma progression: Results from the Early Manifest Glaucoma Trial. Arch. Ophthalmol. 2002, 120, 1268–1279. [Google Scholar] [CrossRef]
- Leske, M.C.; Heijl, A.; Hyman, L.; Bengtsson, B.; Komaroff, E. Predictors of long-term progression in the early manifest glaucoma trial. Ophthalmology 2007, 114, 1965–1972. [Google Scholar] [CrossRef]
- Bengtsson, B.; Leske, M.C.; Hyman, L.; Heijl, A. Fluctuation of intraocular pressure and glaucoma progression in the early manifest glaucoma trial. Ophthalmology 2007, 114, 205–209. [Google Scholar] [CrossRef]
- Garway-Heath, D.F.; Crabb, D.P.; Bunce, C.; Lascaratos, G.; Amalfitano, F.; Anand, N.; Azuara-Blanco, A.; Bourne, R.R.; Broadway, D.C.; Cunliffe, I.A.; et al. Latanoprost for open-angle glaucoma (UKGTS): A randomised, multicentre, placebo-controlled trial. Lancet 2015, 385, 1295–1304. [Google Scholar] [CrossRef]
- Sehi, M.; Grewal, D.S.; Goodkin, M.L.; Greenfield, D.S. Reversal of retinal ganglion cell dysfunction after surgical reduction of intraocular pressure. Ophthalmology 2010, 117, 2329–2336. [Google Scholar] [CrossRef]
- Lee, E.J.; Kim, T.-W.; Weinreb, R.N.; Kim, H. Reversal of lamina cribrosa displacement after intraocular pressure reduction in open-angle glaucoma. Ophthalmology 2013, 120, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Bucolo, C.; Platania, C.B.M.; Drago, F.; Bonfiglio, V.; Reibaldi, M.; Avitabile, T.; Uva, M. Novel therapeutics in glaucoma management. Curr. Neuropharmacol. 2018, 16, 978–992. [Google Scholar] [CrossRef] [PubMed]
- Lusthaus, J.; Goldberg, I. Current management of glaucoma. Med. J. Aust. 2019, 210, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, T.M.; Sit, A.J. Emerging drugs for the treatment of glaucoma: A review of phase II & III trials. Expert Opin. Emerg. Drugs 2022, 27, 321–331. [Google Scholar] [PubMed]
- Civan, M.; Macknight, A.D. The ins and outs of aqueous humor secretion. Exp. Eye Res. 2004, 78, 625–631. [Google Scholar] [CrossRef]
- Acott, T.S.; Vranka, J.A.; Keller, K.E.; Raghunathan, V.; Kelley, M.J. Normal and glaucomatous outflow regulation. Prog. Retin. Eye Res. 2020, 11, 100897. [Google Scholar] [CrossRef]
- Carreon, T.; van der Merwe, E.; Fellman, R.L.; Johnstone, M.; Bhattacharya, S.K. Aqueous outflow—A continuum from trabecular meshwork to episcleral veins. Prog. Retin. Eye Res. 2017, 57, 108–133. [Google Scholar] [CrossRef]
- Abu-Hassan, D.W.; Acott, T.S.; Kelley, M.J. The trabecular meshwork: A basic review of form and function. J. Ocul. Biol. 2014, 2, 9. [Google Scholar]
- Buffault, J.; Labbé, A.; Hamard, P.; Brignole-Baudouin, F.; Baudouin, C. The trabecular meshwork: Structure, function and clinical implications. A review of the literature. J. Fr. Ophtalmol. 2020, 43, e217–e230. [Google Scholar] [CrossRef]
- Van Zyl, T.; Yan, W.; McAdams, A.; Peng, Y.R.; Shekhar, K.; Regev, A.; Juric, D.; Sanes, J.R. Cell atlas of aqueous humor outflow pathways in eyes of humans and four model species provides insight into glaucoma pathogenesis. Proc. Natl. Acad. Sci. USA 2020, 117, 10339–10349. [Google Scholar] [CrossRef]
- Patel, G.; Fury, W.; Yang, H.; Adler, C.; Wei, Y.; Ni, M.; Schmitt, H.; Hu, Y.; Yancopoulos, G.; Stamer, W.D.; et al. Molecular taxonomy of human ocular outflow tissues defined by single-cell transcriptomics. Proc. Natl. Acad. Sci. USA 2020, 117, 12856–12867. [Google Scholar] [CrossRef] [PubMed]
- Overby, D.R.; Zhou, E.H.; Vargas-Pinto, R.; Pedrigi, R.M.; Fuchshofer, R.; Braakman, S.T.; Gupta, R.; Perkumas, K.M.; Sherwood, J.M.; Vahabikashi, A.; et al. Altered mechanobiology of Schlemm’s canal endothelial cells in glaucoma. Proc. Natl. Acad. Sci. USA 2014, 111, 13876–13881. [Google Scholar] [CrossRef] [PubMed]
- Stamer, W.D.; Braakman, S.T.; Zhou, E.H.; Ethier, C.R.; Fredberg, J.J.; Overby, D.R.; Johnson, M. Biomechanics of Schlemm’s canal endothelium and intraocular pressure reduction. Prog. Retin. Eye Res. 2015, 44, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-Y.; Su, G.-Y.; Wei, Z.-Y.; Zhang, Z.J.; Liang, Q.F. Progress in the basic and clinical research on the Schlemm’s canal. Int. J. Ophthalmol. 2020, 13, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Lewczuk, K.; Jabłońska, J.; Konopińska, J.; Mariak, Z.; Rękas, M. Schlemm’s canal: The outflow ‘vessel’. Acta Ophthalmol. 2022, 100, e881–e890. [Google Scholar] [CrossRef] [PubMed]
- Wiederholt, M.; Thieme, H.; Stumpff, F. The regulation of trabecular meshwork and ciliary muscle contractility. Prog. Retin. Eye Res. 2000, 19, 271–295. [Google Scholar] [CrossRef]
- Kim, J.H.; Caprioli, J. Intraocular pressure fluctuation: Is it important? J. Ophthalmic Vis. Res. 2018, 3, 170–174. [Google Scholar]
- Jasien, J.V.; Turner, D.C.; Girkin, C.A.; Downs, J.C. Cyclic pattern of intraocular pressure (IOP) and transient IOP fluctuations in nonhuman primates measured with continuous wireless telemetry. Curr. Eye Res. 2018, 44, 1244–1252. [Google Scholar] [CrossRef]
- Alvarado, J.; Murphy, C.; Juster, R. Trabecular meshwork cellularity in primary open-angle glaucoma and nonglaucomatous normals. Ophthalmology 1984, 91, 564–579. [Google Scholar] [CrossRef]
- Grierson, I.; Howes, R.C. Age-related depletion of the cell population in the human trabecular meshwork. Eye 1987, 1 Pt 2, 204–210. [Google Scholar] [CrossRef]
- Keller, K.E.; Peters, D.M. Pathogenesis of glaucoma: Extracellular matrix dysfunction in the trabecular meshwork—A review. Clin. Exp. Ophthalmol. 2022, 50, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Kasetti, R.B.; Phan, T.N.; Millar, J.C.; Zode, G.S. Expression of mutant myocilin induces abnormal intracellular accumulation of selected extracellular matrix proteins in the trabecular meshwork. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6058–6069. [Google Scholar] [CrossRef] [PubMed]
- Kasetti, R.B.; Maddineni, P.; Millar, J.C.; Clark, A.F.; Zode, G.S. Increased synthesis and deposition of extracellular matrix proteins leads to endoplasmic reticulum stress in the trabecular meshwork. Sci. Rep. 2017, 7, 14951. [Google Scholar] [CrossRef] [PubMed]
- Yemanyi, F.; Vranka, J.; Raghunathan, V.K. Crosslinked extracellular matrix stiffens human trabecular meshwork cells via dysregulating β-catenin and YAP/TAZ signaling pathways. Investig. Ophthalmol. Vis. Sci. 2020, 61, 41. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Johnstone, M.A.; Xin, C.; Padilla, S.; Vranka, J.A.; Acott, T.S.; Zhou, K.; Schwaner, S.A.; Wang, R.K.; Sulchek, T.; et al. Estimating human trabecular meshwork stiffness by numerical modeling and advanced OCT imaging. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4809–4817. [Google Scholar] [CrossRef]
- Borrás, T. A single gene connects stiffness in glaucoma and the vascular system. Exp. Eye Res. 2017, 158, 13–22. [Google Scholar] [CrossRef]
- Morgan, J.T.; Raghunathan, V.K.; Chang, Y.R.; Murphy, C.J.; Russell, P. The intrinsic stiffness of human trabecular meshwork cells increases with senescence. Oncotarget 2015, 6, 15362–15374. [Google Scholar] [CrossRef]
- Bermudez, J.Y.; Montecchi-Palmer, M.; Mao, W.; Clark, A.F. Cross-linked actin networks (CLANs) in glaucoma. Exp. Eye Res. 2017, 159, 16–22. [Google Scholar] [CrossRef]
- Patel, P.D.; Chen, Y.L.; Kasetti, R.B.; Maddineni, P.; Mayhew, W.; Millar, J.C.; Ellis, D.Z.; Sonkusare, S.K.; Zode, G.S. Impaired TRPV4-eNOS signaling in trabecular meshwork elevates intraocular pressure in glaucoma. Proc. Natl. Acad. Sci. USA 2021, 118, e2022461118. [Google Scholar] [CrossRef]
- Lakk, M.; Križaj, D. TRPV4-Rho signaling drives cytoskeletal and focal adhesion remodeling in trabecular meshwork cells. Am. J. Physiol. Cell Physiol. 2021, 320, C1013–C1030. [Google Scholar] [CrossRef]
- Ryskamp, D.A.; Frye, A.M.; Phuong, T.T.; Yarishkin, O.; Jo, A.O.; Xu, Y.; Lakk, M.; Iuso, A.; Redmon, S.N.; Ambati, B.; et al. TRPV4 regulates calcium homeostasis, cytoskeletal remodeling, conventional outflow and intraocular pressure in the mammalian eye. Sci. Rep. 2016, 6, 30583. [Google Scholar] [CrossRef] [PubMed]
- Yarishkin, O.; Phuong, T.T.T.; Baumann, J.M.; De Ieso, M.L.; Vazquez-Chona, F.; Rudzitis, C.N.; Sundberg, C.; Lakk, M.; Stamer, W.D.; Križaj, D. Piezo1 channels mediate trabecular meshwork mechanotransduction and promote aqueous fluid outflow. J. Physiol. 2021, 599, 571–592. [Google Scholar] [CrossRef] [PubMed]
- Dismuke, W.M.; Sharif, N.A.; Ellis, D.Z. Human trabecular meshwork cell volume decrease by NO-independent soluble guanylate cyclase activators YC-1 and BAY-58-2667 involves the BKCa ion channel. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3353–3359. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ellis, D.Z.; Sharif, N.A.; Dismuke, W.M. Endogenous regulation of human Schlemm’s canal cell volume by nitric oxide signaling. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5817–5824. [Google Scholar] [CrossRef]
- Nathanson, J.A. Nitric oxide and nitrovasodilators in the eye: Implications for ocular physiology and glaucoma. J. Glaucoma 1993, 2, 206–210. [Google Scholar] [CrossRef]
- Saccà, S.C.; Pascotto, A.; Camicione, P.; Capris, P.; Izzotti, A. Oxidative DNA damage in the human trabecular meshwork: Clinical correlation in patients with primary open-angle glaucoma. Arch. Ophthalmol. 2005, 123, 458–463. [Google Scholar] [CrossRef]
- He, Y.; Ge, J.; Tombran-Tink, J. Mitochondrial defects and dysfunction in calcium regulation in glaucomatous trabecular meshwork cells. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4912–4922. [Google Scholar] [CrossRef]
- Ying, Y.; Xue, R.; Yang, Y.; Zhang, S.X.; Xiao, H.; Zhu, H.; Li, J.; Chen, G.; Ye, Y.; Yu, M.; et al. Activation of ATF4 triggers trabecular meshwork cell dysfunction and apoptosis in POAG. Aging 2021, 13, 8628–8642. [Google Scholar] [CrossRef]
- Izzotti, A.; Longobardi, M.; Cartiglia, C.; Saccà, S.C. Mitochondrial damage in the trabecular meshwork occurs only in primary open-angle glaucoma and in pseudoexfoliative glaucoma. PLoS ONE 2011, 6, e14567. [Google Scholar] [CrossRef]
- De Groef, L.; Andries, L.; Siwakoti, A.; Geeraerts, E.; Bollaerts, I.; Noterdaeme, L.; Etienne, I.; Papageorgiou, A.P.; Stalmans, I.; Billen, J.; et al. Aberrant collagen composition of the trabecular meshwork results in reduced aqueous humor drainage and elevated IOP in MMP-9 Null Mice. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5984–5995. [Google Scholar] [CrossRef]
- Adornetto, A.; Russo, R.; Parisi, V. Neuroinflammation as a target for glaucoma therapy. Neural Regen. Res. 2019, 14, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Vernazza, S.; Tirendi, S.; Bassi, A.M.; Raverso, C.E.; Saccà, S.C. Neuroinflammation in primary open-angle glaucoma. J. Clin. Med. 2020, 9, 3172. [Google Scholar] [CrossRef] [PubMed]
- Soto, I.; Howell, G.R. The complex role of neuroinflammation in glaucoma. Cold Spring Harb. Perspect. Med. 2014, 4, a017269. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.N.; Inman, D.M.; Dengler-Crish, C.M.; Smith, M.A.; Crish, S.D. Early pro-inflammatory cytokine elevations in the DBA/2J mouse model of glaucoma. J. Neuroinflamm. 2015, 12, 176. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Luo, C.; Cai, J. Neurodegenerative and inflammatory pathway components linked to TNF-alpha/TNFR1 signaling in the glaucomatous human retina. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8442–8454. [Google Scholar] [CrossRef] [PubMed]
- Yerramothu, P.; Vijay, A.K.; Willcox, M.D.P. Inflammasomes, the eye and anti-inflammasome therapy. Eye 2018, 32, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Chintala, S.K. The emerging role of proteases in retinal ganglion cell death. Exp. Eye Res. 2006, 82, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.; Li, F.; Chen, H.; Wang, Y.; Zhu, Y.; Yang, X.; Zhu, J.; Wu, F.; Ouyang, H.; Ge, J.; et al. Caspase-8 promotes NLRP1/NLRP3 inflammasome activation and IL-1β production in acute glaucoma. Proc. Natl. Acad. Sci. USA 2014, 111, 11181–11186. [Google Scholar] [CrossRef]
- Tribble, J.R.; Harder, J.M.; Williams, P.A.; John, S.W.M. Ocular hypertension suppresses homeostatic gene expression in optic nerve head microglia of DBA/2 J mice. Mol. Brain 2020, 13, 81. [Google Scholar] [CrossRef]
- Burgoyne, C.F.; Downs, J.C.; Bellezza, A.J.; Suh, J.K.; Hart, R.T. The optic nerve head as a biomechanical structure; a new paradigm for understanding the role of IOP-related stress and strain in the pathophysiology of glaucomatous optic nerve head damage. Prog. Retinal Eye Res. 2005, 24, 39–73. [Google Scholar] [CrossRef]
- Downs, J.C.; Roberts, M.D.; Sigal, I.A. Glaucomatous cupping of the lamina cribrosa: A review of the evidence for active progressive remodeling as a mechanism. Exp. Eye Res. 2011, 93, 133–140. [Google Scholar] [CrossRef]
- Daguman, I.J.; Delfin, M.S. Correlation of lamina cribosa and standard automated perimeter findings in glaucoma and non-glaucoma patients. J. Ophthal. Studies 2018, 2, 1–5. [Google Scholar]
- Coudrillier, B.; Campbell, I.C.; Read, A.T.; Geraldes, D.M.; Vo, N.T.; Feola, A.; Mulvihill, J.; Albon, J.; Abel, R.L.; Ethier, C.R. Effects of peripapillary scleral stiffening on the deformation of the lamina cribrosa. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2666–2677. [Google Scholar] [CrossRef] [PubMed]
- Jóhannesson, G.; Eklund, A.; Lindén, C. Intracranial and intraocular pressure at the lamina cribrosa: Gradient effects. Curr. Neurol. NeuroSci. Rep. 2018, 18, 25. [Google Scholar] [CrossRef] [PubMed]
- Wostyn, P.; De Groot, V.; Van Dam, D.; Audenaert, K.; Killer, H.E.; De Deyn, P.P. Glaucoma and the role of cerebrospinal fluid dynamics. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6630–6631. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Park, H.-Y.L.; Lee, K.-I.I.; Lee, K.; Shin, H.Y.; Park, C.K. Torsion of the optic nerve head is a prominent feature of normal-tension glaucoma. Investig. Ophthalmol. Vis. Sci. 2015, 2015, 156–163. [Google Scholar] [CrossRef]
- Nickells, R.W.; Howell, G.R.; Soto, I.; John, S.W. Under pressure: Cellular and molecular responses during glaucoma, a common neurodegeneration with axonopathy. Annu. Rev. Neurosci. 2012, 35, 153–179. [Google Scholar] [CrossRef]
- Evangelho, K.; Mogilevskaya, M.; Losada-Barragan, M.; Vargas-Sanchez, J.K. Pathophysiology of primary open-angle glaucoma from a neuroinflammatory and neurotoxicity perspective: A review of the literature. Int. Ophthalmol. 2019, 39, 259–271. [Google Scholar] [CrossRef]
- Fahy, E.T.; Chrysostomou, V.; Crowston, J.G. Impaired axonal transport in glaucoma. Curr. Eye Res. 2016, 41, 273–283. [Google Scholar] [CrossRef]
- Nuschke, A.C.; Farrell, S.R.; Levesque, J.M.; Chauhan, B.C. Assessment of retinal ganglion cell damage in glaucomatous optic neuropathy: Axon transport, injury and soma loss. Exp. Eye Res. 2015, 141, 111–124. [Google Scholar] [CrossRef]
- Li, Y.; Li, D.; Ying, X.; Khaw, P.T.; Raisman, G. An energy theory of glaucoma. Glia 2015, 63, 1537–1552. [Google Scholar] [CrossRef] [PubMed]
- Osborne, N.N.; Nunez-Alvarez, C.; Joglar, B.; Del Olmo-Aguado, S. Glaucoma: Focus on mitochondria in relation to pathogenesis and neuroprotection. Eur. J. Pharmacol. 2016, 787, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Eells, J.T. Mitochondrial dysfunction in the aging retina. Biology 2019, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Silverman, S.M.; Kim, B.J.; Howell, G.R.; Miller, J.; John, S.W.; Wordinger, R.J.; Clark, A.F. C1q propagates microglial activation and neurodegeneration in the visual axis following retinal ischemia/reperfusion injury. Mol. Neurodegener. 2016, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Stasi, K.; Nagel, D.; Yang, X.; Wang, R.F.; Ren, L.; Podos, S.M.; Mittag, T.; Danias, J. Complement component 1Q (C1Q) upregulation in retina of murine, primate, and human glaucomatous eyes. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1024–1029. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.; Alavi, M.V.; Kim, K.Y.; Kang, T.; Scott, R.T.; Noh, Y.H.; Lindsey, J.D.; Wissinger, B.; Ellisman, M.H.; Weinreb, R.N.; et al. A new vicious cycle involving glutamate excitotoxicity, oxidative stress and mitochondrial dynamics. Cell Death Dis. 2011, 2, e240. [Google Scholar] [CrossRef]
- McElnea, E.M.; Quill, B.; Docherty, N.G.; Irnaten, M.; Siah, W.F.; Clark, A.F.; O’Brien, C.J.; Wallace, D.M. Oxidative stress, mitochondrial dysfunction and calcium overload in human lamina cribrosa cells from glaucoma donors. Mol. Vis. 2011, 17, 1182–1189. [Google Scholar]
- Maddineni, P.; Kasetti, R.B.; Patel, P.D.; Millar, J.C.; Kiehlbauch, C.; Clark, A.F.; Zode, G.S. CNS axonal degeneration and transport deficits at the optic nerve head precede structural and functional loss of retinal ganglion cells in a mouse model of glaucoma. Mol. Neurodegener. 2020, 15, 48. [Google Scholar] [CrossRef]
- Skonieczna, K.; Grabska-Liberek, I.; Terelak-Borys, B.; Jamrozy-Witkowska, A. Selected autoantibodies and normal-tension glaucoma. Med. Sci. Monit. 2014, 20, 1201–1219. [Google Scholar]
- Sanes, J.R.; Masland, R.H. The types of retinal ganglion cells: Current status and implications for neuronal classification. Annu. Rev. Neurosci. 2015, 38, 221–246. [Google Scholar] [CrossRef]
- Guo, L.; Moss, S.E.; Alexander, R.A.; Ali, R.R.; Fitzke, F.W.; Cordeiro, M.F. Retinal ganglion cell apoptosis in glaucoma is related to intraocular pressure and IOP-induced effects on extracellular matrix. Investig. Ophthalmol. Vis. Sci. 2005, 46, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Resta, V.; Novelli, E.; Vozzi, G.; Scarpa, C.; Caleo, M.; Ahluwalia, A.; Solini, A.; Santini, E.; Parisi, V.; Di Virgilio, F.; et al. Acute retinal ganglion cell injury caused by intraocular pressure spikes is mediated by endogenous extracellular ATP. Eur. J. Neurosci. 2007, 25, 2741–2754. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Jo, R.E.; Ullian, E.M.; Wong, R.O.; Della Santina, L. Selective vulnerability of specific retinal ganglion cell types and synapses after transient ocular hypertension. J. Neurosci. 2016, 36, 9240–9252. [Google Scholar] [CrossRef] [PubMed]
- Della Santina, L.; Ou, Y. Who’s lost first? Susceptibility of retinal ganglion cell types in experimental glaucoma. Exp. Eye Res. 2017, 158, 43–50. [Google Scholar] [CrossRef]
- Chaphalkar, R.M.; Stankowska, D.L.; He, S.; Kodati, B.; Phillips, N.; Prah, J.; Yang, S.; Krishnamoorthy, R.R. Endothelin-1 mediated decrease in mitochondrial gene expression and bioenergetics contribute to neurodegeneration of retinal ganglion cells. Sci. Rep. 2020, 10, 3571. [Google Scholar] [CrossRef]
- Bhandari, A.; Smith, J.C.; Zhang, Y.; Jensen, A.A.; Reid, L.; Goeser, T.; Fan, S.; Ghate, D.; Van Hook, M.J. Early-stage ocular hypertension alters retinal ganglion cell synaptic transmission in the visual thalamus. Front. Cell. Neurosci. 2019, 13, 426. [Google Scholar] [CrossRef]
- Dvoriantchikova, G.; Ivanov, D. Tumor necrosis factor-alpha mediates activation of NF-κB and JNK signaling cascades in retinal ganglion cells and astrocytes in opposite ways. Eur. J. Neurosci. 2014, 40, 3171–3178. [Google Scholar] [CrossRef]
- Tezel, G.; Yang, X.; Luo, C.; Kain, A.D.; Powell, D.W.; Kuehn, M.H.; Kaplan, H.J. Oxidative stress and the regulation of complement activation in human glaucoma. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5071–5082. [Google Scholar] [CrossRef]
- Cooper, M.L.; Crish, S.D.; Inman, D.M.; Horner, P.J.; Calkins, D.J. Early astrocyte redistribution in the optic nerve precedes axonopathy in the DBA/2J mouse model of glaucoma. Exp. Eye Res. 2016, 150, 22–33. [Google Scholar] [CrossRef]
- Hollander, H.; Makarov, F.; Stefani, F.H.; Stone, J. Evidence of constriction of optic axons at the lamina cribrosa in the normotensive eye in humans and other mammals. Ophthalmic Res. 1995, 127, 296–309. [Google Scholar] [CrossRef]
- Kong, Y.X.; Crowston, J.G.; Vingrys, A.J.; Trounce, I.A.; Bui, V.B. Functional changes in the retina during and after acute intraocular pressure elevation in mice. Investig. Ophthalmol. Vis. Sci. 2010, 50, 5732–5740. [Google Scholar] [CrossRef] [PubMed]
- Ebneter, A.; Casson, R.J.; Wood, J.P.; Chidlow, G. Microglial activation in the visual pathway in experimental glaucoma: Spatiotemporal characterization and correlation with axonal injury. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6448–6460. [Google Scholar] [CrossRef] [PubMed]
- Harwerth, R.S.; Quigley, H.A. Visual field defects and retinal ganglion cell losses in patients with glaucoma. Arch Ophthalmol. 2006, 124, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Tu, S.; Li, K.; Ding, X.; Hu, D.; Li, K.; Ge, J. Relationship between intraocular pressure and retinal nerve fibre thickness loss in a monkey model of chronic ocular hypertension. Eye 2019, 33, 1833–1841. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Weinreb, R.N.; Leung, C.K. Optic nerve head deformation in glaucoma: The temporal relationship between optic nerve head surface depression and retinal nerve fiber layer thinning. Ophthalmology 2014, 121, 2362–2370. [Google Scholar] [CrossRef]
- Sharif, N.A. Glaucomatous optic neuropathy treatment options: The promise of novel therapeutics, techniques and tools to help preserve vision. Neural Regen. Res. 2018, 13, 1145–1150. [Google Scholar] [CrossRef]
- Medeiros, F.A.; Weinreb, R.N.; Zangwill, L.M.; Alencar, L.M.; Sample, P.A.; Vasile, C.; Bowd, C. Long-term intraocular pressure fluctuations and risk of conversion from ocular hypertension to glaucoma. Ophthalmology 2008, 115, 934–940. [Google Scholar] [CrossRef]
- Yucel, Y.H.; Zhang, Q.; Weinreb, R.N.; Kaufman, P.L.; Gupta, N. Atrophy of relay neurons in magno- and parvocellular layers in the lateral geniculate nucleus in experimental glaucoma. Investig. Ophthalmol. Vis. Sci. 2001, 42, 3216–3222. [Google Scholar]
- Gupta, N.; Ly, T.; Zhang, Q.; Kaufman, P.L.; Weinreb, R.N.; Yücel, Y.H. Chronic ocular hypertension induces dendrite pathology in the lateral geniculate nucleus of the brain. Exp. Eye Res. 2007, 84, 176–184. [Google Scholar] [CrossRef]
- Trivedi, V.; Bang, J.W.; Parra, C.; Colbert, M.K.; O’Connell, C.; Arshad, A.; Faiq, M.A.; Conner, I.P.; Redfern, M.S.; Wollstein, G.; et al. Widespread brain reorganization perturbs visuomotor coordination in early glaucoma. Sci. Rep. 2019, 9, 14168. [Google Scholar] [CrossRef]
- Van Hook, M.J.; Monaco, C.; Bierlein, E.R.; Smith, J.C. Neuronal and synaptic plasticity in the visual thalamus in mouse models of glaucoma. Front. Cell. Neurosci. 2021, 14, 626056. [Google Scholar] [CrossRef] [PubMed]
- Hvozda Arana, A.G.; Lasagni Vitar, R.M.; Reides, C.G.; Calabró, V.; Marchini, T.; Lerner, S.F.; Evelson, P.A.; Ferreira, S.M. Mitochondrial function is impaired in the primary visual cortex in an experimental glaucoma model. Arch. Biochem. Biophys. 2021, 701, 108815. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Xie, L.; Dai, C.; Liang, M.; Zhao, L.; Yin, X.; Wang, J. Progressive thinning of visual cortex in primary open-angle glaucoma of varying severity. PLoS ONE 2015, 10, e0121960. [Google Scholar] [CrossRef] [PubMed]
- Crabb, D.P. A view on glaucoma—Are we seeing it clearly? Eye 2016, 30, 304–313. [Google Scholar] [CrossRef]
- Sharif, N.A.; Odani-Kawabata, N.; Lu, F.; Pinchuk, L. FP and EP2 prostanoid receptor agonist drugs and aqueous humor outflow devices for treating ocular hypertension and glaucoma. Exp. Eye Res. 2023, 229, 109415. [Google Scholar] [CrossRef]
- Klimko, P.; Sharif, N.A. Discovery, characterization and clinical utility of prostaglandin agonists for treatment of glaucoma. Br. J. Pharmacol. 2019, 176, 1051–1058. [Google Scholar] [CrossRef]
- Hollo, G.; Topouzis, F.; Fechtner, R.D. Fixed-combination intraocular pressure-lowering therapy for glaucoma and ocular hypertension: Advantages in clinical practice. Expert Opin. Pharmacother. 2014, 15, 1737–1747. [Google Scholar] [CrossRef]
- Asrani, S.; Bacharach, J.; Holland, E.; McKee, H.; Sheng, H.; Lewis, R.A.; Kopczynski, C.C.; Heah, T. Fixed-dose combination of netarsudil and latanoprost in ocular hypertension and open-angle glaucoma: Pooled efficacy/safety analysis of phase 3 MERCURY-1 and -2. Adv. Ther. 2020, 37, 1620–1631. [Google Scholar] [CrossRef]
- Nardi, M.; Casini, G.; Guidi, G.; Figus, M. Emerging surgical therapy in the treatment of glaucoma. Prog. Brain Res. 2015, 221, 341–357. [Google Scholar]
- Ahmed, I.I.K.; Fea, A.; Au, L.; Ang, R.E.; Harasymowycz, P.; Jampel, H.D.; Samuelson, T.W.; Chang, D.F.; Rhee, D.J.; COMPARE Investigators. A prospective randomized trial comparing Hydrus and iStent microinvasive glaucoma surgery implants for standalone treatment of open-angle glaucoma: The COMPARE Study. Ophthalmology 2020, 127, 52–61. [Google Scholar] [CrossRef]
- Lee, R.M.H.; Bouremel, Y.; Eames, I.; Brocchini, S.; Khaw, P.T. Translating minimally invasive glaucoma surgery devices. Clin. Transl. Sci. 2020, 13, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Gazzard, G.; Konstantakopoulou, E.; Garway-Heath, D.; Adeleke, M.; Vickerstaff, V.; Ambler, G.; Hunter, R.; Bunce, C.; Nathwani, N.; Barton, K.; et al. Laser in glaucoma and ocular hypertension (LiGHT) trial: Six-year results of primary selective laser trabeculoplasty versus eye drops for the treatment of glaucoma and ocular hypertension. Ophthalmology 2023, 130, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Newman-Casey, P.A.; Robin, A.L.; Blachley, T.; Farris, K.; Heisler, M.; Resnicow, K.; Lee, P.P. The most common barriers to glaucoma medication adherence. Ophthalmology 2015, 122, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Alm, A.; Grierson, I.; Shields, M.B. Side effects associated with prostaglandin analog therapy. Surv. Ophthalmol. 2008, 53 (Suppl. S1), S93–S105. [Google Scholar] [CrossRef]
- Yeh, P.H.; Cheng, Y.C.; Shie, S.S.; Lee, Y.S.; Shen, S.C.; Chen, H.S.; Wu, W.C.; Su, W.W. Brimonidine related acute follicular conjunctivitis: Onset time and clinical presentations, a long-term follow-up. Medicine 2021, 100, e26724. [Google Scholar] [CrossRef]
- Sharif, N.A. Neuropathology and therapeutics addressing glaucoma, a prevalent sight threatening retina-optic nerve-brain disease. OBM Neurobiol. 2022, 6, 42. [Google Scholar] [CrossRef]
- Sharif, N.A. iDrugs and iDevices discovery and development—Preclinical assays, techniques and animal model studies for ocular hypotensives and neuroprotectants. J. Ocular. Pharmacol. Ther. 2018, 34, 7–39. [Google Scholar] [CrossRef]
- Rao, P.V.; Pattabiraman, P.P.; Kopczynski, C. Role of the Rho GTPase/Rho kinase signaling pathway in pathogenesis and treatment of glaucoma: Bench to bedside research. Exp. Eye Res. 2017, 158, 23–32. [Google Scholar] [CrossRef]
- Sharif, N.A. Rho kinase inhibitor utility for glaucoma and optic neuropathy treatment: Enzymic activity, IOP-lowering and neuroprotection perspectives. Open Access J. Ophthalmol. 2022, 7, 000233. [Google Scholar]
- Ramachandran, C.; Patil, R.V.; Sharif, N.A.; Srinivas, S.P. Effect of elevated intracellular cAMP on actomyosin contraction in bovine trabecular meshwork cells. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1474–1485. [Google Scholar] [CrossRef]
- Ramachandran, C.; Patil, R.V.; Combrink, K.; Sharif, N.A.; Srinivas, S.P. Rho-Rho kinase pathway in the actomyosin contraction and cell-matrix adhesion in immortalized human trabecular meshwork cells. Mol. Vis. 2011, 17, 1877–1890. [Google Scholar] [PubMed]
- Henderson, A.J.; Hadden, M.; Guo, C.; Douglas, N.; Decornez, H.; Hellberg, M.R.; Rusinko, A.; McLaughlin, M.; Sharif, N.; Drace, C.; et al. 2,3-Diaminopyrines as rho kinase inhibitors. Bioorganic Med. Chem. Lett. 2010, 20, 1137–1140. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-H.; Namil, A.; Severns, B.; Ward, J.; Kelly, C.R.; Drace, C.; McLaughlin, M.A.; Yacoub, S.; Li, B.; Patil, R.; et al. In vivo optimization of 2,3-diaminopyrazine Rho kinase inhibitors. Bioorganic Med. Chem. Lett. 2014, 24, 1875–1879. [Google Scholar] [CrossRef] [PubMed]
- Goldhagen, B.; Proia, A.D.; Epstein, D.L.; Rao, P.V. Elevated levels of RhoA in the optic nerve head of human eyes with glaucoma. J. Glaucoma 2012, 21, 530–538. [Google Scholar] [CrossRef]
- Futakuchi, A.; Morimoto, T.; Ikeda, Y.; Tanihara, H.; Inoue, T. Intraocular pressure-lowering effects of ripasudil in uveitic glaucoma, exfoliation glaucoma, and steroid-induced glaucoma patients: ROCK-S, a multicentre historical cohort study. Sci. Rep. 2020, 10, 10308. [Google Scholar] [CrossRef]
- Tanihara, H.; Inoue, T.; Yamamoto, T.; Kuwayama, Y.; Abe, H.; Suganami, H.; Araie, M.; K-115 Clinical Study Group. Additive intraocular pressure-lowering effects of the rho kinase inhibitor ripasudil (K-115) combined with timolol or latanoprost: A report of 2 randomized clinical trials. JAMA Ophthalmol. 2015, 133, 755–761. [Google Scholar] [CrossRef]
- Lin, C.W.; Sherman, B.; Moore, L.A.; Laethem, C.L.; Lu, D.W.; Pattabiraman, P.P.; Rao, P.V.; deLong, M.A.; Kopczynski, C.C. Discovery and preclinical development of netarsudil, a novel ocular hypotensive agent for the treatment of glaucoma. J. Ocul. Pharmacol. Ther. 2018, 34, 40–51. [Google Scholar] [CrossRef]
- Serle, J.B.; Katz, L.J.; McLaurin, E.; Heah, T.; Ramirez-Davis, N.; Usner, D.W.; Novack, G.D.; Kopczynski, C.C.; ROCKET-1 and ROCKET-2 Study Groups. Two phase 3 clinical trials comparing the safety and efficacy of netarsudil to timolol in patients with elevated intraocular pressure: Rho kinase elevated IOP treatment trial 1 and 2 (ROCKET-1 and ROCKET-2). Am. J. Ophthalmol. 2018, 186, 116–127. [Google Scholar] [CrossRef]
- Singh, I.P.; Fechtner, R.D.; Myers, J.S.; Kim, T.; Usner, D.W.; McKee, H.; Sheng, H.; Lewis, R.A.; Heah, T.; Kopczynski, C.C. Pooled efficacy and safety profile of netarsudil ophthalmic solution 0.02% in patients with open-angle glaucoma or ocular hypertension. J. Glaucoma 2020, 29, 878–884. [Google Scholar] [CrossRef]
- Rashad, R.; Zhu, C.; Kupcha, A.C.; Distefano, A.G.; Kefella, H.; Desai, M.A. Partial stenosis and complete punctal closure following topical netarsudil use for glaucoma. J. Glaucoma 2022, 31, 920–925. [Google Scholar] [CrossRef]
- Sit, A.J.; Gupta, D.; Kazemi, A.; McKee, H.; Challa, P.; Liu, K.C.; Lopez, J.; Kopczynski, C.; Heah, T. Netarsudil Improves Trabecular Outflow Facility in Patients with Primary Open Angle Glaucoma or Ocular Hypertension: A Phase 2 Study. Am. J. Ophthalmol. 2021, 226, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Walters, T.R.; Ahmed, I.I.K.; Lewis, R.A.; Usner, D.W.; Lopez, J.; Kopczynski, C.C.; Heah, T.; MERCURY-2 Study Group. Once-Daily Netarsudil/Latanoprost Fixed-Dose Combination for Elevated Intraocular Pressure in the Randomized Phase 3 MERCURY-2 Study. Ophthalmol Glaucoma. 2019, 2, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Ren, R.; Li, G.; Le, T.D.; Kopczynski, C.; Stamer, W.D.; Gong, H. Netarsudil increases outflow facility in human eyes through multiple mechanisms. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6197–6209. [Google Scholar] [CrossRef] [PubMed]
- Van de Velde, S.; De Groef, L.; Stalmans, I.; Moons, L.; Van Hove, I. Towards axonal regeneration and neuroprotection in glaucoma: Rho kinase inhibitors as promising therapeutics. Prog. Neurobiol. 2015, 131, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Sagawa, H.; Terasak, H.; Nakamura, M.; Ichikawa, M.; Ata, T.; Tokita, Y.; Watanabe, M. A novel rock inhibitor, Y-39983, promotes regeneration of crushed axons of retinal ganglion cells into the optic nerve of adult cats. Exp. Neurol. 2007, 205, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.X.; Sang, A.; Wang, Y.; Ho, D.; Douglas, C.; Dia, L.; Goldberg, J.L. Topical administration of a ROCK/NET inhibitor promotes retinal ganglion cell survival and axon regeneration after optic nerve injury. Exp. Eye Res. 2017, 158, 33–42. [Google Scholar] [CrossRef]
- Li, G.; Lee, C.; Read, A.T.; Wang, K.; Ha, J.; Kuhn, M.; Navarro, I.; Cui, J.; Young, K.; Gorijavolu, R. Anti-fibrotic activity of a rho-kinase inhibitor restores outflow function and intraocular pressure homeostasis. Elife 2021, 10, e60831. [Google Scholar] [CrossRef]
- Pattabiraman, P.P.; Rinkoski, T.; Poeschla, E.; Proia, A. RhoA GTPase-induced ocular hypertension in a rodent model is associated with increased fibrogenic activity in the trabecular meshwork. Am. J. Pathol. 2015, 185, 496–512. [Google Scholar] [CrossRef]
- Yamashita, K.; Kotani, Y.; Nakajima, Y.; Shimazawa, M.; Yoshimura, S.; Nakashima, S.; Iwama, T.; Hara, H. Fasudil, a rho kinase (ROCK) inhibitor, protects against ischemic neuronal damage in vitro and in vivo by acting directly on neurons. Brain Res. 2007, 1154, 215–224. [Google Scholar] [CrossRef]
- Tokushige, H.; Waki, M.; Takayama, Y.; Tanihara, H. Effects of Y-39983, a selective rho-associated protein kinase inhibitor, on blood flow in optic nerve head in rabbits and axonal regeneration of retinal ganglion cells in rats. Curr. Eye Res. 2011, 36, 964–970. [Google Scholar] [CrossRef]
- Bastia, E.; Toris, C.B.; Brambilla, S.; Galli, C.; Almirante, N.; Bergamini, M.V.W.; Masini, E.; Sgambellone, S.; Unser, A.M.; Ahmed, F.; et al. NCX 667, a novel nitric oxide donor, lowers intraocular pressure in rabbits, dogs, and non-human primates and enhances TGFβ2-induced outflow in HTM/HSC constructs. Investig. Ophthalmol. Vis. Sci. 2021, 62, 17. [Google Scholar] [CrossRef] [PubMed]
- Nathanson, J.A.; McKee, M. Identification of an extensive system of nitric oxide-producing cells in the ciliary muscle and outflow pathway of the human eye. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1765–1773. [Google Scholar]
- Nathanson, J.A.; McKee, M. Alterations of ocular nitric oxide synthase in human glaucoma. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1774–1784. [Google Scholar]
- Cavet, M.E.; DeCory, H.H. The role of nitric oxide in the intraocular pressure lowering efficacy of latanoprostene bunod: Review of nonclinical studies. J. Ocul. Pharmacol. Ther. 2018, 34, 52–60. [Google Scholar] [CrossRef]
- Kondkar, A.A.; Azad, T.A.; Sultan, T.; Osman, E.A.; Almobarak, F.A.; Al-Obeidan, S.A. Association of endothelial nitric oxide synthase (NOS3) gene polymorphisms with primary open-angle glaucoma in a Saudi cohort. PLoS ONE 2020, 15, e0227417. [Google Scholar] [CrossRef]
- Reina-Torres, E.; De Ieso, M.L.; Pasquale, L.R.; Madekurozwa, M.; van Batenburg-Sherwood, J.; Overby, D.R.; Stamer, W.D. The vital role for nitric oxide in intraocular pressure homeostasis. Prog. Retin. Eye Res. 2021, 83, 100922. [Google Scholar] [CrossRef] [PubMed]
- Ellis, D.; Scheibler, L.; Sharif, N.A. Prostaglandin Conjugates and Derivatives for Treating Glaucoma and Ocular Hypertension. U.S. Patent 9604949 B2, March 2017. [Google Scholar]
- Shahidullah, M.; Mandal, A.; Wei, G.; Delamere, N.A. Nitric oxide regulation of Na, K-ATPase activity in ocular ciliary epithelium involves Src family kinase. J. Cell Physiol. 2014, 229, 343–352. [Google Scholar] [CrossRef]
- Radell, J.E.; Sharma, H.K.; Auyeung, K.L.; Paul, M.E.; Gagliuso, D.J.; Chadha, N.; Tsai, J.C.; Serle, J.B. Two-Year Experience With Latanoprostene Bunod in Clinical Practice. J. Glaucoma 2021, 30, 776–780. [Google Scholar] [CrossRef]
- Xu, D.; Wu, F.; Yu, Y.; Lou, X.; Ye, M.; Zhang, H.; Zhao, Y. Sympathetic activation leads to Schlemm’s canal expansion via increasing vasoactive intestinal polypeptide secretion from trabecular meshwork. Exp. Eye Res. 2022, 224, 109235. [Google Scholar] [CrossRef]
- Roddy, G.W.; Roy Chowdhury, U.; Anderson, K.J.; Rinkoski, T.A.; Hann, C.R.; Chiodo, V.A.; Smith, W.C.; Fautsch, M.P. Transgene expression of Stanniocalcin-1 provides sustained intraocular pressure reduction by increasing outflow facility. PLoS ONE 2022, 17, e0269261. [Google Scholar] [CrossRef]
- Weinreb, R.N.; Scassellati Sforzolini, B.; Vittitow, J.; Liebmann, J. Latanoprostene Bunod 0.024% versus Timolol Maleate 0.5% in Subjects with Open-Angle Glaucoma or Ocular Hypertension: The APOLLO Study. Ophthalmology 2016, 123, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, R.N.; Liebmann, J.M.; Martin, K.R.; Kaufman, P.L.; Vittitow, J.L. Latanoprostene Bunod 0.024% in Subjects With Open-angle Glaucoma or Ocular Hypertension: Pooled Phase 3 Study Findings. J. Glaucoma 2018, 27, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Fingeret, M.; Gaddie, I.B.; Bloomenstein, M. Latanoprostene bunod ophthalmic solution 0.024%: A new treatment option for open-angle glaucoma and ocular hypertension. Clin. Exp. Optom. 2019, 102, 541–550. [Google Scholar] [CrossRef]
- Hoy, S.M. Latanoprostene Bunod Ophthalmic Solution 0.024%: A Review in Open-Angle Glaucoma and Ocular Hypertension. Drugs 2018, 78, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Addis, V.M.; Miller-Ellis, E. Latanoprostene bunod ophthalmic solution 0.024% in the treatment of open-angle glaucoma: Design, development, and place in therapy. Clin. Ophthalmol. 2018, 12, 2649–2657. [Google Scholar] [CrossRef] [PubMed]
- Sharif, N.A.; Williams, G.W.; Kelly, C.R. Bimatoprost and its free acid are prostaglandin FP receptor agonists. Eur. J. Pharmacol. 2001, 432, 211–213. [Google Scholar] [CrossRef]
- Sharif, N.A.; Kelly, C.R.; Crider, J.Y. Human trabecular meshwork cell responses induced by bimatoprost, travoprost, unoprostone, and other FP prostaglandin receptor agonist analogues. Investig. Ophthalmol. Vis. Sci. 2003, 44, 715–721. [Google Scholar] [CrossRef]
- Kelly, C.R.; Williams, G.W.; Sharif, N.A. Real-time intracellular Ca2+ mobilization by travoprost acid, bimatoprost, unoprostone, and other analogs via endogenous mouse, rat, and cloned human FP prostaglandin receptors. J. Pharmacol. Exp. Ther. 2003, 304, 238–245. [Google Scholar] [CrossRef]
- Camras, C.B.; Sharif, N.A.; Wax, M.B.; Stjernschantz, J. Bimatoprost, the prodrug of a prostaglandin analogue. Br. J. Ophthalmol. 2008, 92, 862–863. [Google Scholar]
- Lee, S.S.; Dibas, M.; Almazan, A.; Robinson, M.R. Dose-response of intracameral bimatoprost sustained-release implant and topical bimatoprost in lowering intraocular pressure. J. Ocul. Pharmacol. Ther. 2019, 35, 138–144. [Google Scholar] [CrossRef]
- Lee, S.S.; Burke, J.; Shen, J.; Almazan, A.; Orilla, W.; Hughes, P.; Zhang, J.; Li, H.; Struble, C.; Miller, P.E.; et al. Bimatoprost sustained-release intracameral implant reduces episcleral venous pressure in dogs. Vet. Ophthalmol. 2018, 21, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Brandt, J.D.; Sall, K.; DuBiner, H.; Benza, R.; Alster, Y.; Walker, G.; Semba, C.P.; Collaborators. Six-month intraocular pressure reduction with a topical bimatoprost ocular insert: Results of a phase II randomized controlled study. Ophthalmology 2016, 123, 1685–1694. [Google Scholar] [CrossRef] [PubMed]
- Bacharach, J.; Tatham, A.; Ferguson, G.; Belalcázar, S.; Thieme, H.; Goodkin, M.L.; Chen, M.Y.; Guo, Q.; Liu, J.; Robinson, M.R.; et al. Phase 3, randomized, 20-month study of the efficacy and safety of bimatoprost implant in patients with open-angle glaucoma and ocular hypertension (ARTEMIS 2). Drugs 2021, 81, 2017–2033. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, F.A.; Sheybani, A.; Shah, M.M.; Rivas, M.; Bai, Z.; Werts, E.; Ahmed, I.I.K.; Craven, E.R. Single administration of intracameral bimatoprost implant 10 µg in patients with open-angle glaucoma or ocular hypertension. Ophthalmol. Ther. 2022, 11, 1517–1537. [Google Scholar] [CrossRef] [PubMed]
- Shirley, M. Bimatoprost implant: First approval. Drugs Aging 2020, 37, 457–462. [Google Scholar] [CrossRef]
- Hellberg, M.R.; McLaughlin, M.A.; Sharif, N.A.; DeSantis, L.; Dean, T.R.; Kyba, E.P.; Bishop, J.E.; Klimko, P.G.; Zinke, P.W.; Selliah, R.D.; et al. Identification and characterization of the ocular hypotensive efficacy of travoprost, a potent and selective FP prostaglandin receptor agonist, and AL-6598, a DP prostaglandin receptor agonist. Surv. Ophthalmol. 2002, 47, S13–S33. [Google Scholar] [CrossRef]
- Sharif, N.A.; Williams, G.W.; Crider, J.Y.; Xu, S.X.; Davis, T.L. Molecular pharmacology of the DP/EP2 class prostaglandin AL-6598 and quantitative autoradiographic visualization of DP and EP2 receptor sites in human eyes. J. Ocul. Pharmacol. Ther. 2004, 20, 489–508. [Google Scholar] [CrossRef]
- Nilsson, S.F.; Drecoll, E.; Lutjen-Drecoll, E.; Toris, C.B.; Krauss, A.H.; Kharlamb, A.; Nieves, A.; Guerra, T.; Woodward, D.F. The prostanoid EP2 receptor agonist butaprost increases uveoscleral outflow in the cynomolgus monkey. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4042–4049. [Google Scholar] [CrossRef]
- Prasanna, G.; Carreiro, S.; Anderson, S.; Gukasyan, H.; Sartnurak, S.; Younis, H.; Gale, D.; Xiang, C.; Wells, P.; Dinh, D.; et al. Effect of PF-04217329 a prodrug of a selective prostaglandin EP(2) agonist on intraocular pressure in preclinical models of glaucoma. Exp. Eye Res. 2011, 93, 256–264. [Google Scholar] [CrossRef]
- Schachar, R.A.; Raber, S.; Courtney, R.; Zhang, M. A phase 2, randomized, dose-response trial of taprenepag isopropyl (PF-04217329) versus latanoprost 0.005% in open-angle glaucoma and ocular hypertension. Curr. Eye Res. 2011, 36, 809–817. [Google Scholar] [CrossRef]
- Aguirre, S.A.; Huang, W.; Prasanna, G.; Jessen, B. Corneal neovascularization and ocular irritancy responses in dogs following topical ocular administration of an EP4-prostaglandin E2 agonist. Toxicol. Pathol. 2009, 37, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Krauss, A.H.; Woodward, D.F.; Chen, J.; Gibson, L.L.; Lai, R.K.; Protzman, C.E.; Shan, T.; Williams, L.S.; Gac, T.S.; Burk, R.M. AGN 191976: A novel thromboxane A2-mimetic with ocular hypotensive properties. J. Ocul. Pharmacol. Ther. 1995, 11, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Yamane, S.; Karakawa, T.; Nakayama, S.; Nagai, K.; Moriyuki, K.; Neki, S.; Suto, F.; Kambe, T.; Hirota, Y.; Kawabata, K. IOP-lowering effect of ONO-9054, a novel dual agonist of prostanoid EP3 and FP receptors, in monkeys. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2547–2552. [Google Scholar] [CrossRef] [PubMed]
- Iwamura, R.; Tanaka, M.; Okanari, E.; Kirihara, T.; Odani-Kawabata, N.; Shams, N.; Yoneda, K. Identification of a Selective, Non-Prostanoid EP2 Receptor Agonist for the Treatment of Glaucoma: Omidenepag and its Prodrug Omidenepag Isopropyl. J. Med. Chem. 2018, 61, 6869–6891. [Google Scholar] [CrossRef] [PubMed]
- Kirihara, T.; Taniguchi, T.; Yamamura, K.; Iwamura, R.; Yoneda, K.; Odani-Kawabata, N.; Shimazaki, A.; Matsugi, T.; Shams, N.; Zhang, J.Z. Pharmacologic Characterization of Omidenepag Isopropyl, a Novel Selective EP2 Receptor Agonist, as an Ocular Hypotensive Agent. Investig. Ophthalmol. Vis. Sci. 2018, 59, 145–153. [Google Scholar] [CrossRef]
- Fuwa, M.; Toris, C.B.; Fan, S.; Taniguchi, T.; Ichikawa, M.; Odani-Kawabata, N.; Iwamura, R.; Yoneda, K.; Matsugi, T.; Shams, N.K.; et al. Effects of a novel selective EP2 receptor agonist, omidenepag isopropyl, on aqueous humor dynamics in laser-induced ocular hypertensive monkeys. J. Ocul. Pharmacol. Ther. 2018, 34, 531–537. [Google Scholar] [CrossRef]
- Crider, J.Y.; Griffin, B.W.; Sharif, N.A. Prostaglandin-stimulated adenylyl cyclase activity via a pharmacologically-defined EP2 receptor in human NPE cells. J. Ocular. Pharmacol. Ther. 1998, 14, 293–304. [Google Scholar] [CrossRef]
- Crider, J.Y.; Sharif, N.A. Functional pharmacological evidence for EP2 and EP4 prostanoid receptors in immortalized human trabecular meshwork and non-pigmented ciliary epithelial cells. J. Ocul. Pharmacol. Ther. 2001, 17, 35–46. [Google Scholar] [CrossRef]
- Schlotzer-Schrehardt, U.; Zenkel, M.; Nusing, R.M. Expression and localization of FP and EP prostanoid receptor subtypes in human ocular tissues. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1475–1487. [Google Scholar]
- Rosch, S.R.; Ramer, R.; Brune, K.; Hinz, B. Prostaglandin E2 induces cyclooxygenase-2 expression in human non-pigmented ciliary epithelial cells through activation of p38 and p42/44 mitogen-activated protein kinases. Biochem. Biophys Res. Commun. 2005, 338, 1171–1178. [Google Scholar] [CrossRef]
- Yousufzai, S.Y.; Ye, Z.; Abdel-Latif, A.A. Prostaglandin F2 alpha and its analogs induce release of endogenous prostaglandins in iris and ciliary muscles isolated from cat and other mammalian species. Exp. Eye Res. 1996, 63, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Bergh, K.; Wentzel, P.; Stjernschantz, J. Production of prostaglandin E(2) by iridial melanocytes exposed to latanoprost acid, a prostaglandin F(2 alpha) analogue. J. Ocul. Pharmacol. Ther. 2002, 18, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Uchida, T.; Shimizu, S.; Yamagishi, R.; Tokuoka, S.M.; Kita, Y.; Honjo, M.; Aihara, M. Mechanical stretch induces Ca(2+) influx and extracellular release of PGE2 through Piezo1 activation in trabecular meshwork cells. Sci. Rep. 2021, 2021, 4044. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Woodward, D.F. Prostanoid-induced relaxation of precontracted cat ciliary muscle is mediated by EP2 and DP receptors. Investig. Ophthalmol. Vis. Sci. 1992, 33, 3195–3201. [Google Scholar]
- Goh, Y.; Hotehama, Y.; Mishima, H.K. Characterization of ciliary muscle relaxation induced by various agents in cats. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1188–1192. [Google Scholar]
- Krauss, A.H.; Wiederholt, M.; Sturm, A.; Woodward, D.F. Prostaglandin effects on the contractility of bovine trabecular meshwork and ciliary muscle. Exp. Eye Res. 1997, 64, 447–453. [Google Scholar] [CrossRef]
- Wiederholt, M.; Sturm, A.; Lepple-Wienhues, A. Relaxation of trabecular meshwork and ciliary muscle by release of nitric oxide. Investig. Ophthalmol. Vis. Sci. 1994, 35, 2515–2520. [Google Scholar]
- Anthony, T.L.; Lindsey, J.D.; Aihara, M.; Weinreb, R.N. Detection of prostaglandin EP(1), EP(2), and FP receptor subtypes in human sclera. Investig. Ophthalmol. Vis. Sci. 2001, 42, 3182–3186. [Google Scholar]
- Nakamura, N.; Honjo, M.; Yamagishi, R.; Igarashi, N.; Sakata, R.; Aihara, M. Effects of selective EP2 receptor agonist, omidenepag, on trabecular meshwork cells, Schlemm’s canal endothelial cells and ciliary muscle contraction. Sci. Rep. 2021, 11, 16257. [Google Scholar] [CrossRef]
- Kumon, M.; Fuwa, M.; Shimazaki, A.; Odani-Kawabata, N.; Iwamura, R.; Yoneda, K.; Kato, M. Downregulation of COL12A1 and COL13A1 by a selective EP2 receptor agonist, omidenepag, in human trabecular meshwork cells. PLoS ONE 2023, 18, e0280331. [Google Scholar] [CrossRef]
- Kalouche, G.; Boucher, C.; Coste, A.; Debussche, L.; Orsini, C.; Baudouin, C.; Debeir, T.; Vigé, X.; Rostène, W. Prostaglandin EP2 receptor signaling protects human trabecular meshwork cells from apoptosis induced by ER stress through down-regulation of p53. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2016, 1863, 2322–2332. [Google Scholar] [CrossRef] [PubMed]
- Kalouche, G.; Beguier, F.; Bakria, M.; Melik-Parsadaniantz, S.; Leriche, C.; Debeir, T.; Rostène, W.; Baudouin, C.; Vigé, X. Activation of Prostaglandin FP and EP2 Receptors Differently Modulates Myofibroblast Transition in a Model of Adult Primary Human Trabecular Meshwork Cells. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1816–1825. [Google Scholar] [CrossRef][Green Version]
- Aihara, M.; Lu, F.; Kawata, H.; Tanaka, Y.; Yamamura, K.; Odani-Kawabata, N.; Shams, N.K. Pharmacokinetics, safety, and intraocular pressure-lowering profile of omidenepag isopropyl, a selective, non-prostaglandin, prostanoid EP2 receptor agonist, in healthy Japanese and Caucasian volunteers (Phase I Study). J. Ocul. Pharmacol. Ther. 2019, 35, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Aihara, M.; Lu, F.; Kawata, H.; Iwata, A.; Liu, K.; Odani-Kawabata, N.; Shams, N.K. Phase 2, randomized, dose-finding studies of omidenepag isopropyl, a selective EP2 agonist, in patients with primary open-angle glaucoma or ocular hypertension. J. Glaucoma 2019, 28, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Aihara, M.; Lu, F.; Kawata, H.; Iwata, A.; Odani-Kawabata, N. Twelve-month efficacy and safety of omidenepag isopropyl, a selective EP2 agonist, in open-angle glaucoma and ocular hypertension: The RENGE study. Jpn. J. Ophthalmol. 2021, 65, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Aihara, M.; Lu, F.; Kawata, H.; Iwata, A.; Odani-Kawabata, N.; Shams, N.K. Omidenepag isopropyl versus latanoprost in primary open-angle glaucoma and ocular hypertension: The phase 3 AYAME study. Am. J. Ophthalmol. 2020, 220, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Aihara, M.; Ropo, A.; Lu, F.; Kawata, H.; Iwata, A.; Odani-Kawabata, N.; Shams, N.K. Intraocular pressure-lowering effect of omidenepag isopropyl in latanoprost non-/low-responder patients with primary open-angle glaucoma or ocular hypertension: The FUJI study. Jpn. J. Ophthalmol. 2020, 64, 398–406. [Google Scholar] [CrossRef]
- McLaurin, E.B.; Tepedino, M.E. Omidenepag is Opropyl 0.002% Significantly Lowers Iop in Latanoprost Low/Non-Responders with POAG or OHT: Phase 3 SPECTRUM 5 Study. In Proceedings of the World Ophthalmology Congress, Virtual, September 2022. [Google Scholar]
- Matsuo, M.; Matsuoka, Y.; Tanito, M. Efficacy and patient tolerability of omidenepag isopropyl in the treatment of glaucoma and ocular hypertension. Clin. Ophthalmol. 2022, 16, 1261–1279. [Google Scholar] [CrossRef]
- Miki, A.; Miyamoto, E.; Ishida, N.; Shii, D.; Hori, K.; LESPOIR Research Group. Efficacy and Safety of Omidenepag Isopropyl 0.002% Ophthalmic Solution: A Retrospective Analysis of Real-World Data in Japan. Adv. Ther. 2022, 39, 2085–2095. [Google Scholar] [CrossRef]
- Nakazawa, T.; Takahashi, K.; Kuwayama, Y.; Nomura, A.; Shimada, F. Interim Results of Post-Marketing Observational Study of Omidenepag Isopropyl for Glaucoma and Ocular Hypertension in Japan. Adv. Ther. 2022, 39, 1359–1374. [Google Scholar] [CrossRef]
- Fuwa, M.; Shimazaki, A.; Odani-Kawabata, N.; Kirihara, T.; Taniguchi, T.; Iwamura, R.; Yoneda, K.; Kato, M.; Morishima, K.; Shams, N.K. Additive intraocular pressure-lowering effects of a novel selective EP2 receptor agonist, omidenepag isopropyl, combined with existing antiglaucoma agents in conscious ocular normotensive monkeys. J. Ocul. Pharmacol. Ther. 2021, 37, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Wistrand, P.J.; Stjernschantz, J.; Olsson, K. The incidence and time-course of latanoprost-induced iridial pigmentation as a function of eye color. Surv. Ophthalmol. 1997, 41 (Suppl. S2), S129–S138. [Google Scholar] [CrossRef] [PubMed]
- Kucukevcilioglu, M.; Bayer, A.; Uysal, Y.; Altinsoy, H.I. Prostaglandin associated periorbitopathy in patients using bimatoprost, latanoprost and travoprost. Clin. Exp. Ophthalmol. 2014, 42, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Terao, E.; Nakakura, S.; Nagata, Y.; Dote, S.; Tabuchi, H.; Kiuchi, Y. Evaluation of patterns and correlations of the degree of conjunctival hyperemia induced by omidenepag isopropyl 0.002% and ripasudil 0.4%. Cureus 2020, 12, e10368. [Google Scholar] [CrossRef]
- Esaki, Y.; Katsuta, O.; Kamio, H.; Noto, T.; Mano, H.; Iwamura, R.; Yoneda, K.; Odani-Kawabata, N.; Morishima, K.; Shams, N.K. The antiglaucoma agent and EP2 receptor agonist omidenepag does not affect eyelash growth in mice. J. Ocul. Pharmacol. Ther. 2020, 36, 529–533. [Google Scholar] [CrossRef]
- Inoue, K.; Shiokawa, M.; Katakura, S.; Tsuruoka, M.; Kunimatsu-Sanuki, S.; Shimizu, K.; Ishida, K.; Tomita, G. Periocular adverse reactions to omidenepag isopropyl. Am. J. Ophthalmol. 2022, 237, 114–121. [Google Scholar] [CrossRef]
- Liu, P.; Wang, F.; Song, Y.; Wang, M.; Zhang, X. Current situation and progress of drugs for reducing intraocular pressure. Ther. Adv. Chronic. Dis. 2022, 13, 20406223221140392. [Google Scholar] [CrossRef]
- Wang, T.; Cao, L.; Jiang, Q.; Zhang, T. Topical medication therapy for glaucoma and ocular hypertension. Front. Pharmacol. 2021, 12, 749858. [Google Scholar] [CrossRef]
- Donegan, R.K.; Lieberman, R.L. Discovery of molecular therapeutics for glaucoma: Challenges, successes, and promising directions. J. Med. Chem. 2016, 59, 788–809. [Google Scholar] [CrossRef]
- Bouhenni, R.A.; Dunmire, J.; Sewell, A.; Edward, D.P. Animal models of glaucoma. J. Biomed. Biotechnol. 2012, 2012, 692609. [Google Scholar] [CrossRef]
- Harada, C.; Kimura, A.; Guo, X.; Namekata, K.; Harada, T. Recent advances in genetically modified animal models of glaucoma and their roles in drug repositioning. Br. J. Ophthalmol. 2019, 103, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Struebing, F.L.; Geisert, E.E. What animal models can tell us about glaucoma. Prog. Mol. Biol. Transl. Sci. 2015, 134, 365–380. [Google Scholar] [PubMed]
- Sharif, N.A. Discovery to launch of anti-allergy (Emadine; Patanol/Pataday/Pazeo) and anti-glaucoma (Travatan; Simbrinza) ocular drugs, and generation of novel pharmacological tools such as AL-8810. ACS Pharmacol. Transl. Sci. 2020, 3, 1391–1421. [Google Scholar] [CrossRef] [PubMed]
- Woodward, D.F.; Wang, J.W.; Stamer, W.D.; Lütjen-Drecoll, E.; Krauss, A.H.; Toris, C.B. Antiglaucoma EP2 agonists: A long road that led somewhere. J. Ocul. Pharmacol. Ther. 2019, 35, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M.; Maria, D.N.; Mishra, S.R.; Guragain, D.; Wang, X.; Jablonski, M.M. Once daily pregabalin eye drops for management of glaucoma. ACS Nano 2019, 13, 13728–13744. [Google Scholar] [CrossRef] [PubMed]
- Honjo, M.; Igarashi, N.; Kurano, M.; Yatomi, Y.; Igarashi, K.; Kano, K.; Aoki, J.; Weinreb, R.N.; Aihara, M. Autotaxin–lysophosphatidic acid pathway in intraocular pressure regulation and glaucoma subtypes. Investig. Ophthalmol. Vis. Sci. 2018, 59, 693–701. [Google Scholar] [CrossRef]
- Martínez, T.; González, M.V.; Roehl, I.; Wright, N.; Pañeda, C.; Jiménez, A.I. In vitro and in vivo efficacy of SYL040012, a novel siRNA compound for treatment of glaucoma. Mol. Ther. 2014, 22, 81–91. [Google Scholar] [CrossRef]
- Pfeiffer, N.; Voykov, B.; Renieri, G.; Bell, K.; Richter, P.; Weigel, M.; Thieme, H.; Wilhelm, B.; Lorenz, K.; Feindor, M.; et al. First-in-human phase I study of ISTH0036, an antisense oligonucleotide selectively targeting transforming growth factor beta 2 (TGF-β2), in subjects with open-angle glaucoma undergoing glaucoma filtration surgery. PLoS ONE 2017, 12, e0188899. [Google Scholar] [CrossRef]
- Sun, D.; Zhan, Z.; Zeng, R.; Liu, X.; Wang, B.; Yang, F.; Huang, S.; Li, Y.; Yang, Z.; Su, Y.; et al. Long-term and potent IOP-lowering effect of IκBα-siRNA in a nonhuman primate model of chronic ocular hypertension. iScience 2022, 25, 104149. [Google Scholar] [CrossRef]
- O’Callaghan, J.; Crosbie, D.E.; Cassidy, P.S.; Sherwood, J.M.; Flügel-Koch, C.; Lütjen-Drecoll, E.; Humphries, M.M.; Reina-Torres, E.; Wallace, D.; Kiang, A.S.; et al. Therapeutic potential of AAV-mediated MMP-3 secretion from corneal endothelium in treating glaucoma. Hum. Mol. Genet. 2017, 26, 1230–1246. [Google Scholar] [CrossRef]
- Jain, A.; Zode, G.; Kasetti, R.B.; Ran, F.A.; Yan, W.; Sharma, T.P.; Bugge, K.; Searby, C.C.; Fingert, J.H.; Zhang, F.; et al. CRISPR-Cas9-based treatment of myocilin associated glaucoma. Proc. Natl. Acad. Sci. USA 2017, 114, 11199–11204. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Bell, O.H.; Copland, D.A.; Young, A.; Pooley, J.R.; Maswood, R.; Evans, R.S.; Khaw, P.T.; Ali, R.R.; Dick, A.D.; et al. Gene therapy for glaucoma by ciliary body aquaporin 1 disruption using CRISPR-Cas. Mol. Ther. 2020, 28, 820–829. [Google Scholar] [CrossRef] [PubMed]
- Levin, L.A.; Patrick, C.; Choudry, N.B.; Sharif, N.A.; Goldberg, J.L. Neuroprotection in neurodegenerations of the brain and eye: Lessons from the past and directions for the future. Front. Neurol. 2022, 13, 964197. [Google Scholar] [CrossRef] [PubMed]
- Howell, G.R.; MacNicoll, K.H.; Braine, C.E.; Soto, I.; Macalinao, D.G.; Sousa, G.L.; John, S.W. Combinatorial targeting of early pathways profoundly inhibits neurodegeneration in a mouse model of glaucoma. Neurobiol. Dis. 2014, 71, 44–52. [Google Scholar] [CrossRef]
- He, S.; Stankowska, D.L.; Ellis, D.Z.; Krishnamoorthy, R.R.; Yorio, T. Targets of neuroprotection in glaucoma. J. Ocul. Pharmacol. Ther. 2018, 34, 85–106. [Google Scholar] [CrossRef]
- Boia, R.; Ruzafa, N.; Aires, I.D.; Pereiro, X.; Ambrósio, A.F.; Vecino, E.; Santiago, A.R. Neuroprotective strategies for retinal ganglion cell degeneration: Current status and challenges ahead. Int. J. Mol. Sci. 2020, 21, 2262. [Google Scholar] [CrossRef]
- Williams, P.A.; Harder, J.M.; Cochran, K.E.; Philip, V.M.; Porciatti, V.; Smithies, O.; John, S.W.M. Vitamin B3 modulates mitochondrial vulnerability and prevents glaucoma in aged mice. Science 2017, 355, 756–760. [Google Scholar] [CrossRef]
- Komáromy, A.M.; Koehl, K.L.; Park, S.A. Looking into the future: Gene and cell therapies for glaucoma. Vet. Ophthalmol. 2021, 24 (Suppl. S1), 16–33. [Google Scholar] [CrossRef]
- Zhu, W.; Jain, A.; Gramlich, O.W.; Tucker, B.A.; Sheffield, V.C.; Kuehn, M.H. Restoration of aqueous humor outflow following transplantation of iPSC derived trabecular meshwork cells in a transgenic mouse model of glaucoma. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2054–2062. [Google Scholar] [CrossRef]
- Coulon, S.J.; Schuman, J.S.; Du, Y.; Bahrani Fard, M.R.; Ethier, C.R.; Stamer, W.D. A novel glaucoma approach: Stem cell regeneration of the trabecular meshwork. Prog. Retin. Eye Res. 2022, 90, 101063. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, S.; Jin, Z.B.; Wang, N. Stem cell-based regeneration and restoration for retinal ganglion cell: Recent advancements and current challenges. Biomolecules 2021, 11, 987. [Google Scholar] [CrossRef] [PubMed]
- Sharif, N.A. Electrical, electromagnetic, ultrasound wave therapies and electronic implants for neuronal rejuvenation, neuroprotection, axonal regeneration and IOP reduction. J. Ocul. Pharmacol. Ther. 2023, in press. [CrossRef] [PubMed]
- Batabyal, S.; Kim, S.; Wright, W.; Mohanty, S. Layer-specific nanophotonic delivery of therapeutic opsin-encoding genes into retina. Exp. Eye Res. 2021, 205, 108444. [Google Scholar] [CrossRef] [PubMed]
- Wood, E.H.; Kreymerman, A.; Kowal, T.; Buickians, D.; Sun, Y.; Muscat, S.; Mercola, M.; Moshfeghi, D.M.; Goldberg, J.L. Cellular and subcellular optogenetic approaches towards neuroprotection and vision restoration. Prog. Retin. Eye Res. 2022, 2022, 101153. [Google Scholar] [CrossRef] [PubMed]
- Verta, R.; Saccu, G.; Tanzi, A.; Grange, C.; Buono, L.; Fagoonee, S.; Deregibus, M.C.; Camussi, G.; Scalabrin, S.; Nuzzi, R.; et al. Phenotypic and functional characterization of aqueous humor derived extracellular vesicles. Exp. Eye Res. 2023, 228, 109393. [Google Scholar] [CrossRef]
- Williams, P.A.; Harder, J.M.; Foxworth, N.E.; Cardozo, B.H.; Cochran, K.E.; John, S.W.M. Nicotinamide and WLDS Act Together to Prevent Neurodegeneration in Glaucoma. Front Neurosci. 2017, 11, 232. [Google Scholar] [CrossRef]
- Hui, F.; Tang, J.; Williams, P.A.; Hadoux, X.; Casson, R.J.; Coote, M.; Trounce, I.A.; Martin, K.R.; van Wijngaarden, P.; Crowston, J.G. Improvement in inner retinal function in glaucoma with nicotinamide (vitamin B3) supplementation: A crossover randomized clinical trial. Clin. Exp. Ophthalmol. 2020, 48, 903–914. [Google Scholar] [CrossRef]
- Gaboriau, T.; Dubois, R.; Foucque, B.; Malet, F.; Schweitzer, C. 24-Hour monitoring of intraocular pressure fluctuations using a contact lens sensor: Diagnostic performance for glaucoma progression. Investig. Ophthalmol. Vis. Sci. 2023, 64, 3. [Google Scholar] [CrossRef]
- Cordeiro, M.F.; Hill, D.; Patel, R.; Corazza, P.; Maddison, J.; Younis, S. Detecting retinal cell stress and apoptosis with DARC: Progression from lab to clinic. Prog. Retin. Eye Res. 2022, 86, 100976. [Google Scholar] [CrossRef]
- Wong, D.; Chua, J.; Lin, E.; Tan, B.; Yao, X.; Chong, R.; Sng, C.; Lau, A.; Husain, R.; Aung, T.; et al. Focal structure-function relationships in primary open-angle glaucoma using OCT and OCT-A measurements. Investig. Ophthalmol. Vis. Sci. 2020, 61, 33. [Google Scholar] [CrossRef]
- Zheng, C.; Johnson, T.V.; Garg, A.; Boland, M.V. Artificial intelligence in glaucoma. Curr. Opin. Ophthalmol. 2019, 30, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Kasi, A.; Faiq, M.A.; Chan, K.C. In vivo imaging of structural, metabolic and functional brain changes in glaucoma. Neural Regen. Res. 2019, 14, 446–449. [Google Scholar] [PubMed]
- Torres, L.A.; Hatanaka, M. Correlating structural and functional damage in glaucoma. J. Glaucoma 2019, 28, 1079–1085. [Google Scholar] [CrossRef] [PubMed]






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eyedrop Medication and Year of Health Agency Approval | Common Name and Pharmacological Type of the Drug. (IOP Reduction Achieved in OHT/OAG Patients) | Topical Ocular Dosage Concentration and Form (%, w/v) | Topical Ocular Dosing Frequency | Mode(s) of Action to Reduce IOP | Common Adverse Effects |
|---|---|---|---|---|---|
Glanatec (2014 Japan) | Ripasudil (Rho kinase (ROCK) inhibitor) (3.5–4.5 mmHg IOP reduction; 16–20% reduction) | 0.4% solution | 1 drop twice daily | Enhances conventional (TM) outflow of AQH | Conjunctival congestion, conjunctival inflammation, blepharitis and eye irritation. |
Rhopressa (2017) | Netarsudil (Rho kinase (ROCK) inhibitor) (5 mmHg IOP reduction; 20–25% IOP decrease) | 0.02% solution | 1 drop daily at night | Enhances conventional (TM) outflow of AQH; also decreases episcleral veinous pressure | Conjunctival hyperemia, corneal verticillata, blurred vision, instillation site pain, and conjunctival hemorrhage. |
Vyzulta (2017) | Latanoprostene Bunod (conjugate of latanoprost and an NO-donor agent); (32–34% IOP reduction) | 0.024% solution | 1 drop at night | Enhances AQH outflow via the UVS pathway and via TM/SC pathway | Eye discomfort/irritation, hyperemia, temporary blurred vision, increase in eyelash number/length/thickness and darkening of the eyelashes /eyelids and iris. |
Durysta Implant (2020) | Intracamerally injected sustained-delivery biodegradable polymer containing bimatoprost (7.5 mmHg IOP reduction) | Not applicable | Once implanted into the ANC of the eye (intracameral injection), the drug elutes off the implant over 6 months. | Enhances AQH outflow via the UVS pathway and via TM/SC pathway | Conjunctival hyperemia, foreign body sensation, eye pain, photophobia, conjunctival hemorrhage dry eye, eye irritation increased IOP, corneal endothelial cell loss, vision blurred, iritis, headache. |
| Eybelis (2018 Japan) Omlonti (2022 USA)  | Omidenepag Isopropyl (Non-prostaglandin EP2-receptor selective agonist); (28.6–29.2% IOP reduction) | 0.002% solution | 1 drop daily at night | Enhances AQH outflow via the UVS pathway and via TM/SC pathway | Transient conjunctival hyperemia, corneal thickening. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharif, N.A. Recently Approved Drugs for Lowering and Controlling Intraocular Pressure to Reduce Vision Loss in Ocular Hypertensive and Glaucoma Patients. Pharmaceuticals 2023, 16, 791. https://doi.org/10.3390/ph16060791
Sharif NA. Recently Approved Drugs for Lowering and Controlling Intraocular Pressure to Reduce Vision Loss in Ocular Hypertensive and Glaucoma Patients. Pharmaceuticals. 2023; 16(6):791. https://doi.org/10.3390/ph16060791
Chicago/Turabian StyleSharif, Najam A. 2023. "Recently Approved Drugs for Lowering and Controlling Intraocular Pressure to Reduce Vision Loss in Ocular Hypertensive and Glaucoma Patients" Pharmaceuticals 16, no. 6: 791. https://doi.org/10.3390/ph16060791
APA StyleSharif, N. A. (2023). Recently Approved Drugs for Lowering and Controlling Intraocular Pressure to Reduce Vision Loss in Ocular Hypertensive and Glaucoma Patients. Pharmaceuticals, 16(6), 791. https://doi.org/10.3390/ph16060791