
Lipidomics Profiling of Metformin-Induced Changes in Obesity and Type 2 Diabetes Mellitus: Insights and Biomarker Potential

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Demographics and the Clinical Characteristics of this Study Population
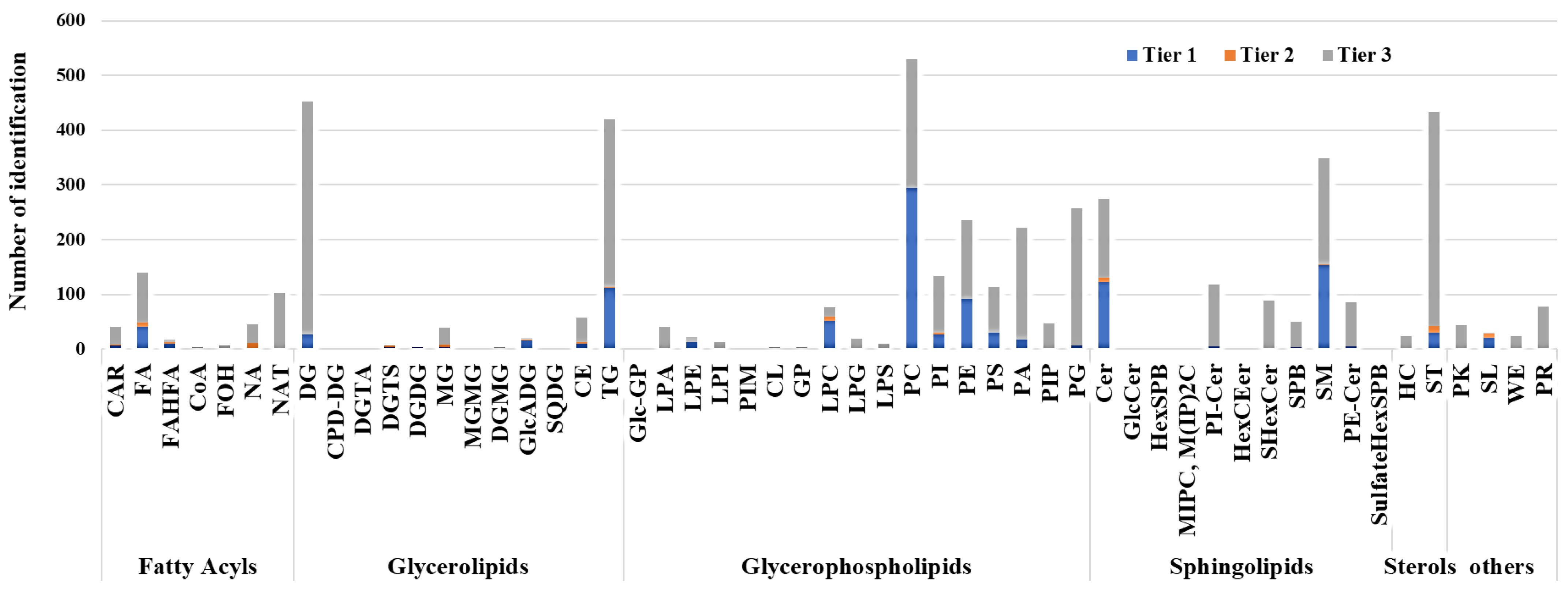
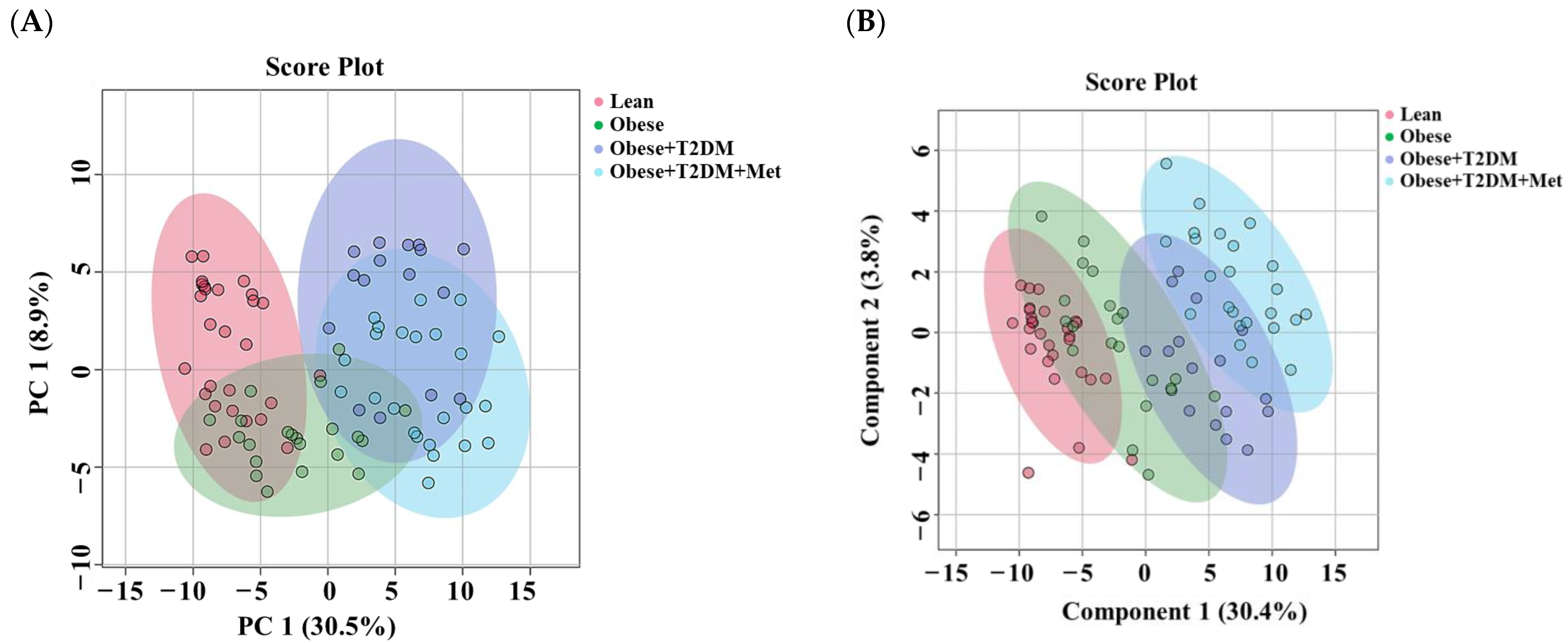
2.2. Lipid Detection and Data Overview Using Multivariate Analysis
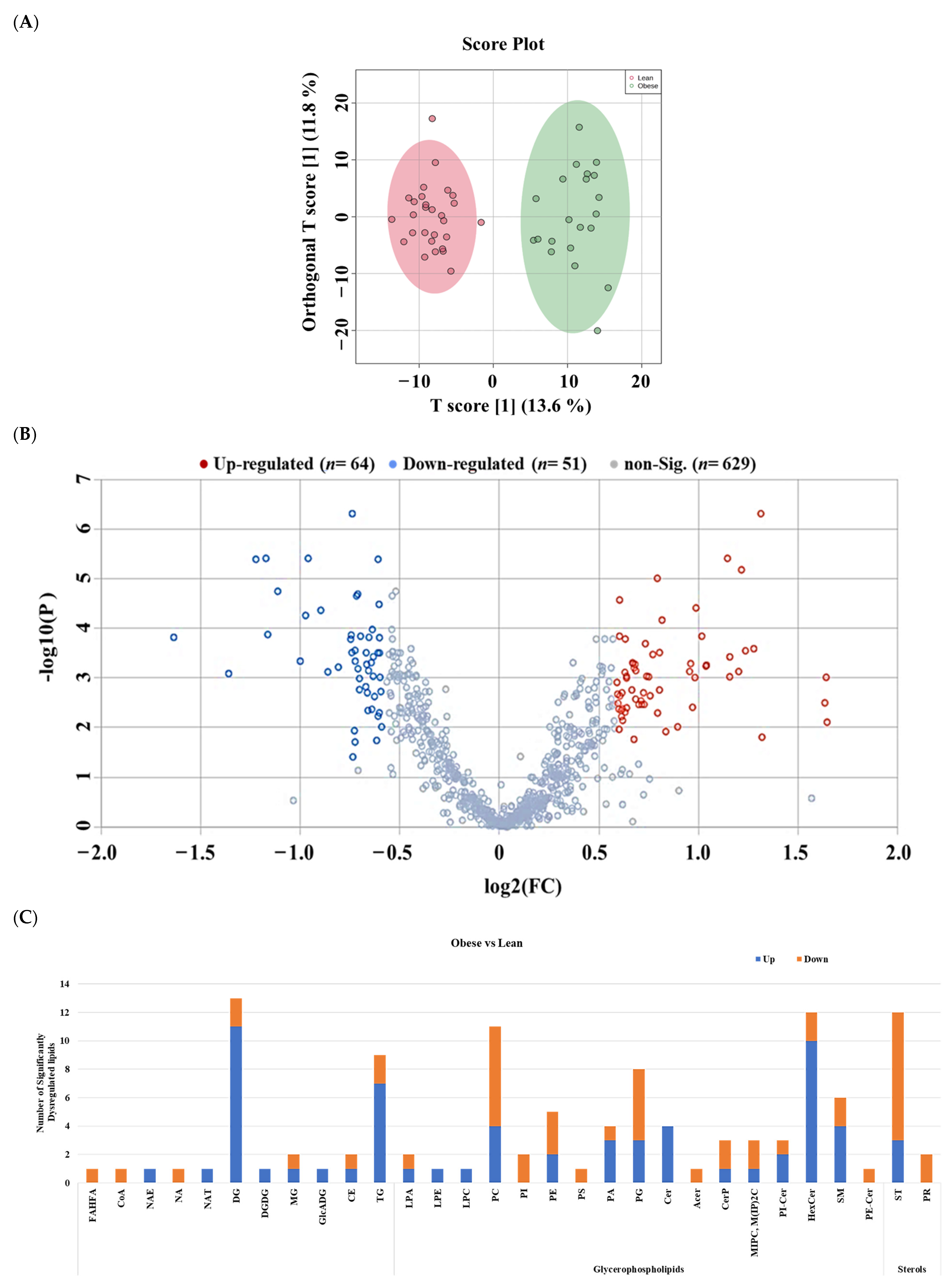
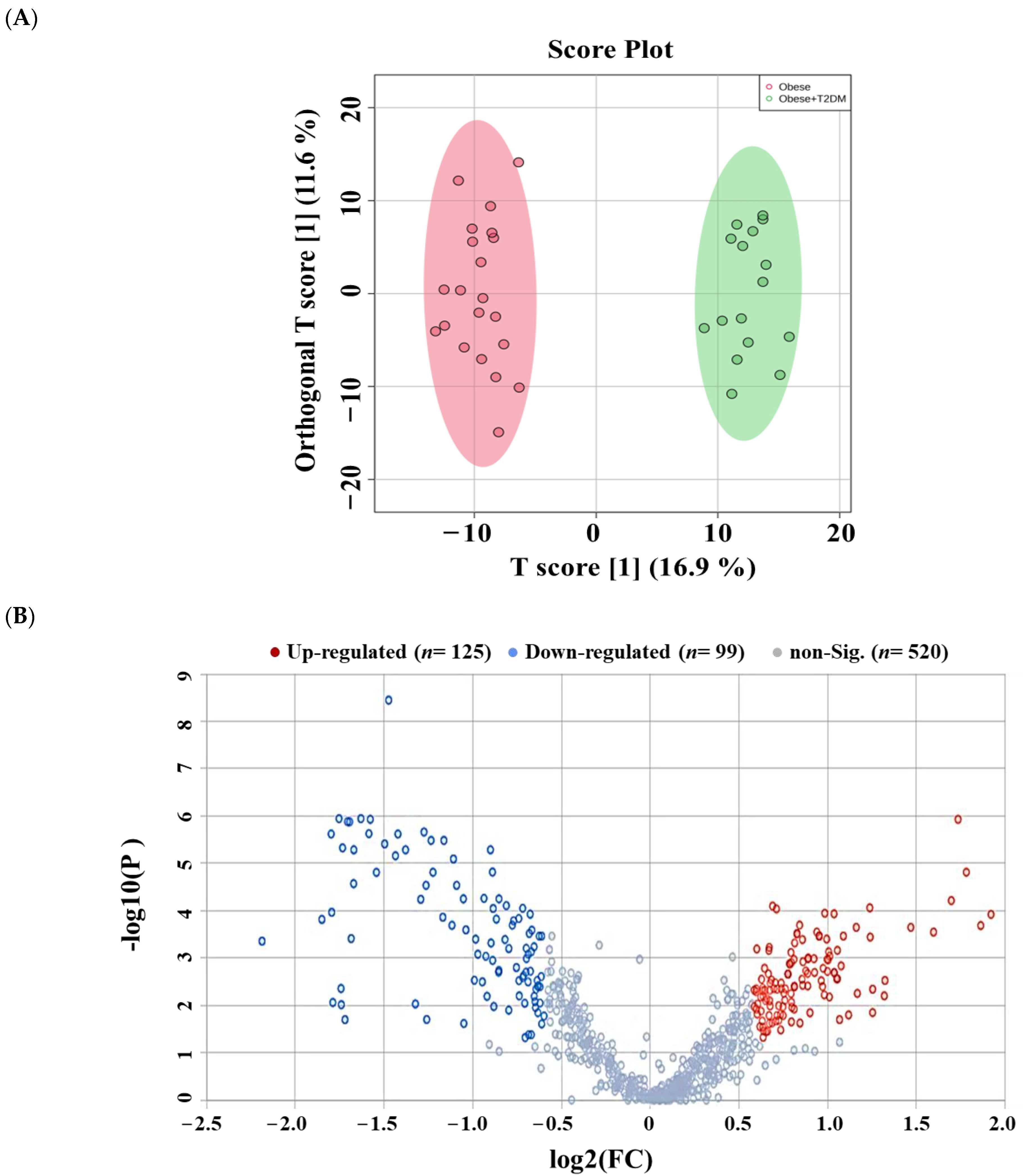
2.3. Lipidomic Alterations Associated with Obesity in Non-Diabetic Subjects
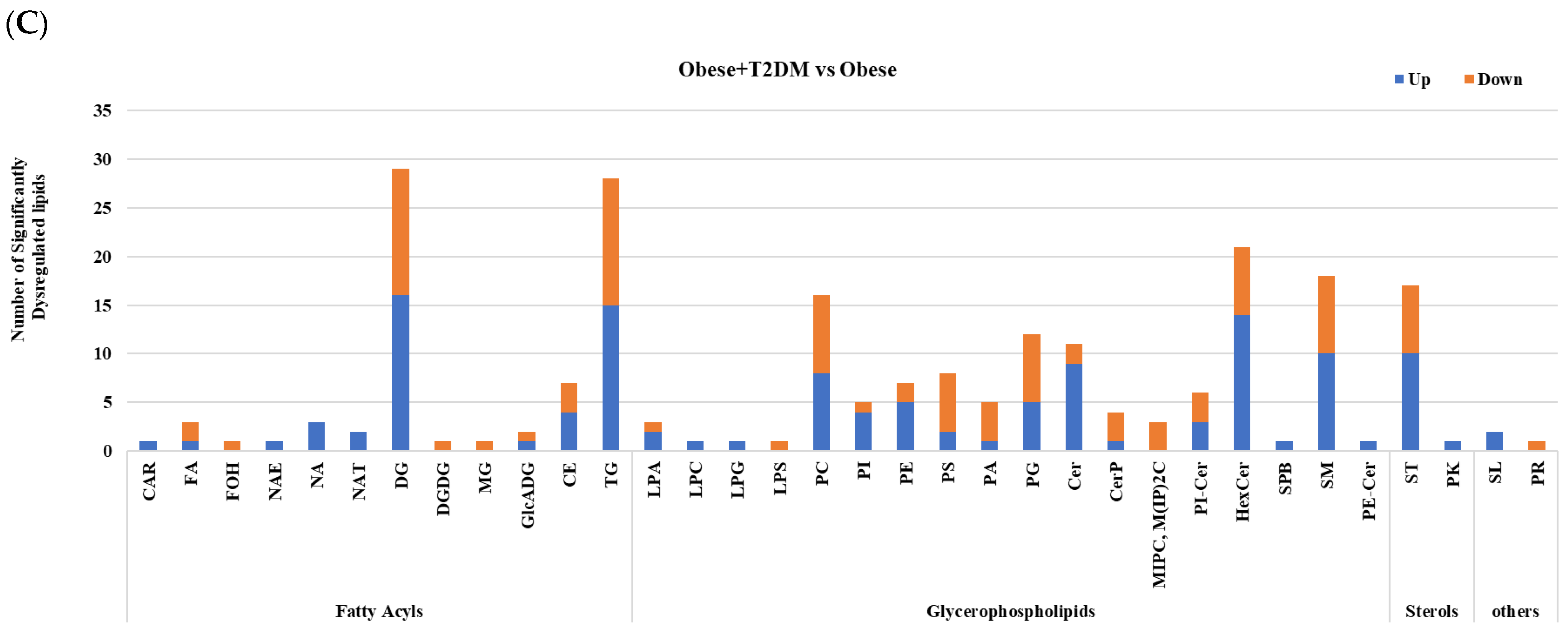
2.4. Lipidomic Alterations Associated with T2DM in Obese Patients
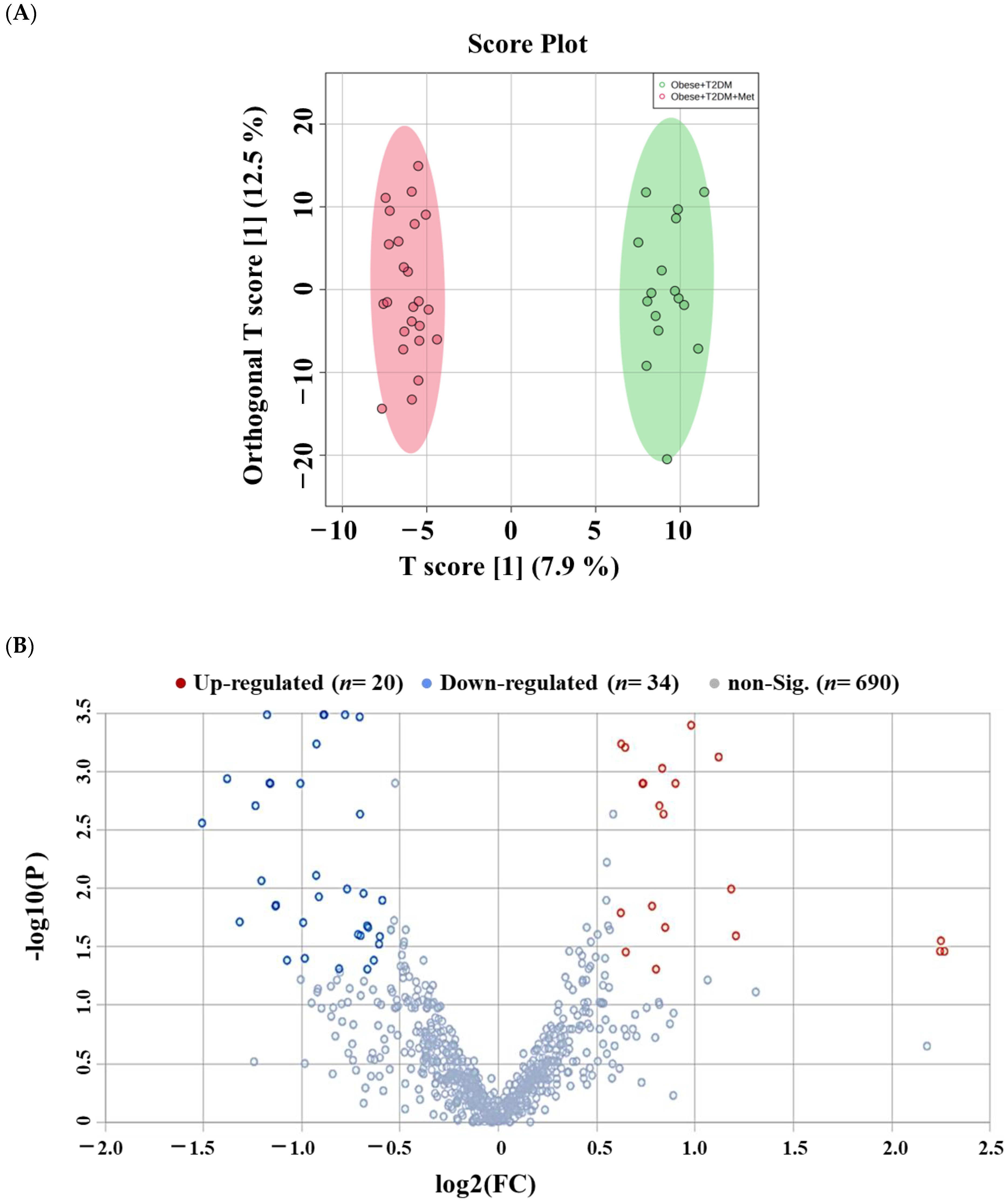
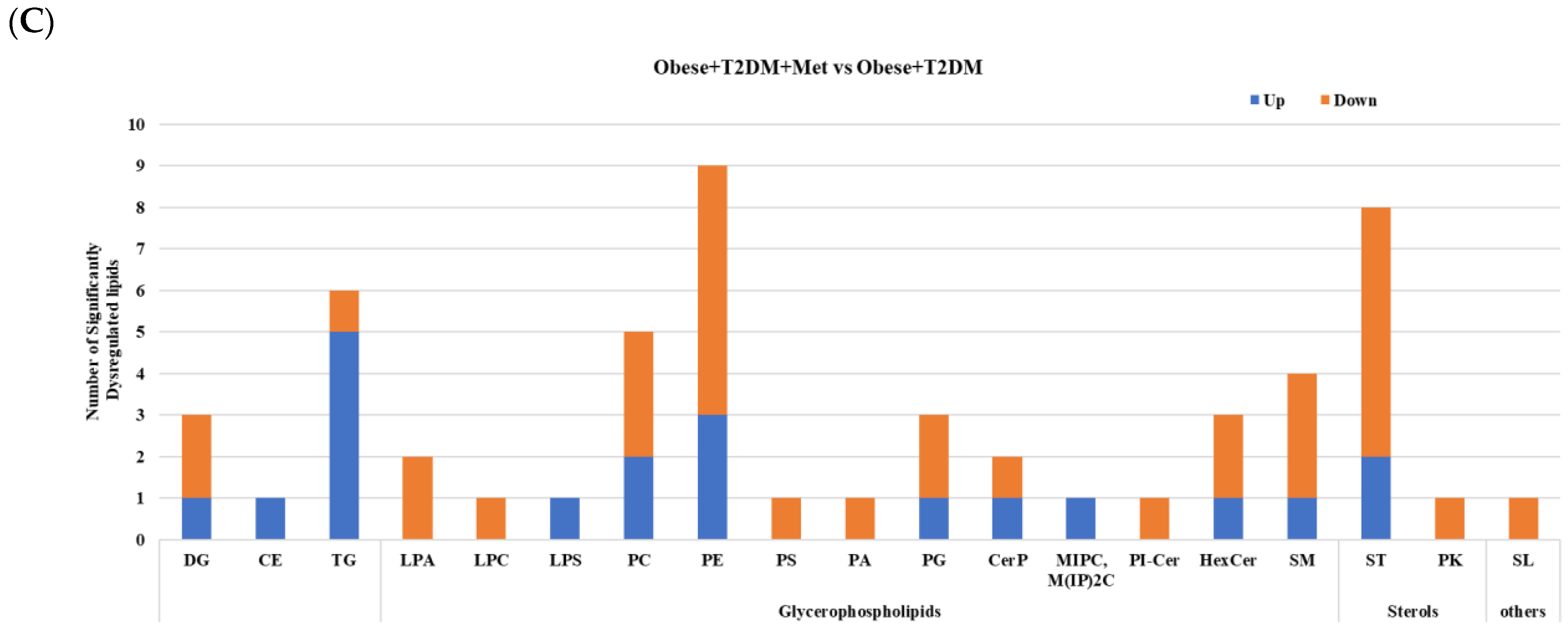
2.5. Lipidomic Alterations Associated with Metformin Administration in Obese Type 2 Diabetic Patients (OT2DM)
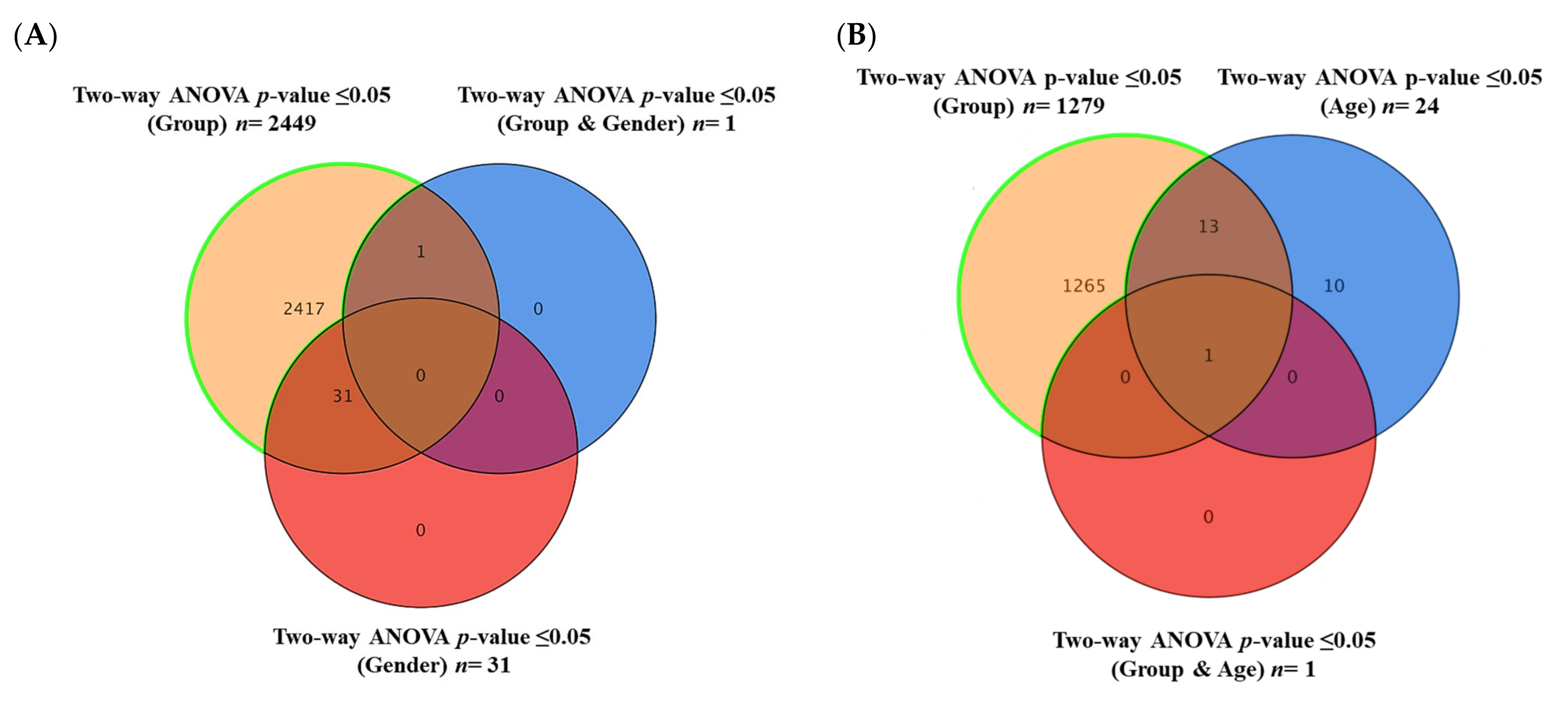
2.6. Commonly Altered Lipid Molecules as Effects of Obesity, T2DM, and Metformin Administration
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Ethics Statement
4.3. Anthropometric Measurements
4.4. Sample Preparation and Liquid Chromatography-Mass Spectrometry (LC-MS) Analysis
4.5. Data Processing
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chatterjee; Khunti, K.; Davies, M.J. Type 2 diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- WHO. Obesity and Overweight [Fact Sheet]. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 5 December 2023).
- Chadt, A.; Scherneck, S.; Joost, H.G.; Al-Hasani, H. Molecular links between Obesity and Diabetes: “Diabesity”. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Wondmkun, Y.T. Obesity, Insulin Resistance, and Type 2 Diabetes: Associations and Therapeutic Implications. Diabetes Metab. Syndr. Obesity Targets Ther. 2020, 13, 3611–3616. [Google Scholar] [CrossRef]
- Kwon, H.; Pessin, J.E. Adipokines Mediate Inflammation and Insulin Resistance. Front. Endocrinol. 2013, 4, 71. [Google Scholar] [CrossRef]
- Ruze, R.; Liu, T.; Zou, X.; Song, J.; Chen, Y.; Xu, R.; Yin, X.; Xu, Q. Obesity and type 2 diabetes mellitus: Connections in epidemiology, pathogenesis, and treatments. Front. Endocrinol. 2023, 14, 1161521. [Google Scholar] [CrossRef]
- Aleidi, S.; Issa, A.; Bustanji, H.; Khalil, M.; Bustanji, Y. Adiponectin serum levels correlate with insulin resistance in type 2 diabetic patients. Saudi Pharm. J. 2015, 23, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Parhofer, K.G. Interaction between Glucose and Lipid Metabolism: More than Diabetic Dyslipidemia. Diabetes Metab. J. 2015, 39, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Shi, Q.; Song, P.; Pan, X.-F.; Yang, X.; Chen, L.; He, Y.; Zong, G.; Zhu, Y.; Su, B.; et al. Global Burden Attributable to High Low-Density Lipoprotein-Cholesterol From 1990 to 2019. Front. Cardiovasc. Med. 2022, 9, 903126. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Aertgeerts, B.; Guyatt, G.; Bekkering, G.E.; Vandvik, P.O.; Khan, S.U.; Rodondi, N.; Jackson, R.; Reny, J.-L.; Al Ansary, L.; et al. PCSK9 inhibitors and ezetimibe for the reduction of cardiovascular events: A clinical practice guideline with risk-stratified recommendations. BMJ 2022, 377, e069066. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Rangel, E.; Inzucchi, S.E. Metformin: Clinical use in type 2 diabetes. Diabetologia 2017, 60, 1586–1593. [Google Scholar] [CrossRef] [PubMed]
- Dludla, P.V.; Nkambule, B.B.; Mazibuko-Mbeje, S.E.; Nyambuya, T.M.; Mxinwa, V.; Mokgalaboni, K.; Ziqubu, K.; Cirilli, I.; Marcheggiani, F.; Louw, J.; et al. Adipokines as a therapeutic target by metformin to improve metabolic function: A systematic review of randomized controlled trials. Pharmacol. Res. 2021, 163, 105219. [Google Scholar] [CrossRef] [PubMed]
- Andrzejewski, S.; Siegel, P.M.; St-Pierre, J. Metabolic Profiles Associated with Metformin Efficacy in Cancer. Front. Endocrinol. 2018, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Pu, R.; Shi, D.; Gan, T.; Ren, X.; Ba, Y.; Huo, Y.; Bai, Y.; Zheng, T.; Cheng, N. Effects of metformin in obesity treatment in different populations: A meta-analysis. Ther. Adv. Endocrinol. Metab. 2020, 11, 2042018820926000. [Google Scholar] [CrossRef] [PubMed]
- Seifarth, C.; Schehler, B.; Schneider, H.J. Effectiveness of Metformin on Weight Loss in Non-Diabetic Individuals with Obesity. Exp. Clin. Endocrinol. Diabetes 2013, 121, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.P. Metformin use in women with polycystic ovary syndrome. Ann. Transl. Med. 2014, 2, 56. [Google Scholar] [PubMed]
- Kristófi, R.; Eriksson, J.W. Metformin as an anti-inflammatory agent: A short review. J. Endocrinol. 2021, 251, R11–R22. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, I.; Hollenberg, M.D.; Ding, H.; Triggle, C.R. A critical review of the evidence that metformin is a putative anti-aging drug that enhances healthspan and extends lifespan. Front. Endocrinol. 2021, 12, 718942. [Google Scholar] [CrossRef]
- Kashi, Z.; Mahrooz, A.; Kianmehr, A.; Alizadeh, A. The Role of Metformin Response in Lipid Metabolism in Patients with Recent-Onset Type 2 Diabetes: HbA1c Level as a Criterion for Designating Patients as Responders or Nonresponders to Metformin. PLoS ONE 2016, 11, e0151543. [Google Scholar] [CrossRef]
- Lin, S.H.; Cheng, P.C.; Tu, S.T.; Hsu, S.R.; Cheng, Y.C.; Liu, Y.H. Effect of metformin monotherapy on serum lipid profile in statin-naïve individuals with newly diagnosed type 2 diabetes mellitus: A cohort study. PeerJ 2018, 6, e4578. [Google Scholar] [CrossRef]
- Yen, F.-S.; Wei, J.C.-C.; Chiu, L.-T.; Hsu, C.-C.; Hwu, C.-M. Cardiovascular outcomes of metformin use in patients with type 2 diabetes and chronic obstructive pulmonary disease. Front. Pharmacol. 2022, 13, 919881. [Google Scholar] [CrossRef]
- Aleidi, S.M.; Dahabiyeh, L.A.; Gu, X.; Al Dubayee, M.; Alshahrani, A.; Benabdelkamel, H.; Mujammami, M.; Li, L.; Aljada, A.; Rahman, A.M.A. Obesity Connected Metabolic Changes in Type 2 Diabetic Patients Treated with Metformin. Front. Pharmacol. 2020, 11, 616157. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Al Dubayee, M.; Alshahrani, A.; Masood, A.; Benabdelkamel, H.; Zahra, M.; Li, L.; Rahman, A.M.A.; Aljada, A. Distinctive Metabolomics Patterns Associated with Insulin Resistance and Type 2 Diabetes Mellitus. Front. Mol. Biosci. 2020, 7, 609806. [Google Scholar] [CrossRef]
- Dahabiyeh, L.A.; Mujammami, M.; Arafat, T.; Benabdelkamel, H.; Alfadda, A.A.; Rahman, A.M.A. A Metabolic Pattern in Healthy Subjects Given a Single Dose of Metformin: A Metabolomics Approach. Front. Pharmacol. 2021, 12, 705932. [Google Scholar] [CrossRef]
- Dahabiyeh, L.A.; Mujammami, M.; AlMalki, R.H.; Arafat, T.; Benabdelkamel, H.; Alfadda, A.A.; Rahman, A.M.A. Lipids Alterations Associated with Metformin in Healthy Subjects: An Investigation Using Mass Spectrometry Shotgun Approach. Int. J. Mol. Sci. 2022, 23, 11478. [Google Scholar] [CrossRef]
- Pernicova, I.; Korbonits, M. Metformin—Mode of action and clinical implications for diabetes and cancer. Nat. Rev. Endocrinol. 2014, 10, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Beyene, H.B.; Olshansky, G.; TSmith, A.A.; Giles, C.; Huynh, K.; Cinel, M.; Mellett, N.A.; Cadby, G.; Hung, J.; Hui, J.; et al. High-coverage plasma lipidomics reveals novel sex-specific lipidomic fingerprints of age and BMI: Evidence from two large population cohort studies. PLoS Biol. 2020, 18, e3000870. [Google Scholar]
- Beyene, H.B.; Olshansky, G.; Giles, C.; Huynh, K.; Cinel, M.; Mellett, N.A.; Smith, A.A.T.; Shaw, J.E.; Magliano, D.J.; Meikle, P.J. Lipidomic Signatures of Changes in Adiposity: A Large Prospective Study of 5849 Adults from the Australian Diabetes, Obesity and Lifestyle Study. Metabolites 2021, 11, 646. [Google Scholar] [CrossRef]
- Rauschert, S.; Uhl, O.; Koletzko, B.; Mori, T.A.; Beilin, L.J.; Oddy, W.H.; Hellmuth, C. Sex differences in the association of phospholipids with components of the metabolic syndrome in young adults. Biol. Sex Differ. 2017, 8, 10. [Google Scholar] [CrossRef]
- Hosseinkhani, S.; Arjmand, B.; Dilmaghani-Marand, A.; Mohammadi Fateh, S.; Dehghanbanadaki, H.; Najjar, N.; Alavi-Moghadam, S.; Ghodssi-Ghassemabadi, R.; Nasli-Esfahani, E.; Farzadfar, F.; et al. Targeted metabolomics analysis of amino acids and acylcarnitines as risk markers for diabetes by LC-MS/MS technique. Sci. Rep. 2022, 12, 8418. [Google Scholar] [CrossRef]
- Holland, W.L.; Summers, S.A. Sphingolipids, Insulin Resistance, and Metabolic Disease: New Insights from in Vivo Manipulation of Sphingolipid Metabolism. Endocr. Rev. 2008, 29, 381–402. [Google Scholar] [CrossRef]
- Amati, F.; Dubé, J.J.; Alvarez-Carnero, E.; Edreira, M.M.; Chomentowski, P.; Coen, P.M.; Switzer, G.E.; Bickel, P.E.; Stefanovic-Racic, M.; Toledo, F.G.; et al. Skeletal muscle triglycerides, diacylglycerols, and ceramides in insulin resistance: Another paradox in endurance-trained athletes? Diabetes 2011, 60, 2588–2597. [Google Scholar] [CrossRef]
- Boini, K.M.; Koka, S.; Xia, M.; Ritter, J.K.; Gehr, T.W.; Li, P.L. Sphingolipids in obesity and related complications. Front. Biosci. Landmark Ed. 2017, 22, 96–116. [Google Scholar] [PubMed]
- Liu, J.; Bai, L.; Wang, W.; Song, Y.; Zhao, W.; Li, Q.; Wu, Q. LC-MS-Based Lipidomic Analysis of Serum Samples from Patients with Type 2 Diabetes Mellitus (T2DM). Dis. Markers 2022, 2022, 5559470. [Google Scholar] [CrossRef]
- Yang, T.; Zhao, J.; Liu, F.; Li, Y. Lipid metabolism and endometrial receptivity. Hum. Reprod. Update 2022, 28, 858–889. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T. Pathophysiology of Diabetic Dyslipidemia. J. Atheroscler. Thromb. 2018, 25, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Patterson, A.D.; Idle, J.R.; Gonzalez, F.J. Xenobiotic metabolomics: Major impact on the metabolome. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 37–56. [Google Scholar] [CrossRef] [PubMed]
- Harper, C.R.; Jacobson, T.A. Using apolipoprotein B to manage dyslipidemic patients: Time for a change? Mayo Clin. Proc. 2010, 85, 440–445. [Google Scholar] [CrossRef]
- Suvitaival, T.; Bondia-Pons, I.; Yetukuri, L.; Pöhö, P.; Nolan, J.J.; Hyötyläinen, T.; Kuusisto, J.; Orešič, M. Lipidome as a predictive tool in progression to type 2 diabetes in Finnish men. Metabolism 2018, 78, 1–12. [Google Scholar] [CrossRef]
- Yun, H.; Sun, L.; Wu, Q.; Zong, G.; Qi, Q.; Li, H.; Zheng, H.; Zeng, R.; Liang, L.; Lin, X. Associations among circulating sphingolipids, β-cell function, and risk of developing type 2 diabetes: A population-based cohort study in China. PLoS Med. 2020, 17, e1003451. [Google Scholar] [CrossRef]
- Morze, J.; Wittenbecher, C.; Schwingshackl, L.; Danielewicz, A.; Rynkiewicz, A.; Hu, F.B.; Guasch-Ferré, M. Metabolomics and Type 2 Diabetes Risk: An Updated Systematic Review and Meta-analysis of Prospective Cohort Studies. Diabetes Care 2022, 45, 1013–1024. [Google Scholar] [CrossRef]
- Sokolowska, E.; Blachnio-Zabielska, A. The Role of Ceramides in Insulin Resistance. Front. Endocrinol. 2019, 10, 577. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.J.; Hajduch, E.; Kular, G.; Hundal, H.S. Ceramide disables 3-phosphoinositide binding to the pleckstrin homology domain of protein kinase B (PKB)/Akt by a PKCzeta-dependent mechanism. Mol. Cell. Biol. 2003, 23, 7794–7808. [Google Scholar] [CrossRef] [PubMed]
- Prentki; Madiraju, S.R. Glycerolipid/free fatty acid cycle and islet β-cell function in health, obesity and diabetes. Mol. Cell. Endocrinol. 2012, 353, 88–100. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, P.D.; Guo, K.; Eid, S.A.; Rumora, A.E.; Hinder, L.M.; Hayes, J.M.; Mendelson, F.E.; Hur, J.; Feldman, E.L. Integrated lipidomic and transcriptomic analyses identify altered nerve triglycerides in mouse models of prediabetes and type 2 diabetes. Dis. Models Mech. 2020, 13, dmm042101. [Google Scholar] [CrossRef]
- Chen, Z.; Liang, Q.; Wu, Y.; Gao, Z.; Kobayashi, S.; Patel, J.; Li, C.; Cai, F.; Zhang, Y.; Liang, C.; et al. Comprehensive lipidomic profiling in serum and multiple tissues from a mouse model of diabetes. Metabolomics 2020, 16, 115. [Google Scholar] [CrossRef]
- Scheidemantle, G.; Duan, L.; Lodge, M.; Cummings, M.J.; Hilovsky, D.; Pham, E.; Wang, X.; Kennedy, A.; Liu, X. Data-dependent and -independent acquisition lipidomics analysis reveals the tissue-dependent effect of metformin on lipid metabolism. Res. Sq. 2023, in press. [Google Scholar] [CrossRef]
- Pradas, I.; Rovira-Llopis, S.; Naudí, A.; Bañuls, C.; Rocha, M.; Hernandez-Mijares, A.; Pamplona, R.; Victor, V.M.; Jové, M. Metformin induces lipid changes on sphingolipid species and oxidized lipids in polycystic ovary syndrome women. Sci. Rep. 2019, 9, 16033. [Google Scholar] [CrossRef]
- Victor, V.M.; Rovira-Llopis, S.; Bañuls, C.; Diaz-Morales, N.; Castelló, R.; Falcón, R.; Gómez, M.; Rocha, M.; Hernández-Mijares, A. Effects of metformin on mitochondrial function of leukocytes from polycystic ovary syndrome patients with insulin resistance. Eur. J. Endocrinol. 2015, 173, 683–691. [Google Scholar] [CrossRef]
- Kim, H.-S.; Ren, G.; Kim, T.; Bhatnagar, S.; Yang, Q.; Bahk, Y.Y.; Kim, J.-A. Metformin reduces saturated fatty acid-induced lipid accumulation and inflammatory response by restoration of autophagic flux in endothelial cells. Sci. Rep. 2020, 10, 13523. [Google Scholar] [CrossRef]
- Zhou, G.; Myers, R.; Li, Y.; Chen, Y.; Shen, X.; Fenyk-Melody, J.; Wu, M.; Ventre, J.; Doebber, T.; Fujii, N.; et al. Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Investig. 2001, 108, 1167–1174. [Google Scholar] [CrossRef]
- Green, C.J.; Marjot, T.; Walsby-Tickle, J.; Charlton, C.; Cornfield, T.; Westcott, F.; Pinnick, K.E.; Moolla, A.; Hazlehurst, J.M.; McCullagh, J.; et al. Metformin maintains intrahepatic triglyceride content through increased hepatic de novo lipogenesis. Eur. J. Endocrinol. 2022, 186, 367–377. [Google Scholar] [CrossRef]
- Gormsen, L.C.; Søndergaard, E.; Christensen, N.L.; Jakobsen, S.; Nielsen, E.H.; Munk, O.L.; Tolbod, L.P.; Jessen, N.; Nielsen, S. Metformin does not affect postabsorptive hepatic free fatty acid uptake, oxidation or resecretion in humans: A 3-month placebo-controlled clinical trial in patients with type 2 diabetes and healthy controls. Diabetes Obes. Metab. 2018, 20, 1435–1444. [Google Scholar] [CrossRef] [PubMed]
- Buzatto, A.Z.; Jabar, M.A.; Nizami, I.; Dasouki, M.; Li, L.; Rahman, A.M.A. Lipidome Alterations Induced by Cystic Fibrosis, CFTR Mutation, and Lung Function. J. Proteome Res. 2021, 20, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Arora, V.; Campbell, J.N.; Chung, M.K. Fight fire with fire: Neurobiology of capsaicin-induced analgesia for chronic pain. Pharmacol. Ther. 2021, 220, 107743. [Google Scholar] [CrossRef] [PubMed]
- Ilonen, J.; Lempainen, J.; Veijola, R. The heterogeneous pathogenesis of type 1 diabetes mellitus. Nat. Rev. Endocrinol. 2019, 15, 635–650. [Google Scholar] [CrossRef] [PubMed]
- Easton, Z.J.W.; Sarr, O.; Zhao, L.; Buzatto, A.Z.; Luo, X.; Zhao, S.; Li, L.; Regnault, T.R.H. An Integrated Multi-OMICS Approach Highlights Elevated Non-Esterified Fatty Acids Impact BeWo Trophoblast Metabolism and Lipid Processing. Metabolites 2023, 13, 883. [Google Scholar] [CrossRef]
- Roychaudhuri, R.; Atashi, H.; Snyder, S.H. Serine Racemase mediates subventricular zone neurogenesis via fatty acid metabolism. Stem Cell Rep. 2023, 18, 1482–1499. [Google Scholar] [CrossRef]
- McCann, J.; Sosa-Miranda, C.D.; Guo, H.; Reshke, R.; Savard, A.; Buzatto, A.Z.; Taylor, J.A.; Li, L.; Gibbings, D.J. Contaminating transfection complexes can masquerade as small extracellular vesicles and impair their delivery of RNA. J. Extracell. Vesicles 2022, 11, e12220. [Google Scholar] [CrossRef]
- Buzatto, A.Z.; Tatlay, J.; Bajwa, B.; Mung, D.; Camicioli, R.; Dixon, R.A.; Li, L. Comprehensive Serum Lipidomics for Detecting Incipient Dementia in Parkinson’s Disease. J. Proteome Res. 2021, 20, 4053–4067. [Google Scholar] [CrossRef]
- Yau, H.; Rivera, K.; Lomonaco, R.; Cusi, K. The future of thiazolidinedione therapy in the management of type 2 diabetes mellitus. Curr. Diabetes Rep. 2013, 13, 329–341. [Google Scholar] [CrossRef]
- Aoyagi, R.; Ikeda, K.; Isobe, Y.; Arita, M. Comprehensive analyses of oxidized phospholipids using a measured MS/MS spectra library. J. Lipid Res. 2017, 58, 2229–2237. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Köfeler, H.C.; Ahrends, R.; Baker, E.S.; Ekroos, K.; Han, X.; Hoffmann, N.; Holčapek, M.; Wenk, M.R.; Liebisch, G. Recommendations for good practice in MS-based lipidomics. J. Lipid Res. 2021, 62, 100138. [Google Scholar] [CrossRef]
- Köfeler, H.C.; Eichmann, T.O.; Ahrends, R.; Bowden, J.A.; Danne-Rasche, N.; Dennis, E.A.; Fedorova, M.; Griffiths, W.J.; Han, X.; Hartler, J.; et al. Quality control requirements for the correct annotation of lipidomics data. Nat. Commun. 2021, 12, 4771. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Non-Diabetic (n = 49) | Diabetic (n = 40) | ||||||
|---|---|---|---|---|---|---|---|---|
| Control (n = 28) | Obese (n = 21) | OT2DM (n = 16) | OT2DMMet (n = 24) | |||||
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |
| Age (Y) | 26 | 11.5 | 34.9 * | 22.4 | 48.7 *§ | 22.9 | 46.4 *§ | 22.2 |
| Gender (M/F) | 17/17 # | - | 5/16 # | - | 11/5 # | - | 9/15 # | - |
| BMI (kg/m2) | 23.1 | 2.9 | 38.7 * | 17.5 | 33.6 * | 14.9 | 39.7 §ǂ | 14.3 |
| FBG (mmol/L) | 5.1 | 1 | 5.3 | 1 | 10.2 *§ | 9.7 | 9.6 *§ | 7.5 |
| HbA1c (%) | 5.6 | 0.5 | 5.2 | 3.4 | 8.4 *§ | 5.6 | 8.6 *§ | 3.9 |
| LDL (mmol/L) | 2.6 | 1.7 | 3.1 | 1.6 | 3.4 * | 1.7 | 2.5 §ǂ | 1.5 |
| HDL (mmol/L) | 1.4 | 0.4 | 1.2 | 0.5 | 1 *§ | 0.4 | 1 *§ | 0.4 |
| TG (mmol/L) | 0.8 | 0.6 | 1.1 | 1 | 2.2 *§ | 1.9 | 1.3 *ǂ | 0.9 |
| Lipid Molecules Characteristics | BMI Panel | T2DM Panel | Metformin Panel | ||||
|---|---|---|---|---|---|---|---|
| Subclass | Level of Identification | Compound | RT (min) | Mass (m/z) | [Obese] vs. [Control] | ([OT2DM] vs. [Obese]) | [OT2DMMet] vs. [OT2DM] |
| FC * | FC * | FC * | |||||
| ST | Tier 3 | ST 26:0;O4;H | 1.5 | 602.4306 | ↑1.94 *** | ↓0.31 *** | ↑2.17 *** |
| MIPC | Tier 3 | MIPC 32:4;O3 | 8.57 | 941.5348 | ↑1.95 *** | ↓0.41 *** | ↑1.74 * |
| PG | Tier 3 | PG 40:5 | 9.58 | 847.548 | ↑3.10 ** | ↓0.29 ** | ↑4.74 * |
| TG | Tier 3 | TG O-76:6 | 13.7 | 1196.091 | ↑3.13 ** | ↓0.30 * | ↑4.76 * |
| Tier 3 | TG 52:4 | 18.8 | 872.7744 | ↑1.96 ** | ↓0.28 *** | ↑2.31 * | |
| PC | Tier 1 | PC O-14:1_22:1 | 12.32 | 772.6243 | ↑2.49 * | ↓0.30 ** | ↑4.82 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mujammami, M.; Aleidi, S.M.; Buzatto, A.Z.; Alshahrani, A.; AlMalki, R.H.; Benabdelkamel, H.; Al Dubayee, M.; Li, L.; Aljada, A.; Abdel Rahman, A.M. Lipidomics Profiling of Metformin-Induced Changes in Obesity and Type 2 Diabetes Mellitus: Insights and Biomarker Potential. Pharmaceuticals 2023, 16, 1717. https://doi.org/10.3390/ph16121717
Mujammami M, Aleidi SM, Buzatto AZ, Alshahrani A, AlMalki RH, Benabdelkamel H, Al Dubayee M, Li L, Aljada A, Abdel Rahman AM. Lipidomics Profiling of Metformin-Induced Changes in Obesity and Type 2 Diabetes Mellitus: Insights and Biomarker Potential. Pharmaceuticals. 2023; 16(12):1717. https://doi.org/10.3390/ph16121717
Chicago/Turabian StyleMujammami, Muhammad, Shereen M. Aleidi, Adriana Zardini Buzatto, Awad Alshahrani, Reem H. AlMalki, Hicham Benabdelkamel, Mohammed Al Dubayee, Liang Li, Ahmad Aljada, and Anas M. Abdel Rahman. 2023. "Lipidomics Profiling of Metformin-Induced Changes in Obesity and Type 2 Diabetes Mellitus: Insights and Biomarker Potential" Pharmaceuticals 16, no. 12: 1717. https://doi.org/10.3390/ph16121717
APA StyleMujammami, M., Aleidi, S. M., Buzatto, A. Z., Alshahrani, A., AlMalki, R. H., Benabdelkamel, H., Al Dubayee, M., Li, L., Aljada, A., & Abdel Rahman, A. M. (2023). Lipidomics Profiling of Metformin-Induced Changes in Obesity and Type 2 Diabetes Mellitus: Insights and Biomarker Potential. Pharmaceuticals, 16(12), 1717. https://doi.org/10.3390/ph16121717