
Soloxolone Methyl Reduces the Stimulatory Effect of Leptin on the Aggressive Phenotype of Murine Neuro2a Neuroblastoma Cells via the MAPK/ERK1/2 Pathway

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
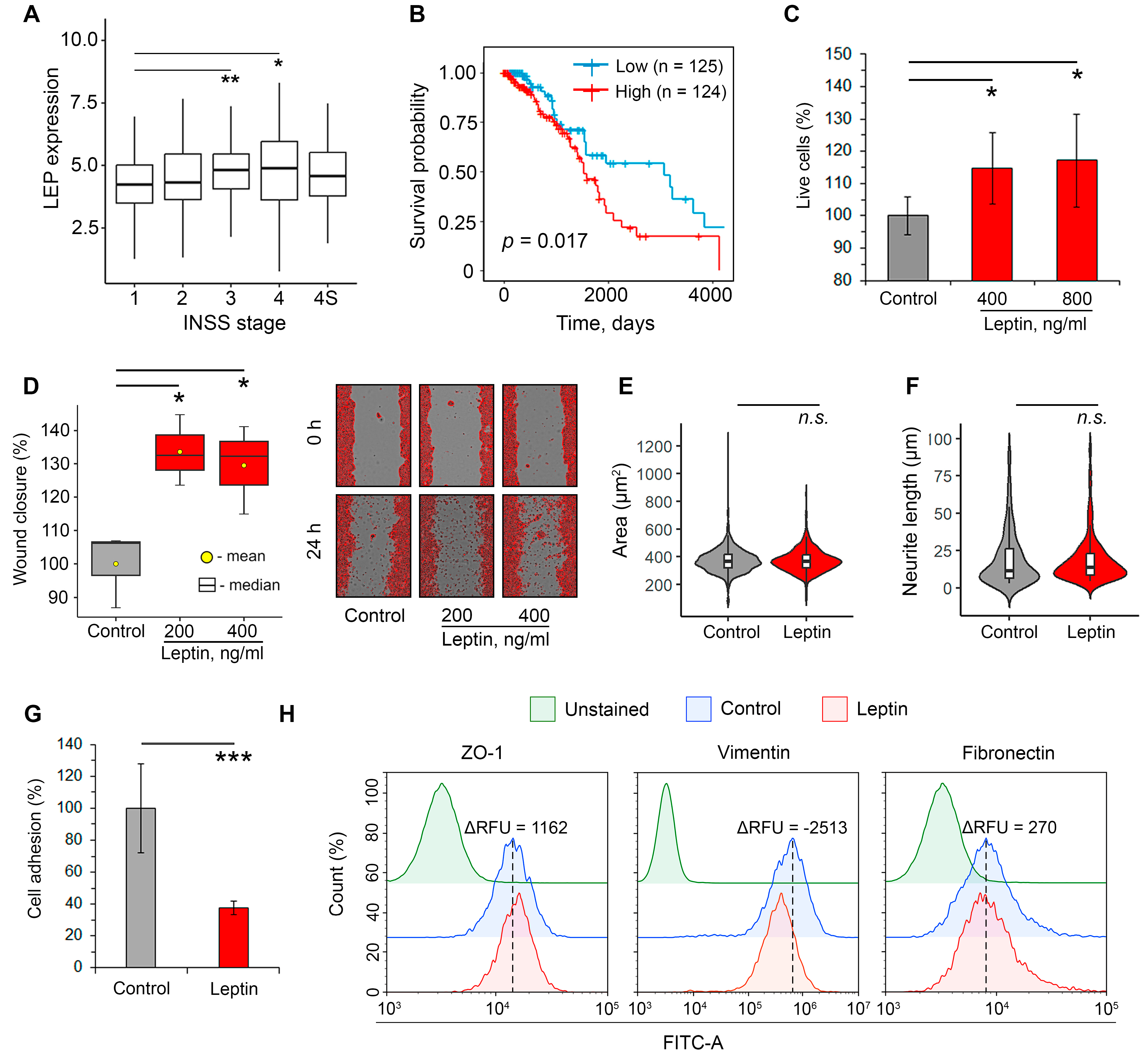
2.1. Leptin Promotes Aggressiveness in Neuroblastoma Cells
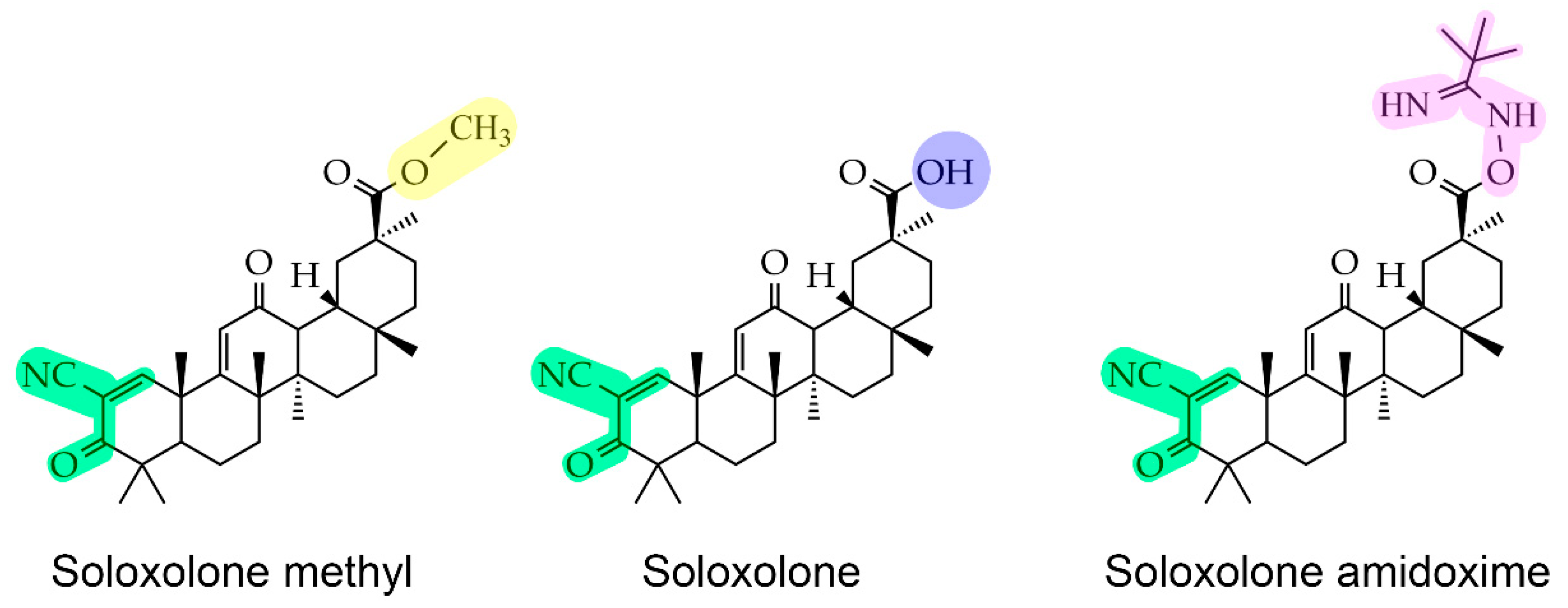
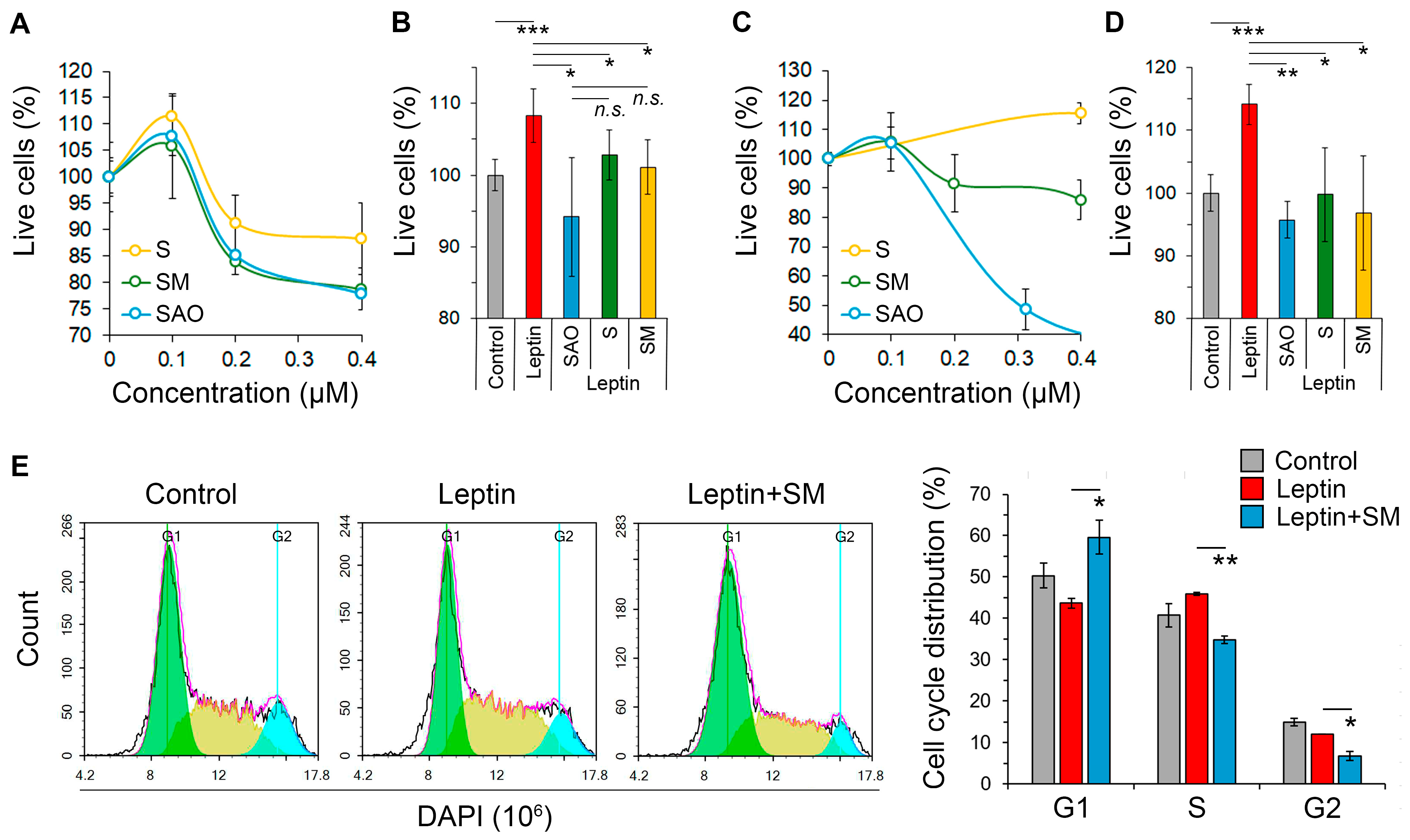
2.2. Cyanoenone-Bearing Triterpenoids Effectively Inhibit the Proliferative Response of Neuroblastoma Cells to Leptin
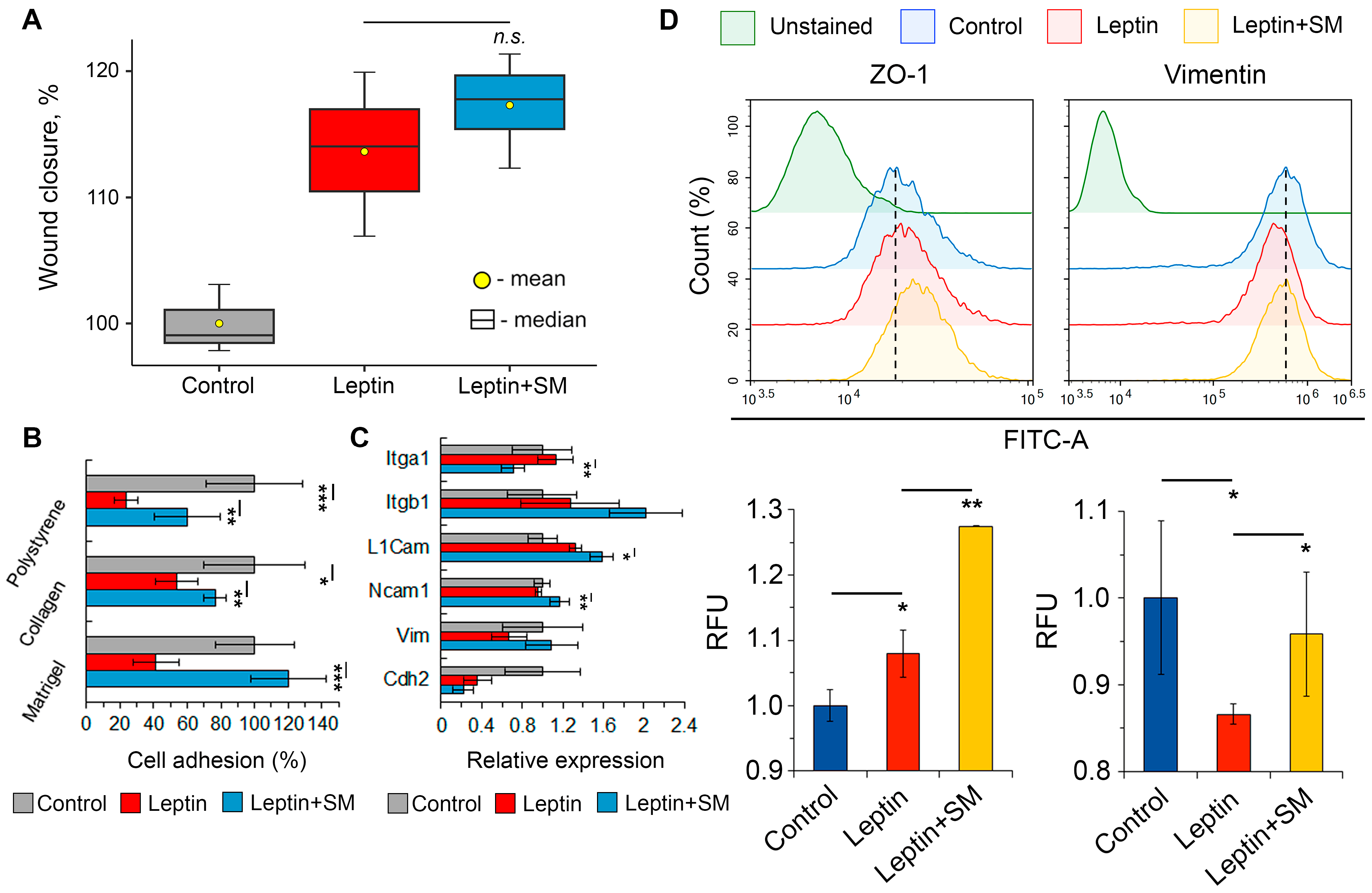
2.3. SM Improves the Adhesiveness of Neuroblastoma Cells Stimulated with Leptin
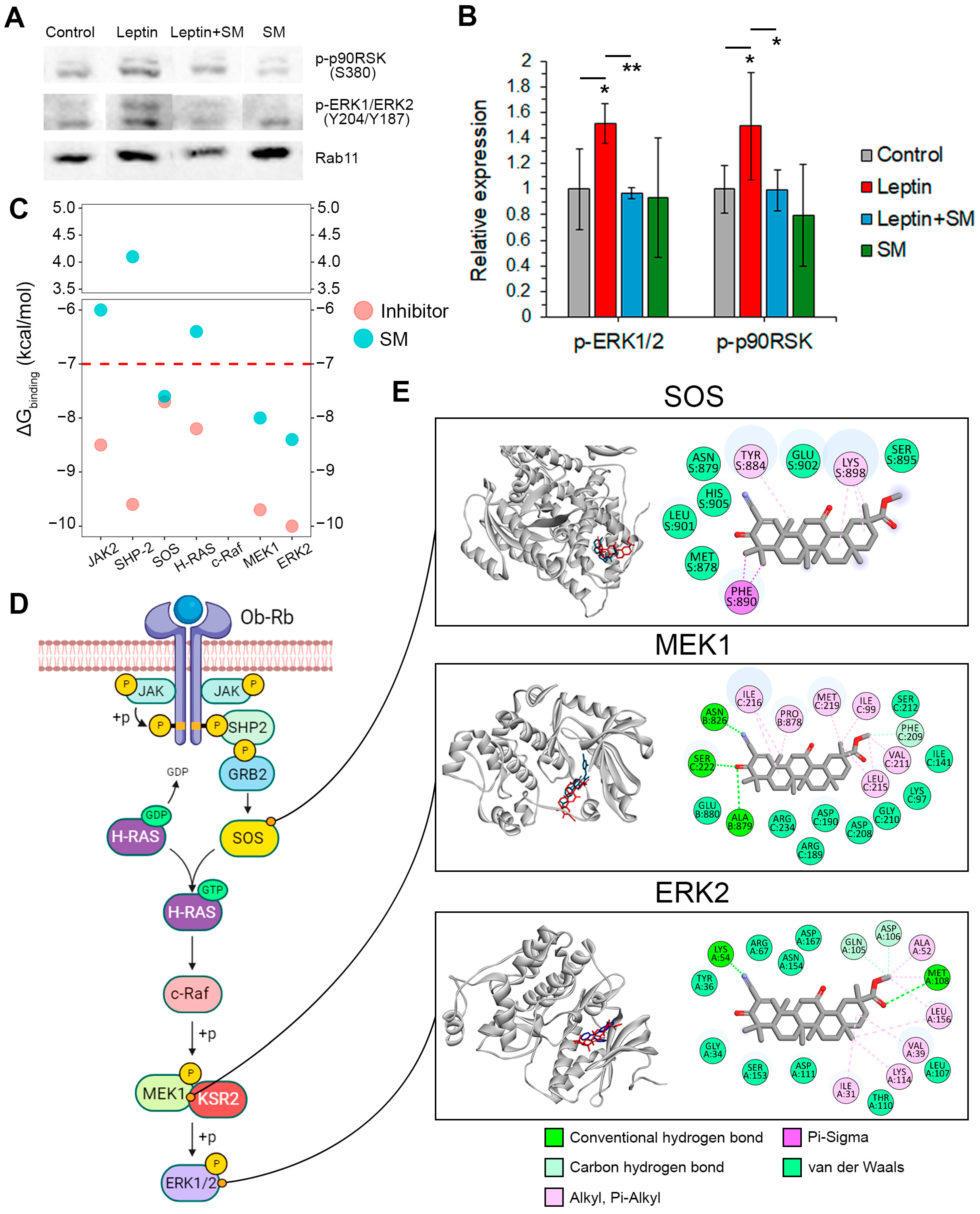
2.4. SM Inhibits the Induction of the ERK1/2 Pathway by Leptin
- The SM binding pocket in SOS was similar to that of the known SOS inhibitor CHEMBL4166974 and contained Pi-alkyl and Pi-sigma interactions with Tyr884, Lys898, and Phe890, which were involved in a hydrophobic pocket at the contact surface of SOS and H-Ras [32].
3. Discussion
Limitations of the Study
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Lines
4.3. Biological Evaluations
4.3.1. Cell Viability Analysis
4.3.2. Flow Cytometry
4.3.3. Wound Healing Assay
4.3.4. Morphological Analysis
4.3.5. Neuritogenesis Assay
4.3.6. Adhesion Assay
4.3.7. RT-qPCR
4.3.8. Western Blotting
4.4. Bioinformatics and Data Analysis
4.4.1. Microarray Data Analysis
4.4.2. Molecular Docking
4.4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CAMs | cell adhesion molecules |
| CDDO-Im | 2-cyano-3, 12-dioxooleana-1, 9-dien-28-imidazolide |
| DMEM | Dulbecco’s modified Eagle’s medium |
| EMT | epithelial-mesenchymal transition |
| ERK1/2 | extracellular signal regulated kinase 1/2 |
| FBS | fetal bovine serum |
| INSS | International Neuroblastoma Staging System |
| L1cam | cell adhesion molecule L1 |
| LEP | leptin |
| MAPK | mitogen-activated protein kinase |
| MEK1 | mitogen-activated protein kinase kinase 1 |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NC | neural crest |
| Ncam1 | neural cell adhesion molecule 1 |
| p90RSK | ribosomal protein S6 kinase A1 |
| ROHAD | rapid-onset obesity with hypothalamic dysfunction, hypoventilation, and autonomic dysregulation |
| S | soloxolone |
| SAO | soloxolone amidoxime |
| SM | soloxolone methyl |
| SOS | son of sevenless homolog 1 |
References
- Maguire, L.H.; Thomas, A.R.; Goldstein, A.M. Tumors of the neural crest: Common themes in development and cancer. Dev. Dyn. 2015, 244, 311–322. [Google Scholar] [CrossRef]
- Johnsen, J.I.; Dyberg, C.; Fransson, S.; Wickström, M. Molecular mechanisms and therapeutic targets in neuroblastoma. Pharmacol. Res. 2018, 131, 164–176. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, X.; Li, N.; Guo, Y.; Yang, X.; Lei, Y. Therapy resistance in neuroblastoma: Mechanisms and reversal strategies. Front. Pharmacol. 2023, 14, 1114295. [Google Scholar] [CrossRef]
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K. Body Fatness and Cancer—Viewpoint of the IARC Working Group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef]
- Olszańska, J.; Pietraszek-Gremplewicz, K.; Nowak, D. Melanoma Progression under Obesity: Focus on Adipokines. Cancers 2021, 13, 2281. [Google Scholar] [CrossRef]
- Lazea, C.; Sur, L.; Florea, M. ROHHAD (Rapid-onset Obesity with Hypoventilation, Hypothalamic Dysfunction, Autonomic Dysregulation) Syndrome—What Every Pediatrician Should Know About the Etiopathogenesis, Diagnosis and Treatment: A Review. Int. J. Gen. Med. 2021, 14, 319–326. [Google Scholar] [CrossRef]
- IJpma, I.; Lequin, M.H.; Nievelstein, R.A.J.; Fiocco, M.; Tissing, W.J.E. Body composition of patients with neuroblastoma using computed tomography. Pediatr. Blood Cancer 2021, 68, e29337. [Google Scholar] [CrossRef]
- Jiménez-Cortegana, C.; López-Saavedra, A.; Sánchez-Jiménez, F.; Pérez-Pérez, A.; Castiñeiras, J.; Virizuela-Echaburu, J.A.; de la Cruz-Merino, L.D.; Sánchez-Margalet, V. Leptin, Both Bad and Good Actor in Cancer. Biomolecules 2021, 11, 913. [Google Scholar] [CrossRef]
- Parida, S.; Siddharth, S.; Sharma, D. Adiponectin, Obesity, and Cancer: Clash of the Bigwigs in Health and Disease. Int. J. Mol. Sci. 2019, 20, 2519. [Google Scholar] [CrossRef]
- Tümmler, C.; Snapkov, I.; Wickström, M.; Moens, U.; Ljungblad, L.; Helena Maria Elfman, L.; Winberg, J.-O.; Kogner, P.; Inge Johnsen, J.; Sveinbjørnsson, B. Inhibition of chemerin/CMKLR1 axis in neuroblastoma cells reduces clonogenicity and cell viability in vitro and impairs tumor growth in vivo. Oncotarget 2017, 8, 95135. [Google Scholar] [CrossRef]
- Ho, P.W.-L.; Liu, H.-F.; Ho, J.W.-M.; Zhang, W.-Y.; Chu, A.C.-Y.; Kwok, K.H.-H.; Ge, X.; Chan, K.-H.; Ramsden, D.B.; Ho, S.-L. Mitochondrial Uncoupling Protein-2 (UCP2) Mediates Leptin Protection Against MPP+ Toxicity in Neuronal Cells. Neurotox. Res. 2010, 17, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Thon, M.; Hosoi, T.; Yoshii, M.; Ozawa, K. Leptin induced GRP78 expression through the PI3K-mTOR pathway in neuronal cells. Sci. Rep. 2014, 4, 7096. [Google Scholar] [CrossRef] [PubMed]
- Avtanski, D.B.; Nagalingam, A.; Kuppusamy, P.; Bonner, M.Y.; Arbiser, J.L.; Saxena, N.K.; Sharma, D. Honokiol abrogates leptin-induced tumor progression by inhibiting Wnt1-MTA1-β-catenin signaling axis in a microRNA-34a dependent manner. Oncotarget 2015, 6, 16396–16410. [Google Scholar] [CrossRef] [PubMed]
- Dalimi-Asl, S.; Babaahmadi-Rezaei, H.; Mohammadzadeh, G. Combination of Silibinin and Curcumin Reduced Leptin Receptor Expression in MCF-7 Human Breast Cancer Cell Line. Iran. J. Med. Sci. 2020, 45, 477–484. [Google Scholar] [PubMed]
- Yu, Y.-M.; Tsai, C.-C.; Tzeng, Y.-W.; Chang, W.-C.; Chiang, S.-Y.; Lee, M.-F. Ursolic acid suppresses leptin-induced cell proliferation in rat vascular smooth muscle cells. Can. J. Physiol. Pharmacol. 2017, 95, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-H.; Cui, X.-Y.; Wang, D.; Jin, Y.; Guan, Y.-X. Anti-obesity effect of escin: A study on high-fat diet-induced obese mice. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 7797–7812. [Google Scholar] [CrossRef]
- Markov, A.V.; Ilyina, A.A.; Salomatina, O.V.; Sen’kova, A.V.; Okhina, A.A.; Rogachev, A.D.; Salakhutdinov, N.F.; Zenkova, M.A. Novel Soloxolone Amides as Potent Anti-Glioblastoma Candidates: Design, Synthesis, In Silico Analysis and Biological Activities In Vitro and In Vivo. Pharmaceuticals 2022, 15, 603. [Google Scholar] [CrossRef]
- Markov, A.V.; Odarenko, K.V.; Sen’kova, A.V.; Salomatina, O.V.; Salakhutdinov, N.F.; Zenkova, M.A. Cyano Enone-Bearing Triterpenoid Soloxolone Methyl Inhibits Epithelial-Mesenchymal Transition of Human Lung Adenocarcinoma Cells In Vitro and Metastasis of Murine Melanoma In Vivo. Molecules 2020, 25, 5925. [Google Scholar] [CrossRef]
- Shin, S.; Wakabayashi, J.; Yates, M.S.; Wakabayashi, N.; Dolan, P.M.; Aja, S.; Liby, K.T.; Sporn, M.B.; Yamamoto, M.; Kensler, T.W. Role of Nrf2 in prevention of high-fat diet-induced obesity by synthetic triterpenoid CDDO-Imidazolide. Eur. J. Pharmacol. 2009, 620, 138–144. [Google Scholar] [CrossRef]
- Clement, E.; Lazar, I.; Muller, C.; Nieto, L. Obesity and melanoma: Could fat be fueling malignancy? Pigment Cell Melanoma Res. 2017, 30, 294–306. [Google Scholar] [CrossRef]
- Babinska, A.; Kaszubowski, M.; Kmieć, P.; Sworczak, K. Selected adipocytokines in patients with an incidentally discovered pheochromocytoma. Minerva Endocrinol. 2020, 45, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Tas, M.L.; Nagtegaal, M.; Kraal, K.C.J.M.; Tytgat, G.A.M.; Abeling, N.G.G.M.; Koster, J.; Pluijm, S.M.F.; Zwaan, C.M.; de Keizer, B.; Molenaar, J.J.; et al. Neuroblastoma stage 4S: Tumor regression rate and risk factors of progressive disease. Pediatr. Blood Cancer 2020, 67, e28061. [Google Scholar] [CrossRef] [PubMed]
- Trottier, M.D.; Naaz, A.; Li, Y.; Fraker, P.J. Enhancement of hematopoiesis and lymphopoiesis in diet-induced obese mice. Proc. Natl. Acad. Sci. USA 2012, 109, 7622–7629. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Jeon, Y.E.; Jung, J.I.; Kim, S.M.; Hong, S.H.; Lee, J.; Hwang, J.S.; Hwang, M.O.; Kwon, K.; Kim, E.J. Anti-obesity effect of Cydonia oblonga Miller extract in high-fat diet-induced obese C57BL/6 mice. J. Funct. Foods 2022, 89, 104945. [Google Scholar] [CrossRef]
- De Melo, T.S.; Lima, P.R.; Carvalho, K.M.M.B.; Fontenele, T.M.; Solon, F.R.N.; Tomé, A.R.; de Lemos, T.L.G.; da Cruz Fonseca, S.G.; Santos, F.A.; Rao, V.S.; et al. Ferulic acid lowers body weight and visceral fat accumulation via modulation of enzymatic, hormonal and inflammatory changes in a mouse model of high-fat diet-induced obesity. Braz. J. Med. Biol. Res. 2017, 50, e5630. [Google Scholar] [CrossRef]
- Russo, V.C.; Metaxas, S.; Kobayashi, K.; Harris, M.; Werther, G.A. Antiapoptotic Effects of Leptin in Human Neuroblastoma Cells. Endocrinology 2004, 145, 4103–4112. [Google Scholar] [CrossRef]
- Paddillaya, N.; Ingale, K.; Gaikwad, C.; Saini, D.K.; Pullarkat, P.; Kondaiah, P.; Menon, G.I.; Gundiah, N. Cell adhesion strength and tractions are mechano-diagnostic features of cellular invasiveness. Soft Matter 2022, 18, 4378–4388. [Google Scholar] [CrossRef]
- Ostrowska-Podhorodecka, Z.; McCulloch, C.A. Vimentin regulates the assembly and function of matrix adhesions. Wound Repair Regen. 2021, 29, 602–612. [Google Scholar] [CrossRef]
- Akrida, I.; Papadaki, H. Adipokines and epithelial-mesenchymal transition (EMT) in cancer. Mol. Cell. Biochem. 2023. [Google Scholar] [CrossRef]
- Olea-Flores, M.; Juárez-Cruz, J.C.; Zuñiga-Eulogio, M.D.; Acosta, E.; García-Rodríguez, E.; Zacapala-Gomez, A.E.; Mendoza-Catalán, M.A.; Ortiz-Ortiz, J.; Ortuño-Pineda, C.; Navarro-Tito, N. New Actors Driving the Epithelial–Mesenchymal Transition in Cancer: The Role of Leptin. Biomolecules 2020, 10, 1676. [Google Scholar] [CrossRef]
- Markov, A.V.; Zenkova, M.A.; Logashenko, E.B. Modulation of Tumour-Related Signaling Pathways by Natural Pentacyclic Triterpenoids and their Semisynthetic Derivatives. Curr. Med. Chem. 2017, 24, 1277–1320. [Google Scholar] [CrossRef] [PubMed]
- Burns, M.C.; Sun, Q.; Daniels, R.N.; Camper, D.; Kennedy, J.P.; Phan, J.; Olejniczak, E.T.; Lee, T.; Waterson, A.G.; Rossanese, O.W.; et al. Approach for targeting Ras with small molecules that activate SOS-mediated nucleotide exchange. Proc. Natl. Acad. Sci. USA 2014, 111, 3401–3406. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R.J. MEK1/2 dual-specificity protein kinases: Structure and regulation. Biochem. Biophys. Res. Commun. 2012, 417, 5–10. [Google Scholar] [CrossRef]
- Khan, Z.M.; Real, A.M.; Marsiglia, W.M.; Chow, A.; Duffy, M.E.; Yerabolu, J.R.; Scopton, A.P.; Dar, A.C. Structural basis for the action of the drug trametinib at KSR-bound MEK. Nature 2020, 588, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R. ERK1/2 MAP kinases: Structure, function, and regulation. Pharmacol. Res. 2012, 66, 105–143. [Google Scholar] [CrossRef]
- Pan, X.; Pei, J.; Wang, A.; Shuai, W.; Feng, L.; Bu, F.; Zhu, Y.; Zhang, L.; Wang, G.; Ouyang, L. Development of small molecule extracellular signal-regulated kinases (ERKs) inhibitors for cancer therapy. Acta Pharm. Sin. B 2022, 12, 2171–2192. [Google Scholar] [CrossRef]
- Gelsomino, L.; Giordano, C.; La Camera, G.; Sisci, D.; Marsico, S.; Campana, A.; Tarallo, R.; Rinaldi, A.; Fuqua, S.; Leggio, A.; et al. Leptin Signaling Contributes to Aromatase Inhibitor Resistant Breast Cancer Cell Growth and Activation of Macrophages. Biomolecules 2020, 10, 543. [Google Scholar] [CrossRef]
- Li, F.; Zhao, S.; Guo, T.; Li, J.; Gu, C. The Nutritional Cytokine Leptin Promotes NSCLC by Activating the PI3K/AKT and MAPK/ERK Pathways in NSCLC Cells in a Paracrine Manner. Biomed Res. Int. 2019, 2019, 2585743. [Google Scholar] [CrossRef]
- Ellerhorst, A.J.; Diwan, H.A.; Dang, M.S.; Uffort, G.D.; Johnson, K.M.; Cooke, P.C.; Grimm, A.E. Promotion of melanoma growth by the metabolic hormone leptin. Oncol. Rep. 2010, 23, 901–907. [Google Scholar] [CrossRef]
- Ara, T.; DeClerck, Y.A. Mechanisms of invasion and metastasis in human neuroblastoma. Cancer Metastasis Rev. 2006, 25, 645–657. [Google Scholar] [CrossRef]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial–mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Gautier, M.; Thirant, C.; Delattre, O.; Janoueix-Lerosey, I. Plasticity in Neuroblastoma Cell Identity Defines a Noradrenergic-to-Mesenchymal Transition (NMT). Cancers 2021, 13, 2904. [Google Scholar] [CrossRef] [PubMed]
- Semina, E.V.; Rubina, K.A.; Shmakova, A.A.; Rysenkova, K.D.; Klimovich, P.S.; Aleksanrushkina, N.A.; Sysoeva, V.Y.; Karagyaur, M.N.; Tkachuk, V.A. Downregulation of uPAR promotes urokinase translocation into the nucleus and epithelial to mesenchymal transition in neuroblastoma. J. Cell. Physiol. 2020, 235, 6268–6286. [Google Scholar] [CrossRef] [PubMed]
- Szemes, M.; Greenhough, A.; Melegh, Z.; Malik, S.; Yuksel, A.; Catchpoole, D.; Gallacher, K.; Kollareddy, M.; Park, J.H.; Malik, K. Wnt Signalling Drives Context-Dependent Differentiation or Proliferation in Neuroblastoma. Neoplasia 2018, 20, 335–350. [Google Scholar] [CrossRef]
- Salomatina, O.V.V.; Sen’kova, A.V.V.; Moralev, A.D.D.; Savin, I.A.A.; Komarova, N.I.I.; Salakhutdinov, N.F.F.; Zenkova, M.A.A.; Markov, A.V.V. Novel Epoxides of Soloxolone Methyl: An Effect of the Formation of Oxirane Ring and Stereoisomerism on Cytotoxic Profile, Anti-Metastatic and Anti-Inflammatory Activities In Vitro and In Vivo. Int. J. Mol. Sci. 2022, 23, 6214. [Google Scholar] [CrossRef]
- Sen’kova, A.V.; Savin, I.A.; Odarenko, K.V.; Salomatina, O.V.; Salakhutdinov, N.F.; Zenkova, M.A.; Markov, A.V. Protective effect of soloxolone derivatives in carrageenan- and LPS-driven acute inflammation: Pharmacological profiling and their effects on key inflammation-related processes. Biomed. Pharmacother. 2023, 159, 114231. [Google Scholar] [CrossRef]
- Markov, A.V.; Odarenko, K.V.; Sen’kova, A.V.; Ilyina, A.A.; Zenkova, M.A. Evaluation of the Antitumor Potential of Soloxolone Tryptamide against Glioblastoma Multiforme Using in silico, in vitro, and in vivo Approaches. Biochem. 2023, 88, 1008–1021. [Google Scholar] [CrossRef]
- Smart, J.A.; Oleksak, J.E.; Hartsough, E.J. Cell Adhesion Molecules in Plasticity and Metastasis. Mol. Cancer Res. 2021, 19, 25–37. [Google Scholar] [CrossRef]
- Liu, F.; Wu, Q.; Dong, Z.; Liu, K. Integrins in cancer: Emerging mechanisms and therapeutic opportunities. Pharmacol. Ther. 2023, 247, 108458. [Google Scholar] [CrossRef]
- Rouaud, F.; Sluysmans, S.; Flinois, A.; Shah, J.; Vasileva, E.; Citi, S. Scaffolding proteins of vertebrate apical junctions: Structure, functions and biophysics. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183399. [Google Scholar] [CrossRef]
- Kinoshita, N.; Yamamoto, T.S.; Yasue, N.; Takagi, C.; Fujimori, T.; Ueno, N. Force-dependent remodeling of cytoplasmic ZO-1 condensates contributes to cell-cell adhesion through enhancing tight junctions. iScience 2022, 25, 103846. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, A.; Hashemy, S.I.; Aghaei, M.; Panjehpour, M. Leptin induces matrix metalloproteinase 7 expression to promote ovarian cancer cell invasion by activating ERK and JNK pathways. J. Cell. Biochem. 2018, 119, 2333–2344. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Cordova, C.; Gutiérrez, B.; Hernández, M.; Nieto, M.L. A dangerous liaison: Leptin and sPLA2-IIA join forces to induce proliferation and migration of astrocytoma cells. PLoS ONE 2017, 12, e0170675. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.Y.; Chen, H.; Li, D.D.; Li, A.L.; Wang, W.Y.; Gu, W. Design, synthesis, and anticancer evaluation of novel quinoline derivatives of ursolic acid with hydrazide, oxadiazole, and thiadiazole moieties as potent MEK inhibitors. J. Enzyme Inhib. Med. Chem. 2019, 34, 955–972. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Liu, C.; Zhang, Y.; Qiu, X.; Zhang, L.; Zhao, H.; Rong, Y.; Sun, Y. Ilexgenin A, a novel pentacyclic triterpenoid extracted from Aquifoliaceae shows reduction of LPS-induced peritonitis in mice. Eur. J. Pharmacol. 2017, 797, 94–105. [Google Scholar] [CrossRef]
- Takahashi, Y.; Okimura, Y.; Mizuno, I.; Iida, K.; Takahashi, T.; Kaji, H.; Abe, H.; Chihara, K. Leptin Induces Mitogen-activated Protein Kinase- dependent Proliferation of C3H10T1/2 Cells. J. Biol. Chem. 1997, 272, 12897–12900. [Google Scholar] [CrossRef]
- Ramaswamy, P.; Nanjaiah, N.D.; Borkotokey, M. Role of MEK-ERK signaling mediated adhesion of glioma cells to extracellular matrix: Possible implication on migration and proliferation. Ann. Neurosci. 2019, 26, 52–56. [Google Scholar] [CrossRef]
- Langlois, B.; Belozertseva, E.; Parlakian, A.; Bourhim, M.; Gao-Li, J.; Blanc, J.; Tian, L.; Coletti, D.; Labat, C.; Ramdame-Cherif, Z.; et al. Vimentin knockout results in increased expression of sub-endothelial basement membrane components and carotid stiffness in mice. Sci. Rep. 2017, 7, 11628. [Google Scholar] [CrossRef]
- Logashenko, E.B.; Salomatina, O.V.; Markov, A.V.; Korchagina, D.V.; Salakhutdinov, N.F.; Tolstikov, G.A.; Vlassov, V.V.; Zenkova, M.A. Synthesis and Pro-Apoptotic Activity of Novel Glycyrrhetinic Acid Derivatives. ChemBioChem 2011, 12, 784–794. [Google Scholar] [CrossRef]
- Meijering, E.; Jacob, M.; Sarria, J.-C.F.; Steiner, P.; Hirling, H.; Unser, M. Design and validation of a tool for neurite tracing and analysis in fluorescence microscopy images. Cytom. Part A 2004, 58A, 167–176. [Google Scholar] [CrossRef]
- RCSB PDB Protein Data Bank. Available online: https://www.rcsb.org/ (accessed on 15 May 2023).
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Odarenko, K.V.; Salomatina, O.V.; Chernikov, I.V.; Salakhutdinov, N.F.; Zenkova, M.A.; Markov, A.V. Soloxolone Methyl Reduces the Stimulatory Effect of Leptin on the Aggressive Phenotype of Murine Neuro2a Neuroblastoma Cells via the MAPK/ERK1/2 Pathway. Pharmaceuticals 2023, 16, 1369. https://doi.org/10.3390/ph16101369
Odarenko KV, Salomatina OV, Chernikov IV, Salakhutdinov NF, Zenkova MA, Markov AV. Soloxolone Methyl Reduces the Stimulatory Effect of Leptin on the Aggressive Phenotype of Murine Neuro2a Neuroblastoma Cells via the MAPK/ERK1/2 Pathway. Pharmaceuticals. 2023; 16(10):1369. https://doi.org/10.3390/ph16101369
Chicago/Turabian StyleOdarenko, Kirill V., Oksana V. Salomatina, Ivan V. Chernikov, Nariman F. Salakhutdinov, Marina A. Zenkova, and Andrey V. Markov. 2023. "Soloxolone Methyl Reduces the Stimulatory Effect of Leptin on the Aggressive Phenotype of Murine Neuro2a Neuroblastoma Cells via the MAPK/ERK1/2 Pathway" Pharmaceuticals 16, no. 10: 1369. https://doi.org/10.3390/ph16101369
APA StyleOdarenko, K. V., Salomatina, O. V., Chernikov, I. V., Salakhutdinov, N. F., Zenkova, M. A., & Markov, A. V. (2023). Soloxolone Methyl Reduces the Stimulatory Effect of Leptin on the Aggressive Phenotype of Murine Neuro2a Neuroblastoma Cells via the MAPK/ERK1/2 Pathway. Pharmaceuticals, 16(10), 1369. https://doi.org/10.3390/ph16101369