
The Influence of Exercise Intensity on Tryptophan Metabolites in Thoroughbred Horses




Abstract
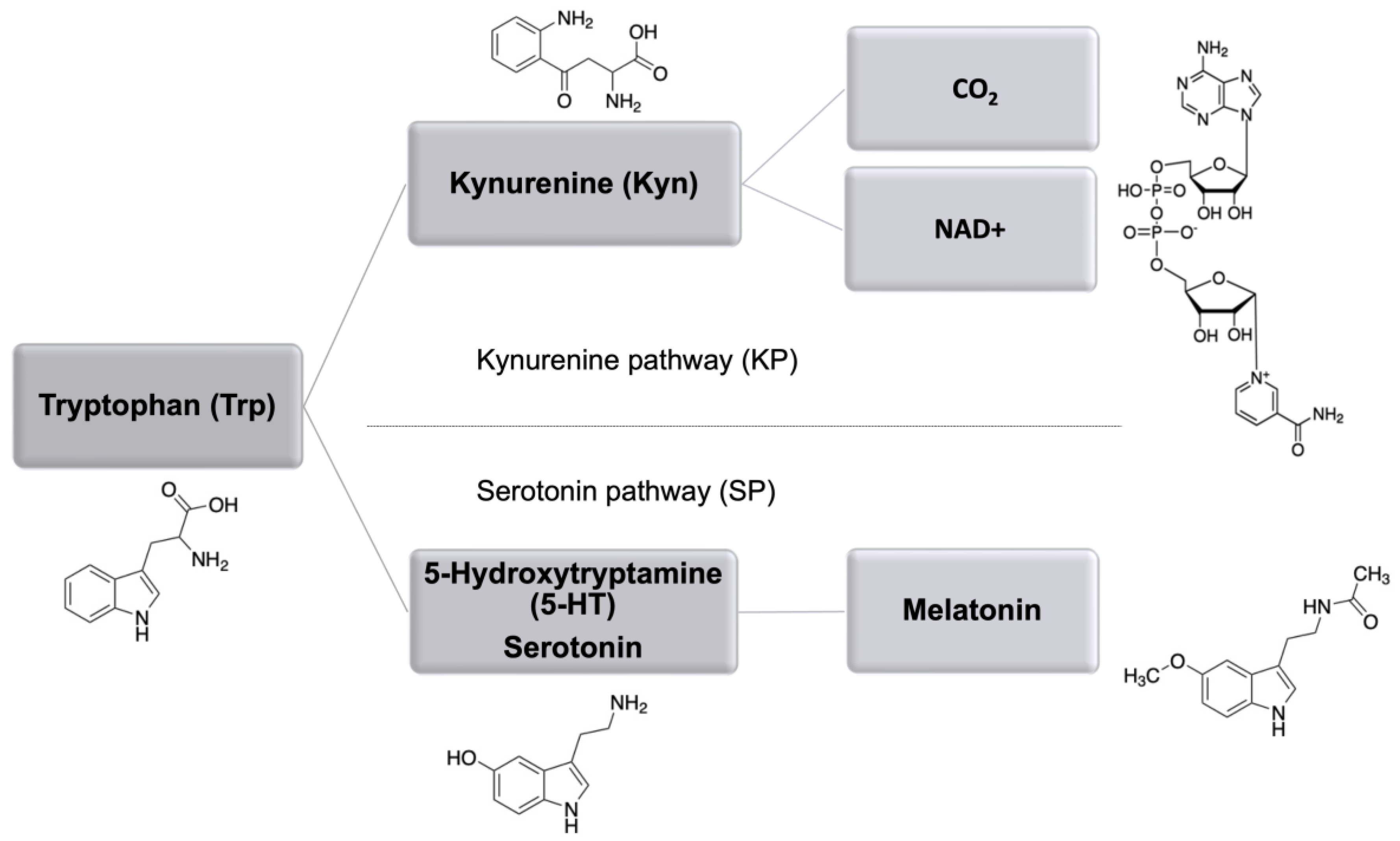
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Horses
4.2. Blood Samples Collection and Analysis
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bender, D.A. Biochemistry of tryptophan in health and disease. Mol. Asp. Med. 1983, 6, 101–197. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A. Kynurenine pathway of tryptophan metabolism: Regulatory and functional aspects. Int. J. Tryptophan Res. 2017, 10, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Song, S.B. Possible adverse effects of high-dose nicotinamide: Mechanisms and safety assessment. Biomolecules 2020, 10, 687. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, K.A.; Ketelhuth, D.F.J. The role of the kynurenine pathway of tryptophan metabolism in cardiovascular disease. Hamostaseologie 2015, 35, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Sas, K.; Robotka, H.; Toldi, J.; Vecsei, L. Mitochondria, metabolic disturbances, oxidative stress and the kynurenine system, with focus on neurodegenerative disorders. J. Neurol. Sci. 2007, 257, 221–239. [Google Scholar] [CrossRef] [PubMed]
- Alberghina, D.; Giannetto, C.; Picione, G. Peripheral serotoninergic response to physical exercise in athletic horses. J. Vet. Sci. 2010, 11, 285–289. [Google Scholar] [CrossRef]
- Valentine-Silva, P.; Raus, J.L. Tryptophan-kynurenine metabolites in exercise and mental health. In Hormones, Metabolism and the Benefits of Exercise. Research and Perspectives in Endocrine Interactions; Springer: Cham, Switzerland, 2017; pp. 83–91. [Google Scholar]
- Arfuso, F.; Assenza, A.; Fazio, F.; Rizzo, M.; Giannetto, C.; Piccione, G. Dynamic change of serum levels of some branched-chain amino acids and tryptophan in athletic horses after different physical exercises. J. Equine Vet. Sci. 2019, 77, 12–16. [Google Scholar] [CrossRef]
- Arfuso, F.; Giannetto, C.; Giudice, E.; Fazio, F.; Panzera, M.; Piccione, G. Peripheral modulators of the central fatigue development and their relationship with athletic performance in jumper horses. Animals 2021, 11, 743. [Google Scholar] [CrossRef]
- Bruschetta, G.; D’Ascola, A.; Medica, P.; Ferlazzo, A.M. Physical exercise affects serotoninergic system in horse leukocytes. J. Equine Vet. Sci. 2020, 88, 102969. [Google Scholar] [CrossRef]
- Medica, P.; Giunta, R.P.; Bruschetta, G.; Ferlazzo, A.M. The influence of training and simulated race on horse plasma. J. Equine Vet. Sci. 2020, 84, 102818. [Google Scholar] [CrossRef] [PubMed]
- Kędzierski, W.; Sadok, I.; Kowalik, S.; Janczarek, I.; Staniszewska, M. Does the type of exercise affect tryptophan catabolism in horses? Animal 2021, 15, 100377. [Google Scholar] [CrossRef] [PubMed]
- Bruschetta, G.; Medica, P.; Fazio, E.; Cravana, C.; Ferlezzo, A.M. The effect of training sessions and feeding regimes on neuromodulator role of serotonin, tryptophan, and β-endorphin of horses. J. Vet. Behav. 2018, 23, 82–86. [Google Scholar] [CrossRef]
- D’Ascola, A.; Bruschetta, G.; Zanghi, G.; Campo, S.; Medica, P.; Campana, S.; Ferlazzo, G.; Gibbs, B.F.; Ferlazzo, A.M. Changes in plasma 5-HT levels and equine leukocyte SERT expression in response to treadmill exercise. Res. Vet. Sci. 2018, 118, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Giannetto, C.; Acri, G.; Giudice, E.; Arfuso, F.; Testagrossa, B.; Piccione, G. Quantifying serum total lipids and tryptophan concentrations by Raman spectroscopy during standardized obstacle course in horses. J. Equine Vet. Sci. 2022, 108, 103820. [Google Scholar] [CrossRef] [PubMed]
- Delarocque, J.; Frers, F.; Feige, K.; Huber, K.; Jung, K.; Warnken, T. Metabolic changes induced by oral glucose tests in horses and their diagnostic use. J. Vet. Intern. Med. 2021, 35, 597–605. [Google Scholar] [CrossRef]
- Noble, G.K.; Brockwell, Y.M.; Munn, K.J.; Harris, P.A.; Davidson, H.P.B.; Li, X.; Zhang, D.; Sillence, M.N. Effects of a commercial dose of L-tryptophan on plasma tryptophan concentrations and behaviour in horses. Equine Vet. J. 2008, 40, 51–56. [Google Scholar] [CrossRef]
- Desmecht, D.; Linden, A.; Amory, H.; Art, T.; Lekeux, P. Relationship of plasma lactate production to cortisol release following competition of different types of sporting events in horses. Vet. Res. Commun. 1996, 20, 371–379. [Google Scholar] [CrossRef]
- Lindner, A.; von Wittke, P.; Schmald, M.; Kusserow, J.; Sommer, H. Maximal lactate concentrations in horses after exercise of rifferent duration and intensity. J. Equine Vet. Sci. 1992, 12, 36–39. [Google Scholar] [CrossRef]
- Bergero, D.; Assenza, A.; Schiavone, A.; Piccione, G.; Perona, G.; Caola, G. Amino acid concentrations in blood serum of horses performing long lasting low-intensity exercise. J. Anim. Physiol. Anim. Nutr. 2005, 89, 146–150. [Google Scholar] [CrossRef]
- Su, C.H.; Chang, H.C.; Hong, C.J. Physical exercise prevents mice from L-kynurenine-induced depression-like behavior. Asian J. Psychiatr. 2020, 48, 101894. [Google Scholar] [CrossRef]
- Knut & Alice Wallenberg Foundation. The Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000117009-KMO (accessed on 21 November 2022).
- Ting, K.K.; Brew, B.J.; Guillemin, G.J. Effect of quinolinic acid on human astrocytes morphology and functions: Implications in Alzheimer’s disease. J. Neuroinflamm. 2009, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Sathyasaikumar, K.V.; Tararina, M.; Wu, H.-Q.; Neale, S.A.; Weisz, F.; Salt, T.E.; Schwarcz, R. Xanthurenic acid formation from 3-Hydroxykynurenine in the mammalian brain: Neurochemical characterization and physiological effects. Neuroscience 2017, 367, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Kędzierski, W.; Strzelec, K.; Cywińska, A.; Kowalik, S. Salivary cortisol concentration in exercised Thoroughbred horses. J. Equine Vet. Sci. 2013, 33, 1106–1109. [Google Scholar] [CrossRef]
- Sadok, I.; Jędruchniewicz, K.; Rawicz-Pruszyński, K.; Staniszewska, M. UHPLC-ESI-MS/MS Quantification of Relevant Substrates and Metabolites of the Kynurenine Pathway Present in Serum and Peritoneal Fluid from Gastric Cancer Patients—Method Development and Validation. Int. J. Mol. Sci. 2021, 22, 6972. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Analyzed Parameters | Low-Intensity Exercise (n = 18) | High-Intensity Exercise (n = 16) | |||||
|---|---|---|---|---|---|---|---|
| Sample A | Sample B | Sample C | Sample A | Sample B | Sample C | p-Value | |
| Lactic acid [mmol/L] | 0.92 ± 0.04 | 1.38 ± 0.08 * | 0.81 ± 0.03 | 0.87 ± 0.04 a | 16.2 ± 0.83 b | 2.32 ± 0.06 c | 0.0007 |
| Other analytes [µmol/L]: | |||||||
| Tryptophan | 118 ± 6.17 | 110 ± 5.28 | 116 ± 5.74 | 126 ± 3.24 a | 106 ± 4.98 b | 132 ± 5.36 a | 0.0010 |
| Kynurenine | 1.39 ± 0.10 | 1.48 ± 0.12 | 1.44 ± 0.11 | 1.47 ± 0.10 a | 1.85 ± 0.16 b | 1.64 ± 0.11 ab | 0.0212 |
| Kynurenic acid | 0.22 ± 0.03 | 0.22 ± 0.04 | 0.22 ± 0.03 | 0.20 ± 0.03 | 0.18 ± 0.03 | 0.19 ± 0.03 | >0.05 |
| 3-Hydroxykynurenine | 0.70 ± 0.04 | 0.62 ± 0.03 | 0.65 ± 0.04 | 0.65 ± 0.07 | 0.68 ± 0.06 | 0.74 ± 0.07 | >0.05 |
| Xanthurenic acid | 0.12 ± 0.01 | 0.13 ± 0.01 | 0.13 ± 0.01 | 0.13 ± 0.01 a | 0.13 ± 0.01 a | 0.17 ± 0.01 b | 0.0007 |
| Nicotinamide | 3.04 ± 0.27 | 2.81 ± 0.27 | 2.92 ± 0.32 | 2.80 ± 0.38 a | 3.97 ± 0.53 b | 3.71 ± 0.43 ab | 0.0371 |
| Serotonin | 0.21 ± 0.03 | 0.21 ± 0.03 | 0.24 ± 0.03 | 0.18 ± 0.02 | 0.21 ± 0.02 | 0.23 ± 0.04 | >0.05 |
| Analytes: | B-A | C-A |
|---|---|---|
| LA (B-A) and: Tryptophan | −0.295 | 0.21 |
| Kynurenine | 0.24 | 0.12 |
| Kynurenic acid | −0.11 | 0.07 |
| 3-Hydroxykynurenine | 0.33 | 0.42 * |
| Xanthurenic acid | 0.02 | 0.63 * |
| Nicotinamide | 0.56 * | 0.61 * |
| Serotonin | 0.19 | 0.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staniszewska, M.; Kowalik, S.; Sadok, I.; Kędzierski, W. The Influence of Exercise Intensity on Tryptophan Metabolites in Thoroughbred Horses. Pharmaceuticals 2023, 16, 107. https://doi.org/10.3390/ph16010107
Staniszewska M, Kowalik S, Sadok I, Kędzierski W. The Influence of Exercise Intensity on Tryptophan Metabolites in Thoroughbred Horses. Pharmaceuticals. 2023; 16(1):107. https://doi.org/10.3390/ph16010107
Chicago/Turabian StyleStaniszewska, Magdalena, Sylwester Kowalik, Ilona Sadok, and Witold Kędzierski. 2023. "The Influence of Exercise Intensity on Tryptophan Metabolites in Thoroughbred Horses" Pharmaceuticals 16, no. 1: 107. https://doi.org/10.3390/ph16010107
APA StyleStaniszewska, M., Kowalik, S., Sadok, I., & Kędzierski, W. (2023). The Influence of Exercise Intensity on Tryptophan Metabolites in Thoroughbred Horses. Pharmaceuticals, 16(1), 107. https://doi.org/10.3390/ph16010107