
Polyphenols as Inhibitors of Antibiotic Resistant Bacteria—Mechanisms Underlying Rutin Interference with Bacterial Virulence

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Polyphenols Are Able to Inhibit Growth of Antibiotic Resistant Bacteria
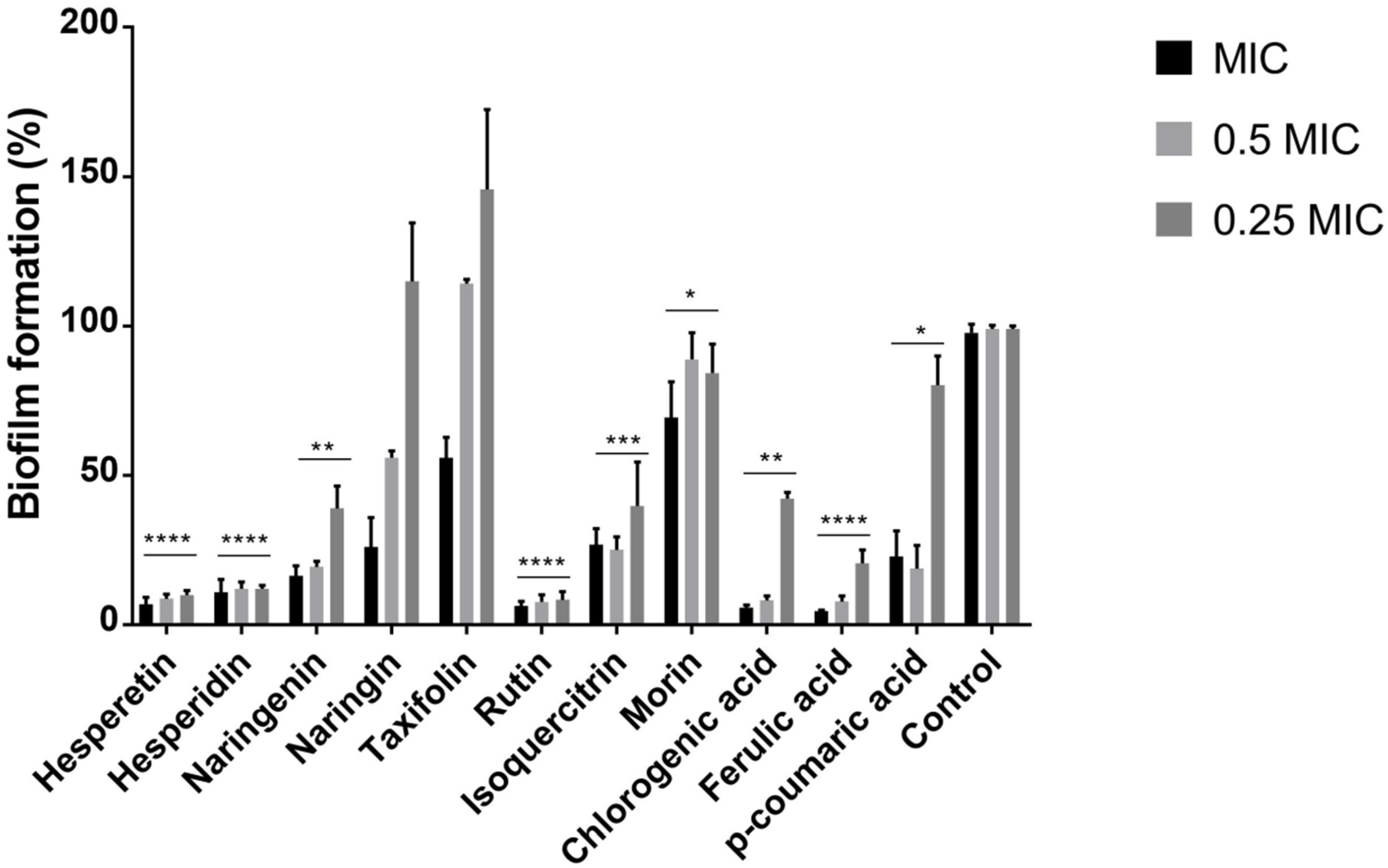
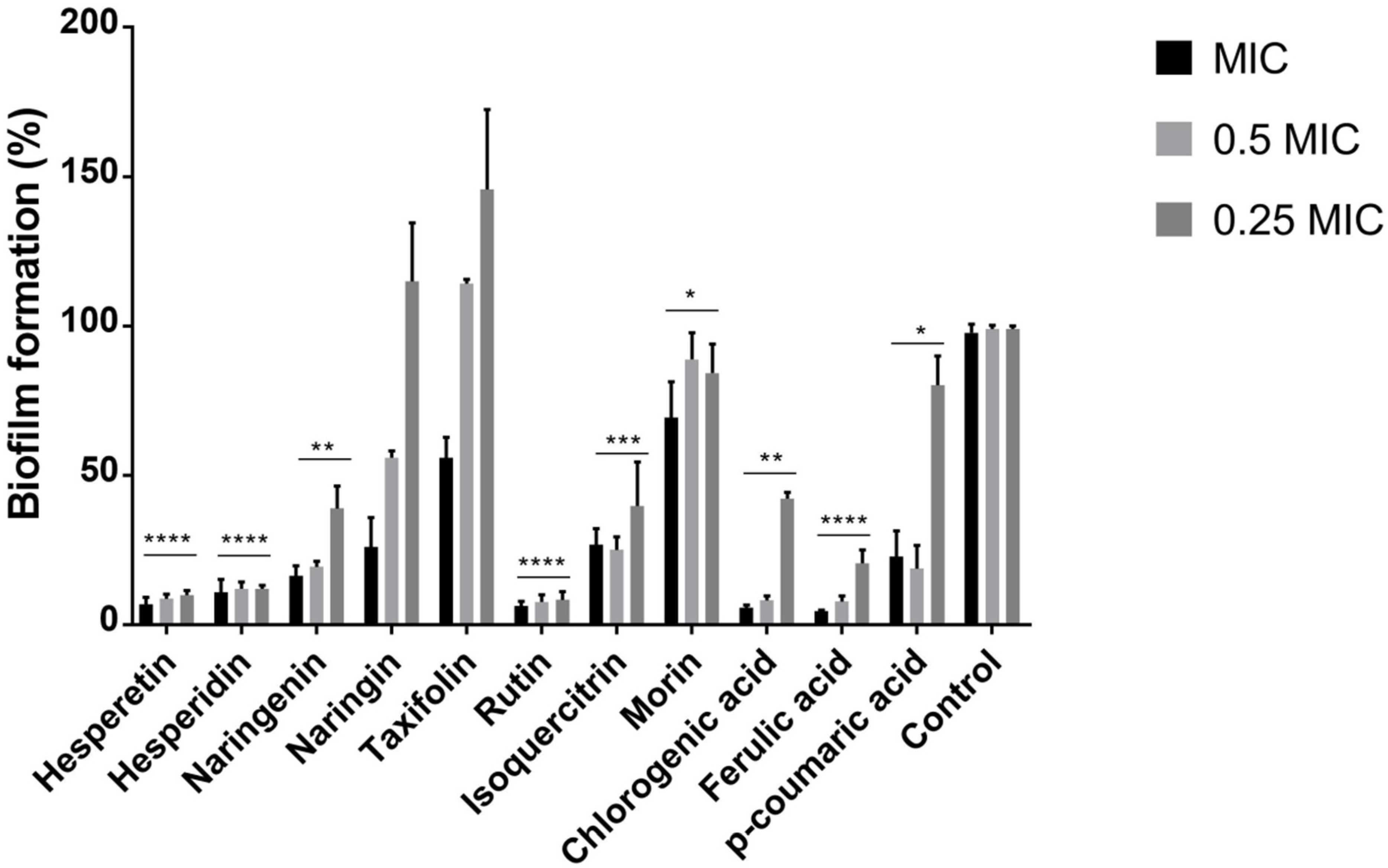
2.2. Polyphenols Reduce the Ability of P. aeruginosa IBRS P001 to Form Biofilms
2.3. Polyphenols Are Mainly Non-Cytotoxic towards HaCaT Cells
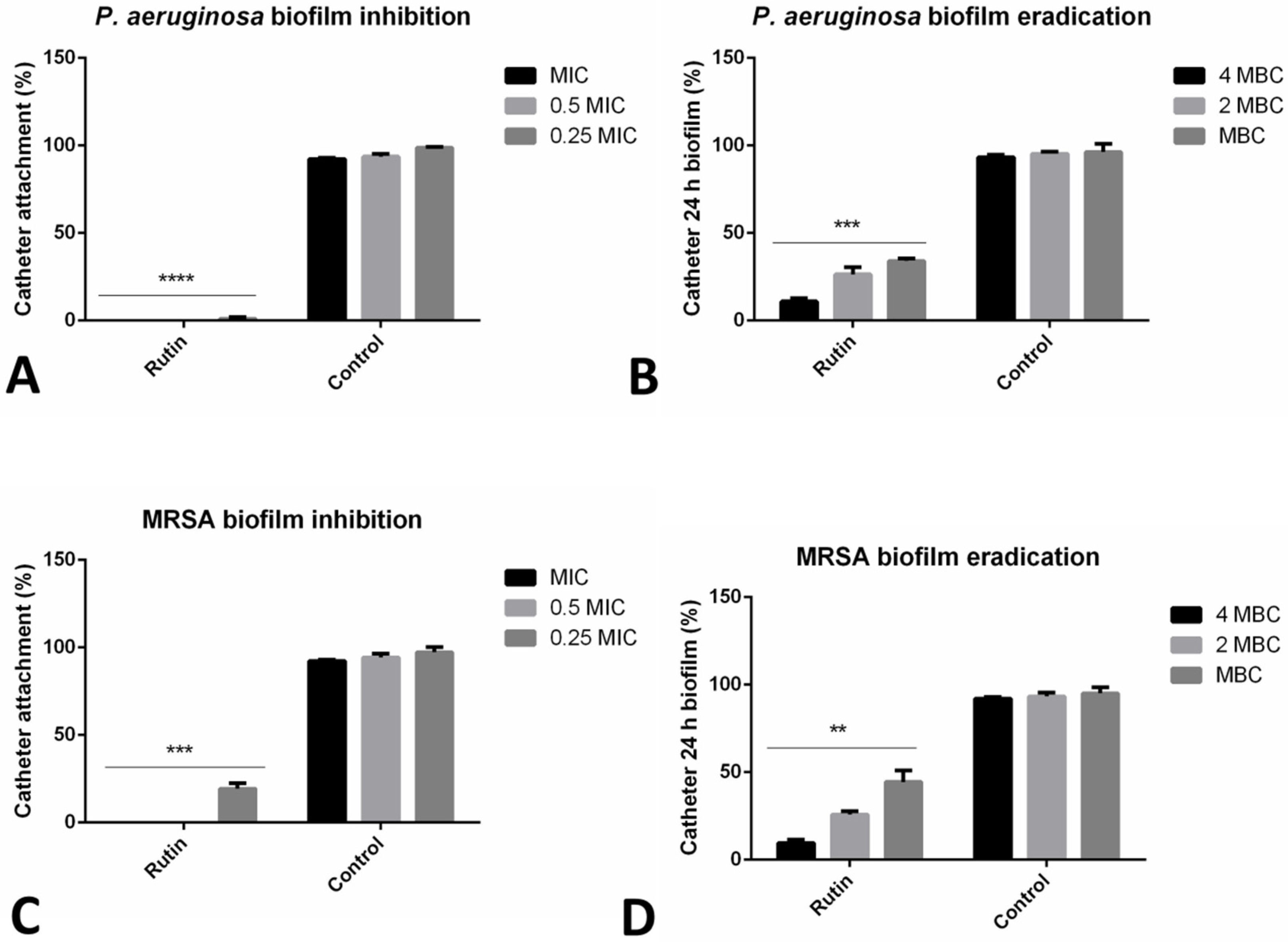
2.4. Rutin Significantly Affected Urinary Catheter Biofilms
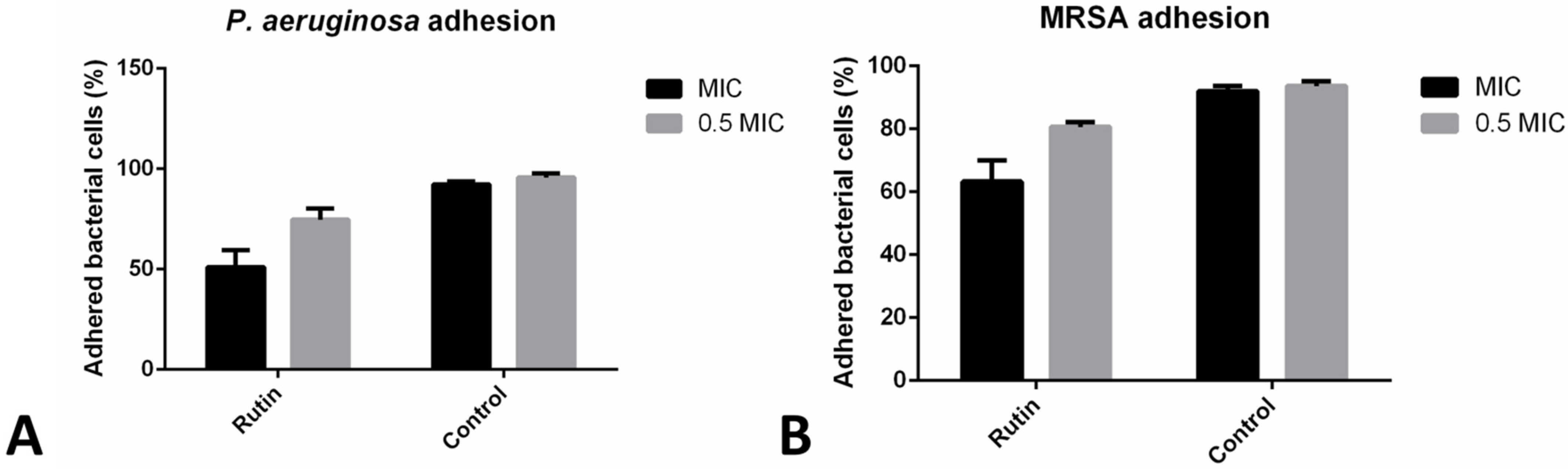
2.5. Co-Incubation with Rutin Reduced Bacterial Adhesion to HaCaT
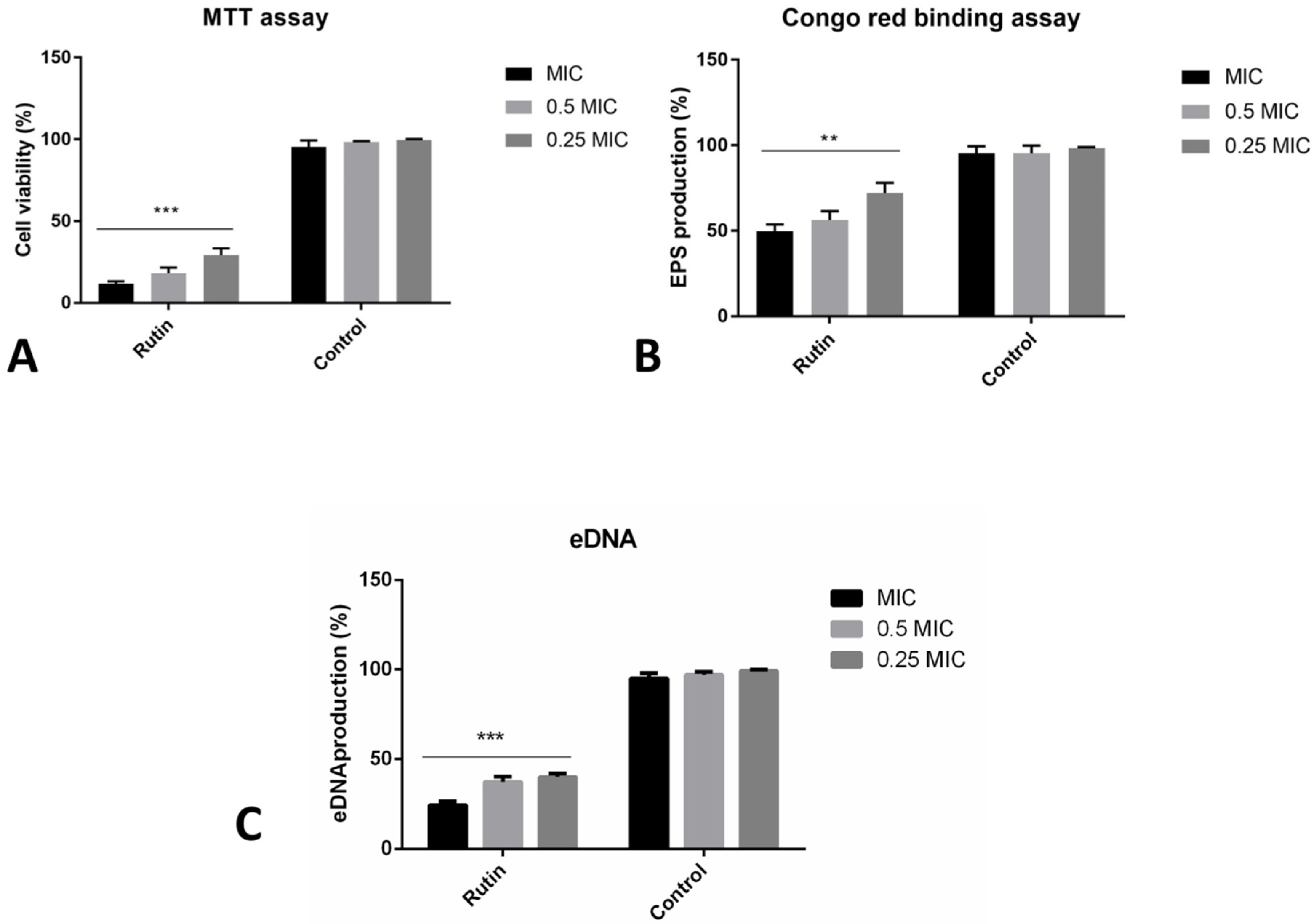
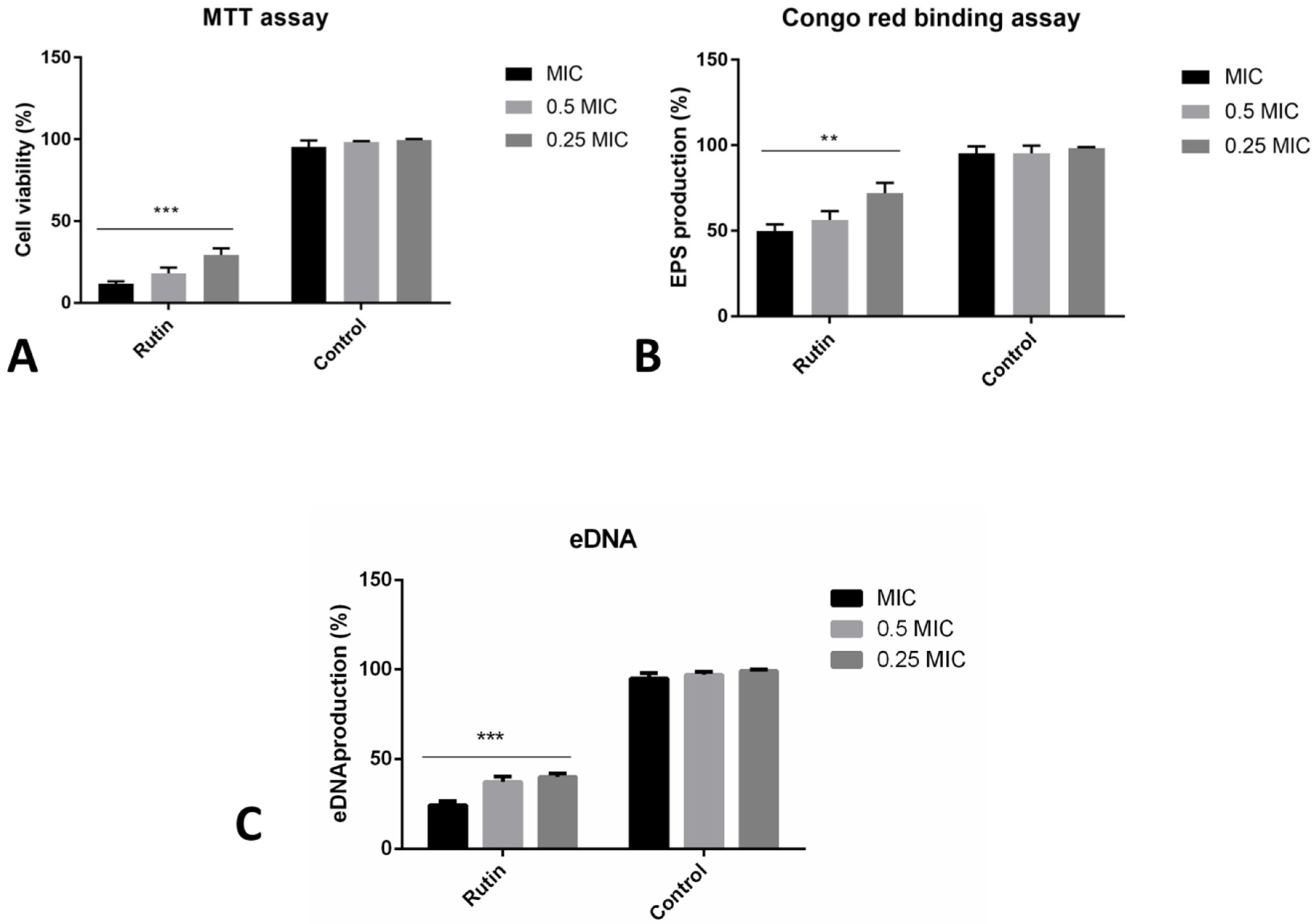
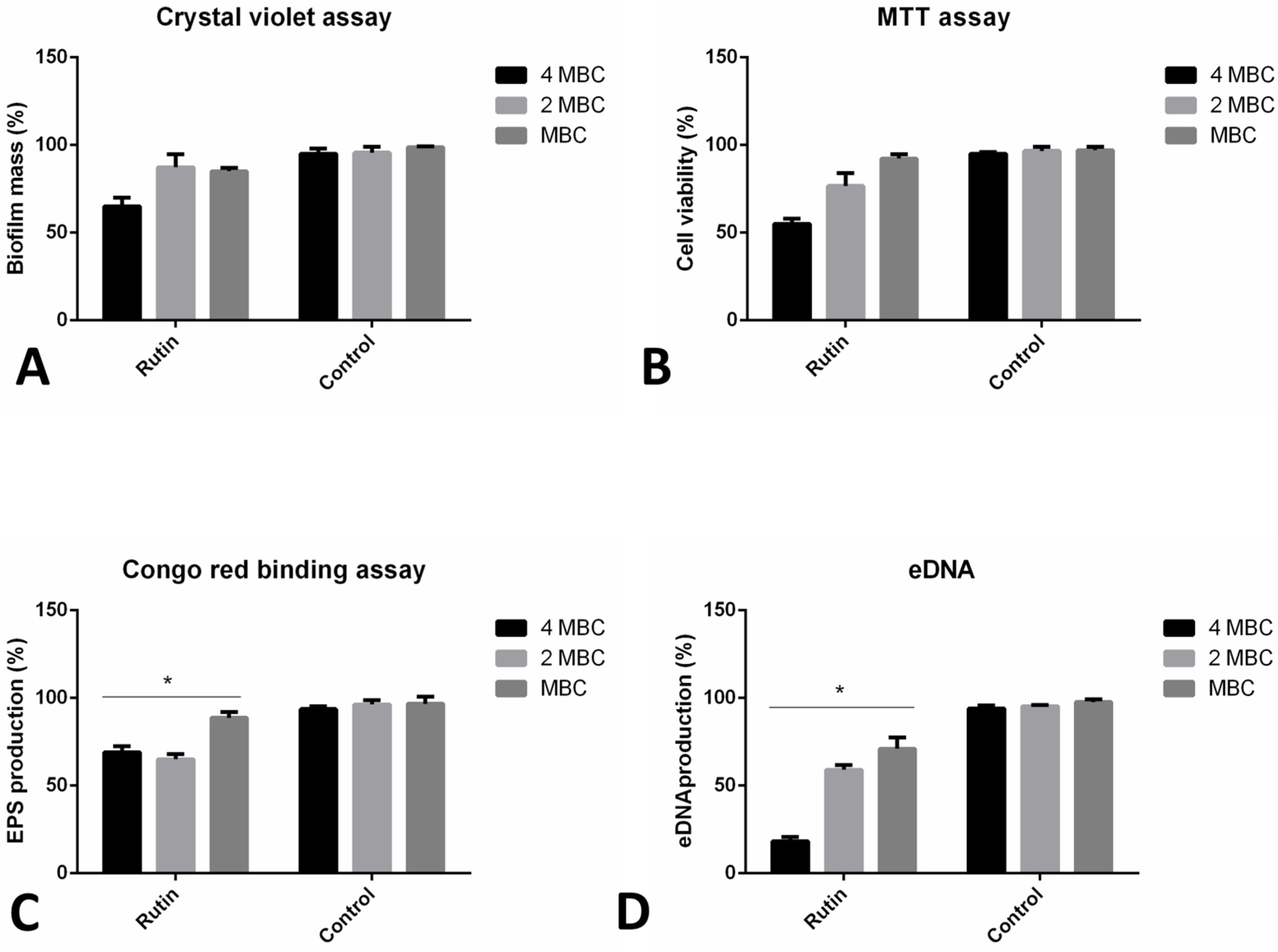
2.6. Co-Incubation with Rutin Affects Different Aspects of Biofilm Formation
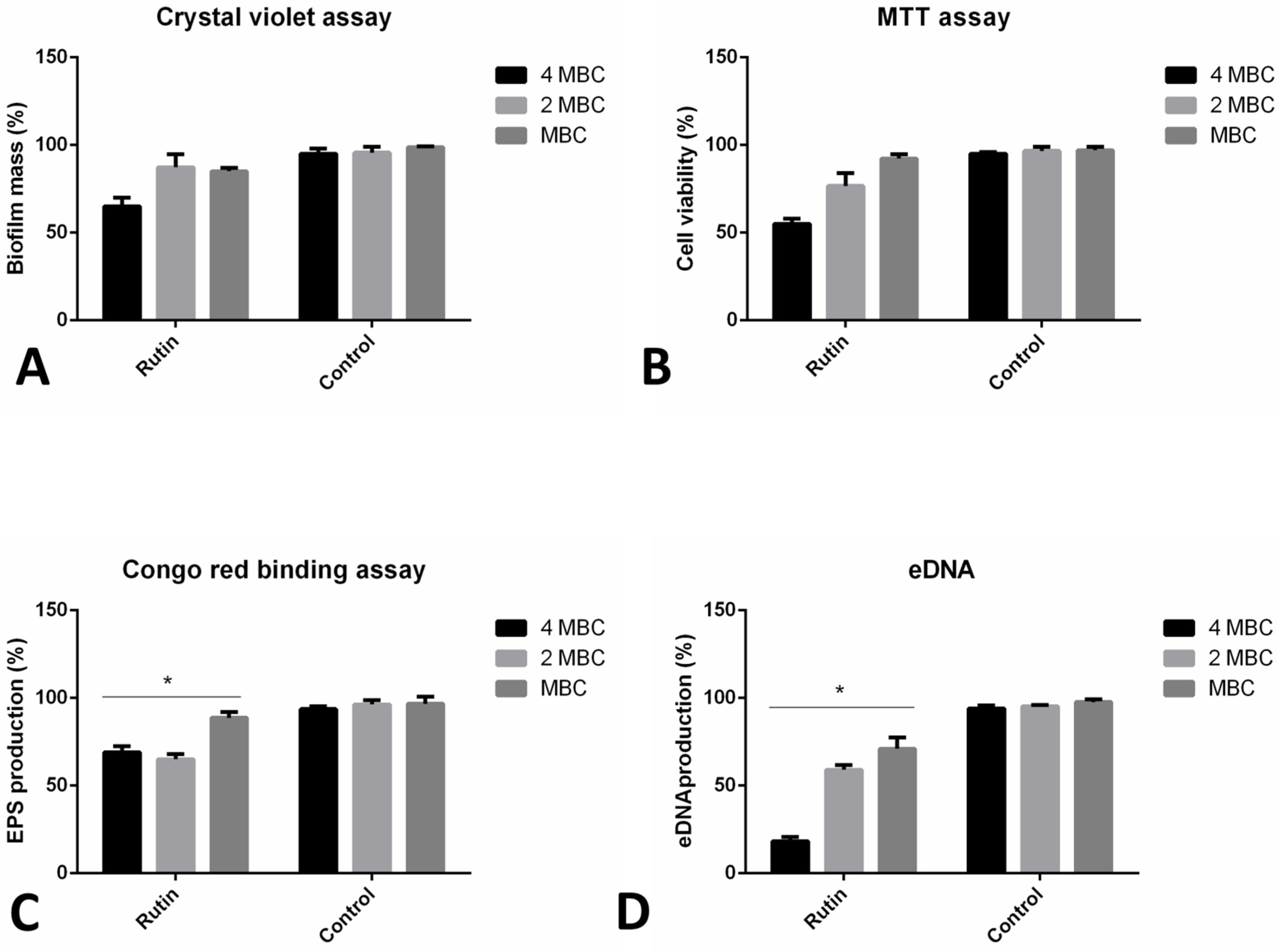
2.7. Rutin Can Eradicate Pre-Formed Biofilms by Diverse Mechanisms
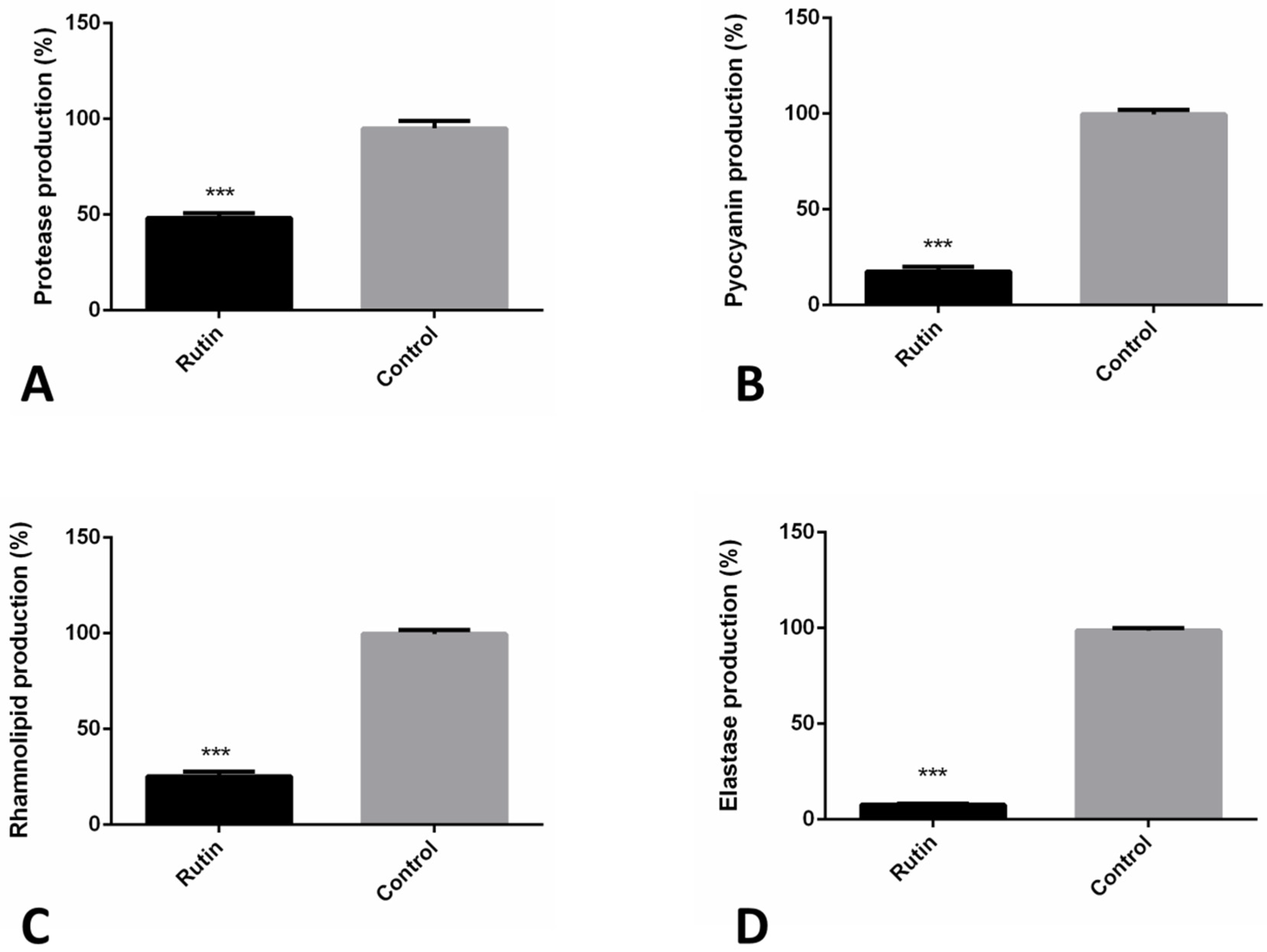
2.8. Rutin Suppresses P. aeruginosa IBRS P001 Virulence Factors Production
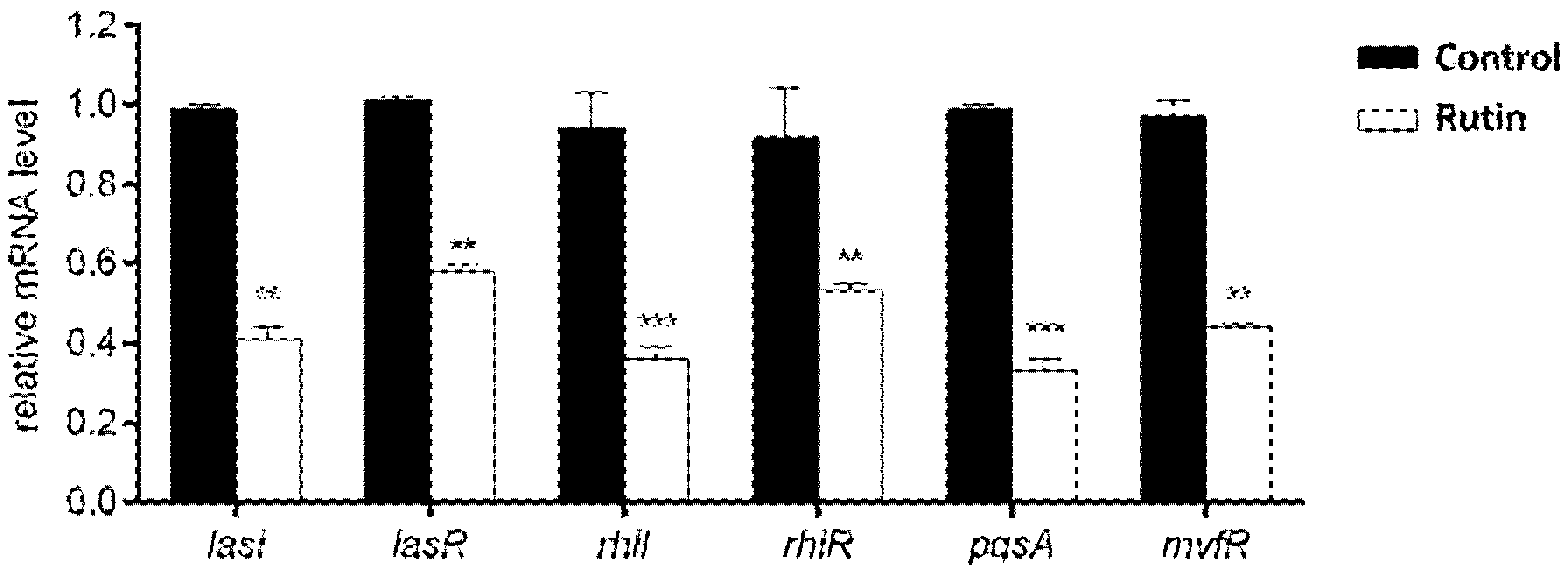
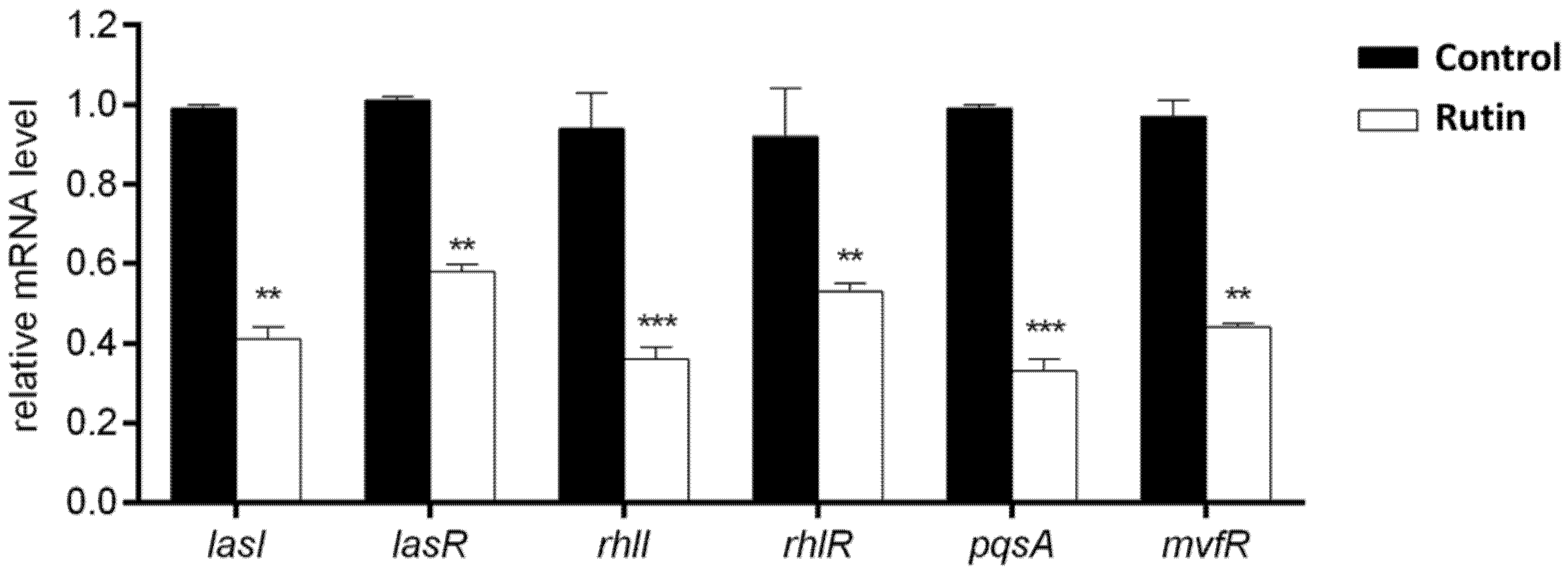
2.9. Rutin Suppresses the P. aeruginosa IBRS P001 QS Systems Expression
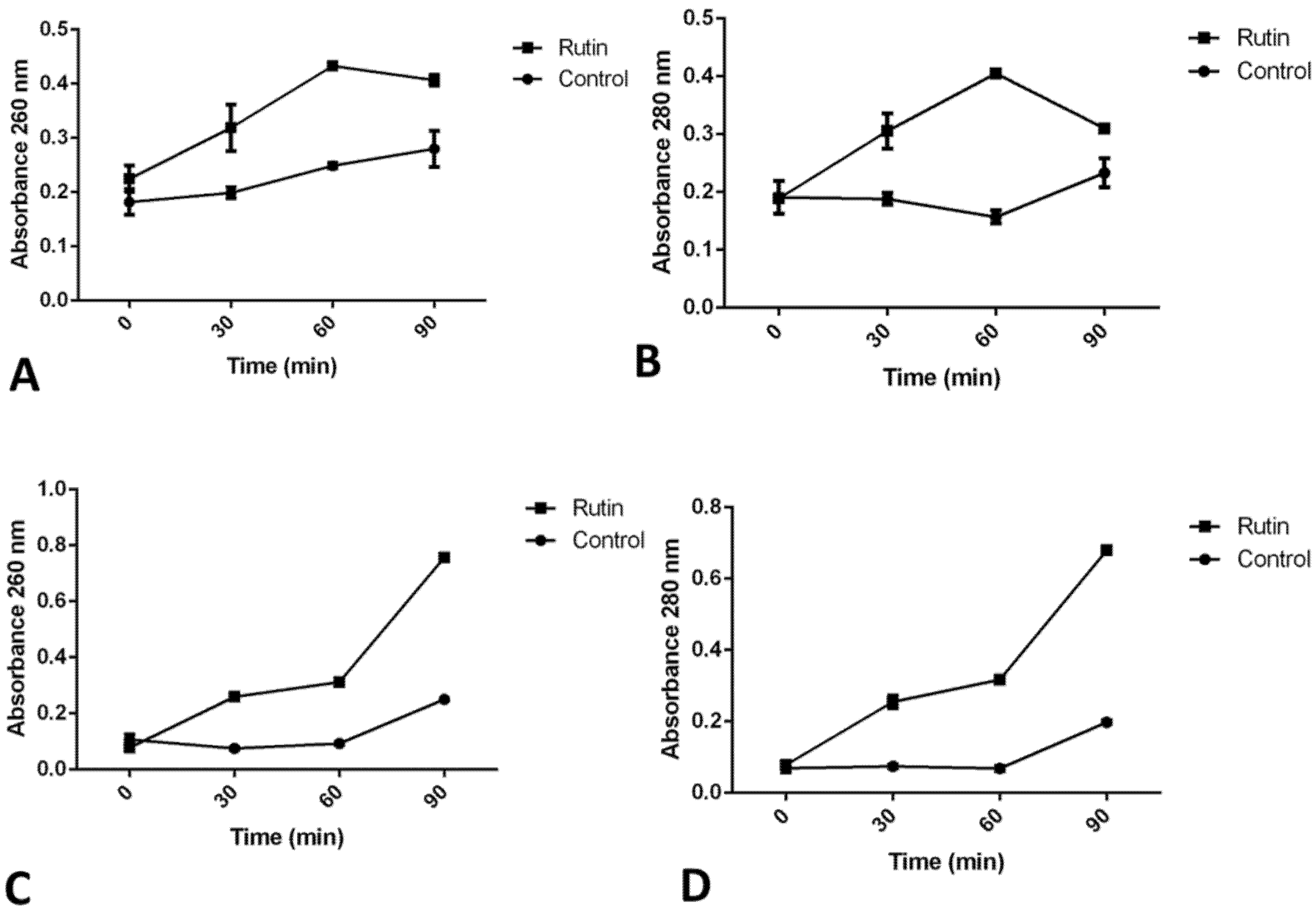
2.10. Rutin’s Impact on Cell Membrane Permeability
3. Discussion
4. Materials and Methods
4.1. Antimicrobial Assay
4.2. Crystal Violet Antibiofilm Assay
4.3. Evaluation of Cytotoxicity in HaCaT Cell Line
4.4. Catheter Biofilm Inhibition/Eradication Model
4.5. P. aeruginosa and MRSA Adhesion to HaCaT Cells
4.6. Mechanisms of Rutin Biofilm Inhibition/Eradication
4.6.1. Biofilm MTT Assay
4.6.2. Congo Red Binding Assay
4.6.3. Quantification of eDNA
4.6.4. Rutin Biofilm Eradication Mechanisms
4.7. Extracellular Virulence Factors Analysis
4.7.1. Protease Assay
4.7.2. Elastase Assay
4.7.3. Pyocyanin Assay
4.7.4. Rhamnolipid Assay
4.8. RT-qPCR Analysis
4.9. Membrane Permeability Assay
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hay, S.I.; Rao, P.C.; Dolecek, C.; Day, N.P.J.; Stergachis, A.; Lopez, A.D.; Murray, C.J.L. Measuring and mapping the global burden of antimicrobial resistance. BMC Med. 2018, 16, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezasoltani, S.; Yadegar, A.; Hatami, B.; Asadzadeh Aghdaei, H.; Zali, M.R. Antimicrobial Resistance as a Hidden Menace Lurking Behind the COVID-19 Outbreak: The Global Impacts of Too Much Hygiene on AMR. Front. Microbiol. 2020, 11, 3097. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.; Larssen, K.W.; Gran, F.W.; Enger, H.; Hæggman, S.; Mäkitalo, B.; Haraldsson, G.; Lindholm, L.; Vuopio, J.; Henius, A.E.; et al. Increasing Incidences and Clonal Diversity of Methicillin-Resistant Staphylococcus aureus in the Nordic Countries-Results From the Nordic MRSA Surveillance. Front. Microbiol. 2021, 12, 1034. [Google Scholar] [CrossRef] [PubMed]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and Treatment of Multidrug-Resistant and Extensively Drug-Resistant Pseudomonas aeruginosa Infections. Clin. Microbiol. Rev. 2019, 32, e00031-19. [Google Scholar] [CrossRef]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2017, 16, 91–102. [Google Scholar] [CrossRef]
- Yan, J.; Bassler, B.L. Surviving as a Community: Antibiotic Tolerance and Persistence in Bacterial Biofilms. Cell Host Microbe 2019, 26, 15–21. [Google Scholar] [CrossRef]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef]
- Uruén, C.; Chopo-Escuin, G.; Tommassen, J.; Mainar-Jaime, R.C.; Arenas, J. Biofilms as Promoters of Bacterial Antibiotic Resistance and Tolerance. Antibiotics 2020, 10, 3. [Google Scholar] [CrossRef]
- Pinto, R.M.; Soares, F.A.; Reis, S.; Nunes, C.; Van Dijck, P. Innovative Strategies Toward the Disassembly of the EPS Matrix in Bacterial Biofilms. Front. Microbiol. 2020, 11, 952. [Google Scholar] [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- DIckey, S.W.; Cheung, G.Y.C.; Otto, M. Different drugs for bad bugs: Antivirulence strategies in the age of antibiotic resistance. Nat. Rev. Drug Discov. 2017, 16, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Martínez, O.F.; Cardoso, M.H.; Ribeiro, S.M.; Franco, O.L. Recent Advances in Anti-virulence Therapeutic Strategies With a Focus on Dismantling Bacterial Membrane Microdomains, Toxin Neutralization, Quorum-Sensing Interference and Biofilm Inhibition. Front. Cell. Infect. Microbiol. 2019, 9, 74. [Google Scholar] [CrossRef] [PubMed]
- Imperi, F.; Fiscarelli, E.V.; Visaggio, D.; Leoni, L.; Visca, P. Activity and impact on resistance development of two antivirulence fluoropyrimidine drugs in Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2019, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Thielen, M.K.; Vaneerd, C.K.; Goswami, M.; Carlson, E.E.; May, J.F. 2-Aminobenzothiazoles Inhibit Virulence Gene Expression and Block Polymyxin Resistance in Salmonella enterica. ChemBioChem 2020, 21, 3500. [Google Scholar] [CrossRef]
- Boibessot, T.; Zschiedrich, C.P.; Lebeau, A.; Bénimèlis, D.; Dunyach-Rémy, C.; Lavigne, J.P.; Szurmant, H.; Benfodda, Z.; Meffre, P. The rational design, synthesis and antimicrobial properties of thiophene derivatives that inhibit bacterial histidine kinases. J. Med. Chem. 2016, 59, 8830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmour, R.; Foster, J.E.; Sheng, Q.; McClain, J.R.; Riley, A.; Sun, P.M.; Ng, W.L.; Yan, D.; Nicas, T.I.; Henry, K.; et al. New Class of Competitive Inhibitor of Bacterial Histidine Kinases. J. Bacteriol. 2005, 187, 8196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carabajal, M.A.; Asquith, C.R.M.; Laitinen, T.; Tizzard, G.J.; Yim, L.; Rial, A.; Chabalgoity, J.A.; Zuercher, W.J.; Véscovi, E.G. Quinazoline-Based Antivirulence Compounds Selectively Target Salmonella PhoP/PhoQ Signal Transduction System. Antimicrob. Agents Chemother. 2020, 64, e01744-19. [Google Scholar] [CrossRef]
- Guarnieri, M.T.; Zhang, L.; Shen, J.; Zhao, R. The Hsp90 Inhibitor Radicicol Interacts with the ATP-Binding Pocket of Bacterial Sensor Kinase PhoQ. J. Mol. Biol. 2008, 379, 82–93. [Google Scholar] [CrossRef]
- Tijjani, H.; Zangoma, M.H.; Mohammed, Z.S.; Obidola, S.M.; Egbuna, C.; Abdulai, S.I. Polyphenols: Classifications, Biosynthesis and Bioactivities. In Functional Foods and Nutraceuticals; Springer: Berlin, Germany, 2020; pp. 389–414. [Google Scholar] [CrossRef]
- Li, A.-N.; Li, S.; Zhang, Y.-J.; Xu, X.-R.; Chen, Y.-M.; Li, H.-B. Resources and Biological Activities of Natural Polyphenols. Nutrients 2014, 6, 6020. [Google Scholar] [CrossRef]
- Smiljković, M.; Kostić, M.; Stojković, D.; Glamočlija, J.; Soković, M. Could flavonoids compete with synthetic azoles in diminishing candida albicans infections? A comparative review based on in vitro studies. Curr. Med. Chem. 2019, 26, 2536–2554. [Google Scholar] [CrossRef]
- Arfaoui, L. Dietary Plant Polyphenols: Effects of Food Processing on Their Content and Bioavailability. Molecules 2021, 26, 2959. [Google Scholar] [CrossRef] [PubMed]
- Baranowska, M.; Koziara, Z.; Suliborska, K.; Chrzanowski, W.; Wormstone, M.; Namieśnik, J.; Bartoszek, A. Interactions between polyphenolic antioxidants quercetin and naringenin dictate the distinctive redox-related chemical and biological behaviour of their mixtures. Sci. Rep. 2021, 11, 12282. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, M.; Kannan, A.; Stojković, D.S.; Glamočlija, J.; Calhelha, R.C.; Ferreira, I.C.F.R.; Sanglard, D.; Soković, M. Flavones, flavonols, and glycosylated derivatives—Impact on candida albicans growth and virulence, expression of cdr1 and erg11, cytotoxicity. Pharmaceuticals 2021, 14, 27. [Google Scholar] [CrossRef]
- Ivanov, M.; Kostić, M.; Stojković, D.; Soković, M. Rosmarinic acid–Modes of antimicrobial and antibiofilm activities of common plant polyphenol. S. Afr. J. Bot. 2022, 146, 521–527. [Google Scholar] [CrossRef]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial properties of polyphenols: Characterization and QSAR (Quantitative structure-activity relationship) models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef]
- Mandalari, G.; Bennett, R.N.; Bisignano, G.; Trombetta, D.; Saija, A.; Faulds, C.B.; Gasson, M.J.; Narbad, A. Antimicrobial activity of flavonoids extracted from bergamot (Citrus bergamia Risso) peel, a byproduct of the essential oil industry. J. Appl. Microbiol. 2007, 103, 2056–2064. [Google Scholar] [CrossRef]
- Attia, G.H.; Marrez, D.A.; Mohammed, M.A.; Albarqi, H.A.; Ibrahim, A.M.; El Raey, M.A. Synergistic Effect of Mandarin Peels and Hesperidin with Sodium Nitrite against Some Food Pathogen Microbes. Molecules 2021, 26, 3186. [Google Scholar] [CrossRef]
- Duda-Madej, A.; Kozłowska, J.; Krzyżek, P.; Anioł, M.; Seniuk, A.; Jermakow, K.; Dworniczek, E. Antimicrobial O-Alkyl Derivatives of Naringenin and Their Oximes Against Multidrug-Resistant Bacteria. Molecules 2020, 25, 3642. [Google Scholar] [CrossRef]
- Dey, P.; Parai, D.; Banerjee, M.; Hossain, S.T.; Mukherjee, S.K. Naringin sensitizes the antibiofilm effect of ciprofloxacin and tetracycline against Pseudomonas aeruginosa biofilm. Int. J. Med. Microbiol. 2020, 310, 151410. [Google Scholar] [CrossRef]
- Wang, L.; Wang, G.; Qu, H.; Wang, K.; Jing, S.; Guan, S.; Su, L.; Li, Q.; Wang, D. Taxifolin, an Inhibitor of Sortase A, Interferes With the Adhesion of Methicillin-Resistant Staphylococcal aureus. Front. Microbiol. 2021, 12, 1876. [Google Scholar] [CrossRef]
- Sathiya Deepika, M.; Thangam, R.; Sakthidhasan, P.; Arun, S.; Sivasubramanian, S.; Thirumurugan, R. Combined effect of a natural flavonoid rutin from Citrus sinensis and conventional antibiotic gentamicin on Pseudomonas aeruginosa biofilm formation. Food Control 2018, 90, 282–294. [Google Scholar] [CrossRef]
- Gao, Z.; Luan, Y.; Yang, P.; Wang, L.; Zhang, H.; Jing, S.; Wang, L.; Wang, T.; Wang, D. Targeting staphylocoagulase with isoquercitrin protects mice from Staphylococcus aureus–induced pneumonia. Appl. Microbiol. Biotechnol. 2020, 104, 3909–3919. [Google Scholar] [CrossRef] [PubMed]
- Kosti, M.; Ivanov, M.; Stojkovi, D.; Sokovi, M. Antibacterial and antibiofilm activity of selected polyphenolic compounds: An in vitro study on Staphylococcus aureus. Res. Artic. Nat. Med. Mater. 2020, 40, 57. [Google Scholar] [CrossRef]
- Wu, S.-C.; Han, F.; Song, M.-R.; Chen, S.; Li, Q.; Zhang, Q.; Zhu, K.; Shen, J.-Z. Natural Flavones from Morus alba against Methicillin-Resistant Staphylococcus aureus via Targeting the Proton Motive Force and Membrane Permeability. J. Agric. Food Chem. 2019, 67, 10222–10234. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Gao, J.; Li, Q.; Guo, T.; Dong, X.; Bai, X.; Yang, J.; Hao, S.; He, F. Synergistic effect of chlorogenic acid and levofloxacin against Klebsiella pneumonia infection in vitro and in vivo. Sci. Rep. 2020, 10, 20013. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Saavedra, M.J.; Simões, M. The activity of ferulic and gallic acids in biofilm prevention and control of pathogenic bacteria. Biofouling 2012, 28, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Li, Q.; Liu, L.; Jin, W.; Wang, J.; Sun, Y. The specific effect of gallic acid on Escherichia coli biofilm formation by regulating pgaABCD genes expression. Appl. Microbiol. Biotechnol. 2018, 102, 1837–1846. [Google Scholar] [CrossRef]
- Kamatham, S.; Kumar, N.; Gudipalli, P. Isolation and characterization of gallic acid and methyl gallate from the seed coats of Givotia rottleriformis Griff. and their anti-proliferative effect on human epidermoid carcinoma A431 cells. Toxicol. Rep. 2015, 2, 520–529. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, M.; Sasaki, N.; Saga, K.; Kaneko, T. Cytotoxicity of flavonoids toward cultured normal human cells. Biol. Pharm. Bull. 2005, 28, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Liu, J.H.; Zhang, J.; Yu, B.Y. Comparative Evaluation of Cytotoxicity and Antioxidative Activity of 20 Flavonoids. J. Agric. Food Chem. 2008, 56, 3876–3883. [Google Scholar] [CrossRef]
- Trautner, B.W.; Darouiche, R.O. Role of biofilm in catheter-associated urinary tract infection. Am. J. Infect. Control 2004, 32, 177. [Google Scholar] [CrossRef] [PubMed]
- Matilla-Cuenca, L.; Gil, C.; Cuesta, S.; Rapún-Araiz, B.; Žiemytė, M.; Mira, A.; Lasa, I.; Valle, J. Antibiofilm activity of flavonoids on staphylococcal biofilms through targeting BAP amyloids. Sci. Rep. 2020, 10, 18968. [Google Scholar] [CrossRef]
- ben Sghaier, M.; Pagano, A.; Mousslim, M.; Ammari, Y.; Kovacic, H.; Luis, J. Rutin inhibits proliferation, attenuates superoxide production and decreases adhesion and migration of human cancerous cells. Biomed. Pharmacother. 2016, 84, 1972–1978. [Google Scholar] [CrossRef]
- Asadi, A.; Razavi, S.; Talebi, M.; Gholami, M. A review on anti-adhesion therapies of bacterial diseases. Infection 2018, 47, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Taniuchi, A.; May, T.; Kawata, K.; Okabe, S. Increased antibiotic resistance of Escherichia coli in mature biofilms. Appl. Environ. Microbiol. 2009, 75, 4093–4100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, J.; Yan, S.; Wang, H.; Zhao, L.; Zhu, H.; Wen, Z. Antimicrobial and antibiofilm effects of total flavonoids from Potentilla kleiniana Wight et Arn on Pseudomonas aeruginosa and its potential application to stainless steel surfaces. LWT 2022, 154, 112631. [Google Scholar] [CrossRef]
- Paczkowski, J.E.; Mukherjee, S.; McCready, A.R.; Cong, J.P.; Aquino, C.J.; Kim, H.; Henke, B.R.; Smith, C.D.; Bassler, B.L. Flavonoids Suppress Pseudomonas aeruginosa Virulence through Allosteric Inhibition of Quorum-sensing Receptors. J. Biol. Chem. 2017, 292, 4064–4076. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.Y.; Yuan, M.; Cui, Z.Q.; Wu, Z.M.; Yu, Z.J.; Song, K.; Tang, B.; Fu, B.D. Rutin inhibits quorum sensing, biofilm formation and virulence genes in avian pathogenic Escherichia coli. Microb. Pathog. 2018, 119, 54–59. [Google Scholar] [CrossRef]
- Smiljkovic, M.; Stanisavljevic, D.; Stojkovic, D.; Petrovic, I.; Vicentic, J.M.; Popovic, J.; Golic Grdadolnik, S.; Markovic, D.; Sanković-Babić, S.; Glamoclija, J.; et al. Apigenin-7-O-glucoside versus apigenin: Insight into the modes of anticandidal and cytotoxic actions. EXCLI J. 2017, 16, 795–807. [Google Scholar] [CrossRef]
- Ivanov, M.; Kannan, A.; Stojković, D.; Glamočlija, J.; Grdadolnik, S.G.; Sanglard, D.; Soković, M. Revealing the astragalin mode of anticandidal action. EXCLI J. 2020, 19, 1436–1445. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, H. Membrane Interactions of Phytochemicals as Their Molecular Mechanism Applicable to the Discovery of Drug Leads from Plants. Molecules 2015, 20, 18923–18966. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, G.R.; Vasconcelos, A.B.S.; Wu, D.T.; Li, H.B.; Antony, P.J.; Li, H.; Geng, F.; Gurgel, R.Q.; Narain, N.; Gan, R.Y. Citrus Flavonoids as Promising Phytochemicals Targeting Diabetes and Related Complications: A Systematic Review of In Vitro and In Vivo Studies. Nutrients 2020, 12, 2907. [Google Scholar] [CrossRef] [PubMed]
- Imani, A.; Maleki, N.; Bohlouli, S.; Kouhsoltani, M.; Sharifi, S.; Maleki Dizaj, S. Molecular mechanisms of anticancer effect of rutin. Phyther. Res. 2021, 35, 2500–2513. [Google Scholar] [CrossRef]
- Budzynska, B.; Faggio, C.; Kruk-Slomka, M.; Samec, D.; Nabavi, S.F.; Sureda, A.; Devi, K.P.; Nabavi, S.M. Rutin as Neuroprotective Agent: From Bench to Bedside. Curr. Med. Chem. 2017, 26, 5152–5164. [Google Scholar] [CrossRef]
- Victor, K.; Boris, L.; Athina, G.; Anthi, P.; Marija, S.; Marina, K.; Oliver, R.; Marina, S. Design, synthesis and antimicrobial activity of usnic acid derivatives. MedChemComm 2018, 9, 870–882. [Google Scholar] [CrossRef]
- Jovcic, B.; Lepsanovic, Z.; Suljagic, V.; Rackov, G.; Begovic, J.; Topisirovic, L.; Kojic, M. Emergence of NDM-1 Metallo-β-Lactamase in Pseudomonas aeruginosa Clinical Isolates from Serbia. Antimicrob. Agents Chemother. 2011, 55, 3929. [Google Scholar] [CrossRef] [Green Version]
- Novovic, K.; Vasiljevic, Z.; Kuzmanovic, M.; Lozo, J.; Begovic, J.; Kojic, M.; Jovcic, B. Novel E. coli ST5123 Containing blaNDM-1 Carried by IncF Plasmid Isolated from a Pediatric Patient in Serbia. Undefined 2016, 22, 707–711. [Google Scholar] [CrossRef]
- Novovic, K.; Mihajlovic, S.; Vasiljevic, Z.; Filipic, B.; Begovic, J.; Jovcic, B. Carbapenem-resistant Acinetobacter baumannii from Serbia: Revision of CarO classification. PLoS ONE 2015, 10, e0122793. [Google Scholar] [CrossRef] [Green Version]
- Jovcic, B.; Novovic, K.; Dekic, S.; Hrenovic, J. Colistin Resistance in Environmental Isolates of Acinetobacter baumannii. Microb. Drug Resist. 2021, 27, 328–336. [Google Scholar] [CrossRef]
- Novović, K.; Trudić, A.; Brkić, S.; Vasiljević, Z.; Kojić, M.; Medić, D.; Ćirković, I.; Jovčića, B. Molecular Epidemiology of Colistin-Resistant, Carbapenemase-Producing Klebsiella pneumoniae in Serbia from 2013 to 2016. Antimicrob. Agents Chemother. 2017, 61, e02550-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirovic, V.; Tomanovic, B.; Lepsanovic, Z.; Jovcic, B.; Kojic, M. Isolation of Klebsiella pneumoniae producing NDM-1 metallo-β-lactamase from the urine of an outpatient baby boy receiving antibiotic prophylaxis. Antimicrob. Agents Chemother. 2012, 56, 6062–6063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipic, B.; Malesevic, M.; Vasiljevic, Z.; Lukic, J.; Novovic, K.; Kojic, M.; Jovcic, B. Uncovering differences in virulence markers associated with Achromobacter species of CF and Non-CF Origin. Front. Cell. Infect. Microbiol. 2017, 7, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jovčić, B.; Novović, K.; Filipić, B.; Velhner, M.; Todorović, D.; Matović, K.; Rašić, Z.; Nikolić, S.; Kiškarolj, F.; Kojić, M. Genomic Characteristics of Colistin-Resistant Salmonella enterica subsp. enterica Serovar Infantis from Poultry Farms in the Republic of Serbia. Antibiotics 2020, 9, 886. [Google Scholar] [CrossRef]
- Haroun, M.; Tratrat, C.; Kolokotroni, A.; Petrou, A.; Geronikaki, A.; Ivanov, M.; Kostic, M.; Sokovic, M.; Carazo, A.; Mladěnka, P.; et al. 5-benzyliden-2-(5-methylthiazol-2-ylimino)thiazolidin-4-ones as antimicrobial agents. design, synthesis, biological evaluation and molecular docking studies. Antibiotics 2021, 10, 309. [Google Scholar] [CrossRef]
- Stojković, D.; Drakulić, D.; Gašić, U.; Zengin, G.; Stevanović, M.; Rajčević, N.; Soković, M. Ononis spinosa L., an edible and medicinal plant: UHPLC-LTQ-Orbitrap/MS chemical profiling and biological activities of the herbal extract. Food Funct. 2020, 11, 7138–7151. [Google Scholar] [CrossRef]
- Häkkinen, S.T.; Soković, M.; Nohynek, L.; Ćirić, A.; Ivanov, M.; Stojković, D.; Tsitko, I.; Matos, M.; Baixinho, J.P.; Ivasiv, V.; et al. Chicory Extracts and Sesquiterpene Lactones Show Potent Activity against Bacterial and Fungal Pathogens. Pharmaceuticals 2021, 14, 941. [Google Scholar] [CrossRef]
- Selvaraj, A.; Valliammai, A.; Premika, M.; Priya, A.; Bhaskar, J.P.; Krishnan, V.; Pandian, S.K. Sapindus mukorossi Gaertn. and its bioactive metabolite oleic acid impedes methicillin-resistant Staphylococcus aureus biofilm formation by down regulating adhesion genes expression. Microbiol. Res. 2021, 242, 126601. [Google Scholar] [CrossRef]
- Alipour, M.; Suntres, Z.E.; Lafrenie, R.M.; Omri, A. Attenuation of Pseudomonas aeruginosa virulence factors and biofilms by co-encapsulation of bismuth-ethanedithiol with tobramycin in liposomes. J. Antimicrob. Chemother. 2010, 65, 684–693. [Google Scholar] [CrossRef]
- Saha, S.; Thavasi, R.; Jayalakshmi, S. Phenazine pigments from Pseudomonas aeruginosa and their application as antibacterial agent and food colourants. Res. J. Microbiol. 2008, 3, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Díaz De Rienzo, M.A.; Kamalanathan, I.D.; Martin, P.J. Comparative study of the production of rhamnolipid biosurfactants by B. thailandensis E264 and P. aeruginosa ATCC 9027 using foam fractionation. Process Biochem. 2016, 51, 820–827. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Malešević, M.; Di Lorenzo, F.; Filipić, B.; Stanisavljević, N.; Novović, K.; Senerovic, L.; Polović, N.; Molinaro, A.; Kojić, M.; Jovčić, B. Pseudomonas aeruginosa quorum sensing inhibition by clinical isolate Delftia tsuruhatensis 11304: Involvement of N-octadecanoylhomoserine lactones. Sci. Rep. 2019, 9, 16465. [Google Scholar] [CrossRef]
- El-Shaer, S.; Shaaban, M.; Barwa, R.; Hassan, R. Control of quorum sensing and virulence factors of Pseudomonas aeruginosa using phenylalanine arginyl β-naphthylamide. J. Med. Microbiol. 2016, 65, 1194–1204. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.L.; Shi, Y.H.; Zhao, W.; Hao, G.; Le, G.W. Insertion mode of a novel anionic antimicrobial peptide MDpep5 (Val-Glu-Ser-Trp-Val) from Chinese traditional edible larvae of housefly and its effect on surface potential of bacterial membrane. J. Pharm. Biomed. Anal. 2008, 48, 1187–1194. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Hesperetin | Hesperidin | Naringenin | Naringin | Taxifolin | Rutin | Isoquercitrin | Morin | Chlorogenic Acid | Ferrulic Acid | p-Coumaric Acid | Gallic Acid | Streptomycin | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Methicillin resistant S. aureus IBRS MRSA 011 | MIC | 0.5 | 0.5 | 0.25 | 0.5 | 0.25 | 0.5 | 0.5 | 0.12 | 0.5 | 0.5 | 0.5 | >1 | 0.1 |
| MBC | 1 | 1 | 0.5 | 1 | 0.5 | 1 | 1 | 0.25 | 1 | 1 | 1 | >1 | 0.8 | |
| P. aeruginosa IBRS P001 | MIC | 0.5 | 0.5 | 0.25 | 0.25 | 0.25 | 0.5 | 0.25 | 0.12 | 0.5 | 0.5 | 0.5 | >1 | 0.05 |
| MBC | 1 | 1 | 0.5 | 0.5 | 0.5 | 1 | 0.5 | 0.25 | 1 | 1 | 1 | >1 | 0.1 | |
| P. aeruginosa MMA83 | MIC | 1 | 1 | 1 | >1 | 1 | 0.5 | 0.25 | 1 | 1 | 1 | 0.5 | 1 | >1 |
| MBC | 1 | 1 | 1 | >1 | >1 | 0.5 | 0.25 | 1 | >1 | 1 | 1 | 1 | >1 | |
| E. coli IBRS E003 | MIC | >1 | >1 | 0.5 | >1 | 0.5 | >1 | >1 | 0.25 | >1 | >1 | >1 | >1 | 0.1 |
| MBC | >1 | >1 | 1 | >1 | 1 | >1 | >1 | 0.5 | >1 | >1 | >1 | >1 | 0.2 | |
| E. coli IMD989 | MIC | >1 | >1 | 1 | >1 | 1 | 1 | 1 | >1 | >1 | 1 | 1 | >1 | 1 |
| MBC | >1 | >1 | >1 | >1 | 1 | 1 | 1 | >1 | >1 | 1 | 1 | >1 | >1 | |
| A. baumannii 6077/12 | MIC | 0.5 | 0.5 | 0.5 | >1 | 0.5 | 0.5 | 0.25 | 0.5 | 1 | 0.5 | 0.5 | 0.5 | >1 |
| MBC | 0.5 | 0.5 | 0.5 | >1 | 1 | 0.5 | 0.5 | 0.5 | 1 | 0.5 | 0.5 | 1 | >1 | |
| A. baumannii S2/2 | MIC | 0.5 | 0.5 | 0.5 | >1 | 0.5 | 0.5 | 0.5 | 0.5 | 1 | 0.5 | 0.5 | 0.5 | >1 |
| MBC | 0.5 | 0.5 | 0.5 | >1 | 0.5 | 0.5 | 0.5 | 0.5 | >1 | 0.5 | 0.5 | 0.5 | >1 | |
| K. pneumoniae Ni9 | MIC | 1 | 1 | 0.25 | >1 | 1 | 1 | 0.5 | 0.5 | >1 | 1 | 1 | 1 | >1 |
| MBC | 1 | 1 | 0.5 | >1 | 1 | 1 | 1 | 1 | >1 | 1 | 1 | 1 | >1 | |
| K. pneumoniae IT977 | MIC | >1 | 1 | 1 | >1 | >1 | 1 | 1 | >1 | >1 | 1 | 1 | 1 | >1 |
| MBC | >1 | 1 | 1 | >1 | >1 | 1 | 1 | >1 | >1 | 1 | 1 | 1 | >1 | |
| A. xylosoxidans 37F | MIC | 1 | 1 | 1 | >1 | 1 | 0.5 | 0.5 | 0.5 | 1 | 1 | 1 | 0.5 | >1 |
| MBC | 1 | 1 | 1 | >1 | 1 | 1 | 0.5 | 1 | >1 | 1 | 1 | 1 | >1 | |
| S. infantis 1371/1 | MIC | >1 | >1 | >1 | >1 | >1 | 1 | 1 | >1 | >1 | 1 | 1 | >1 | >1 |
| MBC | >1 | >1 | >1 | >1 | >1 | 1 | 1 | >1 | >1 | 1 | 1 | >1 | >1 |
| Polyphenol | IC50 (mg/mL) |
|---|---|
| Hesperetin | >1 |
| Hesperidin | >1 |
| Naringenin | 0.528 ± 0.047 |
| Naringin | >1 |
| Taxifolin | 0.495 ± 0.047 |
| Rutin | >1 |
| Isoquercitrin | >1 |
| Morin | 0.347 ± 0.016 |
| Chlorogenic acid | 0.279 ± 0.008 |
| Ferulic acid | >1 |
| p-coumaric acid | >1 |
| Gallic acid | <0.080 |
| Strain | Resistance | Reference |
|---|---|---|
| Methicillin-resistant Staphylococcus aureus IBRS MRSA 011 | cefoxitin | [57] |
| Pseudomonas aeruginosa IBRS P001 | penicillin, ampicillin, amoxicillin, tetracycline, neomycin, gentamicin, ceftriaxone | [57] |
| P. aeruginosa MMA83 | imipenem, meropenem, gentamycin | [58] |
| Escherichia coli IBRS E003 | penicillin, ampicillin, amoxicillin, tetracycline, neomycin, gentamicin, ceftriaxone | [57] |
| E. coli IMD989 | amoxicillin/clavulanate, ampicillin/sulbactam, piperacillin/tazobactam, cefuroxime, cefotaxime, ceftazidime, cefepime, imipenem, meropenem, trimethoprim/sulfamethoxazole, amikacin, gentamicin | [59] |
| Acinetobacter baumannii 6077/12 | amoxicillin/clavulanate, ampicillin/sulbactam, piperacillin/tazobactam, cefoxitin, cefotaxime, ceftazidime, cefepime, aztreonam, imipenem, meropenem | [60] |
| A. baumannii S2/2 | imipenem, meropenem, ciprofloxacin, levofloxacin, amikacin, gentamicin, tobramycin, trimethoprim/sulfamethoxazole, colistin | [61] |
| Klebsiella pneumoniae Ni9 | imipenem, meropenem, colistin | [62] |
| K. pneumoniae IT977 | ampicillin/clavulanate, piperacillin/tazobactam, cefazolin, ceftriaxone, cefepime, aztreonam, ertapenem, imipenem, meropenem, ciprofloxacin, moxifloxacin, gentamicin, tobramycin, nitrofurantoin | [63] |
| Achromobacter xylosoxidans 37F | tetracycline, chloramphenicol, ciprofloxacin, levofloxacin, trimethoprim/sulfamethoxazole | [64] |
| Salmonella Infantis 1371/1 | tetracycline, quinolones, colistin | [65] |
| Gene | Primer Direction | Sequence (5′-3′) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| lasI | Forward | GCGTGCTCAAGTGTTCAAGG | 125 | [74] |
| Reverse | GGGCTTCAGGAGTATCTTCCTGG | |||
| lasR | Forward | CTGTGGATGCTCAAGGACTAC | 133 | [75] |
| Reverse | AACTGGTCTTGCCGATGG | |||
| rhlI | Forward | CCATCCGCAAACCCGCTACATC | 151 | [74] |
| Reverse | CTCCCAGACCGACGGATCGCTCGGC | |||
| rhlR | Forward | GGGCGTGTTCGCCGTCCTGG | 143 | [74] |
| Reverse | GGTATCGCTCCAGCCAGGCCTTG | |||
| pqsA | Forward | GACCGGCTGTATTCGATTC | 74 | [75] |
| Reverse | GCTGAACCAGGGAAAGAAC | |||
| mvfR | Forward | GTCGGGACGGCTACAAGGTCG | 129 | [74] |
| Reverse | GATTGCGCGGACCCTTGTTGAG | |||
| rpsL | Forward | GCAACTATCAACCAGCTGGTG | 231 | [74] |
| Reverse | GCTGTGCTCTTGCAGGTTGTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanov, M.; Novović, K.; Malešević, M.; Dinić, M.; Stojković, D.; Jovčić, B.; Soković, M. Polyphenols as Inhibitors of Antibiotic Resistant Bacteria—Mechanisms Underlying Rutin Interference with Bacterial Virulence. Pharmaceuticals 2022, 15, 385. https://doi.org/10.3390/ph15030385
Ivanov M, Novović K, Malešević M, Dinić M, Stojković D, Jovčić B, Soković M. Polyphenols as Inhibitors of Antibiotic Resistant Bacteria—Mechanisms Underlying Rutin Interference with Bacterial Virulence. Pharmaceuticals. 2022; 15(3):385. https://doi.org/10.3390/ph15030385
Chicago/Turabian StyleIvanov, Marija, Katarina Novović, Milka Malešević, Miroslav Dinić, Dejan Stojković, Branko Jovčić, and Marina Soković. 2022. "Polyphenols as Inhibitors of Antibiotic Resistant Bacteria—Mechanisms Underlying Rutin Interference with Bacterial Virulence" Pharmaceuticals 15, no. 3: 385. https://doi.org/10.3390/ph15030385
APA StyleIvanov, M., Novović, K., Malešević, M., Dinić, M., Stojković, D., Jovčić, B., & Soković, M. (2022). Polyphenols as Inhibitors of Antibiotic Resistant Bacteria—Mechanisms Underlying Rutin Interference with Bacterial Virulence. Pharmaceuticals, 15(3), 385. https://doi.org/10.3390/ph15030385