
The Role of the Iron Protoporphyrins Heme and Hematin in the Antimalarial Activity of Endoperoxide Drugs



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Affinity between Peroxides and Iron Protoporphyrins
2. Introduction to Heme Detoxification
3. Introduction to Redox Chemistry
4. Interplay between Redox Homeostasis and Heme Detoxification
5. Heme Alkylation versus Protein Alkylation: The Debate
6. Frontier Questions and Perspectives
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gonciarz, R.L.; Collisson, E.A.; Renslo, A.R. Ferrous Iron-Dependent Pharmacology. Trends Pharmacol. Sci. 2021, 42, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Meshnick, S.R.; Thomas, A.; Ranz, A.; Xu, C.-M.; Pan, H.-Z. Artemisinin (qinghaosu): The role of intracellular hemin in its mechanism of antimalarial action. Mol. Biochem. Parasitol. 1991, 49, 181–189. [Google Scholar] [CrossRef]
- Meunier, B.; Robert, A. Heme as Trigger and Target for Trioxane-Containing Antimalarial Drugs. Acc. Chem. Res. 2010, 43, 1444–1451. [Google Scholar] [CrossRef] [PubMed]
- Kannan, R.; Sahal, D.; Chauhan, V. Heme-Artemisinin Adducts Are Crucial Mediators of the Ability of Artemisinin to Inhibit Heme Polymerization. Chem. Biol. 2002, 9, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Robert, A.; Bonduelle, C.; Laurent, S.A.-L.; Meunier, B. Heme alkylation by artemisinin and trioxaquines. J. Phys. Org. Chem. 2006, 19, 562–569. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, C.-J.; Chia, W.N.; Loh, C.C.Y.; Li, Z.; Lee, Y.M.; He, Y.; Yuan, L.-X.; Lim, T.K.; Liu, M.; et al. Haem-activated promiscuous targeting of artemisinin in Plasmodium falciparum. Nat. Commun. 2015, 6, 10111. [Google Scholar] [CrossRef]
- Ismail, H.M.; Barton, V.E.; Panchana, M.; Charoensutthivarakul, S.; Biagini, G.A.; Ward, S.A.; O’Neill, P.M. A Click Chemistry-Based Proteomic Approach Reveals that 1,2,4-Trioxolane and Artemisinin Antimalarials Share a Common Protein Alkylation Profile. Angew. Chemie Int. Ed. 2016, 55, 6401–6405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, A.; Meunier, B. Characterization of the First Covalent Adduct between Artemisinin and a Heme Model. J. Am. Chem. Soc. 1997, 119, 5968–5969. [Google Scholar] [CrossRef]
- Zhang, S.; Gerhard, G.S. Heme activates artemisinin more efficiently than hemin, inorganic iron, or hemoglobin. Bioorg. Med. Chem. 2008, 16, 7853–7861. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; He, Y.; Li, Y.; Zhang, X.; Wong, Y.-K.; Shen, S.; Zhong, T.; Zhang, J.; Liu, Q.; Wang, J. Advances in the research on the targets of anti-malaria actions of artemisinin. Pharmacol. Ther. 2020, 216, 107697. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, P.M.; Posner, G.H. A Medicinal Chemistry Perspective on Artemisinin and Related Endoperoxides. J. Med. Chem. 2004, 47, 2945–2964. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, T.; Çapcı Karagöz, A.; Reiter, C.; Tsogoeva, S.B. Artemisinin-Derived Dimers: Potent Antimalarial and Anticancer Agents. J. Med. Chem. 2016, 59, 7360–7388. [Google Scholar] [CrossRef] [PubMed]
- Wittlin, S.; Mäser, P. From Magic Bullet to Magic Bomb: Reductive Bioactivation of Antiparasitic Agents. ACS Infect. Dis. 2021, 7, 2777–2786. [Google Scholar] [CrossRef] [PubMed]
- Choby, J.E.; Skaar, E.P. Heme Synthesis and Acquisition in Bacterial Pathogens. J. Mol. Biol. 2016, 428, 3408–3428. [Google Scholar] [CrossRef] [Green Version]
- Kloehn, J.; Harding, C.R.; Soldati-Favre, D. Supply and demand—Heme synthesis, salvage and utilization by Apicomplexa. FEBS J. 2021, 288, 382–404. [Google Scholar] [CrossRef]
- Pamplona, A.; Ferreira, A.; Balla, J.; Jeney, V.; Balla, G.; Epiphanio, S.; Chora, Â.; Rodrigues, C.D.; Gregoire, I.P.; Cunha-Rodrigues, M.; et al. Heme oxygenase-1 and carbon monoxide suppress the pathogenesis of experimental cerebral malaria. Nat. Med. 2007, 13, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Nasamu, A.S.; Polino, A.J.; Istvan, E.S.; Goldberg, D.E. Malaria parasite plasmepsins: More than just plain old degradative pepsins. J. Biol. Chem. 2020, 295, 8425–8441. [Google Scholar] [CrossRef]
- Rosenthal, P.J. Falcipain cysteine proteases of malaria parasites: An update. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140362. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Spaccapelo, R.; Schwarzer, E.; Sajid, M.; Annoura, T.; Deroost, K.; Ravelli, R.B.G.; Aime, E.; Capuccini, B.; Mommaas-Kienhuis, A.M.; et al. Replication of Plasmodium in reticulocytes can occur without hemozoin formation, resulting in chloroquine resistance. J. Exp. Med. 2015, 212, 893–903. [Google Scholar] [CrossRef] [Green Version]
- Sigala, P.A.; Goldberg, D.E. The Peculiarities and Paradoxes of Plasmodium Heme Metabolism. Annu. Rev. Microbiol. 2014, 68, 259–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slater, A.F.; Swiggard, W.J.; Orton, B.R.; Flitter, W.D.; Goldberg, D.E.; Cerami, A.; Henderson, G.B. An iron-carboxylate bond links the heme units of malaria pigment. Proc. Natl. Acad. Sci. USA 1991, 88, 325–329. [Google Scholar] [CrossRef] [Green Version]
- Kapishnikov, S.; Weiner, A.; Shimoni, E.; Guttmann, P.; Schneider, G.; Dahan-Pasternak, N.; Dzikowski, R.; Leiserowitz, L.; Elbaum, M. Oriented nucleation of hemozoin at the digestive vacuole membrane in Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2012, 109, 11188–11193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moxon, C.A.; Gibbins, M.P.; McGuinness, D.; Milner, D.A.; Marti, M. New Insights into Malaria Pathogenesis. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 315–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Villiers, K.A.; Egan, T.J. Heme Detoxification in the Malaria Parasite: A Target for Antimalarial Drug Development. Acc. Chem. Res. 2021, 54, 2649–2659. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, M.D.; Schneider, T.G.; Taraschi, T.F. A new model for hemoglobin ingestion and transport by the human malaria parasite Plasmodium falciparum. J. Cell Sci. 2008, 121, 1937–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milani, K.J.; Schneider, T.G.; Taraschi, T.F. Defining the Morphology and Mechanism of the Hemoglobin Transport Pathway in Plasmodium falciparum-Infected Erythrocytes. Eukaryot. Cell 2015, 14, 415–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, H.M.; Apte, S.H.; Chojnowski, G.M.; Hänscheid, T.; Rebelo, M.; Grimberg, B.T. Cytometry in Malaria—A Practical Replacement for Microscopy? Curr. Protoc. Cytom. 2013, 65, 11.20.1–11.20.23. [Google Scholar] [CrossRef] [PubMed]
- Buapetch, W.; Maude, R.J.; Silamut, K. A Simplified, Low-Cost Method for Polarized Light Microscopy. Am. J. Trop. Med. Hyg. 2009, 81, 782–783. [Google Scholar] [CrossRef] [Green Version]
- Hänscheid, T.; Egan, T.J.; Grobusch, M.P. Haemozoin: From melatonin pigment to drug target, diagnostic tool, and immune modulator. Lancet Infect. Dis. 2007, 7, 675–685. [Google Scholar] [CrossRef]
- Kapishnikov, S.; Hempelmann, E.; Elbaum, M.; Als-Nielsen, J.; Leiserowitz, L. Malaria Pigment Crystals: The Achilles′ Heel of the Malaria Parasite. ChemMedChem 2021, 16, 1515–1532. [Google Scholar] [CrossRef] [PubMed]
- Warhurst, D.C.; Hockley, D.J. Mode of Action of Chloroquine on Plasmodium berghei and P. cynomolgi. Nature 1967, 214, 935–936. [Google Scholar] [CrossRef] [PubMed]
- Egan, T.J.; Ross, D.C.; Adams, P.A. Quinoline anti-malarial drugs inhibit spontaneous formation of β-haematin (malaria pigment). FEBS Lett. 1994, 352, 54–57. [Google Scholar] [CrossRef] [Green Version]
- Peyton, D.H. Reversed Chloroquine Molecules as a Strategy to Overcome Resistance in Malaria. Curr. Top. Med. Chem. 2012, 12, 400–407. [Google Scholar] [CrossRef] [Green Version]
- Roepe, P.D. To kill or not to kill, that is the question: Cytocidal antimalarial drug resistance. Trends Parasitol. 2014, 30, 130–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olafson, K.N.; Nguyen, T.Q.; Rimer, J.D.; Vekilov, P.G. Antimalarials inhibit hematin crystallization by unique drug–surface site interactions. Proc. Natl. Acad. Sci. USA 2017, 114, 7531–7536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorn, A.; Vippagunta, S.R.; Matile, H.; Jaquet, C.; Vennerstrom, J.L.; Ridley, R.G. An Assessment of Drug-Haematin Binding as a Mechanism for Inhibition of Haematin Polymerisation by Quinoline Antimalarials. Biochem. Pharmacol. 1998, 55, 727–736. [Google Scholar] [CrossRef]
- Parapini, S.; Basilico, N.; Pasini, E.; Egan, T.J.; Olliaro, P.; Taramelli, D.; Monti, D. Standardization of the Physicochemical Parameters to Assess in Vitro the β-Hematin Inhibitory Activity of Antimalarial Drugs. Exp. Parasitol. 2000, 96, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Haynes, R.K.; Monti, D.; Taramelli, D.; Basilico, N.; Parapini, S.; Olliaro, P. Artemisinin Antimalarials Do Not Inhibit Hemozoin Formation. Antimicrob. Agents Chemother. 2003, 47, 1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, C.R.; Sullivan, D.J. Inhibition of heme crystal growth by antimalarials and other compounds: Implications for drug discovery. Biochem. Pharmacol. 2003, 66, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Ribbiso, K.A.; Heller, L.E.; Taye, A.; Julian, E.; Willems, A.V.; Roepe, P.D. Artemisinin-Based Drugs Target the Plasmodium falciparum Heme Detoxification Pathway. Antimicrob. Agents Chemother. 2021, 65, e02137-20. [Google Scholar] [CrossRef]
- Ma, W.; Balta, V.A.; West, R.; Newlin, K.N.; Miljanić, O.Š.; Sullivan, D.J.; Vekilov, P.G.; Rimer, J.D. A second mechanism employed by artemisinins to suppress Plasmodium falciparum hinges on inhibition of hematin crystallization. J. Biol. Chem. 2021, 296, 100123. [Google Scholar] [CrossRef] [PubMed]
- Kapishnikov, S.; Staalsø, T.; Yang, Y.; Lee, J.; Pérez-Berná, A.J.; Pereiro, E.; Yang, Y.; Werner, S.; Guttmann, P.; Leiserowitz, L.; et al. Mode of action of quinoline antimalarial drugs in red blood cells infected by Plasmodium falciparum revealed in vivo. Proc. Natl. Acad. Sci. USA 2019, 116, 22946–22952. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.-L.; Yang, Y.-Z.; Meshnick, S.R. The interaction of artemisinin with malarial hemozoin. Mol. Biochem. Parasitol. 1994, 63, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Heller, L.E.; Goggins, E.; Roepe, P.D. Dihydroartemisinin–Ferriprotoporphyrin IX Adduct Abundance in Plasmodium falciparum Malarial Parasites and the Relationship to Emerging Artemisinin Resistance. Biochemistry 2018, 57, 6935–6945. [Google Scholar] [CrossRef]
- Combrinck, J.M.; Mabotha, T.E.; Ncokazi, K.K.; Ambele, M.A.; Taylor, D.; Smith, P.J.; Hoppe, H.C.; Egan, T.J. Insights into the Role of Heme in the Mechanism of Action of Antimalarials. ACS Chem. Biol. 2013, 8, 133–137. [Google Scholar] [CrossRef] [Green Version]
- Çapcı, A.; Lorion, M.M.; Wang, H.; Simon, N.; Leidenberger, M.; Borges Silva, M.C.; Moreira, D.R.M.; Zhu, Y.; Meng, Y.; Chen, J.Y.; et al. Artemisinin–(Iso)quinoline Hybrids by C−H Activation and Click Chemistry: Combating Multidrug-Resistant Malaria. Angew. Chemie Int. Ed. 2019, 58, 13066–13079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abshire, J.R.; Rowlands, C.J.; Ganesan, S.M.; So, P.T.C.; Niles, J.C. Quantification of labile heme in live malaria parasites using a genetically encoded biosensor. Proc. Natl. Acad. Sci. USA 2017, 114, E2068–E2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monti, D.; Vodopivec, B.; Basilico, N.; Olliaro, P.; Taramelli, D. A Novel Endogenous Antimalarial: Fe(II)-Protoporphyrin IXα (Heme) Inhibits Hematin Polymerization to β-Hematin (Malaria Pigment) and Kills Malaria Parasites. Biochemistry 1999, 38, 8858–8863. [Google Scholar] [CrossRef]
- Iyer, J.K.; Shi, L.; Shankar, A.H.; Sullivan, D.J. Zinc protoporphyrin IX binds heme crystals to inhibit the process of crystallization in Plasmodium falciparum. Mol. Med. 2003, 9, 1. [Google Scholar] [CrossRef]
- Begum, K.; Kim, H.-S.; Kumar, V.; Stojiljkovic, I.; Wataya, Y. In vitro antimalarial activity of metalloporphyrins against Plasmodium falciparum. Parasitol. Res. 2003, 90, 221–224. [Google Scholar] [CrossRef]
- Martiney, J.A.; Cerami, A.; Slater, A.F.G. Inhibition of Hemozoin Formation in Plasmodium falciparum Trophozoite Extracts by Heme Analogs: Possible Implication in the Resistance to Malaria Conferred by the β-Thalassemia Trait. Mol. Med. 1996, 2, 236–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Openshaw, R.; Maepa, K.; Benjamin, S.J.; Wainwright, L.; Combrinck, J.M.; Hunter, R.; Egan, T.J. A Diverse Range of Hemozoin Inhibiting Scaffolds Act on Plasmodium falciparum as Heme Complexes. ACS Infect. Dis. 2021, 7, 362–376. [Google Scholar] [CrossRef] [PubMed]
- Egwu, C.O.; Pério, P.; Augereau, J.-M.; Tsamesidis, I.; Benoit-Vical, F.; Reybier, K. Resistance to artemisinin in falciparum malaria parasites: A redox-mediated phenomenon. Free Radic. Biol. Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Egwu, C.O.; Tsamesidis, I.; Pério, P.; Augereau, J.-M.; Benoit-Vical, F.; Reybier, K. Superoxide: A major role in the mechanism of action of essential antimalarial drugs. Free Radic. Biol. Med. 2021, 167, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.; Sigala, P.A.; Miura, K.; Morrisey, J.M.; Mather, M.W.; Crowley, J.R.; Henderson, J.P.; Goldberg, D.E.; Long, C.A.; Vaidya, A.B. The Heme Biosynthesis Pathway Is Essential for Plasmodium falciparum Development in Mosquito Stage but Not in Blood Stages. J. Biol. Chem. 2014, 289, 34827–34837. [Google Scholar] [CrossRef] [Green Version]
- Rathnapala, U.L.; Goodman, C.D.; McFadden, G.I. A novel genetic technique in Plasmodium berghei allows liver stage analysis of genes required for mosquito stage development and demonstrates that de novo heme synthesis is essential for liver stage development in the malaria parasite. PLoS Pathog. 2017, 13, e1006396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, A.; Coppel, Y.; Meunier, B. Alkylation of heme by the antimalarial drug artemisinin. Chem. Commun. 2002, 5, 414–415. [Google Scholar] [CrossRef] [PubMed]
- Laurent, S.A.-L.; Robert, A.; Meunier, B. C10-Modified Artemisinin Derivatives: Efficient Heme-Alkylating Agents. Angew. Chemie 2005, 117, 2096–2099. [Google Scholar] [CrossRef]
- Stocks, P.A.; Bray, P.G.; Barton, V.E.; Al-Helal, M.; Jones, M.; Araujo, N.C.; Gibbons, P.; Ward, S.A.; Hughes, R.H.; Biagini, G.A.; et al. Evidence for a Common Non-Heme Chelatable-Iron-Dependent Activation Mechanism for Semisynthetic and Synthetic Endoperoxide Antimalarial Drugs. Angew. Chemie Int. Ed. 2007, 46, 6278–6283. [Google Scholar] [CrossRef]
- Bousejra-El Garah, F.; Wong, M.H.-L.; Amewu, R.K.; Muangnoicharoen, S.; Maggs, J.L.; Stigliani, J.-L.; Park, B.K.; Chadwick, J.; Ward, S.A.; O’Neill, P.M. Comparison of the Reactivity of Antimalarial 1,2,4,5-Tetraoxanes with 1,2,4-Trioxolanes in the Presence of Ferrous Iron Salts, Heme, and Ferrous Iron Salts/Phosphatidylcholine. J. Med. Chem. 2011, 54, 6443–6455. [Google Scholar] [CrossRef] [PubMed]
- Creek, D.J.; Charman, W.N.; Chiu, F.C.K.; Prankerd, R.J.; Dong, Y.; Vennerstrom, J.L.; Charman, S.A. Relationship between Antimalarial Activity and Heme Alkylation for Spiro- and Dispiro-1,2,4-Trioxolane Antimalarials. Antimicrob. Agents Chemother. 2008, 52, 1291–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, A.; Claparols, C.; Witkowski, B.; Benoit-Vical, F. Correlation between Plasmodium yoelii nigeriensis Susceptibility to Artemisinin and Alkylation of Heme by the Drug. Antimicrob. Agents Chemother. 2013, 57, 3998–4000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heller, L.E.; Roepe, P.D. Quantification of Free Ferriprotoporphyrin IX Heme and Hemozoin for Artemisinin Sensitive versus Delayed Clearance Phenotype Plasmodium falciparum Malarial Parasites. Biochemistry 2018, 57, 6927–6934. [Google Scholar] [CrossRef]
- Heller, L.E.; Roepe, P.D. Artemisinin-Based Antimalarial Drug Therapy: Molecular Pharmacology and Evolving Resistance. Trop. Med. Infect. Dis. 2019, 4, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loup, C.; Lelièvre, J.; Benoit-Vical, F.; Meunier, B. Trioxaquines and Heme-Artemisinin Adducts Inhibit the In Vitro Formation of Hemozoin Better than Chloroquine. Antimicrob. Agents Chemother. 2007, 51, 3768–3770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, K.A.; Ziegler, J.; Evans, C.A.; Wright, D.W. Metalloporphyrins inhibit β-hematin (hemozoin) formation. J. Inorg. Biochem. 2000, 78, 109–115. [Google Scholar] [CrossRef]
- Alves, E.; Iglesias, B.A.; Deda, D.K.; Budu, A.; Matias, T.A.; Bueno, V.B.; Maluf, F.V.; Guido, R.V.C.; Oliva, G.; Catalani, L.H.; et al. Encapsulation of metalloporphyrins improves their capacity to block the viability of the human malaria parasite Plasmodium falciparum. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Delves, M.; Plouffe, D.; Scheurer, C.; Meister, S.; Wittlin, S.; Winzeler, E.A.; Sinden, R.E.; Leroy, D. The Activities of Current Antimalarial Drugs on the Life Cycle Stages of Plasmodium: A Comparative Study with Human and Rodent Parasites. PLoS Med. 2012, 9, e1001169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coghi, P.; Basilico, N.; Taramelli, D.; Chan, W.-C.; Haynes, R.K.; Monti, D. Interaction of Artemisinins with Oxyhemoglobin Hb-Fe II, Hb-Fe II, CarboxyHb-Fe II, Heme-Fe II, and Carboxyheme Fe II: Significance for Mode of Action and Implications for Therapy of Cerebral Malaria. ChemMedChem 2009, 4, 2045–2053. [Google Scholar] [CrossRef] [PubMed]
- Parapini, S.; Olliaro, P.; Navaratnam, V.; Taramelli, D.; Basilico, N. Stability of the Antimalarial Drug Dihydroartemisinin under Physiologically Relevant Conditions: Implications for Clinical Treatment and Pharmacokinetic and In Vitro Assays. Antimicrob. Agents Chemother. 2015, 59, 4046–4052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, G.; Gao, Y.; Liu, S.; Shui, S.; Liu, G. pH-dependent rearrangement determines the iron-activation and antitumor activity of artemisinins. Free Radic. Biol. Med. 2021, 163, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.K.; McChesney, J.D.; Chi, H.T. Decomposition of Arteether in Simulated Stomach Acid Yielding Compounds Retaining Antimalarial Activity. Pharm. Res. An Off. J. Am. Assoc. Pharm. Sci. 1993, 10, 662–666. [Google Scholar]
- Wu, Y.; Wu, R.W.K.; Cheu, K.W.; Williams, I.D.; Krishna, S.; Slavic, K.; Gravett, A.M.; Liu, W.M.; Wong, H.N.; Haynes, R.K. Methylene Homologues of Artemisone: An Unexpected Structure-Activity Relationship and a Possible Implication for the Design of C10-Substituted Artemisinins. ChemMedChem 2016, 11, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Pacorel, B.; Leung, S.C.; Stachulski, A.V.; Davies, J.; Vivas, L.; Lander, H.; Ward, S.A.; Kaiser, M.; Brun, R.; O’Neill, P.M. Modular Synthesis and in Vitro and in Vivo Antimalarial Assessment of C-10 Pyrrole Mannich Base Derivatives of Artemisinin. J. Med. Chem. 2010, 53, 633–640. [Google Scholar] [CrossRef]
- Jourdan, J.; Walz, A.; Matile, H.; Schmidt, A.; Wu, J.; Wang, X.; Dong, Y.; Vennerstrom, J.L.; Schmidt, R.S.; Wittlin, S.; et al. Stochastic Protein Alkylation by Antimalarial Peroxides. ACS Infect. Dis. 2019, 5, 2067–2075. [Google Scholar] [CrossRef] [PubMed]
- Klayman, D.L. Qinghaosu (Artemisinin): An Antimalarial Drug from China. Science 1985, 228, 1049–1055. [Google Scholar] [CrossRef] [Green Version]
- Posner, G.H.; Oh, C.H.; Wang, D.; Gerena, L.; Milhous, W.K.; Meshnick, S.R.; Asawamahasadka, W. Mechanism-Based Design, Synthesis, and in vitro Antimalarial Testing of New 4-Methylated Trioxanes Structurally Related to Artemisinin: The Importance of a Carbon-Centered Radical for Antimalarial Activity. J. Med. Chem. 1994, 37, 1256–1258. [Google Scholar] [CrossRef] [PubMed]
- Lichorowic, C.L.; Zhao, Y.; Maher, S.P.; Padín-Irizarry, V.; Mendiola, V.C.; de Castro, S.T.; Worden, J.A.; Casandra, D.; Kyle, D.E.; Manetsch, R. Synthesis of Mono- and Bisperoxide-Bridged Artemisinin Dimers to Elucidate the Contribution of Dimerization to Antimalarial Activity. ACS Infect. Dis. 2021, 7, 2013–2024. [Google Scholar] [CrossRef]
- Jortzik, E.; Becker, K. Thioredoxin and glutathione systems in Plasmodium falciparum. Int. J. Med. Microbiol. 2012, 302, 187–194. [Google Scholar] [CrossRef]
- Giannangelo, C.; Anderson, D.; Wang, X.; Vennerstrom, J.L.; Charman, S.A.; Creek, D.J. Ozonide Antimalarials Alkylate Heme in the Malaria Parasite Plasmodium falciparum. ACS Infect. Dis. 2019, 5, 2076–2086. [Google Scholar] [CrossRef] [PubMed]
- Heller, L.; Roepe, P.D.; de Dios, A.C. Artesunate activation by heme in an aqueous medium. Inorganica Chim. Acta 2019, 496, 119029. [Google Scholar] [CrossRef] [PubMed]
- Eckstein-Ludwig, U.; Webb, R.J.; van Goethem, I.D.A.; East, J.M.; Lee, A.G.; Kimura, M.; O’Neill, P.M.; Bray, P.G.; Ward, S.A.; Krishna, S. Artemisinins target the SERCA of Plasmodium falciparum. Nature 2003, 424, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Sissoko, A.; Vásquez-Ocmín, P.; Maciuk, A.; Barbieri, D.; Neveu, G.; Rondepierre, L.; Grougnet, R.; Leproux, P.; Blaud, M.; Hammad, K.; et al. A Chemically Stable Fluorescent Mimic of Dihydroartemisinin, Artemether, and Arteether with Conserved Bioactivity and Specificity Shows High Pharmacological Relevance to the Antimalarial Drugs. ACS Infect. Dis. 2020, 6, 1532–1547. [Google Scholar] [CrossRef] [PubMed]
- Jida, M.; Sanchez, C.P.; Urgin, K.; Ehrhardt, K.; Mounien, S.; Geyer, A.; Elhabiri, M.; Lanzer, M.; Davioud-Charvet, E. A Redox-Active Fluorescent pH Indicator for Detecting Plasmodium falciparum Strains with Reduced Responsiveness to Quinoline Antimalarial Drugs. ACS Infect. Dis. 2017, 3, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Woodland, J.G.; Hunter, R.; Smith, P.J.; Egan, T.J. Chemical Proteomics and Super-resolution Imaging Reveal That Chloroquine Interacts with Plasmodium falciparum Multidrug Resistance-Associated Protein and Lipids. ACS Chem. Biol. 2018, 13, 2939–2948. [Google Scholar] [CrossRef]
- Adjalley, S.H.; Johnston, G.L.; Li, T.; Eastman, R.T.; Ekland, E.H.; Eappen, A.G.; Richman, A.; Sim, B.K.L.; Lee, M.C.S.; Hoffman, S.L.; et al. Quantitative assessment of Plasmodium falciparum sexual development reveals potent transmission-blocking activity by methylene blue. Proc. Natl. Acad. Sci. USA 2011, 108, E1214–E1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quadros, H.C.; Çapcı, A.; Herrmann, L.; D’Alessandro, S.; Fontinha, D.; Azevedo, R.; Villarreal, W.; Basilico, N.; Prudêncio, M.; Tsogoeva, S.B.; et al. Studies of Potency and Efficacy of an Optimized Artemisinin-Quinoline Hybrid against Multiple Stages of the Plasmodium Life Cycle. Pharmaceuticals 2021, 14, 1129. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, G.; Santha Kumar, T.R.; Bona, R.; Camarda, G.; Calabretta, M.M.; Cevenini, L.; Davioud-Charvet, E.; Becker, K.; Cara, A.; Fidock, D.A.; et al. A high susceptibility to redox imbalance of the transmissible stages of Plasmodium falciparum revealed with a luciferase-based mature gametocyte assay. Mol. Microbiol. 2017, 104, 306–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haynes, R.; Cheu, K.-W.; N’Da, D.; Coghi, P.; Monti, D. Considerations on the Mechanism of Action of Artemisinin Antimalarials: Part 1—The ’Carbon Radical’ and ’Heme’ Hypotheses. Infect. Disord. Drug Targets. 2013, 13, 217–277. [Google Scholar] [CrossRef] [PubMed]
- Tsogoeva, S.B. Recent Progress in the Development of Synthetic Hybrids of Natural or Unnatural Bioactive Compounds for Medicinal Chemistry. Mini-Reviews Med. Chem. 2010, 10, 773–793. [Google Scholar] [CrossRef]
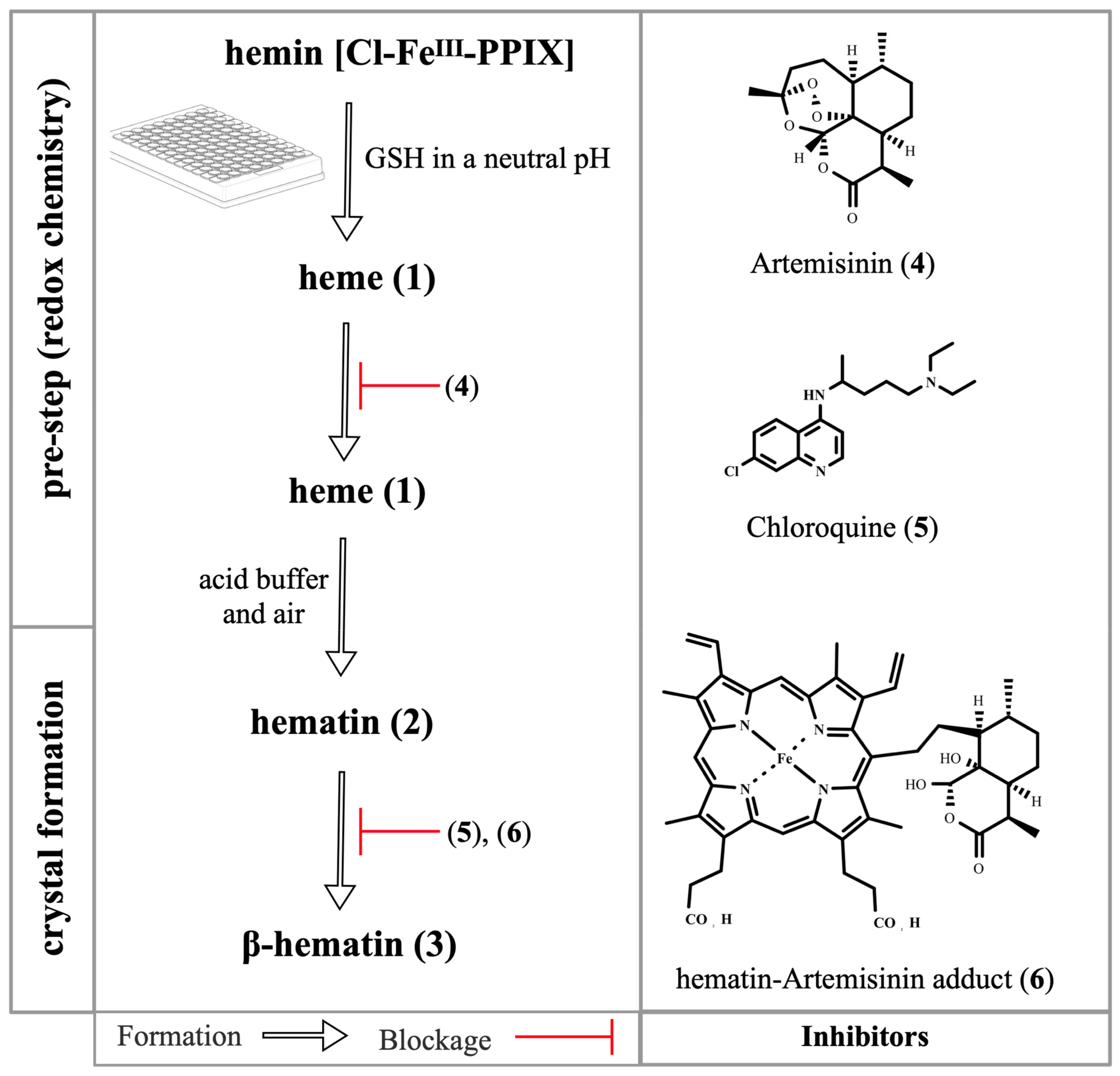
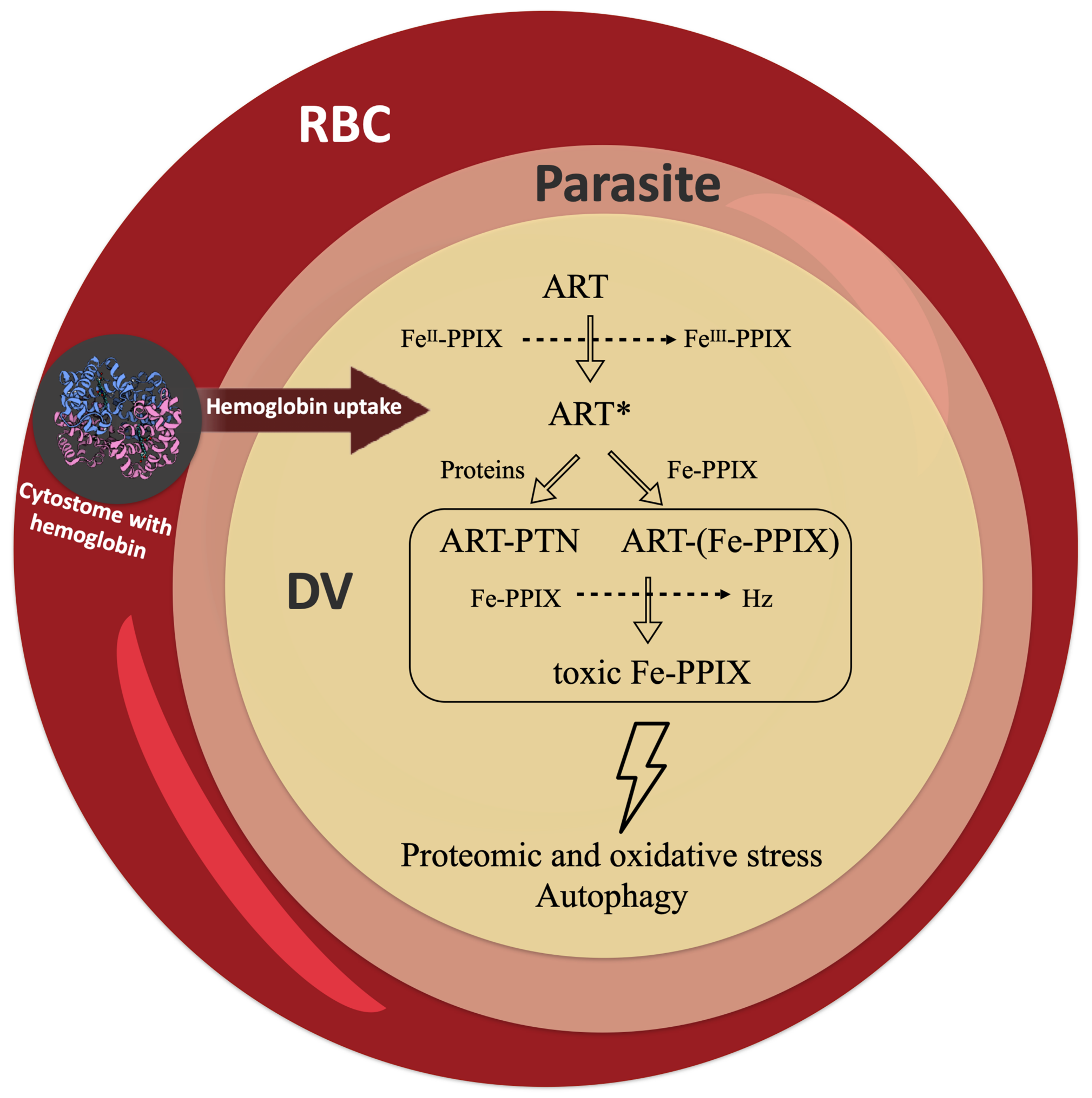
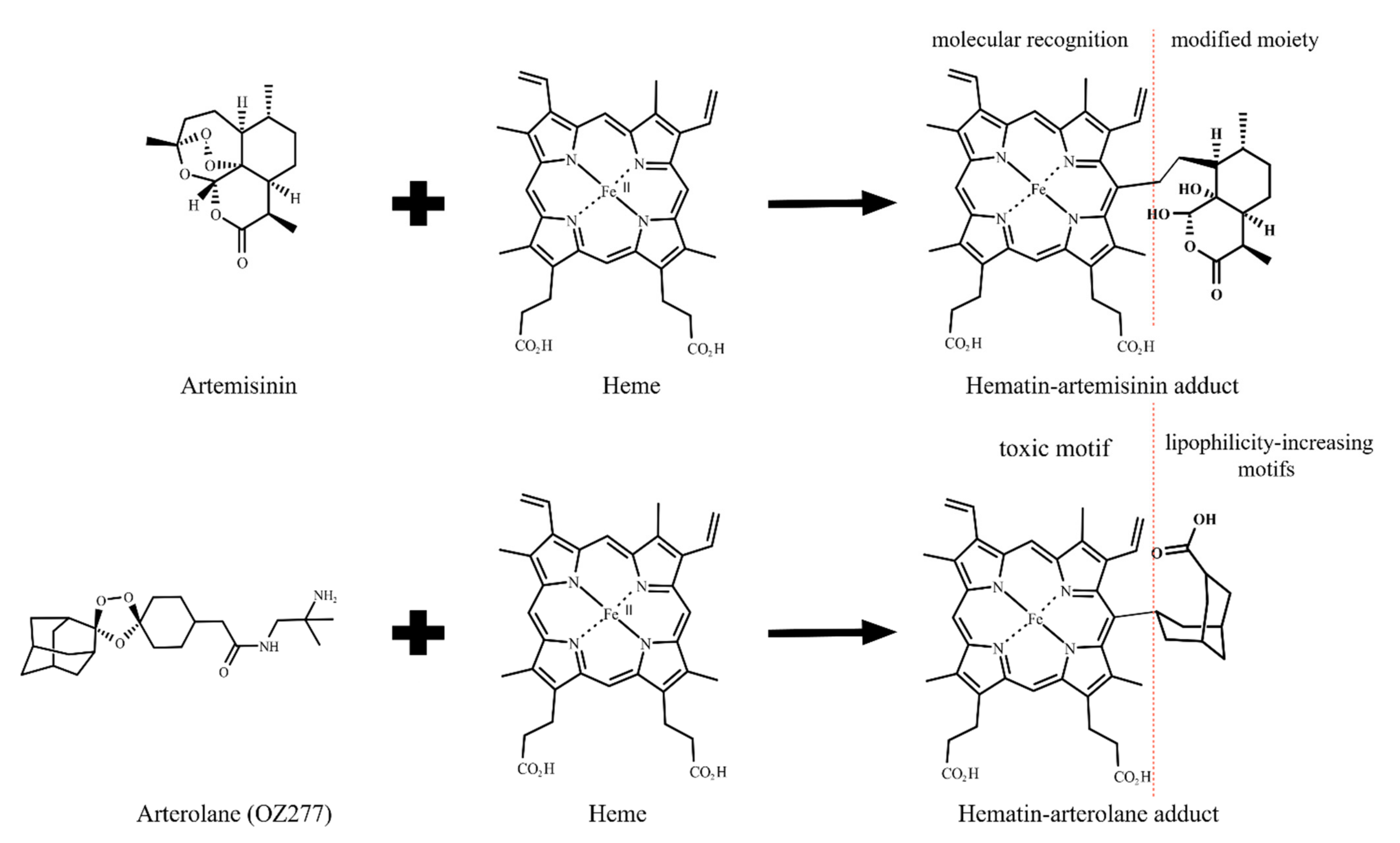
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 22 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quadros, H.C.; Silva, M.C.B.; Moreira, D.R.M. The Role of the Iron Protoporphyrins Heme and Hematin in the Antimalarial Activity of Endoperoxide Drugs. Pharmaceuticals 2022, 15, 60. https://doi.org/10.3390/ph15010060
Quadros HC, Silva MCB, Moreira DRM. The Role of the Iron Protoporphyrins Heme and Hematin in the Antimalarial Activity of Endoperoxide Drugs. Pharmaceuticals. 2022; 15(1):60. https://doi.org/10.3390/ph15010060
Chicago/Turabian StyleQuadros, Helenita C., Mariana C. B. Silva, and Diogo R. M. Moreira. 2022. "The Role of the Iron Protoporphyrins Heme and Hematin in the Antimalarial Activity of Endoperoxide Drugs" Pharmaceuticals 15, no. 1: 60. https://doi.org/10.3390/ph15010060
APA StyleQuadros, H. C., Silva, M. C. B., & Moreira, D. R. M. (2022). The Role of the Iron Protoporphyrins Heme and Hematin in the Antimalarial Activity of Endoperoxide Drugs. Pharmaceuticals, 15(1), 60. https://doi.org/10.3390/ph15010060