
ICG-001, an Inhibitor of the β-Catenin and cAMP Response Element-Binding Protein Dependent Gene Transcription, Decreases Proliferation but Enhances Migration of Osteosarcoma Cells


 ,
,  ,
,  ,
,
Abstract

1. Introduction
2. Results
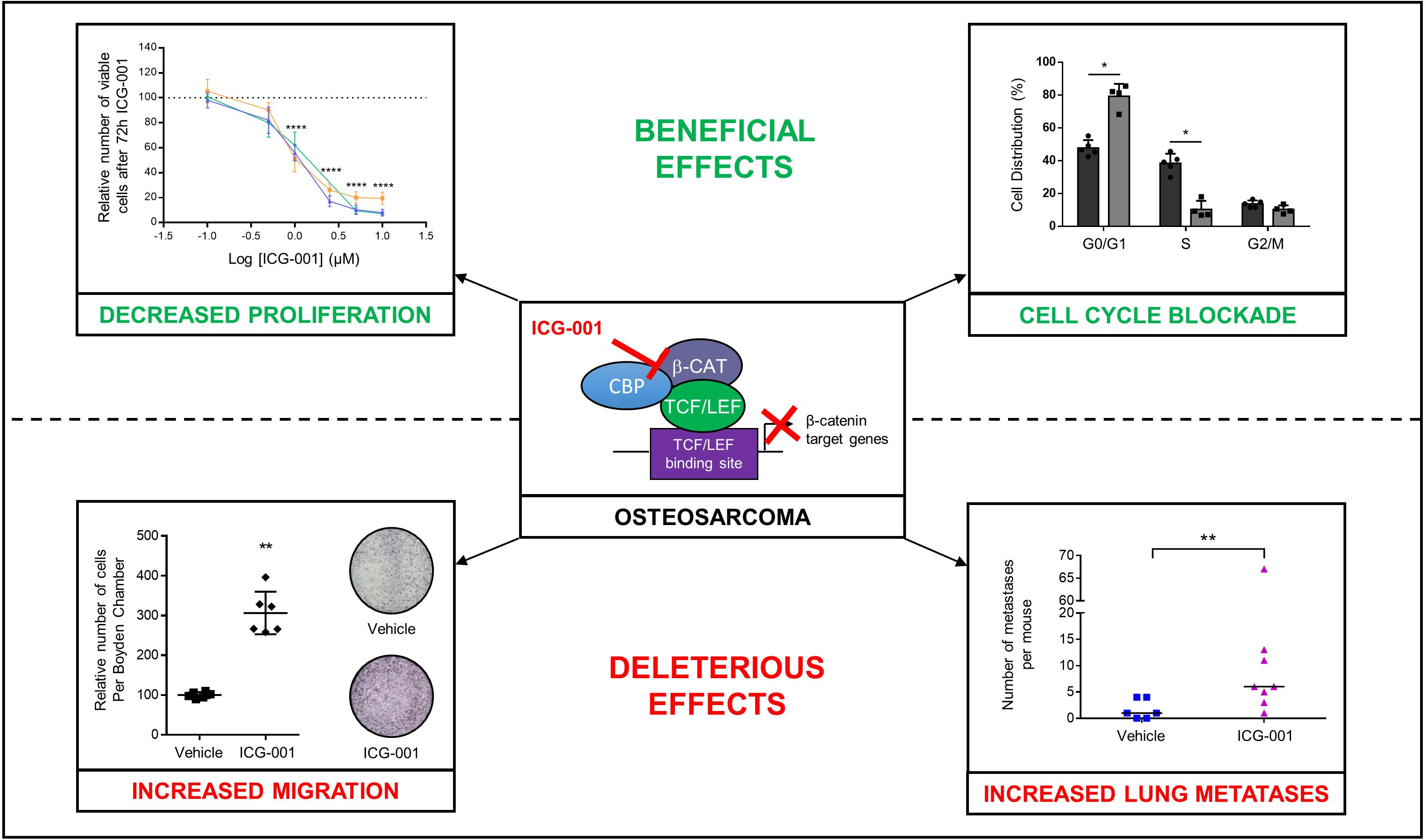
2.1. Osteosarcoma Cell Lines Expressing High β-Catenin Levels Are Sensitive to ICG-001
2.2. ICG-001 Induced a Cell Cycle Blockade in the G0/G1 Phase in Osteosarcoma Cells
2.3. ICG-001 Stimulates Osteosarcoma Cell Migration
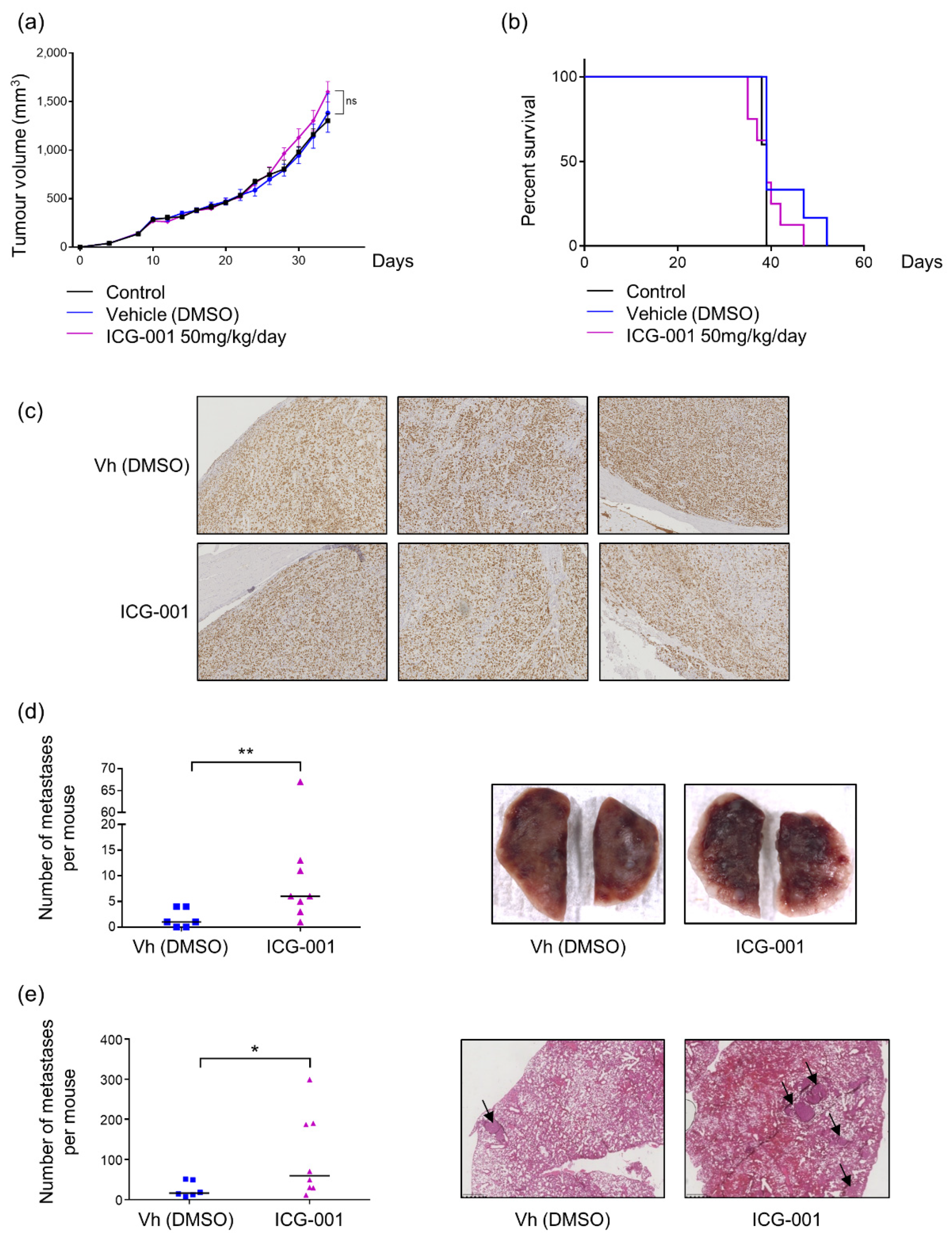
2.4. ICG-001 Stimulates Metastatic Dissemination to Lungs in a Pre-Clinical Mouse Model of Osteosarcoma
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Chemicals Reagents
4.2. RNA Seq Analysis
4.3. Luciferase Gene Reporter Assay
4.4. RT-qPCR
4.5. Western Blot
4.6. Viability Assay (Crystal Violet Staining)
4.7. Trypan Blue Counting
4.8. Caspase-3 Activity
4.9. Cell Cycle Analysis
4.10. Thymidine Synchronization
4.11. Migration Assay
4.12. Real-Time Migration Assay (xCELLigence®)
4.13. Osteosarcoma Xenograft Murine Model
4.14. Histological Analysis
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| APC | Adenomatous Polyposis Coli |
| ATCC | American Type Culture Collection |
| BCA | BiCinchoninic acid Assay |
| BSA | Bovine Serum Albumin |
| CBP | cAMP response element-binding protein |
| CDD | Charged Coupled Device |
| CDK | Cyclin-Dependent Kinase |
| CK1α | Casein Kinase 1α |
| CST | Cell Signaling Technology |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DMSO | Dimethyl sulfoxide |
| DPBS | Dulbecco’s Phosphate Buffer Saline |
| EDTA | EthyleneDiamineTetraacetic Acid |
| FBS | Fetal Bovine Serum |
| Fzd | Frizzled |
| GSEA | Gene Set enrichment analysis |
| GSK3β | Glycogen Synthase Kinase 3 β |
| HEK293 | Human Embryonic Kidney |
| HE | Hematoxylin-Eosin |
| HRP | Horseradish Peroxidase |
| LRP | low-density lipoprotein receptor related protein |
| MSCs | mesenchymal stem cells |
| miR | micro-RNAs |
| PMSF | phenylmethylsulforyl fluoride |
| PVDF | PolyVinyliDene Fluoride |
| RIPA | Radio-Immuno-Precipitation Assay |
| RPKM | Reads Per Kilobase Million |
| SDS-PAGE | Sodium Dodecyl Sulfate-Polyacrylamide gel |
| SOST | Sclerostin |
| SPARCL1 | Secreted Protein Acidic and Rich in Cysteine Like-1 |
| TBS | Tris-Buffered Saline |
| TCF/LEF | T cell factor/lymphoid enhancer factor |
References
- Desandes, E.; Lacour, B.; Belot, A.; Molinie, F.; Delafosse, P.; Tretarre, B.; Velten, M.; Sauleau, E.-A.; Woronoff, A.-S.; Guizard, A.-V.; et al. Cancer Incidence and Survival in Adolescents and Young Adults in France, 2000–2008. Pediatr. Hematol. Oncol. 2013, 30, 291–306. [Google Scholar] [CrossRef]
- Broadhead, M.L.; Clark, J.C.M.; Myers, D.E.; Dass, C.R.; Choong, P.F.M. The Molecular Pathogenesis of Osteosarcoma: A Review. Sarcoma 2011, 2011. [Google Scholar] [CrossRef]
- Mirabello, L.; Troisi, R.J.; Savage, S.A. International Osteosarcoma Incidence Patterns in Children and Adolescents, Middle Ages, and Elderly Persons. Int. J. Cancer 2009, 125, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Mirabello, L.; Troisi, R.J.; Savage, S.A. Osteosarcoma Incidence and Survival Rates from 1973 to 2004: Data from the Surveillance, Epidemiology, and End Results Program. Cancer 2009, 115, 1531–1543. [Google Scholar] [CrossRef]
- Strauss, S.J.; Whelan, J.S. Current Questions in Bone Sarcomas. Curr. Opin. Oncol. 2018, 30, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.D.; Nusse, R. Wnt Signaling: Multiple Pathways, Multiple Receptors, and Multiple Transcription Factors. J. Biol. Chem. 2006, 281, 22429–22433. [Google Scholar] [CrossRef] [PubMed]
- Stamos, J.L.; Weis, W.I. The β-Catenin Destruction Complex. Cold Spring Harb. Perspect. Biol. 2013, 5, a007898. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/Beta-Catenin Signaling: Components, Mechanisms, and Diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef]
- Anastas, J.N.; Moon, R.T. WNT Signalling Pathways as Therapeutic Targets in Cancer. Nat. Rev. Cancer 2013, 13, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Duchartre, Y.; Kim, Y.-M.; Kahn, M. The Wnt Signaling Pathway in Cancer. Crit. Rev. Oncol. Hematol. 2016, 99, 141–149. [Google Scholar] [CrossRef]
- Chen, C.; Zhao, M.; Tian, A.; Zhang, X.; Yao, Z.; Ma, X. Aberrant Activation of Wnt/β-Catenin Signaling Drives Proliferation of Bone Sarcoma Cells. Oncotarget 2015, 6, 17570–17583. [Google Scholar] [CrossRef]
- Iwaya, K.; Ogawa, H.; Kuroda, M.; Izumi, M.; Ishida, T.; Mukai, K. Cytoplasmic and/or Nuclear Staining of Beta-Catenin Is Associated with Lung Metastasis. Clin. Exp. Metastasis 2003, 20, 525–529. [Google Scholar] [CrossRef]
- Liu, W.; Zhao, Z.; Wang, Y.; Li, W.; Su, Q.; Jia, Q.; Zhang, J.; Zhang, X.; Shen, J.; Yin, J. Dioscin Inhibits Stem-Cell-like Properties and Tumor Growth of Osteosarcoma through Akt/GSK3/β-Catenin Signaling Pathway. Cell Death Dis. 2018, 9, 343. [Google Scholar] [CrossRef]
- Lu, Y.; Guan, G.-F.; Chen, J.; Hu, B.; Sun, C.; Ma, Q.; Wen, Y.-H.; Qiu, X.-C.; Zhou, Y. Aberrant CXCR4 and β-Catenin Expression in Osteosarcoma Correlates with Patient Survival. Oncol. Lett. 2015, 10, 2123–2129. [Google Scholar] [CrossRef]
- Cai, Y.; Mohseny, A.B.; Karperien, M.; Hogendoorn, P.C.W.; Zhou, G.; Cleton-Jansen, A.-M. Inactive Wnt/Beta-Catenin Pathway in Conventional High-Grade Osteosarcoma. J. Pathol. 2010, 220, 24–33. [Google Scholar] [CrossRef]
- Du, X.; Yang, J.; Yang, D.; Tian, W.; Zhu, Z. The Genetic Basis for Inactivation of Wnt Pathway in Human Osteosarcoma. BMC Cancer 2014, 14, 450. [Google Scholar] [CrossRef] [PubMed]
- Emami, K.H.; Nguyen, C.; Ma, H.; Kim, D.H.; Jeong, K.W.; Eguchi, M.; Moon, R.T.; Teo, J.-L.; Oh, S.W.; Kim, H.Y.; et al. A Small Molecule Inhibitor of Beta-Catenin/CREB-Binding Protein Transcription. Proc. Natl. Acad. Sci. USA 2004, 101, 12682–12687. [Google Scholar] [CrossRef] [PubMed]
- Arensman, M.D.; Telesca, D.; Lay, A.R.; Kershaw, K.M.; Wu, N.; Donahue, T.R.; Dawson, D.W. The CREB-Binding Protein Inhibitor ICG-001 Suppresses Pancreatic Cancer Growth. Mol. Cancer Ther. 2014, 13, 2303–2314. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, H.; Zheng, P.; Zheng, Y.; Luo, Q.; Xie, G.; Ma, Y.; Shen, L. ICG-001 Suppresses Growth of Gastric Cancer Cells and Reduces Chemoresistance of Cancer Stem Cell-like Population. J. Exp. Clin. Cancer Res. 2017, 36, 125. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Ikoma, A.; Shibakawa, M.; Shimoda, S.; Harada, K.; Saio, M.; Imamura, J.; Osawa, Y.; Kimura, M.; Nishikawa, K.; et al. Safety, Tolerability, and Preliminary Efficacy of the Anti-Fibrotic Small Molecule PRI-724, a CBP/β-Catenin Inhibitor, in Patients with Hepatitis C Virus-Related Cirrhosis: A Single-Center, Open-Label, Dose Escalation Phase 1 Trial. EBioMedicine 2017, 23, 79–87. [Google Scholar] [CrossRef]
- Ho, X.D.; Phung, P.; Le, V.Q.; Nguyen, V.H.; Reimann, E.; Prans, E.; Kõks, G.; Maasalu, K.; Le, N.T.; Trinh, L.H.; et al. Whole Transcriptome Analysis Identifies Differentially Regulated Networks between Osteosarcoma and Normal Bone Samples. Exp. Biol. Med. 2017, 242, 1802–1811. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Li, L.; Yan, W. Knockdown of TC-1 Enhances Radiosensitivity of Non-Small Cell Lung Cancer via the Wnt/β-Catenin Pathway. Biol. Open 2016, 5, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Stratford, E.W.; Daffinrud, J.; Munthe, E.; Castro, R.; Waaler, J.; Krauss, S.; Myklebost, O. The Tankyrase-Specific Inhibitor JW74 Affects Cell Cycle Progression and Induces Apoptosis and Differentiation in Osteosarcoma Cell Lines. Cancer Med. 2014, 3, 36–46. [Google Scholar] [CrossRef]
- Guo, Y.; Rubin, E.M.; Xie, J.; Zi, X.; Hoang, B.H. Dominant Negative LRP5 Decreases Tumorigenicity and Metastasis of Osteosarcoma in an Animal Model. Clin. Orthop. Relat. Res. 2008, 466, 2039–2045. [Google Scholar] [CrossRef] [PubMed]
- Shimozaki, S.; Yamamoto, N.; Domoto, T.; Nishida, H.; Hayashi, K.; Kimura, H.; Takeuchi, A.; Miwa, S.; Igarashi, K.; Kato, T.; et al. Efficacy of Glycogen Synthase Kinase-3β Targeting against Osteosarcoma via Activation of β-Catenin. Oncotarget 2016, 7, 77038–77051. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Goldstein, S.D.; Trucco, M.; Bautista Guzman, W.; Hayashi, M.; Loeb, D.M. A Monoclonal Antibody against the Wnt Signaling Inhibitor Dickkopf-1 Inhibits Osteosarcoma Metastasis in a Preclinical Model. Oncotarget 2016, 7, 21114–21123. [Google Scholar] [CrossRef] [PubMed]
- Qie, S.; Diehl, J.A. Cyclin D1, Cancer Progression, and Opportunities in Cancer Treatment. J. Mol. Med. 2016, 94, 1313–1326. [Google Scholar] [CrossRef]
- Boulaire, J.; Fotedar, A.; Fotedar, R. The Functions of the Cdk-Cyclin Kinase Inhibitor P21WAF1. Pathol. Biol. 2000, 48, 190–202. [Google Scholar]
- Gartel, A.L.; Tyner, A.L. Transcriptional Regulation of the P21((WAF1/CIP1)) Gene. Exp. Cell Res. 1999, 246, 280–289. [Google Scholar] [CrossRef]
- Ramazzotti, G.; Billi, A.M.; Manzoli, L.; Mazzetti, C.; Ruggeri, A.; Erneux, C.; Kim, S.; Suh, P.-G.; Cocco, L.; Faenza, I. IPMK and β-Catenin Mediate PLC-Β1-Dependent Signaling in Myogenic Differentiation. Oncotarget 2016, 7, 84118–84127. [Google Scholar] [CrossRef]
- Kamei, J.; Toyofuku, T.; Hori, M. Negative Regulation of P21 by Beta-Catenin/TCF Signaling: A Novel Mechanism by Which Cell Adhesion Molecules Regulate Cell Proliferation. Biochem. Biophys. Res. Commun. 2003, 312, 380–387. [Google Scholar] [CrossRef]
- Quasnichka, H.; Slater, S.C.; Beeching, C.A.; Boehm, M.; Sala-Newby, G.B.; George, S.J. Regulation of Smooth Muscle Cell Proliferation by Beta-Catenin/T-Cell Factor Signaling Involves Modulation of Cyclin D1 and P21 Expression. Circ. Res. 2006, 99, 1329–1337. [Google Scholar] [CrossRef]
- Xu, J.; Chen, Y.; Huo, D.; Khramtsov, A.; Khramtsova, G.; Zhang, C.; Goss, K.H.; Olopade, O.I. β-Catenin Regulates c-Myc and CDKN1A Expression in Breast Cancer Cells. Mol. Carcinog. 2016, 55, 431–439. [Google Scholar] [CrossRef]
- Verrecchia, F.; Rédini, F. Transforming Growth Factor-β Signaling Plays a Pivotal Role in the Interplay Between Osteosarcoma Cells and Their Microenvironment. Front. Oncol. 2018, 8, 133. [Google Scholar] [CrossRef]
- Duronio, R.J.; Xiong, Y. Signaling Pathways That Control Cell Proliferation. Cold Spring Harb. Perspect. Biol. 2013, 5, a008904. [Google Scholar] [CrossRef]
- Zou, J.; Zhang, W.; Li, X.-L. Effects of SOST Gene Silencing on Proliferation, Apoptosis, Invasion, and Migration of Human Osteosarcoma Cells Through the Wnt/β-Catenin Signaling Pathway. Calcif. Tissue Int. 2017, 100, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Martins-Neves, S.R.; Paiva-Oliveira, D.I.; Fontes-Ribeiro, C.; Bovée, J.V.M.G.; Cleton-Jansen, A.-M.; Gomes, C.M.F. IWR-1, a Tankyrase Inhibitor, Attenuates Wnt/β-Catenin Signaling in Cancer Stem-like Cells and Inhibits in Vivo the Growth of a Subcutaneous Human Osteosarcoma Xenograft. Cancer Lett. 2018, 414, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Chen, A.; Chen, J.; Yu, T.; Guo, F. SiRNA-Mediated Silencing of Beta-Catenin Suppresses Invasion and Chemosensitivity to Doxorubicin in MG-63 Osteosarcoma Cells. Asian Pac. J. Cancer Prev. 2011, 12, 239–245. [Google Scholar] [PubMed]
- Fang, F.; Van Cleave, A.; Helmuth, R.; Torres, H.; Rickel, K.; Wollenzien, H.; Sun, H.; Zeng, E.; Zhao, J.; Tao, J. Targeting the Wnt/β-Catenin Pathway in Human Osteosarcoma Cells. Oncotarget 2018, 9, 36780–36792. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.-J.; Jiang, Y.-Q.; Xu, N.-W.; Li, Q.; Zhang, Q.; Wang, S.-Y.; Li, J.; Wang, Y.-H.; Zhang, Y.-L.; Jiang, S.-H.; et al. SPARCL1 Suppresses Osteosarcoma Metastasis and Recruits Macrophages by Activation of Canonical WNT/β-Catenin Signaling through Stabilization of the WNT-Receptor Complex. Oncogene 2018, 37, 1049–1061. [Google Scholar] [CrossRef]
- Willert, J.; Epping, M.; Pollack, J.R.; Brown, P.O.; Nusse, R. A Transcriptional Response to Wnt Protein in Human Embryonic Carcinoma Cells. BMC Dev. Biol. 2002, 2, 8. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | MG63 | KHOS | 143B |
|---|---|---|---|
| IC50 (24 h) | 1.11 µM | 1.27 µM | 1.12 µM |
| IC50 (48 h) | 0.87 µM | 1.11 µM | 1.59 µM |
| IC50 (72 h) | 0.83 µM | 1.05 µM | 1.24 µM |
| Genes | Primers Sequences |
|---|---|
| h18S | Qf: cgattggatggtttagtgagg |
| Qr: agttcgaccgtcttctcagc | |
| hAXIN2 | Qf: atgattccatgtccatgacg |
| Qr: cttcacactgcgatgcattt | |
| hβ2M | Qf: ttctggcctggaggctatc |
| Qr: tcaggaaatttgactttccattc | |
| hBIRC5 | Qf: aggaccaccgcatctctacat |
| Qr: aagtctggctcgttctcagtg | |
| hGAPDH | Qf: tgggtgtgaaccatgagaagtatg |
| Qr: ggtgcaggaggcattgct |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danieau, G.; Morice, S.; Renault, S.; Brion, R.; Biteau, K.; Amiaud, J.; Cadé, M.; Heymann, D.; Lézot, F.; Verrecchia, F.; et al. ICG-001, an Inhibitor of the β-Catenin and cAMP Response Element-Binding Protein Dependent Gene Transcription, Decreases Proliferation but Enhances Migration of Osteosarcoma Cells. Pharmaceuticals 2021, 14, 421. https://doi.org/10.3390/ph14050421
Danieau G, Morice S, Renault S, Brion R, Biteau K, Amiaud J, Cadé M, Heymann D, Lézot F, Verrecchia F, et al. ICG-001, an Inhibitor of the β-Catenin and cAMP Response Element-Binding Protein Dependent Gene Transcription, Decreases Proliferation but Enhances Migration of Osteosarcoma Cells. Pharmaceuticals. 2021; 14(5):421. https://doi.org/10.3390/ph14050421
Chicago/Turabian StyleDanieau, Geoffroy, Sarah Morice, Sarah Renault, Régis Brion, Kevin Biteau, Jérôme Amiaud, Marie Cadé, Dominique Heymann, Frédéric Lézot, Franck Verrecchia, and et al. 2021. "ICG-001, an Inhibitor of the β-Catenin and cAMP Response Element-Binding Protein Dependent Gene Transcription, Decreases Proliferation but Enhances Migration of Osteosarcoma Cells" Pharmaceuticals 14, no. 5: 421. https://doi.org/10.3390/ph14050421
APA StyleDanieau, G., Morice, S., Renault, S., Brion, R., Biteau, K., Amiaud, J., Cadé, M., Heymann, D., Lézot, F., Verrecchia, F., Rédini, F., & Brounais-Le Royer, B. (2021). ICG-001, an Inhibitor of the β-Catenin and cAMP Response Element-Binding Protein Dependent Gene Transcription, Decreases Proliferation but Enhances Migration of Osteosarcoma Cells. Pharmaceuticals, 14(5), 421. https://doi.org/10.3390/ph14050421