
Sterigmatocystin Limits Plasmodium falciparum Proliferation and Transmission


Abstract
:1. Introduction
2. Results
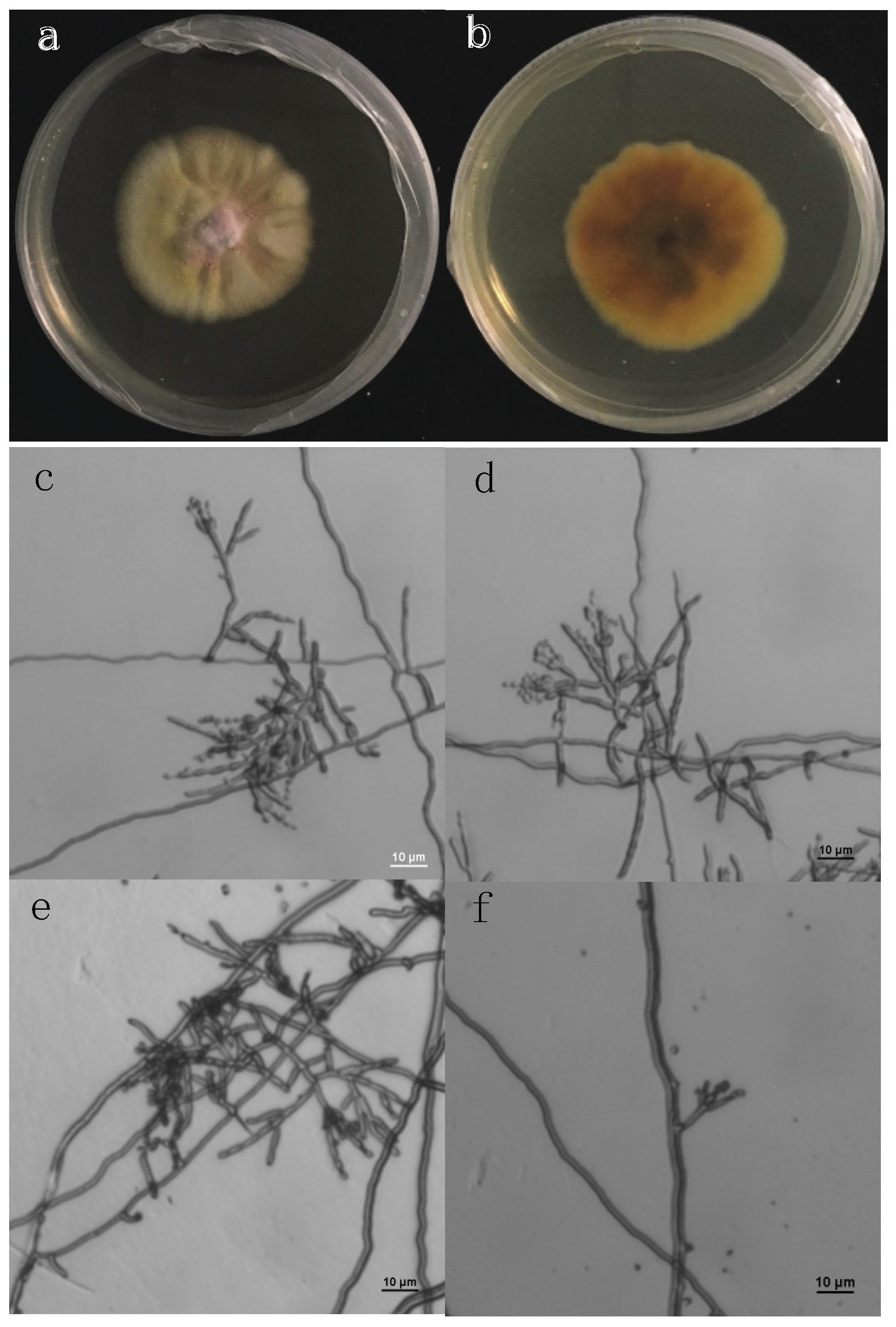
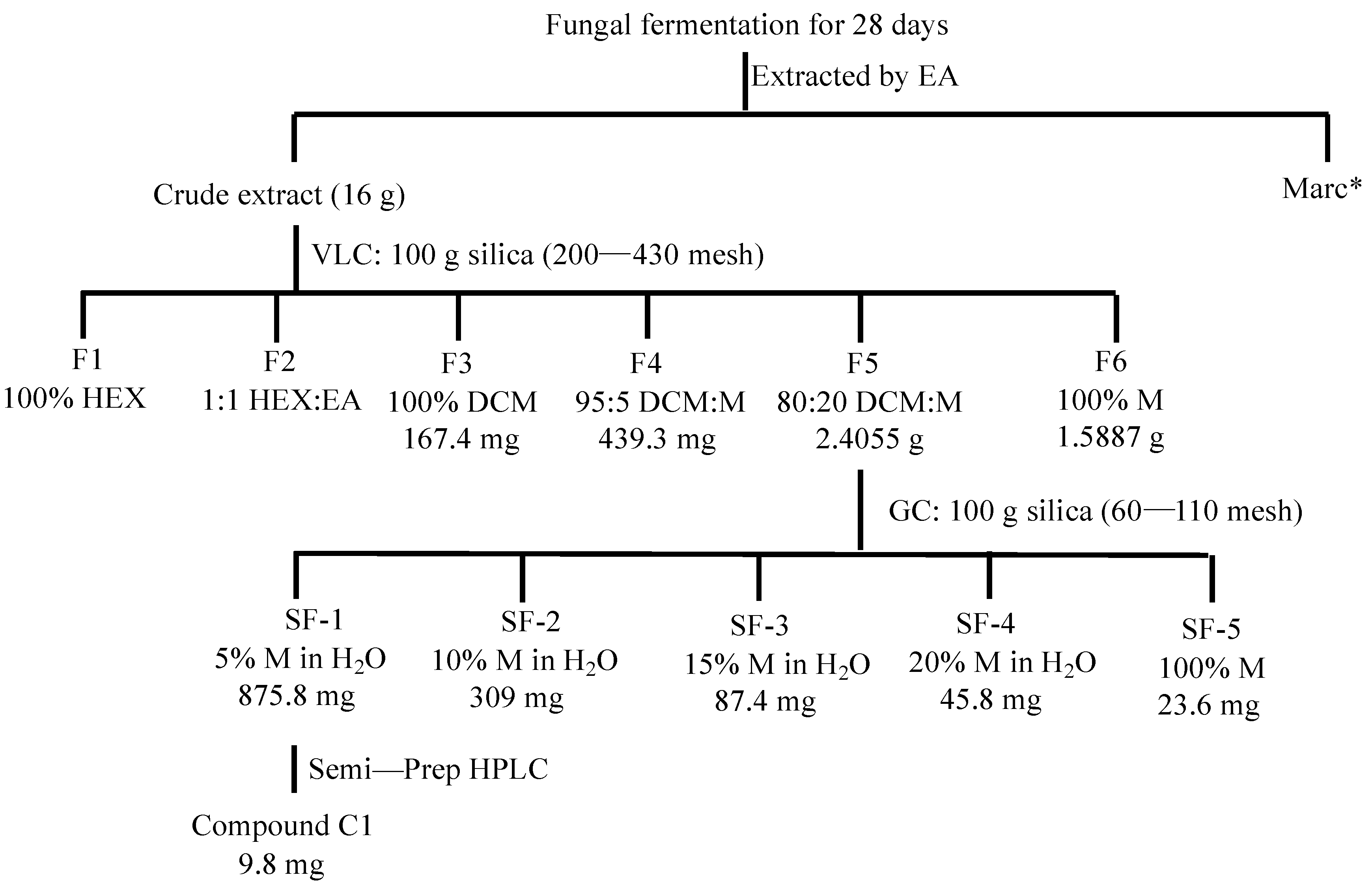
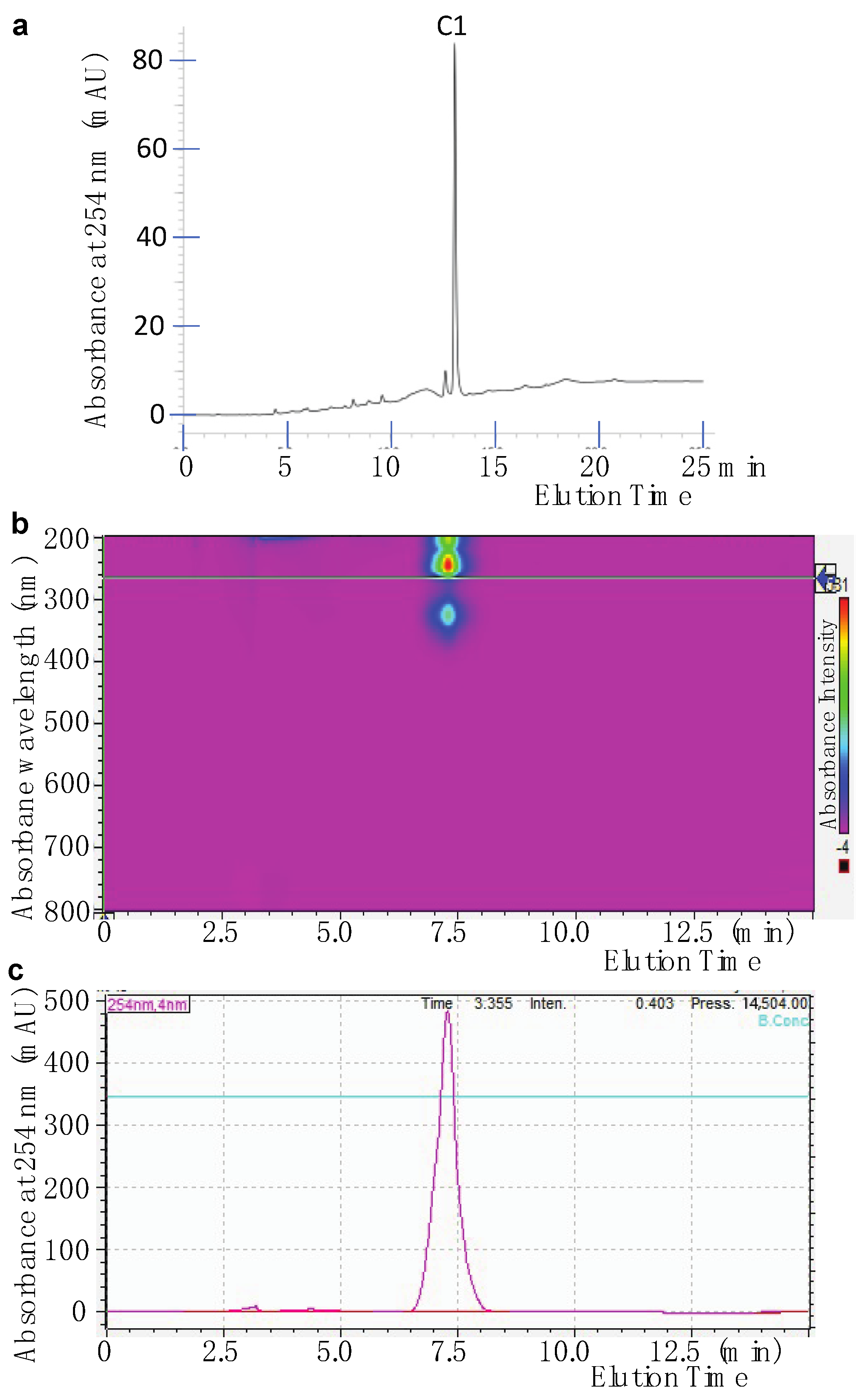
2.1. Isolation of Active Compounds from the Candidate Fungal Extract
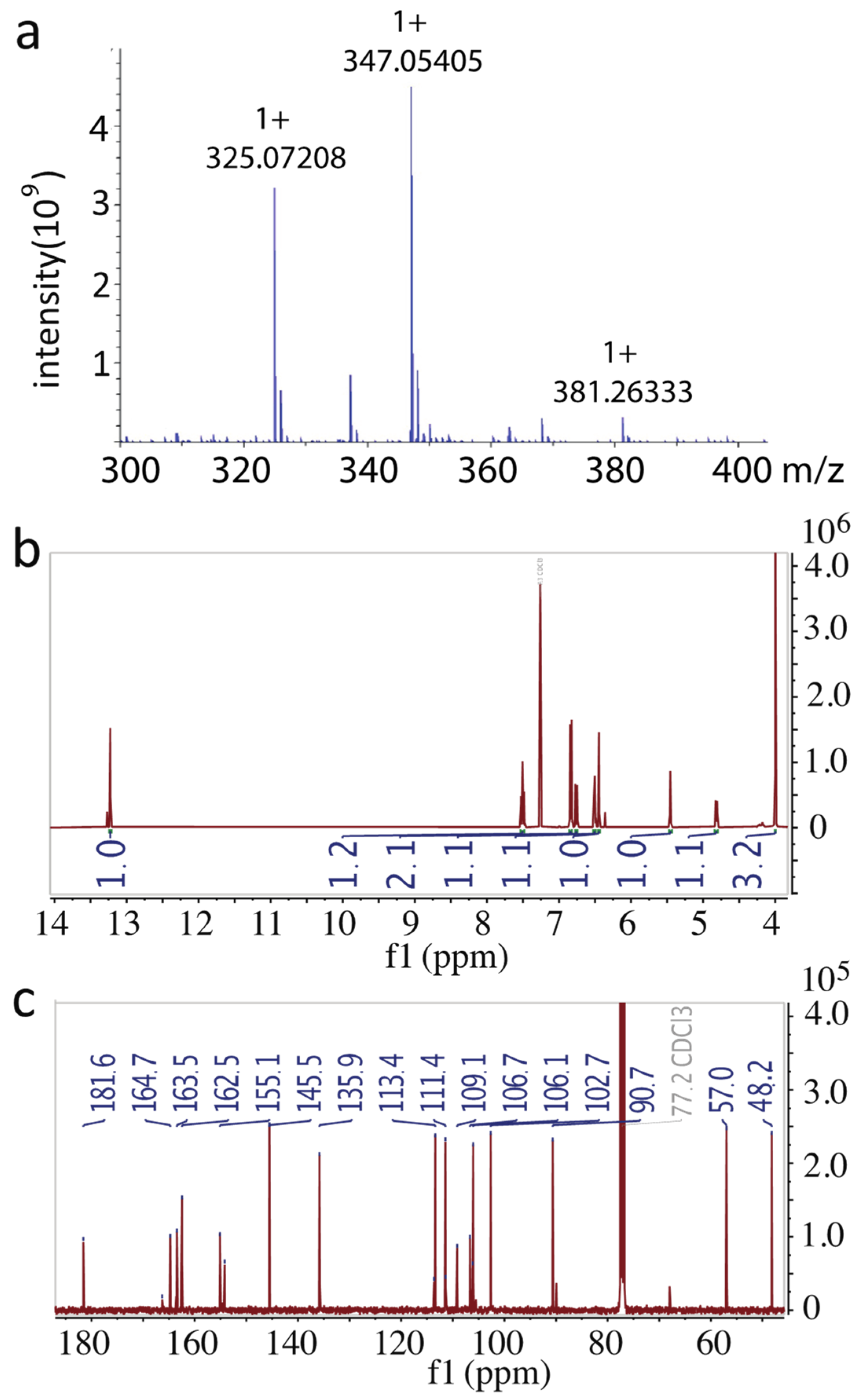
2.2. Identification of the Purified Compound
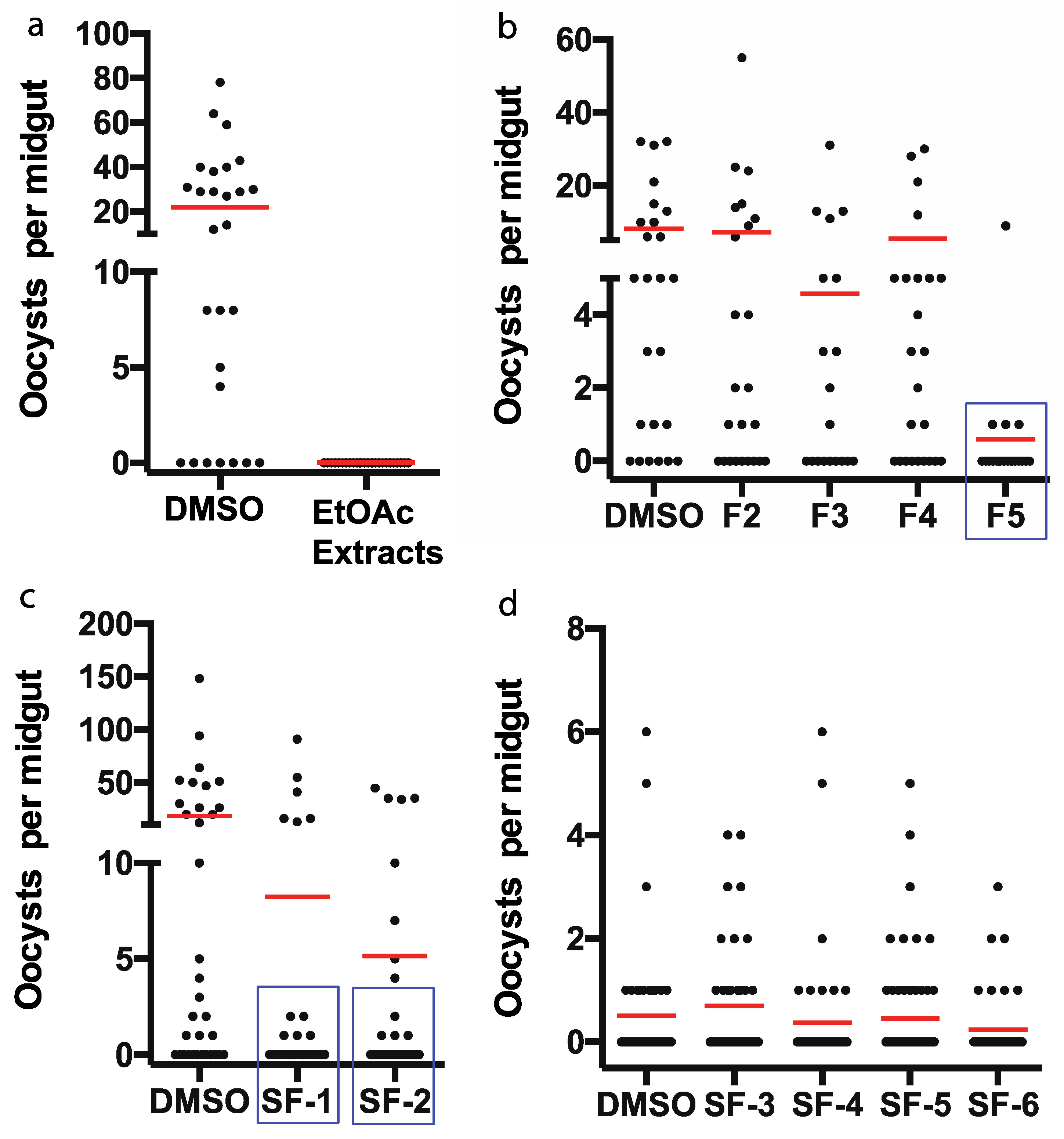
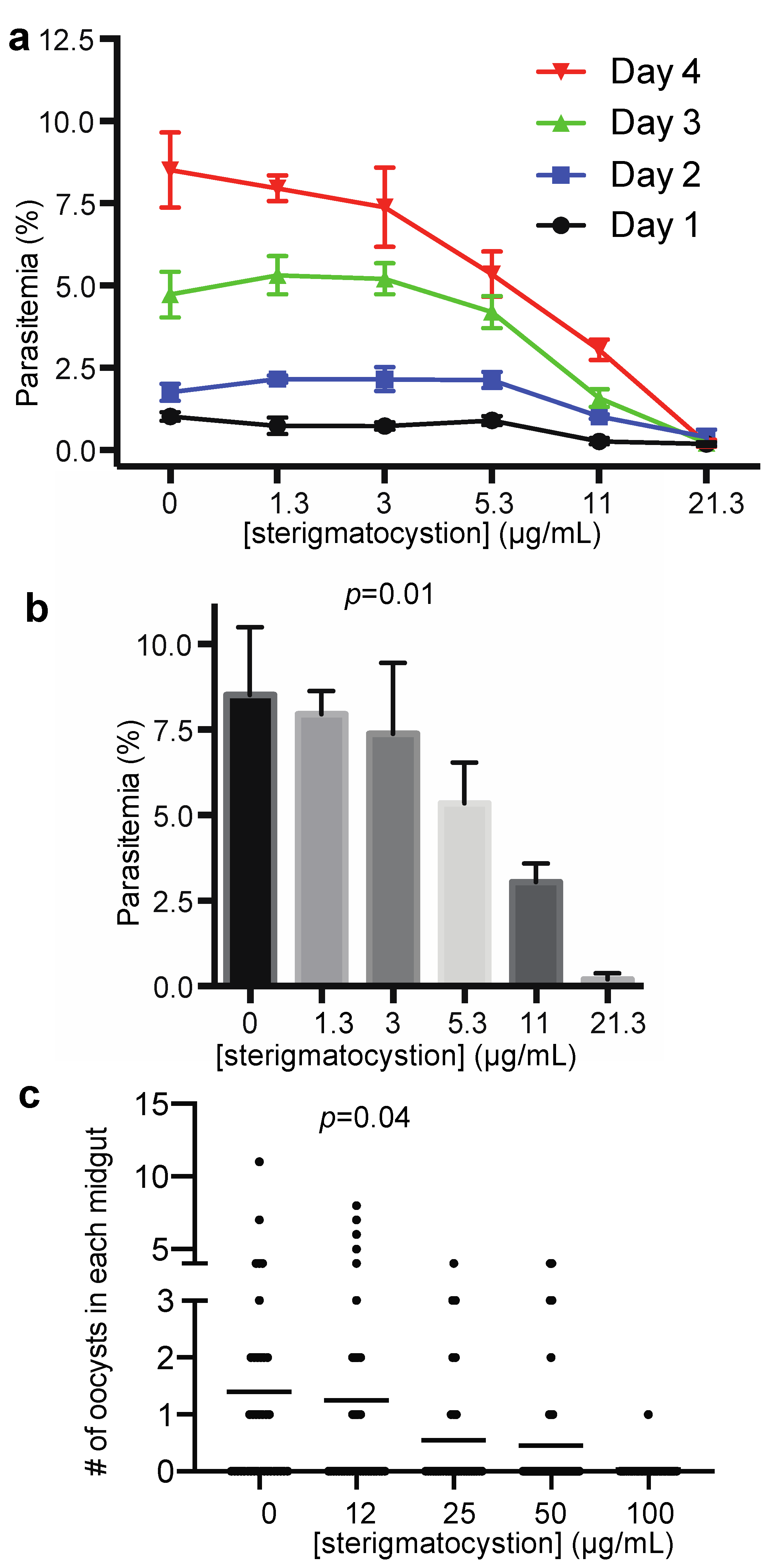
2.3. The Antimalarial Properties of the Pure Compound
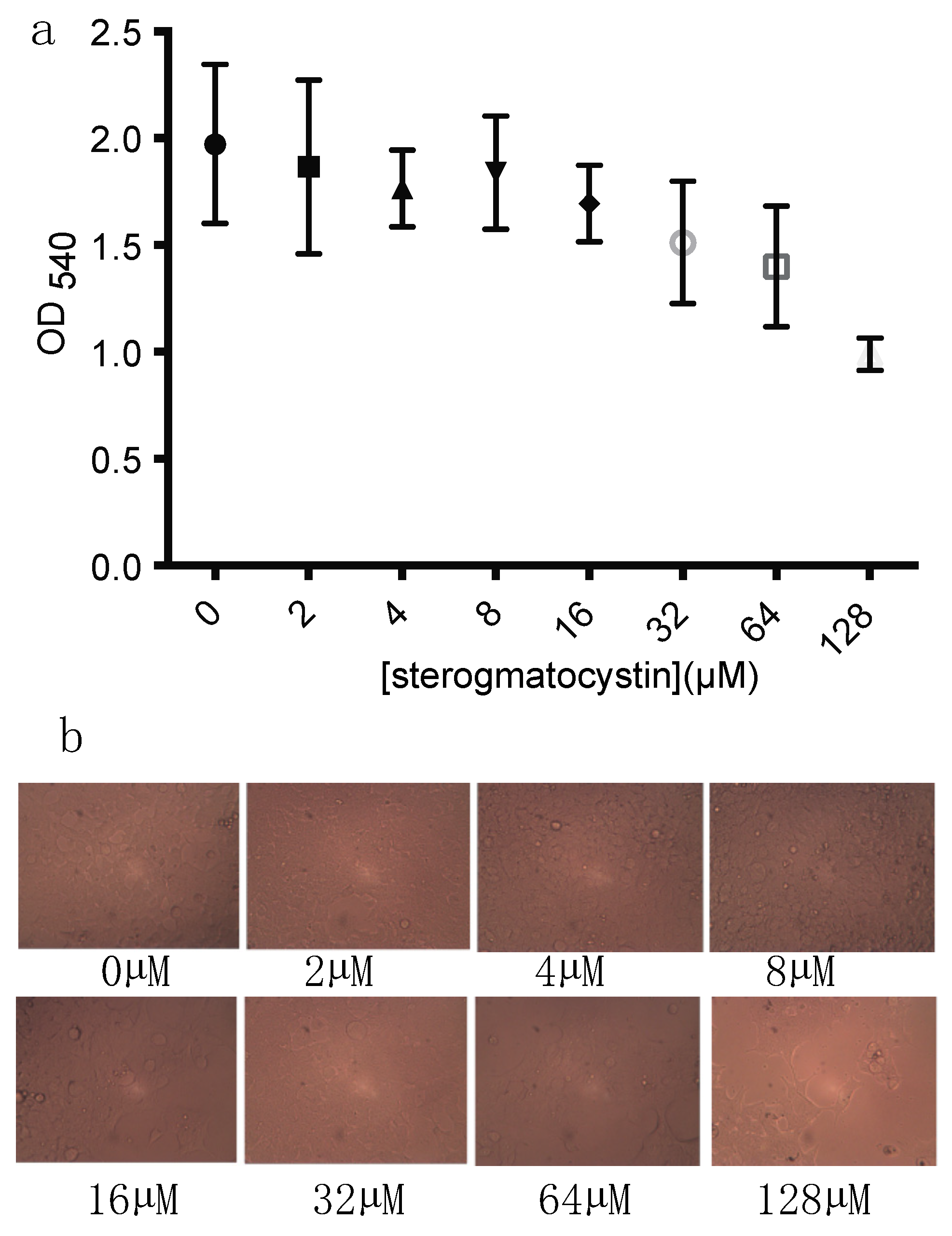
2.4. The Cytotoxicity of Sterigmatocystin
3. Discussion
4. Materials and Methods
4.1. Cloning and Sequencing ITS of a Fungal Species
4.2. Preparation of Crude Fungal Extract
4.3. Isolation of Active Compounds
4.4. High-Resolution Mass Spectrum
4.5. Nuclear Magnetic Resonance (NMR) Analysis
4.6. Asexual Plasmodium Falciparum Growth Inhibition Assays
4.7. Analyzing the Effect of Active Compounds on P. falciparum Infection in Anopheles Gambiae
4.8. Cytotoxic Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. World Malaria Report 2020. 2020, pp. 1–299. Available online: https://www.who.int/publications/i/item/9789240015791 (accessed on 26 November 2021).
- Moxon, C.A.; Gibbins, M.P.; McGuinness, D.; Milner, D.A.; Marti, M. New Insights into Malaria Pathogenesis. Annu. Rev. Pathol-Mech. 2020, 15, 315–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duru, V.; Witkowski, B.; Menard, D. Plasmodium falciparum Resistance to Artemisinin Derivatives and Piperaquine: A Major Challenge for Malaria Elimination in Cambodia. Am. J. Trop. Med. Hyg. 2016, 95, 1228–1238. [Google Scholar] [CrossRef]
- Schueffler, A.; Anke, T. Fungal natural products in research and development. Nat. Prod. Rep. 2014, 31, 1425–1448. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Huang, J.X.; Zhou, X.F.; Lin, X.P.; Liu, J.; Liao, S.R.; Wang, J.F.; Liu, F.A.; Tao, H.M.; Liu, Y.H. The Fungal Metabolites with Potential Antiplasmodial Activity. Curr. Med. Chem. 2018, 25, 3796–3825. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; He, Z.; Xue, J.; Chen, X.; Wei, X. beta-Resorcylic acid lactones from a Paecilomyces fungus. J. Nat. Prod. 2010, 73, 885–889. [Google Scholar] [CrossRef]
- Calcul, L.; Waterman, C.; Ma, W.S.; Lebar, M.D.; Harter, C.; Mutka, T.; Morton, L.; Maignan, P.; Van Olphen, A.; Kyle, D.E.; et al. Screening mangrove endophytic fungi for antimalarial natural products. Mar. Drugs 2013, 11, 5036–5050. [Google Scholar] [CrossRef]
- Isaka, M.; Palasarn, S.; Sriklung, K.; Kocharin, K. Cyclohexadepsipeptides from the insect pathogenic fungus Hirsutella nivea BCC 2594. J. Nat. Prod. 2005, 68, 1680–1682. [Google Scholar] [CrossRef]
- Niu, G.; Wang, B.; Zhang, G.; King, J.B.; Cichewicz, R.H.; Li, J. Targeting mosquito FREP1 with a fungal metabolite blocks malaria transmission. Sci. Rep. 2015, 5, 14694. [Google Scholar] [CrossRef] [Green Version]
- Niu, G.D.; Hao, Y.; Wang, X.H.; Gao, J.M.; Li, J. Fungal Metabolite Asperaculane B Inhibits Malaria Infection and Transmission. Molecules 2020, 25, 3018. [Google Scholar] [CrossRef]
- Niu, G.D.; Wang, X.H.; Hao, Y.; Kandel, S.; Niu, G.M.; Raptis, R.G.; Li, J. A novel fungal metabolite inhibits Plasmodium falciparum transmission and infection. Parasite Vector 2021, 14, 1–11. [Google Scholar] [CrossRef]
- Ateba, J.E.T.; Toghueo, R.M.K.; Awantu, A.F.; Mba’ning, B.M.; Gohlke, S.; Sahal, D.; Rodrigues-Filho, E.; Tsamo, E.; Boyom, F.F.; Sewald, N.; et al. Antiplasmodial Properties and Cytotoxicity of Endophytic Fungi from Symphonia globulifera (Clusiaceae). J. Fungi 2018, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- Niu, G.; Annamalai, T.; Wang, X.; Li, S.; Munga, S.; Niu, G.; Tse-Dinh, Y.C.; Li, J. A diverse global fungal library for drug discovery. PeerJ 2020, 8, e10392. [Google Scholar] [CrossRef]
- Sreeja-Raju, A.; Christopher, M.; Kooloth-Valappil, P.; Kuni-Parambil, R.; Gokhale, D.V.; Sankar, M.; Abraham, A.; Pandey, A.; Sukumaran, R.K. Penicillium janthinellum NCIM1366 shows improved biomass hydrolysis and a larger number of CAZymes with higher induction levels over Trichoderma reesei RUT-C30. Biotechnol. Biofuels 2020, 13, 196. [Google Scholar] [CrossRef]
- Nordberg, H.; Cantor, M.; Dusheyko, S.; Hua, S.; Poliakov, A.; Shabalov, I.; Smirnova, T.; Grigoriev, I.V.; Dubchak, I. The genome portal of the Department of Energy Joint Genome Institute: 2014 updates. Nucleic. Acids Res. 2014, 42, D26–D31. [Google Scholar] [CrossRef] [PubMed]
- Nieto CH, D.; Granero, A.M.; Zon, M.A.; Fernandez, H. Sterigmatocystin: A mycotoxin to be seriously considered. Food Chem. Toxicol. 2018, 118, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Rank, C.; Nielsen, K.F.; Larsen, T.O.; Varga, J.; Samson, R.A.; Frisvad, J.C. Distribution of sterigmatocystin in filamentous fungi. Fungal Biol. 2011, 115, 406–420. [Google Scholar] [CrossRef]
- Zingales, V.; Fernandez-Franzon, M.; Ruiz, M.J. Sterigmatocystin: Occurrence. toxicity and molecular mechanisms of action—A review. Food Chem. Toxicol. 2020, 146, 111802. [Google Scholar] [CrossRef] [PubMed]
- Zingales, V.; Fedeli, C.; Fernandez-Franzon, M.; Ruiz, M.J. Cytotoxic effects of individual and combined sterigmatocystin and nivalenol on liver hepatocellular carcinoma cells. Food Chem. Toxicol. 2020, 143, 111473. [Google Scholar] [CrossRef] [PubMed]
- Anninou, N.; Chatzaki, E.; Papachristou, F.; Pitiakoudis, M.; Simopoulos, C. Mycotoxins’ Activity at Toxic and Sub-Toxic Concentrations: Differential Cytotoxic and Genotoxic Effects of Single and Combined Administration of Sterigmatocystin. Ochratoxin A and Citrinin on the Hepatocellular Cancer Cell Line Hep3B. Int. J. Environ. Res. Public Health 2014, 11, 1855–1872. [Google Scholar] [CrossRef]
- Fujii, K.; Kurata, H.; Odashima, S.; Hatsuda, Y. Tumor induction by a single subcutaneous injection of sterigmatocystin in newborn mice. Cancer Res. 1976, 36, 1615–1618. [Google Scholar]
- Rajachan, O.A.; Kanokmedhakul, K.; Soytong, K.; Kanokmedhakul, S. Mycotoxins from the Fungus Botryotrichum piluliferum. J. Agric. Food Chem. 2017, 65, 1337–1341. [Google Scholar] [CrossRef] [PubMed]
- Moreno, E.; Varughese, T.; Spadafora, C.; Arnold, A.E.; Coley, P.D.; Kursar, T.A.; Gerwick, W.H.; Cubilla-Rios, L. Chemical constituents of the new endophytic fungus Mycosphaerella sp. nov. and their anti-parasitic activity. Nat. Prod. Commun. 2011, 6, 835–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | The Inhibitory Ratio of the Growth of Asexual Stage of P. falciparum (%) (30 µg/mL) |
|---|---|
| Crude extract | 100.0 |
| F1 (100% Hexane) | 31.5 |
| F2 (50:50 Hexane:EtOAc) | 60.6 |
| F3 (100% DCM) | 93.1 |
| F4 (95:5 DCM:MeOH) | 95.0 |
| F5 (80:20 DCM:MeOH) | 100.0 |
| F6 (100% MeOH) | 38.1 |
| SF-1 (5% MeOH) | 100.0 |
| SF-2 (10% MeOH) | 95.7 |
| SF-3 (15% MeOH) | 77.9 |
| SF-4 (20% MeOH) | 82.1 |
| SF-5 (100% MeOH) | 80.7 |
| Position | δC | δH (J in Hz) | Structure |
|---|---|---|---|
| 1 | 181.6, C | 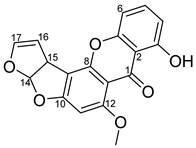 | |
| 2 | 109.1, C | ||
| 3 | 162.5, C | ||
| 4 | 113.4, CH | 6.84, d (7.3) | |
| 5 | 135.9, CH | 7.51, t (8.5) | |
| 6 | 106.7, CH | 6.84, d (7.3) | |
| 7 | 155.1, C | ||
| 8 | 154.2, C | ||
| 9 | 106.5, C | ||
| 10 | 164.7, C | ||
| 11 | 90.7, CH | 6.44, s | |
| 12 | 163.5, C | ||
| 13 | 106.5, C | ||
| 14 | 111.4, CH | 6.76, dd (0.9, 8.4) | |
| 15 | 48.2, CH | 4.82, dt (7.2, 2.3) | |
| 16 | 102.7, CH | 5.45, t (2.5) | |
| 17 | 145.5, CH | 6.51, m | |
| OCH3 | 57.0, CH3 | 4.00, s |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, G.; Kalani, K.; Wang, X.; Li, J. Sterigmatocystin Limits Plasmodium falciparum Proliferation and Transmission. Pharmaceuticals 2021, 14, 1238. https://doi.org/10.3390/ph14121238
Niu G, Kalani K, Wang X, Li J. Sterigmatocystin Limits Plasmodium falciparum Proliferation and Transmission. Pharmaceuticals. 2021; 14(12):1238. https://doi.org/10.3390/ph14121238
Chicago/Turabian StyleNiu, Guodong, Komal Kalani, Xiaohong Wang, and Jun Li. 2021. "Sterigmatocystin Limits Plasmodium falciparum Proliferation and Transmission" Pharmaceuticals 14, no. 12: 1238. https://doi.org/10.3390/ph14121238
APA StyleNiu, G., Kalani, K., Wang, X., & Li, J. (2021). Sterigmatocystin Limits Plasmodium falciparum Proliferation and Transmission. Pharmaceuticals, 14(12), 1238. https://doi.org/10.3390/ph14121238